Seminal Plasma Triggers the Differential Expression of the Glucocorticoid Receptor (NR3C1/GR) in the Rabbit Reproductive Tract

 , , ,
, , ,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals
2.3. Experimental Design
2.3.1. Experiment 1: Analysis of Gene Expression Differences in the Does’ Reproductive Tract, at 20 h Post-Mating (Whole Semen) or Seminal Plasma (SP) Infusion (Sperm-Free)
2.3.2. Experiment 2: Analysis of Gene Expression Differences in the Reproductive Tract of Mated Rabbit Females from 10 h Post-mating to up to 72 h Post-mating
2.4. Mating and Semen Collection
2.5. Collection of Tissues and Embryos
2.6. Real Time Quantitative PCR Analyses
2.7. Statistical Analyses
3. Results
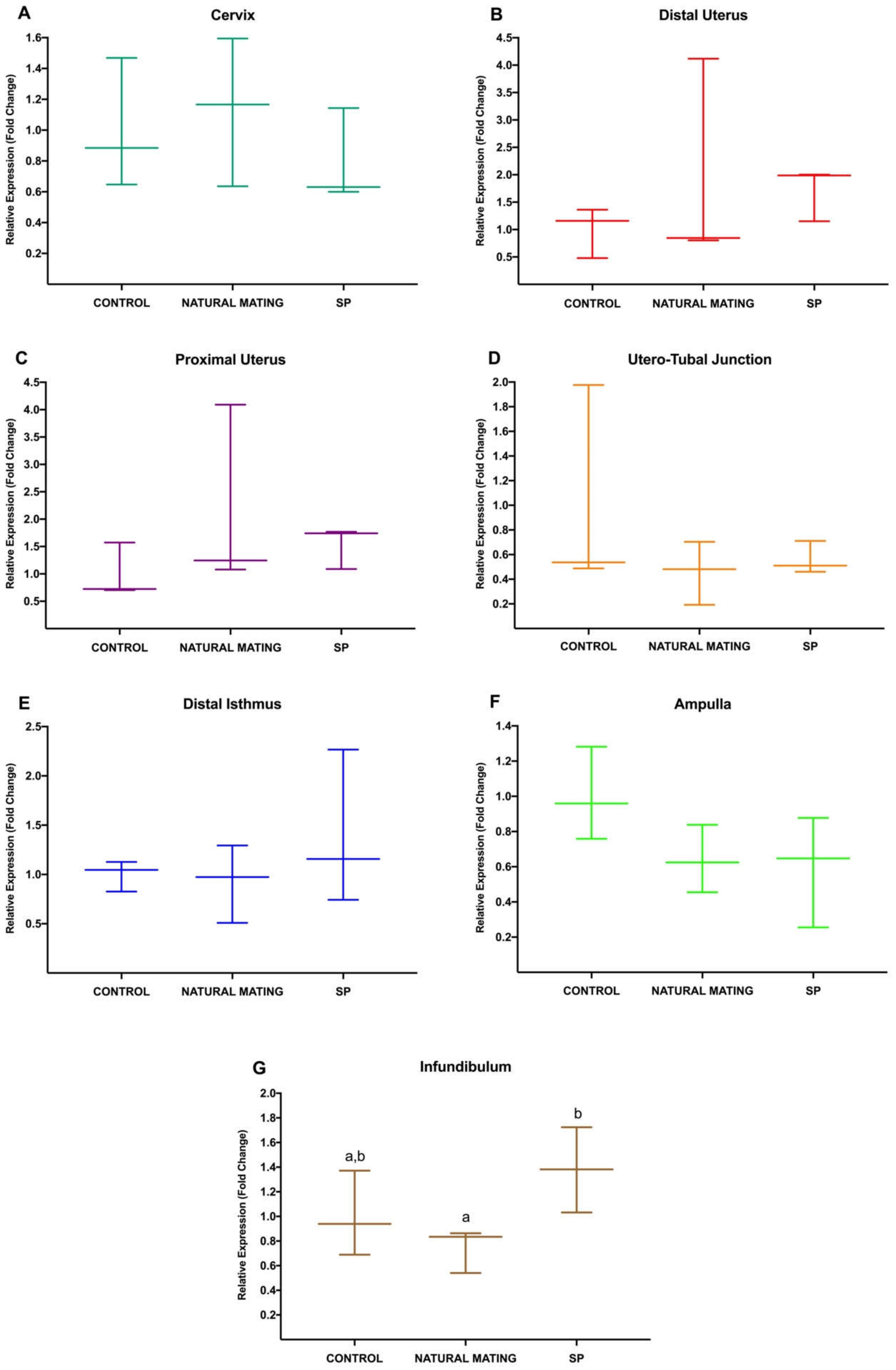
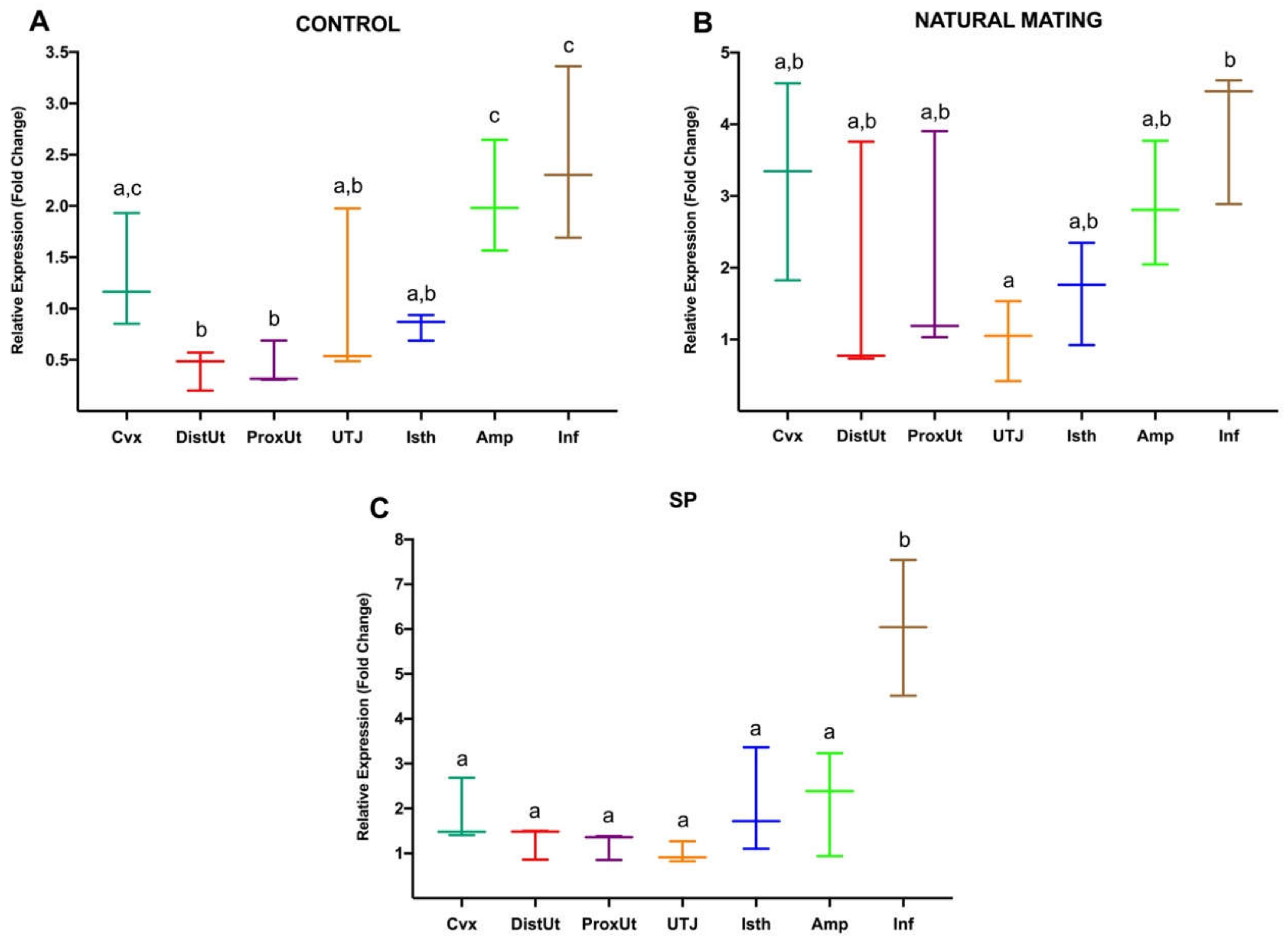
3.1. Experiment 1: Differential Gene Expression in Rabbit Female Reproductive Tract at 20 h after Natural Mating or Infusion of Sperm-Free Seminal Plasma
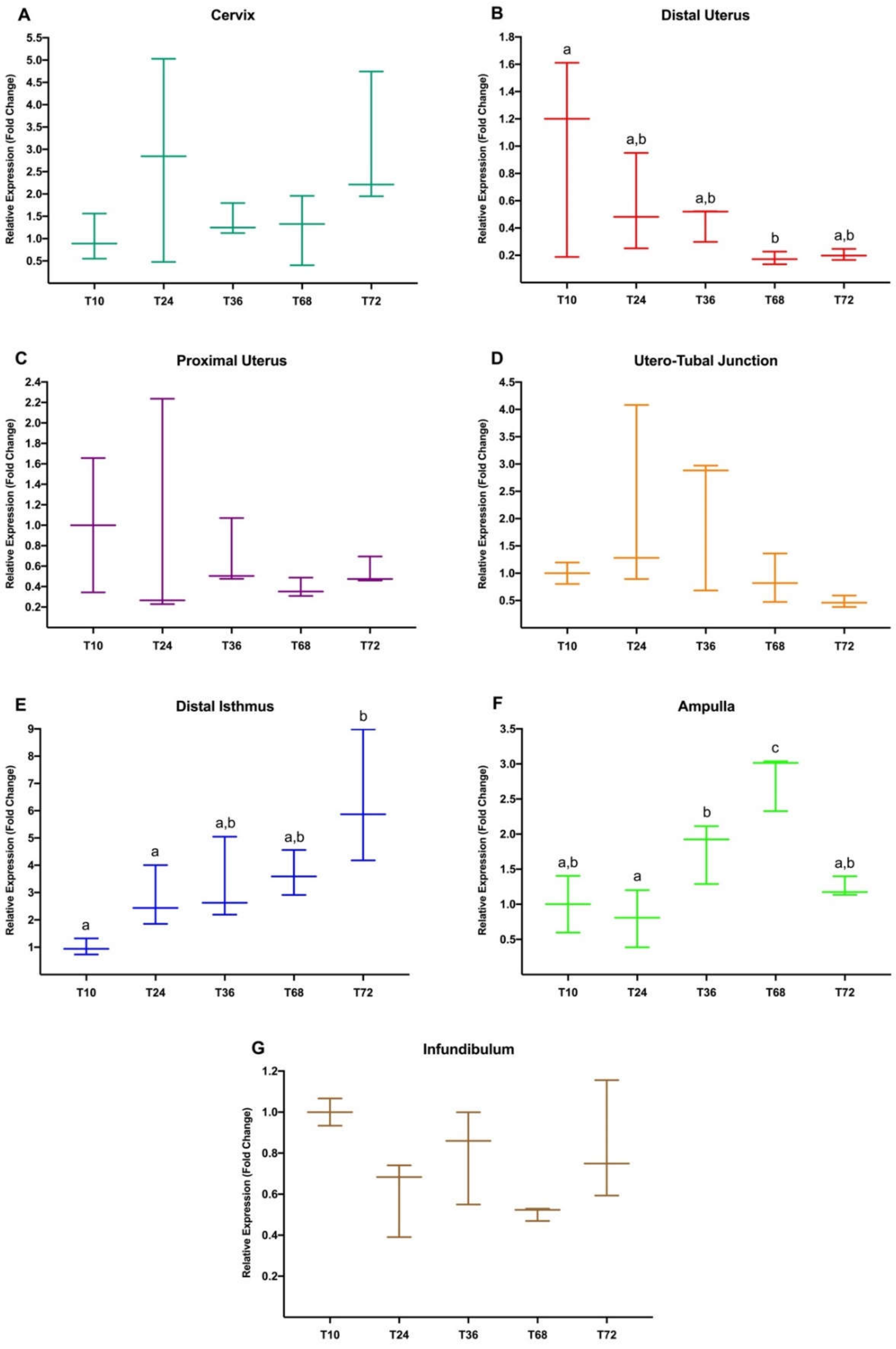
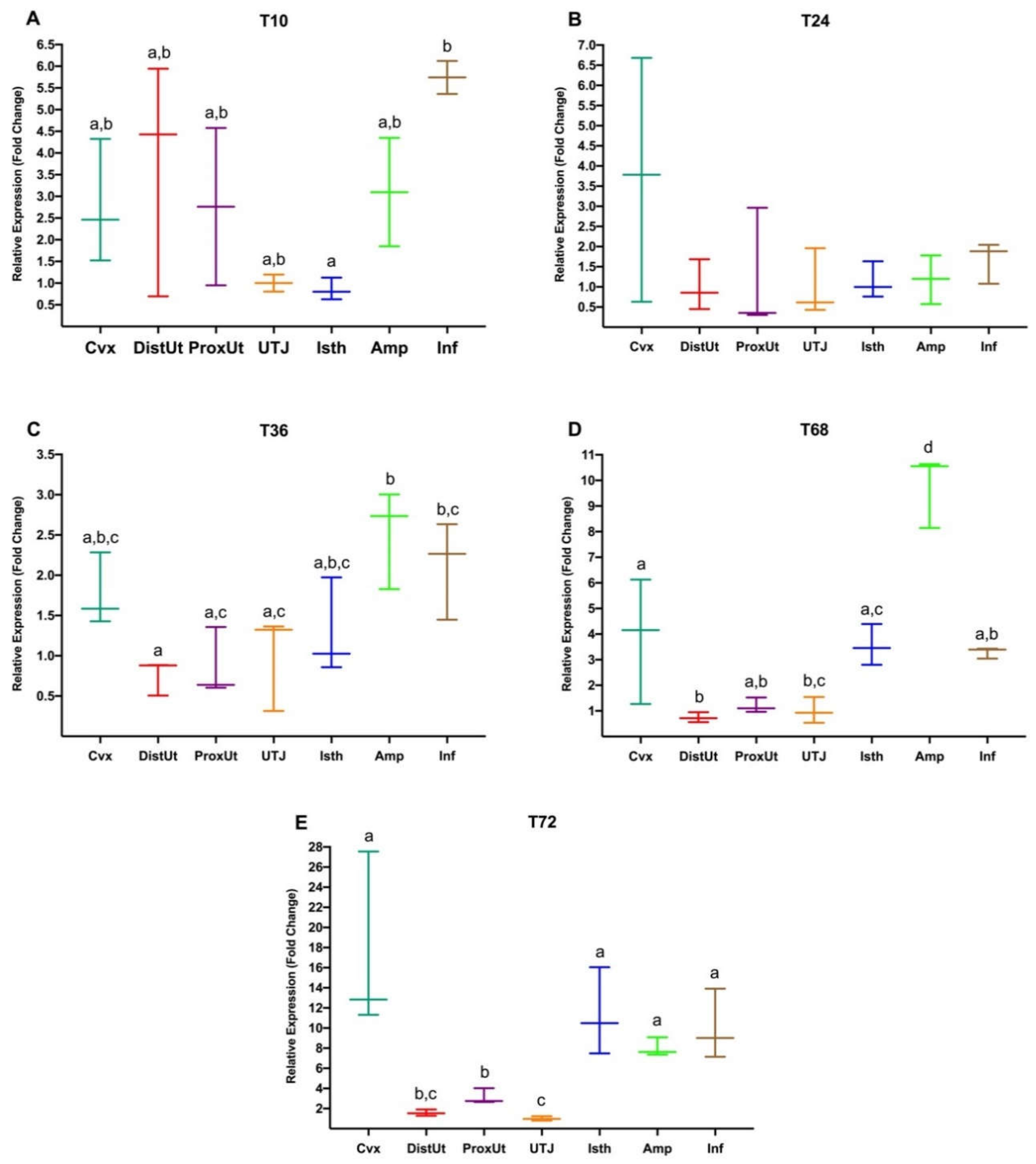
3.2. Experiment 2: Differential Gene Expression in Rabbit Female Reproductive Tract from 10 h to up to 72 h in Response to Natural Mating
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tablado, Z.; Revilla, E.; Palomares, F. Breeding like rabbits: Global patterns of variability and determinants of European wild rabbit reproduction. Ecography 2009, 32, 310–320. [Google Scholar] [CrossRef]
- Roy-Dufresne, E.; Lurgi, M.; Brown, S.C.; Wells, K.; Cooke, B.; Mutze, G.; Peacock, D.; Cassey, P.; Berman, D.; Brook, B.W.; et al. The Australian national rabbit database: 50 year of population monitoring of an invasive species. Ecology 2019, 100, e02750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petraci, M.; Cavani, C. Rabbit meat processing: Historical perspective to future directions. World Rabbit Sci. 2013, 21, 217–226. [Google Scholar]
- Li, S.; Zeng, W.; Li, R.; Hoffman, L.C.; He, Z.; Sun, Q.; Li, H. Rabbit meat production and processing in China. Meat Sci. 2018, 145, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Püschel, B.; Daniel, N.; Bitzer, E.; Blum, M.; Renard, J.P.; Viebahn, C. The rabbit (Oryctolagus cuniculus): A model for mammalian reproduction and early embryology. Cold Spring Harb. Protoc. 2010, 5, 1–6. [Google Scholar]
- Fischer, B.; Chavatte-Palmer, P.; Viebahn, C.; Santos, A.N.; Duranthon, V. Rabbit as a reproductive model for human health. Reproduction 2012, 144, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorge-Neto, P.N.; Luczinski, T.C.; Araújo, G.R.D.; Salomão Júnior, J.A.; Traldi, A.D.S.; Santos, J.A.M.D.; Requena, L.A.; Gianni, M.C.M.; Deco-Souza, T.D.; Pizzutto, C.S.; et al. Can jaguar (Panthera onca) ovulate without copulation? Theriogenology 2020, 147, 57–61. [Google Scholar] [CrossRef]
- Silva, M.; Paiva, L.; Ratto, M.H. Ovulation mechanism in South American Camelids: The active role of β-NGF as the chemical signal eliciting ovulation in llamas and alpacas. Theriogenology 2020, 150, 280–287. [Google Scholar] [CrossRef]
- Adams, G.; Ratto, M.; Silva, M.; Carrasco, R. Ovulation-inducing factor (OIF/NGF) in seminal plasma: A review and update. Reprod. Domest. Anim. 2016, 51, 4–17. [Google Scholar] [CrossRef]
- Pelican, K.M.; Wildt, D.E.; Pukazhenthi, B.; Howard, J.G. Ovarian control for assisted reproduction in the domestic cat and wild felids. Theriogenology 2006, 66, 37–48. [Google Scholar] [CrossRef]
- Garcia-Dominguez, X.; Marco-Jimenez, F.; Viudes-de-Castro, M.P.; Vicente, J.S. Minimally invasive embryo transfer and embryo vitrification at the optimal embryo stage in rabbit model. J. Vis. Exp. 2019, e58055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piles, M.; Tusell, L.; Lavara, R.; Baselga, M. Breeding programmes to improve male reproductive performance and efficiency of insemination dose production in paternal lines: Feasibility and limitations. World Rabbit Sci. 2013, 21, 61–75. [Google Scholar] [CrossRef] [Green Version]
- Bakker, J.; Baum, M.J. Neuroendocrine regulation of GnRH release in induced ovulators. Front. Neuroendocrinol. 2000, 21, 220–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratto, M.H.; Berland, M.; Silva, M.E.; Adams, G.P. New insights of the role of β-NGF in the ovulation mechanism of induced ovulating species. Reproduction 2019, 157, R199–R207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brun, J.M.; Sanchez, A.; Ailloud, E.; Saleil, G.; Theau-Clément, M. Genetic parameters of rabbit semen traits and male fertilising ability. Anim. Reprod. Sci. 2016, 166, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.A. Seminal plasma and male factor signalling in the female reproductive tract. Cell Tissue Res. 2005, 322, 43–52. [Google Scholar] [CrossRef]
- Parada-Bustamante, A.; Oróstica, M.L.; Reuquen, P.; Zuñiga, L.M.; Cardenas, H.; Orihuela, P.A. The role of mating in oviduct biology. Mol. Reprod. Dev. 2016, 83, 875–883. [Google Scholar] [CrossRef]
- Whirledge, S.; Kisanga, E.P.; Taylor, R.N.; Cidlowski, J.A. Pioneer factors FOXA1 and FOXA2 assist selective glucocorticoid receptor signaling in human endometrial cells. Endocrinology 2017, 158, 4076–4092. [Google Scholar] [CrossRef] [Green Version]
- Whirledge, S.; Cidlowski, J.A. Glucocorticoids and reproduction: Traffic control on the road to reproduction. Trends Endocrinol. Metab. 2017, 28, 399–415. [Google Scholar] [CrossRef]
- Wang, J.-C.; Harris, C. Glucocorticoid Signaling from Molecules to Mice to Man; Springer: New York, NY, USA, 2015; ISBN 9781493928941. [Google Scholar]
- Whirledge, S.; Cidlowski, J.A. A role for glucocorticoids in stress-impaired reproduction: Beyond the hypothalamus and pituitary. Endocrinology 2013, 154, 4450–4468. [Google Scholar] [CrossRef] [Green Version]
- Whirledge, S.; Cidlowski, J.A. Glucocorticoids, stress, and fertility. Minerva Endocrinol. 2010, 35, 109–125. [Google Scholar] [PubMed]
- Ratman, D.; Vanden Berghe, W.; Dejager, L.; Libert, C.; Tavernier, J.; Beck, I.M.; de Bosscher, K. How glucocorticoid receptors modulate the activity of other transcription factors: A scope beyond tethering. Mol. Cell. Endocrinol. 2013, 380, 41–54. [Google Scholar] [CrossRef]
- Cain, D.W.; Cidlowski, J.A. Immune regulation by glucocorticoids. Nat. Rev. Immunol. 2017, 17, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Bekhbat, M.; Rowson, S.A.; Neigh, G.N. Checks and balances: The glucocorticoid receptor and NFĸB in good times and bad. Front. Neuroendocrinol. 2017, 46, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Conca, M.; Gardela, J.; Alvarez-Rodríguez, M.; Mogas, T.; López-Béjar, M. Immunofluorescence analysis of NR3C1 receptor following cortisol exposure during bovine in vitro oocyte maturation. Anim. Reprod. Sci. 2019, 16, 753. [Google Scholar]
- Yang, J.-G.; Chen, W.-Y.; Li, P.S. Effects of glucocorticoids on maturation of pig oocytes and their subsequent fertilizing capacity in vitro1. Biol. Reprod. 1999, 60, 929–936. [Google Scholar] [CrossRef] [Green Version]
- Whirledge, S.D.; Oakley, R.H.; Myers, P.H.; Lydon, J.P.; DeMayo, F.; Cidlowski, J.A. Uterine glucocorticoid receptors are critical for fertility in mice through control of embryo implantation and decidualization. Proc. Natl. Acad. Sci. USA 2015, 112, 15166–15171. [Google Scholar] [CrossRef] [Green Version]
- Wochnik, G.M.; Rüegg, J.; Abel, G.A.; Schmidt, U.; Holsboer, F.; Rein, T. FK506-binding proteins 51 and 52 differentially regulate dynein interaction and nuclear translocation of the glucocorticoid receptor in mammalian cells. J. Biol. Chem. 2005, 280, 4609–4616. [Google Scholar] [CrossRef] [Green Version]
- Petta, I.; Dejager, L.; Ballegeer, M.; Lievens, S.; Tavernier, J.; de Bosscher, K.; Libert, C. The interactome of the glucocorticoid receptor and its influence on the actions of glucocorticoids in combatting inflammatory and infectious diseases. Microbiol. Mol. Biol. Rev. 2016, 80, 495–522. [Google Scholar] [CrossRef] [Green Version]
- Simmons, R.M.; Satterfield, M.C.; Welsh, T.H.; Bazer, F.W.; Spencer, T.E. HSD11B1, HSD11B2, PTGS2, and NR3C1 expression in the peri-implantation ovine uterus: Effects of pregnancy, progesterone, and interferon Tau1. Biol. Reprod. 2010, 82, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Siemieniuch, M.J.; Majewska, M.; Takahashi, M.; Sakatani, M.; Łukasik, K.; Okuda, K.; Skarzynski, D.J. Are glucocorticoids auto- and/or paracrine factors in early bovine embryo development and implantation? Reprod. Biol. 2010, 10, 249–256. [Google Scholar] [CrossRef]
- Majewska, M.; Lee, H.Y.; Tasaki, Y.; Acosta, T.J.; Szostek, A.Z.; Siemieniuch, M.; Okuda, K.; Skarzynski, D.J. Is cortisol a modulator of interferon tau action in the endometrium during early pregnancy in cattle? J. Reprod. Immunol. 2012, 93, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Lindblom, B.; Hamberger, L.; Ljung, B. Contractile patterns of isolated oviductal smooth muscle under different hormonal conditions. Fertil. Steril. 1980, 33, 283–287. [Google Scholar] [CrossRef]
- Wånggren, K.; Stavreus-Evers, A.; Olsson, C.; Andersson, E.; Gemzell-Danielsson, K. Regulation of muscular contractions in the human Fallopian tube through prostaglandins and progestagens. Hum. Reprod. 2008, 23, 2359–2368. [Google Scholar] [CrossRef] [Green Version]
- Andersen, Y.C. Possible new mechanism of cortisol action in female reproductive organs: Physiological implications of the free hormone hypothesis. J. Endocrinol. 2002, 173, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Conca, M.; Gardela, J.; Martínez, C.A.; Wright, D.; López-Bejar, M.; Rodríguez-Martínez, H.; Álvarez-Rodríguez, M. Natural mating differentially triggers expression of glucocorticoid receptor (NR3C1)-related genes in the preovulatory porcine female reproductive tract. Int. J. Mol. Sci. 2020, 21, 4437. [Google Scholar] [CrossRef]
- Gong, S.; Sun, G.-Y.; Zhang, M.; Yuan, H.-J.; Zhu, S.; Jiao, G.-Z.; Luo, M.-J.; Tan, J.-H. Mechanisms for the species difference between mouse and pig oocytes in their sensitivity to glucorticoids. Biol. Reprod. 2017, 96, 1019–1030. [Google Scholar] [CrossRef]
- Scarlet, D.; Ille, N.; Ertl, R.; Alves, B.G.; Gastal, G.D.A.; Paiva, S.O.; Gastal, M.O.; Gastal, E.L.; Aurich, C. Glucocorticoid metabolism in equine follicles and oocytes. Domest. Anim. Endocrinol. 2017, 59, 11–22. [Google Scholar] [CrossRef]
- Da Costa, N.N.; Brito, K.N.L.; Santana, P.D.P.B.; Cordeiro, M.D.S.; Silva, T.V.G.; Santos, A.X.; Ramos, P.D.C.; Santos, S.D.S.D.; King, W.A.; Miranda, M.D.S.; et al. Effect of cortisol on bovine oocyte maturation and embryo development in vitro. Theriogenology 2016, 85, 323–329. [Google Scholar] [CrossRef]
- Ruiz-Conca, M.; Alvarez-Rodriguez, M.; Mogas, T.; Gardela, J. NR3C1 expression in response to cortisol during bovine in vitro oocyte maturation. In Proceedings of the 15 th International Congress of the Spanish Society for Animal Reproduction (AERA), Toledo, Spain, 7–9 November 2019; p. 129. [Google Scholar]
- Carneiro, M.; Piorno, V.; Rubin, C.-J.; Alves, J.M.; Ferrand, N.; Alves, P.C.; Andersson, L. Candidate genes underlying heritable differences in reproductive seasonality between wild and domestic rabbits. Anim. Genet. 2015, 46, 418–425. [Google Scholar] [CrossRef]
- Gardela, J.; Jauregi-Miguel, A.; Martinez, C.A.; Rodriguez-Martinez, H.; Lopez-Bejar, M.; Alvarez-Rodriguez, M. Semen modulates the expression of NGF, ABHD2, VCAN, and CTEN in the reproductive tract of female rabbits. Genes 2020, 11, 758. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Computing, R.F.S. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Pinheiro, J.C.; DebRoy, S.S.; Sarkar, D.; Bates, D.M. Nlme: Linear and nonlinear mixed effects models. R Package Version 2020, 3, 111. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dukelow, W.R.; Williams, W.L. Survival of capacitated spermatozoa in the oviduct of the rabbit. Reproduction 1967, 14, 477–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbot, P.; Geiske, C.; Knoll, M. Oocyte pickup by the mammalian oviduct. Mol. Biol. Cell 1999, 10, 5–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schjenken, J.E.; Robertson, S.A. The female response to seminal fluid. Physiol. Rev. 2020, 100, 1077–1117. [Google Scholar] [CrossRef]
- Robertson, S.A. Seminal fluid signaling in the female reproductive tract: Lessons from rodents and pigs. J. Anim. Sci. 2007, 85, E36–E44. [Google Scholar] [CrossRef]
- O’Leary, S.; Jasper, M.J.; Warnes, G.M.; Armstrong, D.T.; Robertson, S.A. Seminal plasma regulates endometrial cytokine expression, leukocyte recruitment and embryo development in the pig. Reproduction 2004, 128, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, D.J.; Macpherson, A.M.; Tremellen, K.P.; Robertson, S.A. Seminal plasma differentially regulates inflammatory cytokine gene expression in human cervical and vaginal epithelial cells. Mol. Hum. Reprod. 2007, 13, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Castellini, C.; Mourvaki, E.; Cardinali, R.; Collodel, G.; Lasagna, E.; del Vecchio, M.T.; dal Bosco, A. Secretion patterns and effect of prostate-derived granules on the sperm acrosome reaction of rabbit buck. Theriogenology 2012, 78, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, M.J.B.; Arruda-Alencar, J.M.; Martins, J.A.M.; Viana, A.G.A.; Viana Neto, A.M.; Rêgo, J.P.A.; Oliveira, R.V.; Lobo, M.; Moreira, A.C.O.; Moreira, R.A.; et al. Major seminal plasma proteome of rabbits and associations with sperm quality. Theriogenology 2019, 128, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Schjenken, J.E.; Robertson, S.A. Seminal fluid and immune adaptation for pregnancy - comparative biology in mammalian species. Reprod. Domest. Anim. 2014, 49, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.A.; Cambra, J.M.; Gil, M.A.; Parrilla, I.; Alvarez-Rodriguez, M.; Rodriguez-Martinez, H.; Cuello, C.; Martinez, E.A. Seminal plasma induces overexpression of genes associated with embryo development and implantation in day-6 porcine blastocysts. Int. J. Mol. Sci. 2020, 21, 3662. [Google Scholar] [CrossRef]
- Martinez, C.A.; Cambra, J.M.; Parrilla, I.; Roca, J.; Ferreira-Dias, G.; Pallares, F.J.; Lucas, X.; Vazquez, J.M.; Martinez, E.A.; Gil, M.A.; et al. Seminal plasma modifies the transcriptional pattern of the endometrium and advances embryo development in pigs. Front. Vet. Sci. 2019, 6, 465. [Google Scholar] [CrossRef] [Green Version]
- Waberski, D. Effects of semen components on ovulation and fertilization. J. Reprod. Fertil. Suppl. 1997, 52, 105. [Google Scholar]
- Maranesi, M.; Petrucci, L.; Leonardi, L.; Piro, F.; Rebollar, P.G.; Millán, P.; Cocci, P.; Vullo, C.; Parillo, F.; Moura, A.; et al. New insights on a NGF-mediated pathway to induce ovulation in rabbits (Oryctolagus cuniculus). Biol. Reprod. 2018, 98, 634–643. [Google Scholar] [CrossRef] [Green Version]
- Kershaw-Young, C.M.; Druart, X.; Vaughan, J.; Maxwell, W.M.C. β-Nerve growth factor is a major component of alpaca seminal plasma and induces ovulation in female alpacas. Reprod. Fertil. Dev. 2012, 24, 1093–1097. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.; Niño, A.; Guerra, M.; Letelier, C.; Valderrama, X.P.; Adams, G.P.; Ratto, M.H. Is an ovulation-inducing factor (OIF) present in the seminal plasma of rabbits? Anim. Reprod. Sci. 2011, 127, 213–221. [Google Scholar] [CrossRef]
- Garcia-Garcia, R.M.; Masdeu, M.d.M.; Sanchez Rodriguez, A.; Millan, P.; Arias-Alvarez, M.; Sakr, O.G.; Bautista, J.M.; Castellini, C.; Lorenzo, P.L.; Rebollar, P.G. B-nerve growth factor identification in male rabbit genital tract and seminal plasma and its role in ovulation induction in rabbit does. Ital. J. Anim. Sci. 2018, 17, 442–453. [Google Scholar] [CrossRef]
- Bomsel-Helmreich, O.; Huyen, L.V.N.; Durand-Gasselin, I. Effects of varying doses of HCG on the evolution of preovulatory rabbit follicles and oocytes. Hum. Reprod. 1989, 4, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Duffy, D.M.; Ko, C.; Jo, M.; Brannstrom, M.; Curry, T.E. Ovulation: Parallels With inflammatory processes. Endocr. Rev. 2019, 40, 369–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, J.A.S.; Liu, Z.; Shimada, M. Immune-like mechanisms in ovulation. Trends Endocrinol. Metab. 2008, 19, 191–196. [Google Scholar] [CrossRef]
- Espey, L.L. Ovulation as an inflammatory reaction—A hypothesis. Biol. Reprod. 1980, 22, 73–106. [Google Scholar] [CrossRef] [PubMed]
- Gross, K.L.; Cidlowski, J.A. Tissue-specific glucocorticoid action: A family affair. Trends Endocrinol. Metab. 2008, 19, 331–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, M.; Lamont, M.C.; van den Driesche, S.; Mary, N.; Thong, K.J.; Hillier, S.G.; Duncan, W.C. Role of luteal glucocorticoid metabolism during maternal recognition of pregnancy in women. Endocrinology 2007, 148, 5769–5779. [Google Scholar] [CrossRef] [Green Version]
- Harlow, C.R.; Jenkins, J.M.; Winston, R.M.L. Increased follicular fluid total and free cortisol levels during the luteinizing hormone surge. Fertil. Steril. 1997, 68, 48–53. [Google Scholar] [CrossRef]
- Thurston, L.M.; Abayasekara, D.R.E.; Michael, A.E. 11β-hydroxysteroid dehydrogenase expression and activities in bovine granulosa cells and corpora lutea implicate corticosteroids in bovine ovarian physiology. J. Endocrinol. 2007, 193, 299–310. [Google Scholar] [CrossRef]
- Chin, E.; Jonas, K.C.; Abayasekara, D.R.E.; Michael, A.E. Expression of 11β-hydroxysteroid dehydrogenase (11βHSD) proteins in luteinizing human granulosa-lutein cells. BioScientifica 2003, 178, 127–135. [Google Scholar]
- Gong, H.; Jarzynka, M.J.; Cole, T.J.; Jung, H.L.; Wada, T.; Zhang, B.; Gao, J.; Song, W.C.; DeFranco, D.B.; Cheng, S.Y.; et al. Glucocorticoids antagonize estrogens by glucocorticoid receptor-mediated activation of estrogen sulfotransferase. Cancer Res. 2008, 68, 7386–7393. [Google Scholar] [CrossRef] [Green Version]
- Rhen, T.; Grissom, S.; Afshari, C.; Cidlowski, J.A. Dexamethasone blocks the rapid biological effects of 17β-estradiol in the rat uterus without antagonizing its global genomic actions. FASEB J. 2003, 17, 1849–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witorsch, R.J. Effects of elevated glucocorticoids on reproduction and development: Relevance to endocrine disruptor screening. Crit. Rev. Toxicol. 2016, 46, 420–436. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.S.; Majeska, R.J.; Ma, Y.; LaChapelle, L.; Guller, S. Steroid regulation of human placental integrins: Suppression of α2 integrin expression in cytotrophoblasts by glucocorticoids. Endocrinology 1999, 140, 3904–3908. [Google Scholar] [CrossRef] [PubMed]
- Bitman, J.; Cecil, H.C. Differential inhibition by cortisol of estrogen-stimulated uterine responses. Endocrinology 1967, 80, 423–429. [Google Scholar] [CrossRef]
- Johnson, D.C.; Dey, S.K. Role of histamine in implantation: Dexamethasone inhibits estradiol-induced implantation in the rat. Biol. Reprod. 1980, 22, 1136–1141. [Google Scholar] [CrossRef] [PubMed]
- Rhen, T.; Cidlowski, J.A. Estrogens and glucocorticoids have opposing effects on the amount and latent activity of complement proteins in the rat uterus. Biol. Reprod. 2006, 74, 265–274. [Google Scholar] [CrossRef]
- Higgins, B.D.; Kane, M.T. Inositol transport in mouse oocytes and preimplantation embryos: Effects of mouse strain, embryo stage, sodium and the hexose transport inhibitor, phloridzin. Reproduction-Cambridge 2003, 125, 111–118. [Google Scholar] [CrossRef]
- Greenwald, G.S. A study of the transport of ova through the rabbit oviduct. Fertil. Steril. 1961, 12, 80–95. [Google Scholar] [CrossRef]
- Cole, H.H.; Cupps, P.T. Reproduction in Domestic Animals, 2nd ed.; Academic Press: New York, NY, USA, 1969. [Google Scholar]
- Barton, B.E.; Herrera, G.G.; Anamthathmakula, P.; Rock, J.K.; Willie, A.M.; Harris, E.A.; Takemaru, K.I.; Winuthayanon, W. Roles of steroid hormones in oviductal function. Reproduction 2020, 159, R125–R137. [Google Scholar] [CrossRef]
- Goodwin, J.E.; Zhang, J.; Geller, D.S. A critical role for vascular smooth muscle in acute glucocorticoid-lnduced hypertension. J. Am. Soc. Nephrol. 2008, 19, 1291–1299. [Google Scholar] [CrossRef] [Green Version]
- Rog-Zielinska, E.A.; Thomson, A.; Kenyon, C.J.; Brownstein, D.G.; Moran, C.M.; Szumska, D.; Michailidou, Z.; Richardson, J.; Owen, E.; Watt, A.; et al. Glucocorticoid receptor is required for foetal heart maturation. Hum. Mol. Genet. 2013, 22, 3269–3282. [Google Scholar] [CrossRef] [Green Version]
- Spilman, C.H.; Harper, M.J.K. Effects of prostaglandins on oviductal motility and egg transport. Gynecol. Obstet. Investig. 1975, 6, 186–205. [Google Scholar] [CrossRef] [PubMed]
- Blair, W.D.; Beck, L.R. In vivo effects of prostaglandin F2α and E2 on contractility and diameter of the rabbit oviduct using intraluminal transducers. Biol. Reprod. 1977, 16, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Peeters, B.W.M.M.; Ruigt, G.S.F.; Craighead, M.; Kitchener, P. Differential effects of the new glucocorticoid receptor antagonist ORG 34517 and RU486 (mifepristone) on glucocorticoid receptor nuclear translocation in the AtT20 cell line. Ann. N. Y. Acad. Sci. 2008, 1148, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Xi, M.; Yang, L.Z.; Hsieh, C.M.; Y Shen, L.L. The effect of RU486 on contractility of rabbit oviduct smooth muscle. Sheng Li Xue Bao Acta Physiol. Sin. 1996, 48, 277–283. [Google Scholar]
- Fernandez-Fuertes, B.; Rodríguez-Alonso, B.; Sánchez, J.M.; Simintiras, C.A.; Lonergan, P.; Rizos, D. Looking at the big picture: Understanding how the oviduct’s dialogue with gametes and the embryo shapes reproductive success. Anim. Reprod. 2018, 15, 751–764. [Google Scholar] [CrossRef]
- Tetsuka, M.; Tanakadate, M. Activation of hsd11b1 in the bovine cumulus-oocyte complex during ivm and ivf. Endocr. Connect. 2019, 8, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.Y. Effect of glucocorticoids on spontaneous and follicle-stimulating hormone induced oocyte maturation in mouse oocytes during culture. J. Steroid Biochem. Mol. Biol. 2003, 85, 423–427. [Google Scholar] [CrossRef]
- Scammell, J.G.; Denny, W.B.; Valentine, D.L.; Smiths, D.F. Overexpression of the FK506-binding immunophilin FKBP51 is the common cause of glucocorticoid resistance in three New World primates. Gen. Comp. Endocrinol. 2001, 124, 152–165. [Google Scholar] [CrossRef]
- Ratajczak, T.; Cluning, C.; Ward, B.K. Steroid receptor-associated immunophilins: A gateway to steroid signalling. Clin. Biochem. Rev. 2015, 36, 31–52. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Product Size (bp) | Efficiency (%) |
|---|---|---|---|
| NR3C1 | F: CACAACTCACCCCAACACTG | 212 | 89.6 |
| R: CAGGAGGGTCATTTGGTCAT | |||
| β-ACTIN | F: commercial, not available | 120 | 88.6 |
| R: commercial, not available |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Conca, M.; Gardela, J.; Jauregi-Miguel, A.; Martinez, C.A.; Rodríguez-Martinez, H.; López-Béjar, M.; Alvarez-Rodriguez, M. Seminal Plasma Triggers the Differential Expression of the Glucocorticoid Receptor (NR3C1/GR) in the Rabbit Reproductive Tract. Animals 2020, 10, 2158. https://doi.org/10.3390/ani10112158
Ruiz-Conca M, Gardela J, Jauregi-Miguel A, Martinez CA, Rodríguez-Martinez H, López-Béjar M, Alvarez-Rodriguez M. Seminal Plasma Triggers the Differential Expression of the Glucocorticoid Receptor (NR3C1/GR) in the Rabbit Reproductive Tract. Animals. 2020; 10(11):2158. https://doi.org/10.3390/ani10112158
Chicago/Turabian StyleRuiz-Conca, Mateo, Jaume Gardela, Amaia Jauregi-Miguel, Cristina A. Martinez, Heriberto Rodríguez-Martinez, Manel López-Béjar, and Manuel Alvarez-Rodriguez. 2020. "Seminal Plasma Triggers the Differential Expression of the Glucocorticoid Receptor (NR3C1/GR) in the Rabbit Reproductive Tract" Animals 10, no. 11: 2158. https://doi.org/10.3390/ani10112158