Daily Activity Patterns and Co-Occurrence of Duikers Revealed by an Intensive Camera Trap Survey across Central African Rainforests

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
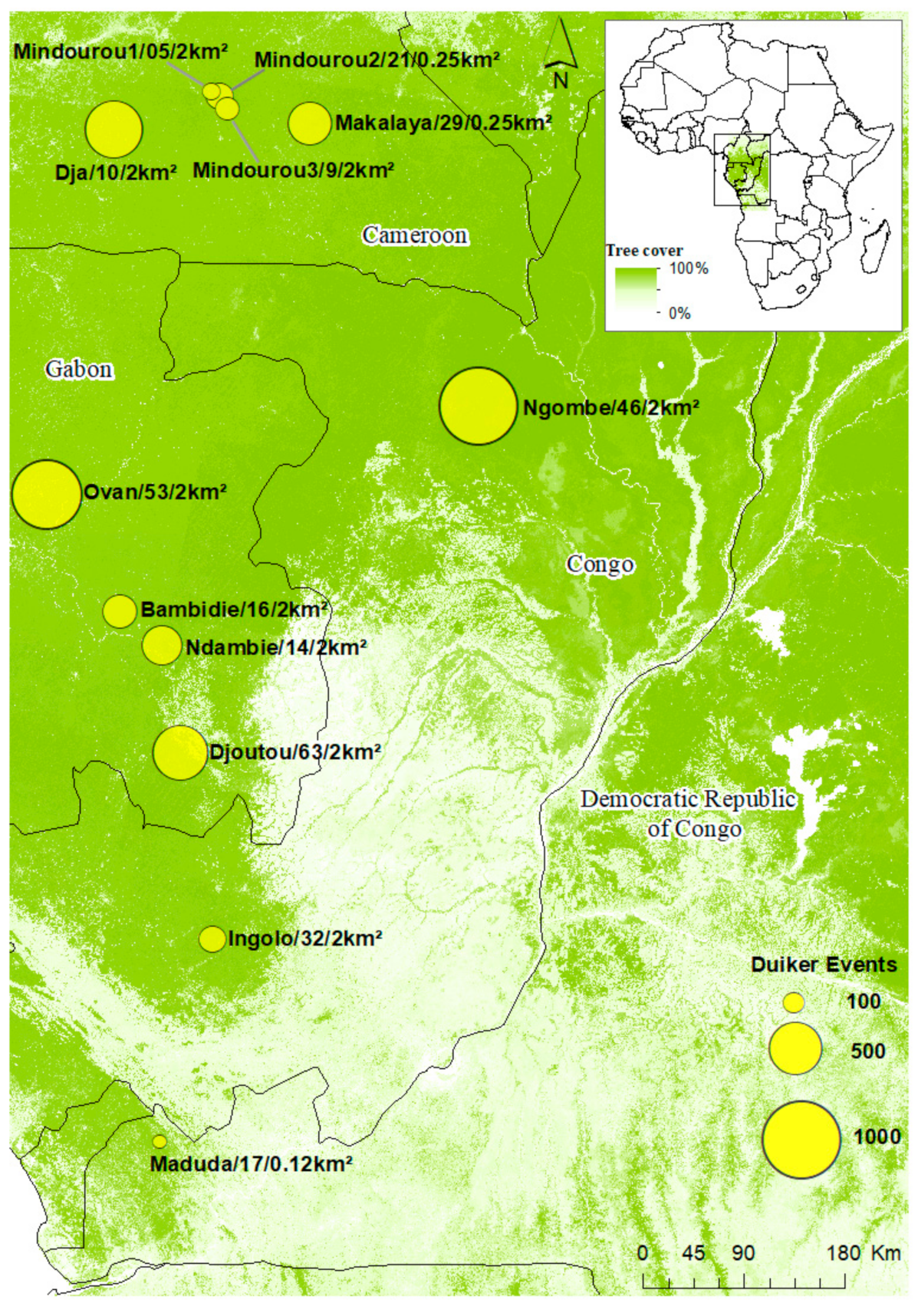
2.1. Study Area
2.2. Camera Traps Data Collection
- A photo of several individuals (multiple individuals in the same image) of the same species was treated as a single individual event [49].
- A photo of two species was treated as two separate independent events.
2.3. Statistical Analysis
2.3.1. Activity Periods
2.3.2. Overlap of Activity Patterns
2.3.3. Spatial Co-Occurrence Patterns
3. Results
3.1. Inventory Data
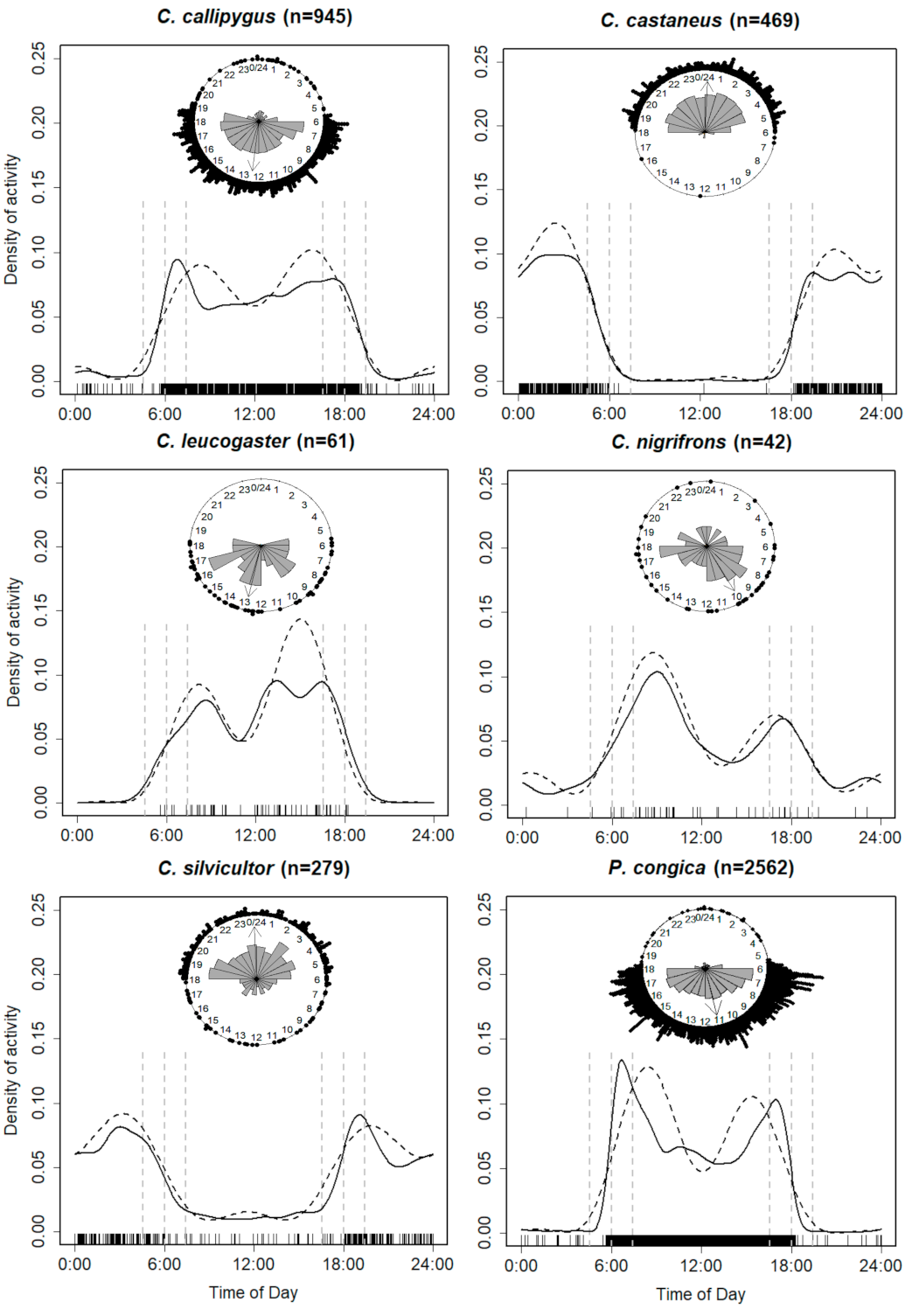
3.2. Species-Specific Activity Periods
3.3. Temporal Overlap Patterns
3.4. Spatial Co-Occurrence Patterns
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hempson, G.P.; Archibald, S.; Bond, W.J.A. Continent-Wide Assessment of the Form and Intensity of Large Mammal Herbivory in Africa. Science 2015, 350, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, B.J.; Robinson, T.J. Retrieval of Four Adaptive Lineages in Duiker Antelope: Evidence from Mitochondrial DNA Sequences and Fluorescence in Situ Hybridization. Mol. Phylogenet. Evol. 2001, 20, 409–425. [Google Scholar] [CrossRef] [PubMed]
- Ntie, S.; Johnston, A.R.; Mickala, P.; Bowkett, A.E.; Jansen van Vuuren, B.; Colyn, M.; Telfer, P.; Maisels, F.; Hymas, O.; Rouyer, R.L.; et al. Molecular Diagnostic for Identifying Central African Forest Artiodactyls from Faecal Pellets. Anim. Conserv. 2010, 13, 80–89. [Google Scholar] [CrossRef]
- Johnston, A.R.; Anthony, N.M.A. Multi-Locus Species Phylogeny of African Forest Duikers in the Subfamily Cephalophinae: Evidence for a Recent Radiation in the Pleistocene. BMC Evol. Biol. 2012, 12, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groves, C.; Grubb, P. Ungulate Taxonomy; Johns Hopkins University Press: Baltimore, ML, USA, 2011; p. 317. [Google Scholar]
- Fimbel, C.; Curran, B.; Usongo, L. Enhancing the sustainability of duiker hunting through community participation and controlled access in the Lobéké region of southeastern Cameroon. In Hunting for Sustainability in Tropical Forest; Robinson, J.G., Bennett, E.L., Eds.; Colombia University Press: New York, NY, USA, 2000; pp. 356–374. [Google Scholar]
- Van Vliet, N.; Zundel, S.; Miquel, C.; Taberlet, P.; Nasi, R. Distinguishing Dung from Blue, Red and Yellow-Backed Duikers through Noninvasive Genetic Techniques. Afr. J. Ecol. 2007, 46, 411–417. [Google Scholar] [CrossRef]
- Ghassemi-Khademi, T.; Hamidi, K. Re-Evaluation of Molecular Phylogeny of the Subfamily Cephalophinae (Bovidae: Artiodactyla); with Notes on Diversification of Body Size. Jordan J. Biol. Sci. 2019, 12, 637–647. [Google Scholar]
- Castelló, J.R. Bovids of the World: Antelopes, Gazelles, Cattle, Goats, Sheep, and Relatives; Princeton University Press: Princeton, NJ, USA, 2016; Volume 104, p. 665. [Google Scholar]
- Hedwig, D.; Kienast, I.; Bonnet, M.; Curran, B.K.; Courage, A.; Boesch, C.; Kühl, H.S.; King, T. A Camera Trap Assessment of the Forest Mammal Community within the Transitional Savannah-Forest Mosaic of the Batéké Plateau National Park, Gabon. Afr. J. Ecol. 2018, 56, 777–790. [Google Scholar] [CrossRef] [Green Version]
- Mockrin, M.H. Duiker Demography and Dispersal under Hunting in Northern Congo. Afr. J. Ecol. 2010, 48, 239–247. [Google Scholar] [CrossRef]
- Yasuoka, H.; Hirai, M.; Kamgaing, T.O.W.; Dzefack, Z.C.B.; Kamdoum, E.C.; Bobo, K.S. Changes in the Composition of Hunting Catches in Southeastern Cameroon: A Promising Approach for Collaborative Wildlife Management between Ecologists and Local Hunters. Ecol. Soc. 2015, 20. [Google Scholar] [CrossRef] [Green Version]
- Vanthomme, H.; Bellé, B.; Forget, P.-M. Bushmeat Hunting Alters Recruitment of Large-Seeded Plant Species in Central Africa. Biotropica 2010, 42, 672–679. [Google Scholar] [CrossRef]
- Newing, H. Bushmeat Hunting and Management: Implications of Duiker Ecology and Interspecific Competition. Biodivers. Conserv. 2001, 10, 99–118. [Google Scholar] [CrossRef]
- Azevedo, F.C.; Lemos, F.G.; Freitas-Junior, M.C.; Rocha, D.G.; Azevedo, F.C.C. Puma Activity Patterns and Temporal Overlap with Prey in a Human-Modified Landscape at Southeastern Brazil. J. Zool. 2018, 305, 246–255. [Google Scholar] [CrossRef]
- Lashley, M.A.; Cove, M.V.; Chitwood, M.C.; Penido, G.; Gardner, B.; Deperno, C.S.; Moorman, C.E. Estimating Wildlife Activity Curves: Comparison of Methods and Sample Size. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynam, A.J.; Jenks, K.E.; Tantipisanuh, N.; Chutipong, W.; Ngoprasert, D.; Gale, G.A.; Steinmetz, R.; Sukmasuang, R.; Bhumpakphan, N.; Grassman, L.I.; et al. Terrestrial activity patterns of wild cats from camera-trapping. Raffles Bull. Zool. 2013, 61, 407–415. [Google Scholar]
- Rowcliffe, J.M.; Kays, R.; Kranstauber, B.; Carbone, C.; Jansen, P.A. Quantifying Levels of Animal Activity Using Camera Trap Data. Methods Ecol. Evol. 2014, 5, 1170–1179. [Google Scholar] [CrossRef] [Green Version]
- Schoener, T.W. Resource Partitioning in Ecological Communities. Science 1974, 185, 27–39. [Google Scholar] [CrossRef]
- Kneitel, J.M.; Chase, J.M. Trade-Offs in Community Ecology: Linking Spatial Scales and Species Coexistence. Ecol. Lett. 2004, 7, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.H. The Comparison of Usage and Availability Measurements for Evaluating Resource Preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- Halle, S.; Stenseth, N.C. (Eds.) Introduction. In Activity Patterns in Small Mammals—An Ecological Approach; Springer: Berlin, Germany, 2000; Volume 141, pp. 3–17. [Google Scholar]
- Bennie, J.J.; Duffy, J.P.; Inger, R.; Gaston, K.J. Biogeography of Time Partitioning in Mammals. Proc. Natl. Acad. Sci. USA 2014, 111, 13727–13732. [Google Scholar] [CrossRef] [Green Version]
- Ridout, M.S.; Linkie, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 2009, 14, 322–337. [Google Scholar] [CrossRef]
- Weitzman, M.S. Measures of the Overlap of Income Distribution of White and Negro Families in the United States; Bureau of the Census: Washington, DC, USA, 1970; Volume 22. [Google Scholar]
- Veech, J.A.A. Probabilistic Model for Analysing Species Co-Occurrence. Glob. Ecol. Biogeogr. 2013, 22, 252–260. [Google Scholar] [CrossRef]
- Rota, C.T.; Ferreira, M.A.R.; Kays, R.W.; Forrester, T.D.; Kalies, L.; Mcshea, W.J.; Parsons, A.W.; Millspaugh, J.J. A Multispecies Occupancy Model for Two or More Interacting Species. Methods Ecol. Evol. 2016, 7, 1164–1173. [Google Scholar] [CrossRef]
- Bourgoin, G.; Garel, M.; Blanchard, P.; Dubray, D.; Maillard, D.; Gaillard, J.M. Daily Responses of Mouflon (Ovis Gmelini Musimon × Ovis Sp.) Activity to Summer Climatic Conditions. Can. J. Zool. 2011, 89, 765–773. [Google Scholar] [CrossRef] [Green Version]
- Frey, S.; Fisher, J.T.; Burton, A.C.; Volpe, J.P. Investigating Animal Activity Patterns and Temporal Niche Partitioning Using Camera-Trap Data: Challenges and Opportunities. Remote Sens. Ecol. Conserv. 2017, 3, 123–132. [Google Scholar] [CrossRef]
- Saint-Girons, M.C. Le rythme circadian d’activité chez les mammifères holarctiques. Mém. Mus. Natl. Hist. Nat. Sér. A Zool. 1966, 40, 101–187. (In French) [Google Scholar]
- Johnsingh, T.A.J. Large Mammalian Prey-Predators in Bandipur. J. Bombay Nat. Hist. Soc. 1983, 80, 1–57. [Google Scholar]
- Bowland, A.E.; Perrin, M.R. Temporal and Spatial Patterns in Blue Duikers Philatomba Monticola and Red Duikers Cephalophus natalensis. J. Zool. 1995, 237, 487–498. [Google Scholar] [CrossRef]
- Dubost, G. L‘écologie et La Vie Sociale Du Céphalophe Bleu (Cephalophus Monticola Thunberg), Petit Ruminant Forestier Africain. Z. Tierpsychol. 1980, 54, 205–266. [Google Scholar] [CrossRef]
- Bessone, M.; Kühl, H.S.; Hohmann, G.; Herbinger, I.; N’Goran, K.P.; Asanzi, P.; da Costa, P.B.; Dérozier, V.; Fotsing, E.D.B.; Beka, B.I.; et al. Drawn out of the Shadows: Surveying Secretive Forest Species with Camera Trap Distance Sampling. J. Appl. Ecol. 2020, 57, 963–974. [Google Scholar] [CrossRef] [Green Version]
- Harmsen, B.J.; Foster, R.J.; Silver, S.C.; Ostro, L.E.T.; Doncaster, C.P. Jaguar and Puma Activity Patterns in Relation to Their Main Prey. Mamm. Biol. 2011, 76, 320–324. [Google Scholar] [CrossRef]
- Feer, F. Stratégies Écologiques de Deux Espèces de Bovidés Sympatriques de La Forêt Sempervirente Africaine (Cephalophus Callipygus et C. Dorsalis): Influence du rythme d’activité. Ph.D. Thesis, Université Pierre et Marie Curie, Paris, France, 1988. (In French). [Google Scholar]
- Feer, F. Comparaison Des Régimes Alimentaires de Cephalophus Callipygus et C. Dorsalis. Bovidés Sympatriques de La Forêt Sempervirente Africaine. Mammalia 1989, 53, 563–604. [Google Scholar] [CrossRef]
- Kranz, K.R.; Lumpkin, S. Notes on the yellow-backed duiker Cephalophus sylvicultor in captivity with comments on its natural history. Int. Zoo Yearb. 1982, 22, 232–240. [Google Scholar] [CrossRef]
- Lumpkin, S.; Kranz, K.R. Cephalophus Sylvicultor. Mamm. Species 1984, 225, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gessner, J.; Buchwald, R.; Wittemyer, G. Assessing Species Occurrence and Species-Specific Use Patterns of Bais (Forest Clearings) in Central Africa with Camera Traps. Afr. J. Ecol. 2014, 52, 59–68. [Google Scholar] [CrossRef]
- Nakashima, Y.; Hongo, S.; Akomo-Okoue, E.F. Landscape-Scale Estimation of Forest Ungulate Density and Biomass Using Camera Traps: Applying the REST Model. Biol. Conserv. 2020, 241, e108381. [Google Scholar] [CrossRef]
- Illius, A.W.; Gordon, I.J. Modelling the Nutritional Ecology of Ungulate Herbivores: Evolution of Body Size and Competitive Interactions. Oecologia 1992, 89, 428–434. [Google Scholar] [CrossRef] [PubMed]
- White, F. The vegetation of Africa: A descriptive memoir to accompany the UNESCO/AETFAT/UNSO vegetation map of Africa. Nat. Resour. Res. 1983, 20, 351–356. [Google Scholar]
- Gond, V.; Fayolle, A.; Pennec, A.; Cornu, G.; Mayaux, P.; Camberlin, P.; Doumenge, C.; Fauvet, N.; Gourlet-Fleury, S. Vegetation Structure and Greenness in Central Africa from Modis Multi-Temporal Data. Philos. Trans. R. Soc. B: Biol. Sci. 2013, 368, e20120309. [Google Scholar] [CrossRef] [Green Version]
- Van Schaik, C.P.; Griffiths, M. Activity Periods of Indonesian Rain Forest Mammals. Biotropica 1996, 28, e105. [Google Scholar] [CrossRef]
- Noor, A.; Mir, Z.R.; Veeraswami, G.G.; Habib, B. Activity Patterns and Spatial Co-Occurrence of Sympatric Mammals in the Moist Temperate Forest of the Kashmir Himalaya, India. Folia Zool. 2017, 66, 231–241. [Google Scholar] [CrossRef]
- Camera Base. Available online: http://www.atrium-biodiversity.org/tools/camerabase/ (accessed on 4 June 2019).
- Groves, C.P.; Leslie, D.M., Jr. Family Bovidae (hollow-horned ruminants). In Handbook of the Mammals of the World, 2nd ed.; Wilson, D.E., Mittermeier, R.A., Eds.; Lynx Edicions: Barcelona, Spain, 2011; Volume 2, pp. 755–773. [Google Scholar]
- Caravaggi, A.; Gatta, M.; Vallely, M.C.; Hogg, K.; Freeman, M.; Fadaei, E.; Dick, J.T.A.; Montgomery, W.I.; Reid, N.; Tosh, D.G. Seasonal and Predator-Prey Effects on Circadian Activity of Free-Ranging Mammals Revealed by Camera Traps. PeerJ 2018, 6, e5287. [Google Scholar] [CrossRef] [PubMed]
- Cusack, J.J.; Dickman, A.J.; Rowcliffe, J.M.; Carbone, C.; Macdonald, D.W.; Coulson, T. Random versus Game Trail-Based Camera Trap Placement Strategy for Monitoring Terrestrial Mammal Communities. PLoS ONE 2015, 10, e0126373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, P.; Iezzi, M.E.; de Angelo, C.; Varela, D.; di Bitetti, M.S.; Paviolo, A. Effects of Human Impacts on Habitat Use, Activity Patterns and Ecological Relationships among Medium and Small Felids of the Atlantic Forest. PLoS ONE 2018, 13, e0200806. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-Resolution Global Maps of 21st-Century Forest Cover Change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USNO. Available online: https://aa.usno.navy.mil/faq/docs/world_tzones.php. (accessed on 4 April 2019).
- Gómez, H.; Wallace, R.B.; Ayala, G.; Tejada, R. Dry Season Activity Periods of Some Amazonian Mammals. Stud. Neotrop. Fauna Environ. 2005, 40, 91–95. [Google Scholar] [CrossRef]
- Ross, J.; Hearn, A.J.; Johnson, P.J.; Macdonald, D.W. Activity patterns and temporal avoidance by prey in response to Sunda clouded leopard predation risk. J. Zool. Lond. 2013, 290, 96–106. [Google Scholar] [CrossRef]
- Pewsey, A.; Neuhäuser, M.; Ruxton, G.D. Circular Statistics in R, 1st ed.; Oxford University Press: New York, NY, USA, 2013; p. 183. [Google Scholar]
- Landler, L.; Ruxton, G.D.; Malkemper, E.P. The Hermans-Rasson Test as a Powerful Alternative to the Rayleigh Test for Circular Statistics in Biology. BMC Ecol. 2019, 19. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Durán, J.J. Circular Distributions Based on Nonnegative Trigonometric Sums. Biometrics 2004, 60, 499–503. [Google Scholar] [CrossRef]
- Linkie, M.; Ridout, M.S. Assessing Tiger-Prey Interactions in Sumatran Rainforests. J. Zool. 2011, 284, 224–229. [Google Scholar] [CrossRef]
- Monterroso, P.; Alves, P.C.; Ferreras, P. Plasticity in Circadian Activity Patterns of Mesocarnivores in Southwestern Europe: Implications for Species Coexistence. Behav. Ecol. Sociobiol. 2014, 68, 1403–1417. [Google Scholar] [CrossRef] [Green Version]
- Overview of the Overlap Package. Available online: https://cran.r-project.org/web/packages/overlap/vignettes/overlap.pdf (accessed on 18 July 2019).
- Havmøller, R.W. Assessing the Activity Pattern Overlap among Leopards (Panthera Pardus), Potential Prey and Competitors in a Complex Landscape in Tanzania. J. Zool. 2020, 311, 175–182. [Google Scholar] [CrossRef]
- Griffith, D.M.; Veech, J.A.; Marsh, C.J. Cooccur: Probabilistic Species Co-Occurrence Analysis in R. J. Stat. Softw. 2016, 69. [Google Scholar] [CrossRef]
- The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 10 April 2016).
- Dray, S.; Dufour, A.B. The Ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Package ‘CircMLE’. Available online: https://cran.r-project.org/web/packages/CircMLE/CircMLE.pdf (accessed on 28 October 2020).
- Package ‘Circular’. Available online: https://cran.r-project.org/web/packages/circular/circular.pdf (accessed on 10 May 2019).
- Package ‘Activity’. Available online: https://cran.r-project.org/web/packages/activity/activity.pdf (accessed on 4 September 2020).
- Kingdon, J. The Kingdon Field Guide to African Mammals, 2nd ed.; Bloomsbury Publishing: London, UK, 2015. [Google Scholar]
- Mori, E.; Nerva, L.; Lovari, S. Reclassification of the Serows and Gorals: The End of a Neverending Story? Mamm. Rev. 2019, 49, 256–262. [Google Scholar] [CrossRef]
- Stockman, A.; Sharpe, L.T. Into the Twilight Zone: The Complexities of Mesopic Vision and Luminous Efficiency. Ophthal. Physl. opt. 2006, 26, 225–239. [Google Scholar] [CrossRef]
- Dubost, G. The Size of African Forest Artiodactyls as Determined by the Vegetation Structure. Afr. J. Ecol. 1979, 17, 1–17. [Google Scholar] [CrossRef]
- Kingdon, J.; Lahm, S.A. Cephalophus silvicultor Yellow-backed Duiker. In Mammals of Africa: Volume VI: Pigs, Hippopotamuses, Chevrotain, Giraffes, Deer and Bovids; Kingdon, J., Hoffmann, M., Eds.; Bloomsbury Publishing: London, UK, 2013; Volume 6, pp. 288–293. [Google Scholar]
- De Sá Alves, L.C.P.; Andriolo, A. Camera traps used on the mastofaunal survey of Araras Biological Reserve. IEF-RJ. Rev. Bras. Zoociências 2005, 7, 231–246. [Google Scholar]
- Lahm, S.A. Ecology and Economics of Human/Wildlife Interaction in North-East Gabon. Ph.D. Thesis, New York University, New York, NY, USA, 1993. [Google Scholar]
- Croes, B.M.; Laurance, W.F.; Lahm, S.A.; Tchignoumba, L.; Alonso, A.; Lee, M.E.; Campbell, P.; Buij, R. The Influence of Hunting on Antipredator Behavior in Central African Monkeys and Duikers. Biotropica 2007, 39, 257–263. [Google Scholar] [CrossRef]
- Gautier-Hion, A.; Gautier, J.-P. Cephalophus ogilbyi crusalbum Grubb 1978, described from coastal Gabon, is quite common in the forêt des abeilles, central Gabon. Rev. Ecol. 1994, 49, 177–180. [Google Scholar]
- Houngbégnon, F.G.A.; Sonké, B.; Vermeulen, C.; Doucet, J.L. State of Knowledge on Duikers (Genera Cephalophus and Philantomba) of Central Africa Rainforests. A Review. Biotechnol. Agron. Soc. Environ. 2019, 23, 104–124. [Google Scholar]
- Veech Research Group in Ecology. Available online: http://ecology.wp.txstate.edu/publications-2/ (accessed on 28 October 2019).




{kind=link}
{kind=link}
{kind=link}
| Sites | Cephalophus callipygus R | C. castaneus R | C. leucogaster R | C. nigrifrons R | C. silvicultor Y | Philantomba congica B |
|---|---|---|---|---|---|---|
| Dja CMR | 70 | 30 | 0 | 1 | 60 | 417 |
| Makalaya CMR | 140 | 30 | 1 | 1 | 32 | 151 |
| Mindourou 1 CMR | 7 | 13 | 0 | 0 | 6 | 42 |
| Mindourou 2 CMR | 27 | 17 | 0 | 1 | 14 | 99 |
| Mindourou 3 CMR | 12 | 13 | 0 | 0 | 4 | 86 |
| Ingolo COG | 1 | 35 | 0 | 0 | 10 | 113 |
| Ngombe COG | 230 | 148 | 1 | 0 | 55 | 545 |
| Bambidie GBN | 100 | 45 | 12 | 1 | 14 | 52 |
| Djoutou GBN | 102 | 31 | 19 | 3 | 19 | 359 |
| Ndambie GBN | 116 | 67 | 25 | 0 | 30 | 77 |
| Ovan GBN | 140 | 36 | 3 | 8 | 35 | 601 |
| Maduda RDC | 0 | 4 | 0 | 27 | 0 | 20 |
| Total | 945 | 469 | 61 | 42 | 279 | 2562 |
| C. callipygus | C. castaneus | C. leucogaster | C. silvicultor | P. congica | |
|---|---|---|---|---|---|
| C. callipygus | plt = 0.999 pgt < 0.001 | plt = 0.991 pgt = 0.025 | plt = 0.999 pgt < 0.001 | plt = 0.999 pgt = 0.001 | |
| C. castaneus | 0.048 | plt = 0.875 pgt = 0.229 | plt = 0.962 pgt = 0.062 | plt = 0.875 pgt = 0.199 | |
| C. leucogaster | 0.017 | 0.007 | plt = 0.105 pgt = 0.956 | plt = 0.958 pgt = 0.141 | |
| C. silvicultor | 0.056 | 0.022 | −0.011 | plt = 0.735 pgt = 0.376 | |
| P. congica | 0.034 | 0.010 | 0.008 | 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Houngbégnon, F.G.A.; Cornelis, D.; Vermeulen, C.; Sonké, B.; Ntie, S.; Fayolle, A.; Fonteyn, D.; Lhoest, S.; Evrard, Q.; Yapi, F.; et al. Daily Activity Patterns and Co-Occurrence of Duikers Revealed by an Intensive Camera Trap Survey across Central African Rainforests. Animals 2020, 10, 2200. https://doi.org/10.3390/ani10122200
Houngbégnon FGA, Cornelis D, Vermeulen C, Sonké B, Ntie S, Fayolle A, Fonteyn D, Lhoest S, Evrard Q, Yapi F, et al. Daily Activity Patterns and Co-Occurrence of Duikers Revealed by an Intensive Camera Trap Survey across Central African Rainforests. Animals. 2020; 10(12):2200. https://doi.org/10.3390/ani10122200
Chicago/Turabian StyleHoungbégnon, Fructueux G. A., Daniel Cornelis, Cédric Vermeulen, Bonaventure Sonké, Stephan Ntie, Adeline Fayolle, Davy Fonteyn, Simon Lhoest, Quentin Evrard, Fabrice Yapi, and et al. 2020. "Daily Activity Patterns and Co-Occurrence of Duikers Revealed by an Intensive Camera Trap Survey across Central African Rainforests" Animals 10, no. 12: 2200. https://doi.org/10.3390/ani10122200