Chloroquine Downregulation of Intestinal Autophagy to Alleviate Biological Stress in Early-Weaned Piglets

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals Experimental Design, and Diets
2.2. Sample Collection and Preparation
2.3. Western Blotting Analysis
2.4. Transmission Electron Microscope
2.5. Serum Inflammatory Cytokines, Immune Factors, Diamine Oxidase, and D-lactate
2.6. Plasma Antioxidative Capacity
2.7. Intestinal Morphology
2.8. Real-Time Quantitative Reverse Transcriptase PCR
2.9. Statistical Analysis
3. Results
3.1. Growth Performance
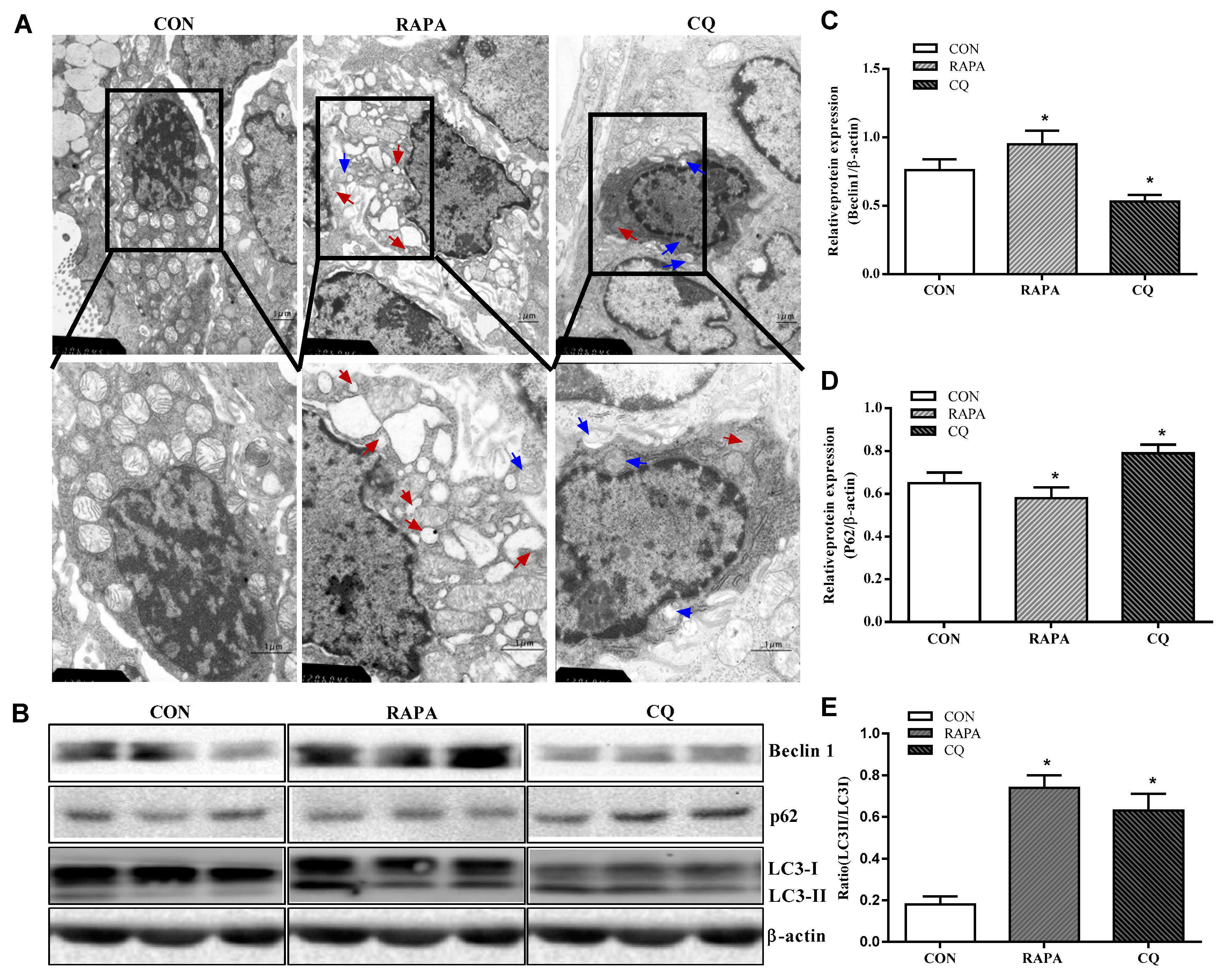
3.2. Autophagy Analysis
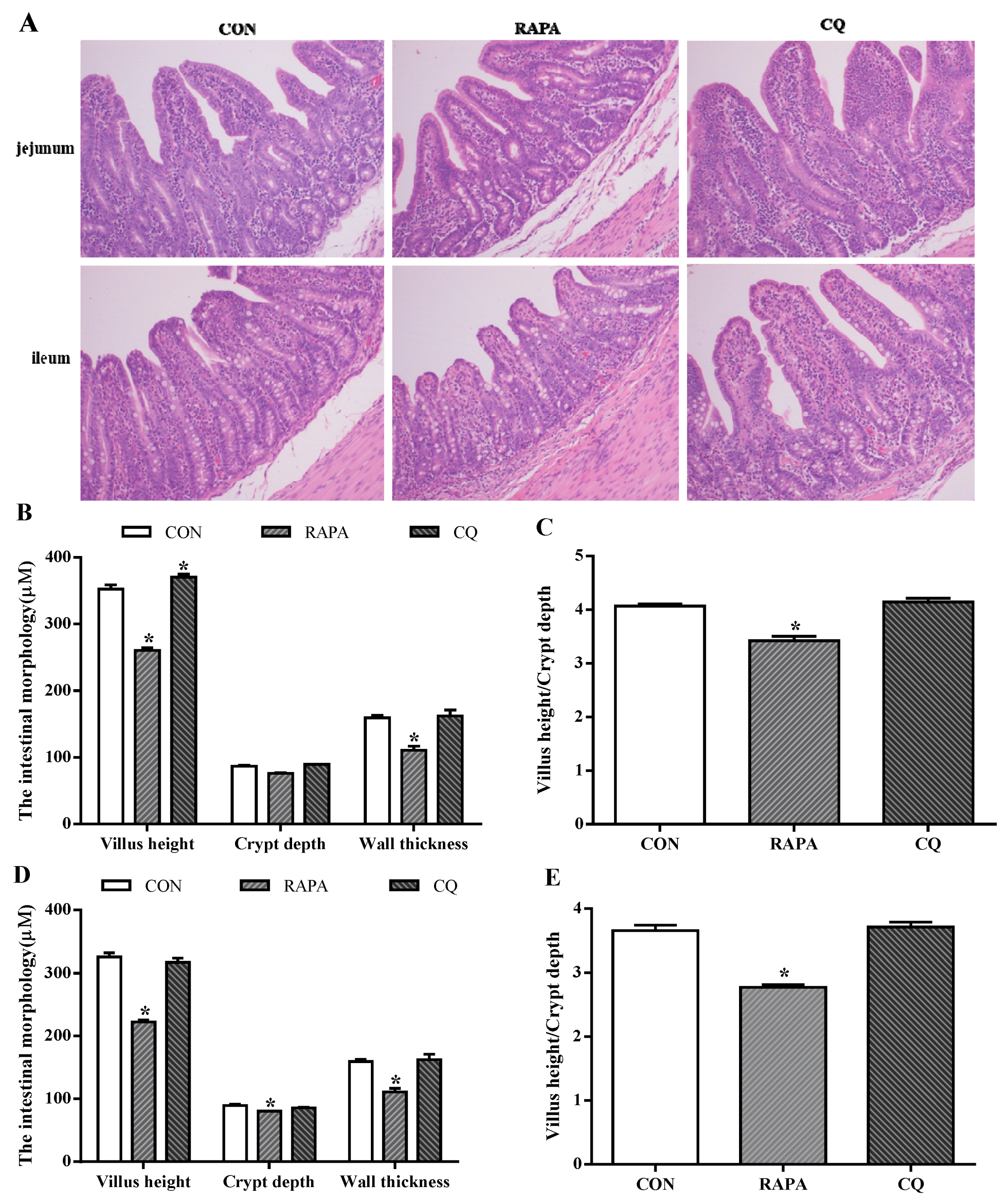
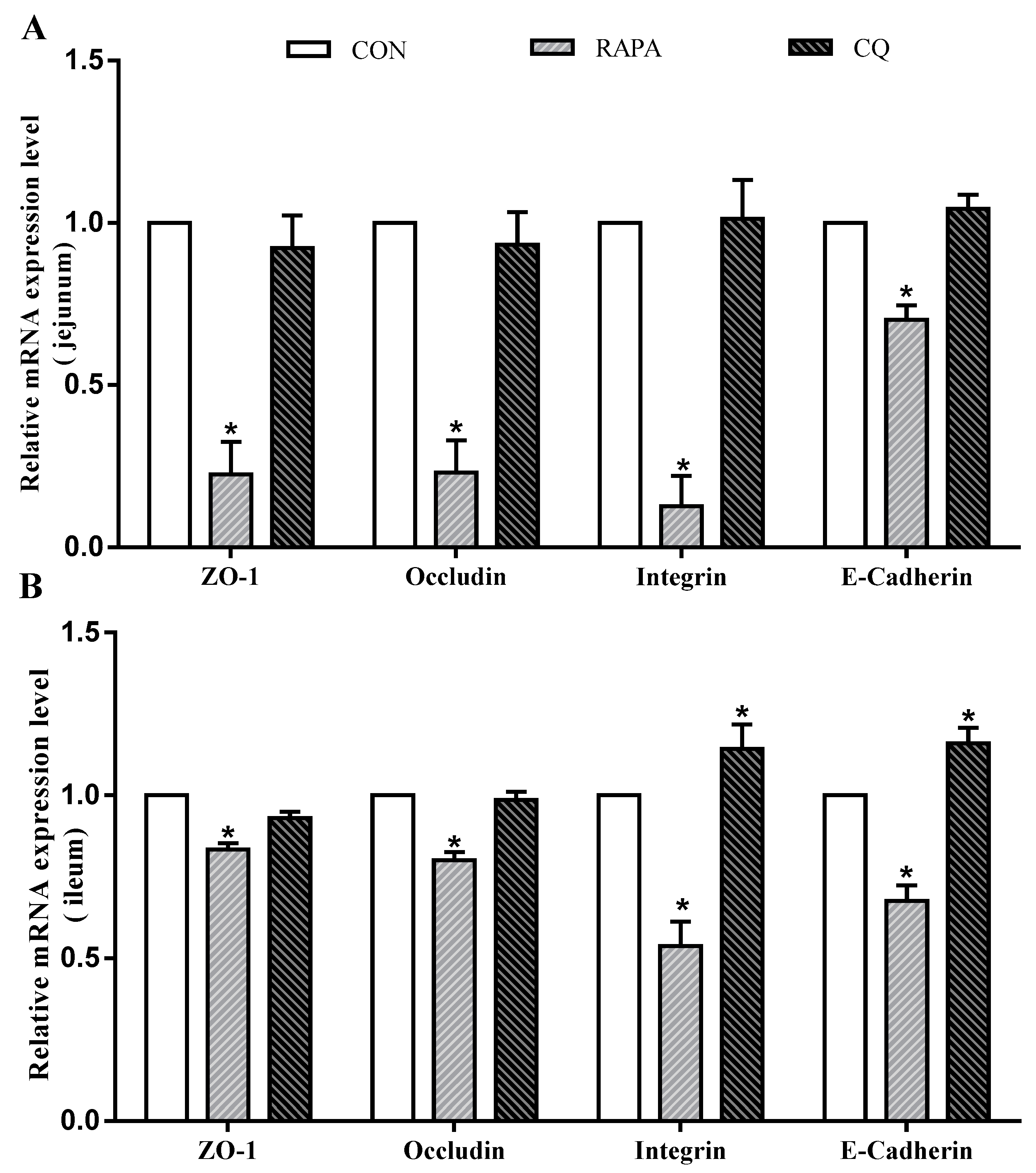
3.3. Intestinal Morphology and Permeability
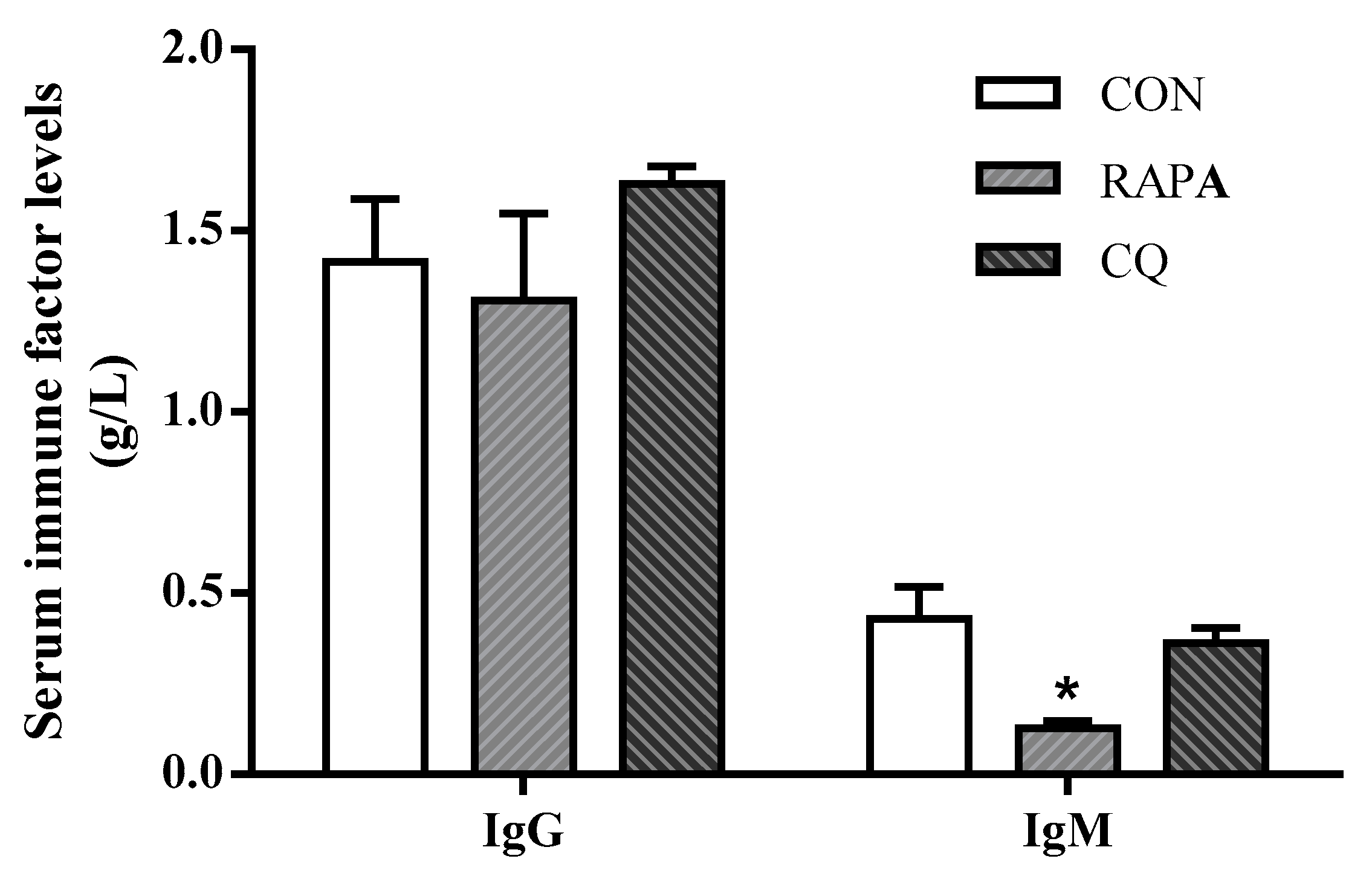
3.4. Inflammatory Cytokine, Immune, and Antioxidant Factor Profiles
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Kick, A.R.; Tompkins, M.B.; Flowers, W.L.; Whisnant, C.S.; Almond, G.W. Effects of Stress Associated with Weaning on the Adaptive Immune System in Pigs. J. Anim. Sci. 2012, 90, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early Weaning Increases Intestinal Permeability, Alters Expression of Cytokine and Tight Junction Proteins, and Activates Mitogen-Activated Protein Kinases in Pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moeser, A.J.; Pohl, C.S.; Rajput, M. Weaning Stress and Gastrointestinal Barrier Development: Implications for Lifelong Gut Health in Pigs. Anim. Nutr. 2017, 3, 313–321. [Google Scholar] [CrossRef]
- Xiao, H.; Tan, B.E.; Wu, M.M.; Yin, Y.L.; Li, T.J.; Yuan, D.X.; Li, L. Effects of Composite Antimicrobial Peptides in Weanling Piglets Challenged with Deoxynivalenol: Ii. Intestinal Morphology and Function. J. Anim. Sci. 2013, 91, 4750–4756. [Google Scholar] [CrossRef]
- Cao, S.T.; Wang, C.C.; Wu, H.; Zhang, Q.H.; Jiao, L.F.; Hu, C.H. Weaning Disrupts Intestinal Antioxidant Status, Impairs Intestinal Barrier and Mitochondrial Function, and Triggers Mitophagy in Piglets. J. Anim. Sci. 2018, 96, 1073–1083. [Google Scholar] [CrossRef]
- Xiao, H.; Wu, M.M.; Tan, B.E.; Yin, Y.L.; Li, T.J.; Xiao, D.F.; Li, L. Effects of Composite Antimicrobial Peptides in Weanling Piglets Challenged with Deoxynivalenol: I. Growth Performance, Immune Function, and Antioxidation Capacity. J. Anim. Sci. 2013, 91, 4772–4780. [Google Scholar] [CrossRef]
- Galluzzi, L.; Pietrocola, F.; Levine, B.; Kroemer, G. Metabolic Control of Autophagy. Cell 2014, 159, 1263–1276. [Google Scholar] [CrossRef] [Green Version]
- Grizotte-Lake, M.; Vaishnava, S. Autophagy: Suicide Prevention Hotline for the Gut Epithelium. Cell Host Microbe 2018, 23, 147–148. [Google Scholar] [CrossRef] [Green Version]
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and Molecular Mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Huett, A.; Xavier, R.J. Autophagy at the Gut Interface: Mucosal Responses to Stress and the Consequences for Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2010, 16, 152–174. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Li, J.; Li, F.; Hu, C.A.; Liao, P.; Tan, K.; Tan, B.; Xiong, X.; Liu, G.; Li, T.; et al. Autophagy Protects Intestinal Epithelial Cells against Deoxynivalenol Toxicity by Alleviating Oxidative Stress Via Ikk Signaling Pathway. Free Radic. Biol. Med. 2015, 89, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Randall-Demllo, S.; Chieppa, M.; Eri, R. Intestinal Epithelium and Autophagy: Partners in Gut Homeostasis. Front. Immunol. 2013, 4, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshaer, D.; Begun, J. The Role of Barrier Function, Autophagy, and Cytokines in Maintaining Intestinal Homeostasis. Semin. Cell Dev. Biol. 2017, 61, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, K.K.; Stappenbeck, T.S. Autophagy and Intestinal Homeostasis. Annu. Rev. Physiol. 2013, 75, 241–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Klionsky, D.J. Regulation Mechanisms and Signaling Pathways of Autophagy. Annu. Rev. Genet. 2009, 43, 67–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalbandian, A.; Llewellyn, K.J.; Nguyen, C.; Yazdi, P.G.; Kimonis, V.E. Rapamycin and Chloroquine: The in Vitro and in Vivo Effects of Autophagy-Modifying Drugs Show Promising Results in Valosin Containing Protein Multisystem Proteinopathy. PLoS ONE 2015, 10, e0122888. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.Q.; Zhao, J.P.; Fan, X.X.; Liu, G.H.; Jiao, H.C.; Wang, X.J.; Sun, S.H.; Lin, H. Rapamycin, a Specific Inhibitor of the Target of Rapamycin Complex 1, Disrupts Intestinal Barrier Integrity in Broiler Chicks. J. Anim. Physiol. Anim. Nutr. 2016, 100, 323–330. [Google Scholar] [CrossRef]
- Makky, K.; Tekiela, J.; Mayer, A.N. Target of Rapamycin (Tor) Signaling Controls Epithelial Morphogenesis in the Vertebrate Intestine. Dev. Biol. 2007, 303, 501–513. [Google Scholar] [CrossRef] [Green Version]
- Nagar, J.; Ranade, S.; Kamath, V.; Singh, S.; Karunanithi, P.; Subramani, S.; Venkatesh, K.; Srivastava, R.; Dudhgaonkar, S.; Vikramadithyan, R.K. Therapeutic Potential of Chloroquine in a Murine Model of Inflammatory Bowel Disease. Int. Immunopharmacol. 2014, 21, 328–335. [Google Scholar] [CrossRef]
- Li, P.; Hao, L.; Guo, Y.Y.; Yang, G.L.; Mei, H.; Li, X.H.; Zhai, Q.X. Chloroquine Inhibits Autophagy and Deteriorates the Mitochondrial Dysfunction and Apoptosis in Hypoxic Rat Neurons. Life Sci. 2018, 202, 70–77. [Google Scholar] [CrossRef]
- Wu, F.; Wei, X.; Wu, Y.; Kong, X.; Hu, A.; Tong, S.; Liu, Y.; Gong, F.; Xie, L.; Zhang, J.; et al. Chloroquine Promotes the Recovery of Acute Spinal Cord Injury by Inhibiting Autophagy-Associated Inflammation and Endoplasmic Reticulum Stress. J. Neurotrauma. 2018, 35, 1329–1344. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tan, B.; Huang, B.; Li, J.; Wang, J.; Liao, P.; Guan, G.; Ji, P.; Yin, Y. Involvement of CaSR activation in the alleviation of intestinal inflammation in piglets model by dietary aromatic amino acid supplementation. Br. J. Nutr. 2018, 120, 1321–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Yao, K.; Wang, L.; Ding, B.; Fu, D.; Liu, Y.; Zhu, H.; Liu, J.; Li, Y.; Kang, P.; et al. Effects of Alpha-Ketoglutarate on Energy Status in the Intestinal Mucosa of Weaned Piglets Chronically Challenged with Lipopolysaccharide. Br. J. Nutr. 2011, 106, 357–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Li, F.; Liu, H.; Li, Y.; Liu, Y.; Kong, X.; Zhang, Y.; Deng, D.; Tang, Y.; Feng, Z.; et al. Nutritional and Regulatory Roles of Leucine in Muscle Growth and Fat Reduction. Front. Biosci. 2015, 20, 796–813. [Google Scholar]
- Chen, J.; Su, W.; Kang, B.; Jiang, Q.; Zhao, Y.; Fu, C.; Yao, K. Supplementation with Alpha-Ketoglutarate to a Low-Protein Diet Enhances Amino Acid Synthesis in Tissues and Improves Protein Metabolism in the Skeletal Muscle of Growing Pigs. Amino Acids 2018, 50, 1525–1537. [Google Scholar] [CrossRef]
- Wang, J.; Li, G.R.; Tan, B.E.; Xiong, X.; Kong, X.F.; Xiao, D.F.; Xu, L.W.; Wu, M.M.; Huang, B.; Kim, S.W.; et al. Oral Administration of Putrescine and Proline During the Suckling Period Improves Epithelial Restitution after Early Weaning in Piglets. J. Anim. Sci. 2015, 93, 1679–1688. [Google Scholar] [CrossRef]
- Coleman, C.M.; Olivier, A.K.; Jacobus, J.A.; Mapuskar, K.A.; Mao, G.; Martin, S.M.; Riley, D.P.; Gius, D.; Spitz, D.R. Superoxide Mediates Acute Liver Injury in Irradiated Mice Lacking Sirtuin 3. Antioxid. Redox Signal. 2014, 20, 1423–1435. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.Q.; Varma, M.V.; Wolford, A.; Ryder, T.; Di, L.; Feng, B.; Terra, S.G.; Sagawa, K.; Kalgutkar, A.S. Pharmacokinetics and Disposition of the Thiouracil Derivative Pf-06282999, an Orally Bioavailable, Irreversible Inactivator of Myeloperoxidase Enzyme, across Animals and Humans. Drug Metab. Dispos. 2016, 44, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.R. Intestinal Mucosal Barrier Function in Health and Disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Yu, Y.; Shiou, S.R.; Guo, Y.; Lu, L.; Westerhoff, M.; Sun, J.; Petrof, E.O.; Claud, E.C. Erythropoietin Protects Epithelial Cells from Excessive Autophagy and Apoptosis in Experimental Neonatal Necrotizing Enterocolitis. PLoS ONE 2013, 8, e69620. [Google Scholar] [CrossRef] [Green Version]
- Baxt, L.A.; Xavier, R.J. Role of Autophagy in the Maintenance of Intestinal Homeostasis. Gastroenterology 2015, 149, 553–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.E.; Duan, X.; Lin, D.; Li, T.; Luo, D.; Wang, L.; Lian, K. Rapamycin-Induced Autophagy Activity Promotes Bone Fracture Healing in Rats. Exp. Ther. Med. 2015, 10, 1327–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, C.; Li, F.; Long, Y.; Zheng, J. Chloroquine Attenuates Lipopolysaccharide-Induced Inflammatory Responses through Upregulation of Usp25. Can. J. Physiol. Pharmacol. 2017, 95, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Wu, N.; Wang, Y.; Zhao, H.; Zhang, L.; Li, T.; Zhao, M. Chloroquine Attenuates Paraquat-Induced Lung Injury in Mice by Altering Inflammation, Oxidative Stress and Fibrosis. Int. Immunopharmacol. 2017, 46, 16–22. [Google Scholar] [CrossRef]
- Siracusa, R.; Paterniti, I.; Bruschetta, G.; Cordaro, M.; Impellizzeri, D.; Crupi, R.; Cuzzocre, S.; Esposito, E. The Association of Palmitoylethanolamide with Luteolin Decreases Autophagy in Spinal Cord Injury. Mol. Neurobiol. 2016, 53, 3783–3792. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession No. | Primers | Sequences (5′-3′) |
|---|---|---|---|
| E-cadherin | NM_001163060.1 | Forward | GAAGGAGGTGGAGAAGAGGAC |
| Reverse | AGAGTCATAAGGTGGGGCAGT | ||
| Occludin | NM_001163647.2 | Forward | AGAGTCATAAGGTGGGGCAGT |
| Reverse | CGCCCGTCGTGTAGTCTGTC | ||
| ZO-1 | XM_005659811.1 | Forward | TACCCTGCGGCTGGAAGA |
| Reverse | GGACGGGACCTGCTCATAACT | ||
| Integrin | NM_213968.1 | Forward | GCAGTTTCAAGGTCAAGATGG |
| Reverse | AGCAGGAGGAAGATGAGCAG | ||
| β-actin | XM_003124280.3 | Forward | GGATGCAGAAGGAGATCACG |
| Reverse | ATCTGCTGGAAGGTGGACAG |
| Items | Dietary Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | RAPA | CQ | |||
| Day 1 body weight, kg | 7.14 | 6.78 | 6.86 | 0.15 | 0.63 |
| Day 7 body weight, kg | 8.04 | 6.78 * | 7.84 | 0.20 | <0.01 |
| Day 14 body weight, kg | 9.57 | 6.80 * | 9.99 | 0.40 | <0.01 |
| Average daily gain, g | 173.43 | 1.19 * | 223.29 * | 26.13 | <0.01 |
| Average daily feed intake, g | 261.67 | 122.50 | 254.50 | 16.53 | <0.01 |
| Feed efficiency, g gain/g feed | 0.60 | 0.13 | 0.91 | 0.11 | <0.01 |
| Items | Dietary Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | RAPA | CQ | |||
| DAO, mmol/L | 1.40 | 0.72 * | 1.35 | 0.22 | < 0.01 |
| D-lactate, μg/mL | 77.75 | 95.67 * | 63.53 * | 9.30 | < 0.01 |
| Items | Dietary Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | RAPA | CQ | |||
| IL-6 | 669.74 | 705.83 * | 565.70 * | 42.01 | 0.04 |
| IL-12 | 106.20 | 123.07 * | 104.06 | 24.60 | <0.01 |
| IL-1β | 339.61 | 376.88 * | 231.75 * | 44.53 | <0.01 |
| TNF-α | 110.68 | 125.06 | 96.10 | 8.53 | <0.01 |
| IL-8 | 38.48 | 44.40 * | 23.56 * | 6.20 | <0.01 |
| IFN-γ | 25.59 | 27.19 | 13.34 * | 4.37 | <0.01 |
| TGF-β | 454.20 | 216.89 * | 549.37 * | 47.00 | <0.01 |
| IL-10 | 107.62 | 91.83 | 106.80 | 5.13 | 0.20 |
| Items | Dietary treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | RAPA | CQ | |||
| SOD, U/ml | 74.38 | 64.28 * | 71.86 | 1.17 | < 0.01 |
| MDA, nmol/ml | 3.65 | 4.56 * | 3.43 | 0.16 | < 0.01 |
| GST, U/ml | 34.32 | 26.61 * | 42.60 * | 1.72 | < 0.01 |
| GSH-PX, U/ml | 105.39 | 86.33 * | 114.52 * | 3.07 | < 0.01 |
| T-AOC, mmol/L | 0.22 | 0.18 | 0.22 | 0.01 | 0.02 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, S.; Tang, S.; Chang, M.; Qi, M.; Li, J.; Tan, B.; Gao, Q.; Zhang, S.; Li, X.; Yin, Y.; et al. Chloroquine Downregulation of Intestinal Autophagy to Alleviate Biological Stress in Early-Weaned Piglets. Animals 2020, 10, 290. https://doi.org/10.3390/ani10020290
Liao S, Tang S, Chang M, Qi M, Li J, Tan B, Gao Q, Zhang S, Li X, Yin Y, et al. Chloroquine Downregulation of Intestinal Autophagy to Alleviate Biological Stress in Early-Weaned Piglets. Animals. 2020; 10(2):290. https://doi.org/10.3390/ani10020290
Chicago/Turabian StyleLiao, Simeng, Shengguo Tang, Meinan Chang, Ming Qi, Jianjun Li, Bie Tan, Qian Gao, Shuo Zhang, Xiaozhen Li, Yulong Yin, and et al. 2020. "Chloroquine Downregulation of Intestinal Autophagy to Alleviate Biological Stress in Early-Weaned Piglets" Animals 10, no. 2: 290. https://doi.org/10.3390/ani10020290