Assessment of the Stress Response in North American Deermice: Laboratory and Field Validation of Two Enzyme Immunoassays for Fecal Corticosterone Metabolites


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Acquisition and Husbandry of Laboratory Deermice
2.2. Fecal Sample Collection
2.3. Acclimation Period
2.4. Dexamethasone Suppression Challenge
2.5. ACTH Stimulation Challenge
2.6. Field Validation
2.7. Processing of Feces and Extraction of FCMs
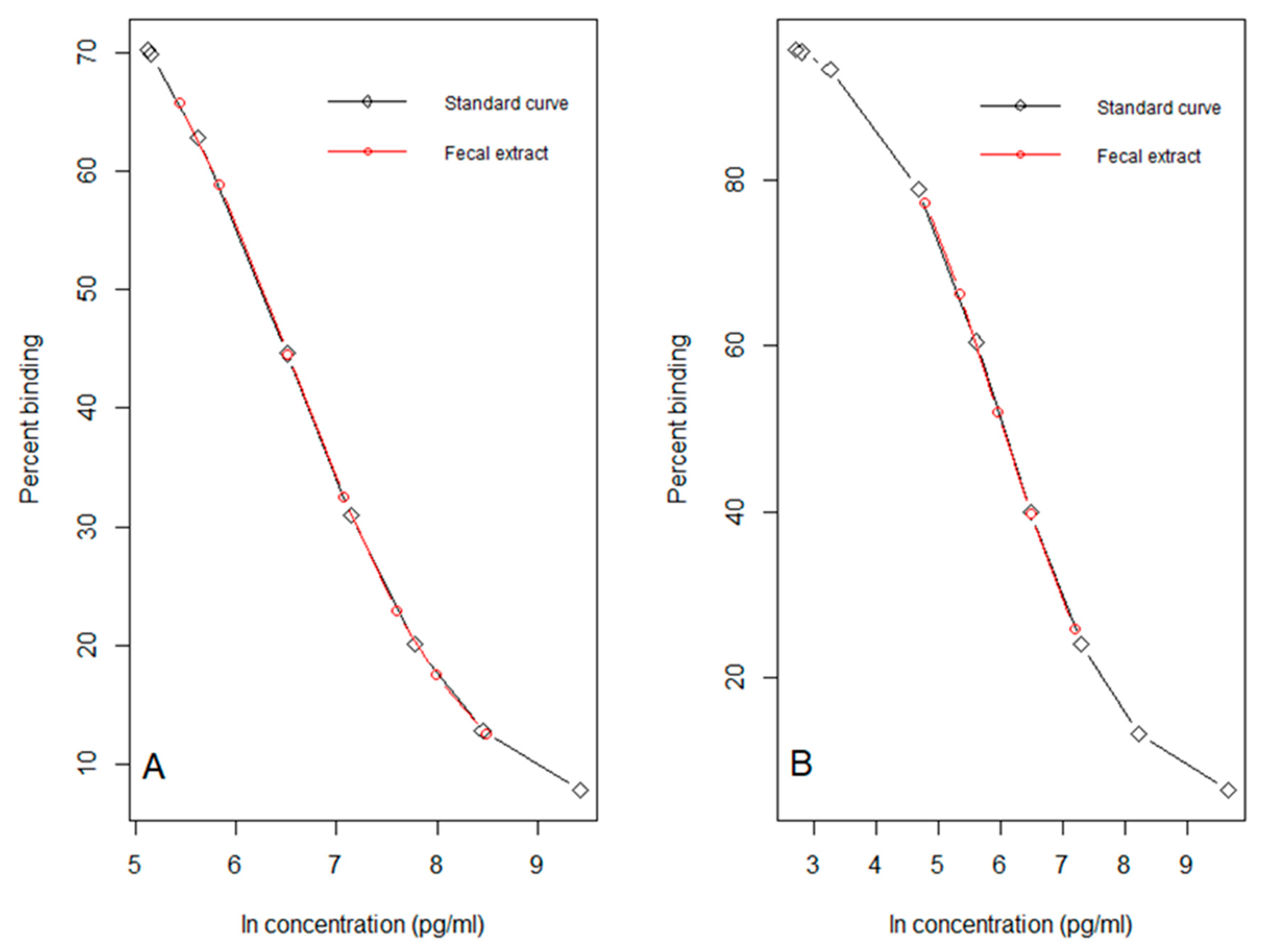
2.8. Immunoassay Methods
2.9. Statistical Analyses
3. Results
3.1. Acclimation
3.2. Dexamethasone Suppression Test
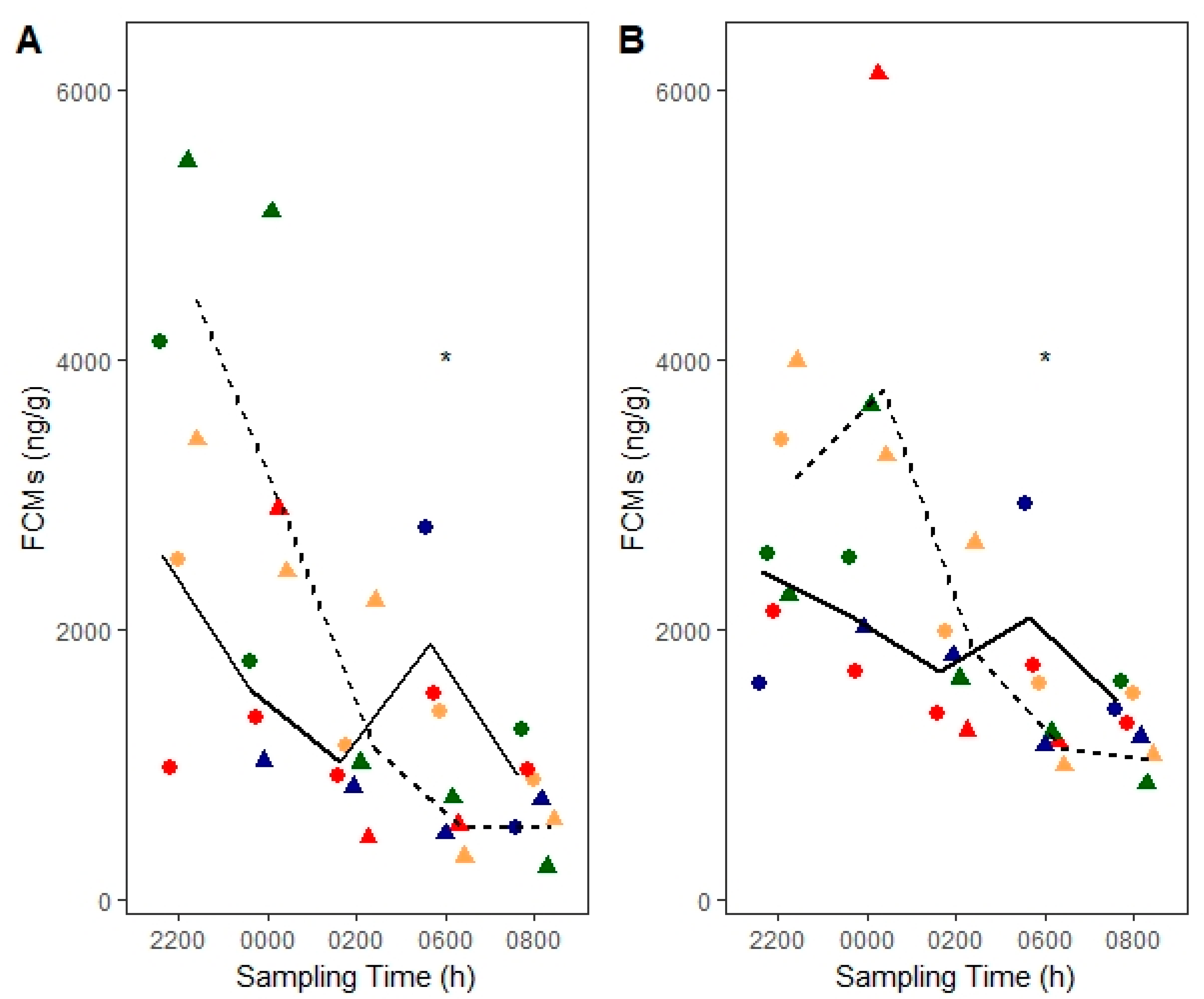
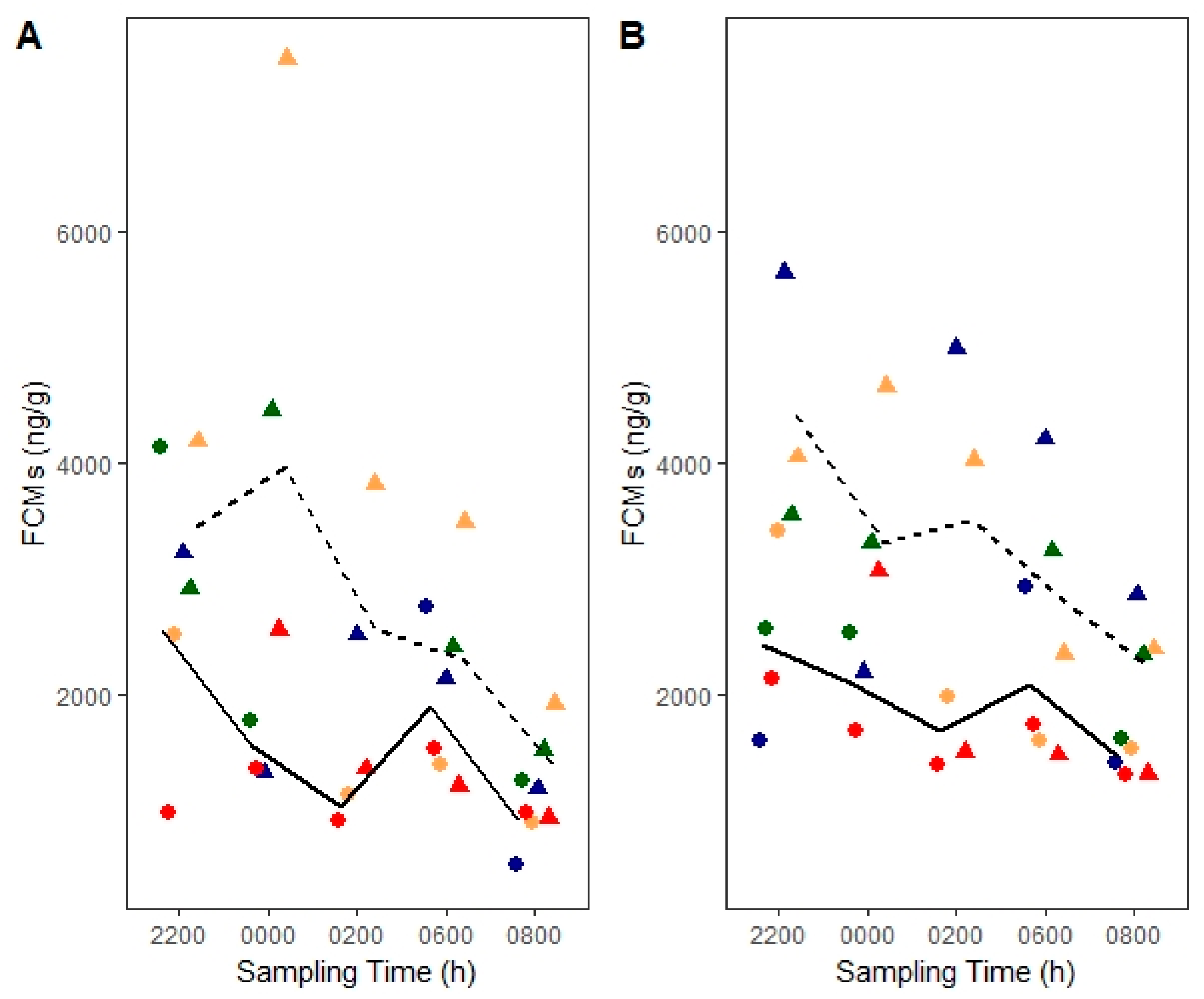
3.3. ACTH Stimulation Test
3.4. Field Validation
4. Discussion
4.1. Diurnal Rhythm and Sex Effects
4.2. Suppression of Adrenocortical Activity
4.3. Stimulation of Adrenocortical Activity
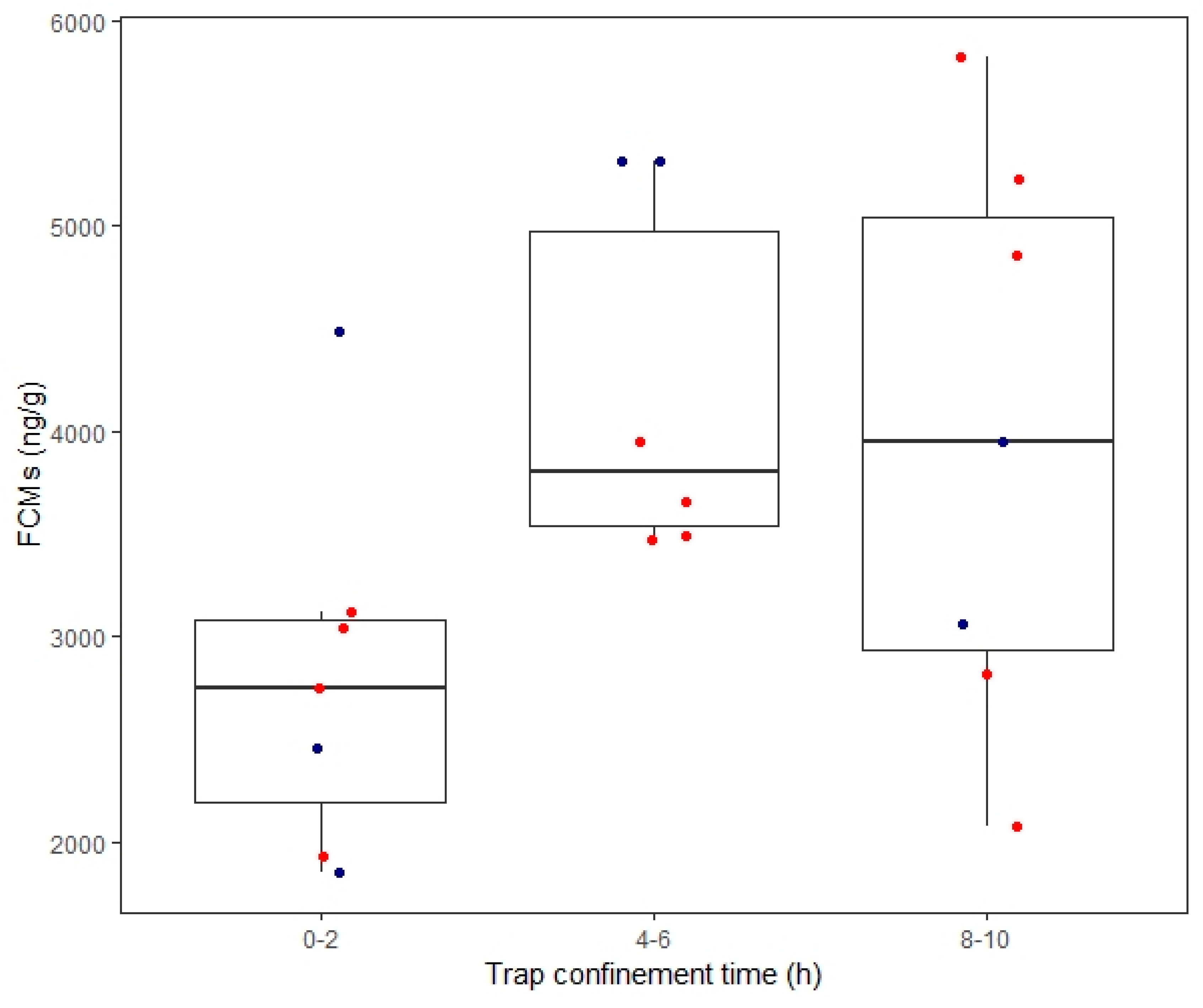
4.4. Trap-Induced Effects on FCMs
4.5. Drying Effects on FCMs
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sheriff, M.J.; Dantzer, B.; Delehanty, B.; Palme, R.; Boonstra, R. Measuring stress in wildlife: Techniques for quantifying glucocorticoids. Oecologia 2011, 166, 869–887. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, B.; Fletcher, Q.E.; Boonstra, R.; Sheriff, M.J. Measures of physiological stress: A transparent or opaque window into the status, management and conservation of species? Conserv. Physiol. 2014, 2, cou023. [Google Scholar] [CrossRef] [PubMed]
- Touma, C.; Palme, R. Measuring fecal glucocorticoid metabolites in mammals and birds: The importance of validation. Ann. NY Acad. Sci. 2005, 1046, 54–74. [Google Scholar] [CrossRef]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Fanson, K.V.; Best, E.C.; Bunce, A.; Fanson, B.G.; Hogan, L.A.; Keeley, T.; Narayan, E.J.; Palme, R.; Parrott, M.L.; Sharp, T.M. One size does not fit all: Monitoring faecal glucocorticoid metabolites in marsupials. Gen. Comp. Endocrinol. 2017, 244, 146–156. [Google Scholar] [CrossRef]
- Heistermann, M.; Palme, R.; Ganswindt, A. Comparison of different enzyme immunoassays for assessment of adrenocortical activity in primates based on fecal analysis. Am. J. Primatol. 2006, 68, 257. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Prescott, J.; Cogswell, A.C.; Oko, L.; Mirowsky-Garcia, K.; Galvez, A.P.; Hjelle, B. Regulatory T cell-like responses in deer mice persistently infected with Sin Nombre virus. Proc. Natl. Acad. Sci. USA 2007, 104, 15496–15501. [Google Scholar] [CrossRef] [Green Version]
- Mahalingam, S.; McClelland, G.B.; Scott, G.R. Evolved changes in the intracellular distribution and physiology of muscle mitochondria in high-altitude native deer mice. J. Physiol. (Lond.) 2017, 595, 4785–4801. [Google Scholar] [CrossRef]
- Luis, A.D.; Douglass, R.J.; Mills, J.N.; Bjørnstad, O.N. Environmental fluctuations lead to predictability in Sin Nombre hantavirus outbreaks. Ecology 2015, 96, 1691–1701. [Google Scholar] [CrossRef]
- Harper, J.M.; Austad, S.N. Fecal glucocorticoids: A noninvasive method of measuring adrenal activity in wild and captive rodents. Physiol. Biochem. Zool. 2000, 73, 12–22. [Google Scholar] [CrossRef]
- Hayssen, V.; Harper, J.; DeFina, R. Fecal corticosteroids in agouti and non-agouti deer mice (Peromyscus maniculatus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 132, 439–446. [Google Scholar] [CrossRef]
- Pedersen, A.B.; Greives, T.J. The interaction of parasites and resources cause crashes in a wild mouse population. J. Anim. Ecol. 2008, 77, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Fredebaugh-Siller, S.; Suski, C.; Zuckerman, Z.; Schooley, R. Ecological correlates of stress for a habitat generalist in a biofuels landscape. Can. J. Zool. 2013, 91, 853–858. [Google Scholar] [CrossRef]
- Bradley, E.L.; Terman, C.R. A comparison of the adrenal histology, reproductive condition, and serum corticosterone concentrations of prairie deermice (Peromyscus maniculatus bairdii) in captivity. J. Mammal. 1981, 62, 353–361. [Google Scholar] [CrossRef]
- Touma, C.; Sachser, N.; Möstl, E.; Palme, R. Effects of sex and time of day on metabolism and excretion of corticosterone in urine and feces of mice. Gen. Comp. Endocrinol. 2003, 130, 267–278. [Google Scholar] [CrossRef]
- Wielebnowski, N.; Watters, J. Applying fecal endocrine monitoring to conservation and behavior studies of wild mammals: Important considerations and preliminary tests. Isr. J. Ecol. Evol. 2007, 53, 439–460. [Google Scholar] [CrossRef]
- Delehanty, B.; Boonstra, R. Impact of live trapping on stress profiles of Richardson’s ground squirrel (Spermophilus richardsonii). Gen. Comp. Endocrinol. 2009, 160, 176–182. [Google Scholar] [CrossRef]
- Bosson, C.; Islam, Z.; Boonstra, R. The impact of live trapping and trap model on the stress profiles of North American red squirrels. J. Zool. 2012, 288, 159–169. [Google Scholar] [CrossRef]
- Millspaugh, J.J.; Washburn, B.E. Use of fecal glucocorticoid metabolite measures in conservation biology research: Considerations for application and interpretation. Gen. Comp. Endocrinol. 2004, 138, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Palme, R.; Rettenbacher, S.; Touma, C.; El-Bahr, S.; Möstl, E. Stress hormones in mammals and birds: Comparative aspects regarding metabolism, excretion, and noninvasive measurement in fecal samples. Ann. NY Acad. Sci. 2005, 1040, 162–171. [Google Scholar] [CrossRef]
- Harper, J.M.; Austad, S.N. Effect of capture and season on fecal glucocorticoid levels in deer mice (Peromyscus maniculatus) and red-backed voles (Clethrionomys gapperi). Gen. Comp. Endocrinol. 2001, 123, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Montiglio, P.; Pelletier, F.; Palme, R.; Garant, D.; Réale, D.; Boonstra, R. Noninvasive monitoring of fecal cortisol metabolites in the eastern chipmunk (Tamias striatus): Validation and comparison of two enzyme immunoassays. Physiol. Biochem. Zool. 2012, 85, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, Q.E.; Boonstra, R. Impact of live trapping on the stress response of the meadow vole (Microtus pennsylvanicus). J. Zool. 2006, 270, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Edwards, P.D.; Dean, E.K.; Palme, R.; Boonstra, R. Assessing space use in meadow voles: The relationship to reproduction and the stress axis. J. Mammal. 2019, 100, 4–12. [Google Scholar] [CrossRef]
- Touma, C.; Palme, R.; Sachser, N. Analyzing corticosterone metabolites in fecal samples of mice: A noninvasive technique to monitor stress hormones. Horm. Behav. 2004, 45, 10–22. [Google Scholar] [CrossRef]
- Harris, B.N.; Saltzman, W.; de Jong, T.R.; Milnes, M.R. Hypothalamic–pituitary–adrenal (HPA) axis function in the California mouse (Peromyscus californicus): Changes in baseline activity, reactivity, and fecal excretion of glucocorticoids across the diurnal cycle. Gen. Comp. Endocrinol. 2012, 179, 436–450. [Google Scholar] [CrossRef] [Green Version]
- Hare, J.F.; Ryan, C.P.; Enright, C.; Gardiner, L.E.; Skyner, L.J.; Berkvens, C.N.; Anderson, W.G. Validation of a radioimmunoassay-based fecal corticosteroid assay for Richardson’s ground squirrels Urocitellus richardsonii and behavioural correlates of stress. Curr. Zool. 2014, 60, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Fairbairn, D.J. The spring decline in deer mice: Death or dispersal? Can. J. Zool. 1977, 55, 84–92. [Google Scholar] [CrossRef]
- Mills, J.N.; Yates, T.L.; Childs, J.E.; Parmenter, R.R.; Ksiazek, T.G.; Rollin, P.E.; Peters, C. Guidelines for working with rodents potentially infected with hantavirus. J. Mammal. 1995, 76, 716–722. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, B. Fitting Linear Mixed-Effects Models using lme4. J. Stat. Soft. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Soft. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R. Emmeans: Estimated Marginal Means aka Least-Squares Means. 2018. R Package Version 1.3.1. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 1 May 2020).
- Siswanto, H.; Hau, J.; Carlsson, H.E.; Goldkuhl, R.; Abelson, K.S. Corticosterone concentrations in blood and excretion in faeces after ACTH administration in male Sprague-Dawley rats. In Vivo 2008, 22, 435–440. [Google Scholar] [PubMed]
- Bosson, C.O.; Palme, R.; Boonstra, R. Assessment of the stress response in Columbian ground squirrels: Laboratory and field validation of an enzyme immunoassay for fecal cortisol metabolites. Physiol. Biochem. Zool. 2009, 82, 291–301. [Google Scholar] [CrossRef]
- Sheriff, M.J.; Bosson, C.O.; Krebs, C.J.; Boonstra, R. A non-invasive technique for analyzing fecal cortisol metabolites in snowshoe hares (Lepus americanus). J. Comp. Physiol. B 2009, 179, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, B.; McAdam, A.G.; Palme, R.; Fletcher, Q.E.; Boutin, S.; Humphries, M.M.; Boonstra, R. Fecal cortisol metabolite levels in free-ranging North American red squirrels: Assay validation and the effects of reproductive condition. Gen. Comp. Endocrinol. 2010, 167, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Bauer, B.; Palme, R.; Machatschke, I.H.; Dittami, J.; Huber, S. Non-invasive measurement of adrenocortical and gonadal activity in male and female guinea pigs (Cavia aperea f. porcellus). Gen. Comp. Endocrinol. 2008, 156, 482–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, J.M.; Austad, S.N. Fecal corticosteroid levels in free-living populations of deer mice (Peromyscus maniculatus) and southern red-backed voles (Clethrionomys gapperi). Am. Midl. Nat. 2004, 152, 400–409. [Google Scholar] [CrossRef]
- Lepschy, M.; Touma, C.; Hruby, R.; Palme, R. Non-invasive measurement of adrenocortical activity in male and female rats. Lab. Anim. 2007, 41, 372–387. [Google Scholar] [CrossRef] [Green Version]
- Nováková, M.; Palme, R.; Kutalová, H.; Janský, L.; Frynta, D. The effects of sex, age and commensal way of life on levels of fecal glucocorticoid metabolites in spiny mice (Acomys cahirinus). Physiol. Behav. 2008, 95, 187–193. [Google Scholar] [CrossRef]
- Sipari, S.; Ylönen, H.; Palme, R. Excretion and measurement of corticosterone and testosterone metabolites in bank voles (Myodes glareolus). Gen. Comp. Endocrinol. 2017, 243, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Fauteux, D.; Gauthier, G.; Berteaux, D.; Bosson, C.; Palme, R.; Boonstra, R. Assessing stress in Arctic lemmings: Fecal metabolite levels reflect plasma free corticosterone levels. Physiol. Biochem. Zool. 2017, 90, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Millspaugh, J.J.; Washburn, B.E.; Milanick, M.A.; Slotow, R.; van Dyk, G. Effects of heat and chemical treatments on fecal glucocorticoid measurements: Implications for sample transport. Wildl. Soc. Bull. 2003, 31, 399–406. [Google Scholar]
- Goymann, W. On the use of non-invasive hormone research in uncontrolled, natural environments: The problem with sex, diet, metabolic rate and the individual. Methods Ecol. Evol. 2012, 3, 757–765. [Google Scholar] [CrossRef]
- Lexen, E.; El-Bahr, S.; Sommerfeld-Stur, I.; Palme, R.; Möstl, E. Monitoring the adrenocortical response to disturbances in sheep by measuring glucocorticoid metabolites in the faeces. Wien. Tierarztl. Monat. 2008, 95, 64. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Treatment 1 | Immunoassay | Sample Collection 2 Schedule (h Post Treatment) |
|---|---|---|---|
| May 2–3 | Acclimation | Corticosterone EIA | 70, 72, 74, 76, 80, 82 |
| Group-specific EIA | 70, 72, 74, 76, 78, 80, 82 | ||
| May 3–4 | Adrenal suppression | Corticosterone EIA | 2, 4, 6, 10, 12 |
| Group-specific EIA | 2, 4, 6, 10, 12 | ||
| May 5–6 | Adrenal stimulation | Corticosterone EIA | 2, 4, 6, 10, 12 |
| Group-specific EIA | 2, 4, 6, 10, 12 |
| Sex | Reproductive 1 | FCMs at 0–4 h (ng/g) | FCMs Overnight (ng/g) | FCM Difference 2 |
|---|---|---|---|---|
| Male | No | 21,034 | 49,709 | 28,676 |
| Male | No | 5407 | 76,071 | 70,663 |
| Male | Yes | 11,224 | 185,928 | 174,704 |
| Male | Yes | 4649 | 91,514 | 86,865 |
| Female | Yes | 37,172 | 37,776 | 604 |
| Female | Yes | 3611 | 41,678 | 38,067 |
| Female | Yes | 9704 | 82,299 | 72,595 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eleftheriou, A.; Palme, R.; Boonstra, R. Assessment of the Stress Response in North American Deermice: Laboratory and Field Validation of Two Enzyme Immunoassays for Fecal Corticosterone Metabolites. Animals 2020, 10, 1120. https://doi.org/10.3390/ani10071120
Eleftheriou A, Palme R, Boonstra R. Assessment of the Stress Response in North American Deermice: Laboratory and Field Validation of Two Enzyme Immunoassays for Fecal Corticosterone Metabolites. Animals. 2020; 10(7):1120. https://doi.org/10.3390/ani10071120
Chicago/Turabian StyleEleftheriou, Andreas, Rupert Palme, and Rudy Boonstra. 2020. "Assessment of the Stress Response in North American Deermice: Laboratory and Field Validation of Two Enzyme Immunoassays for Fecal Corticosterone Metabolites" Animals 10, no. 7: 1120. https://doi.org/10.3390/ani10071120