Enhancing Essential Grains Yield for Sustainable Food Security and Bio-Safe Agriculture through Latest Innovative Approaches

 , , ,
, , ,  and
and 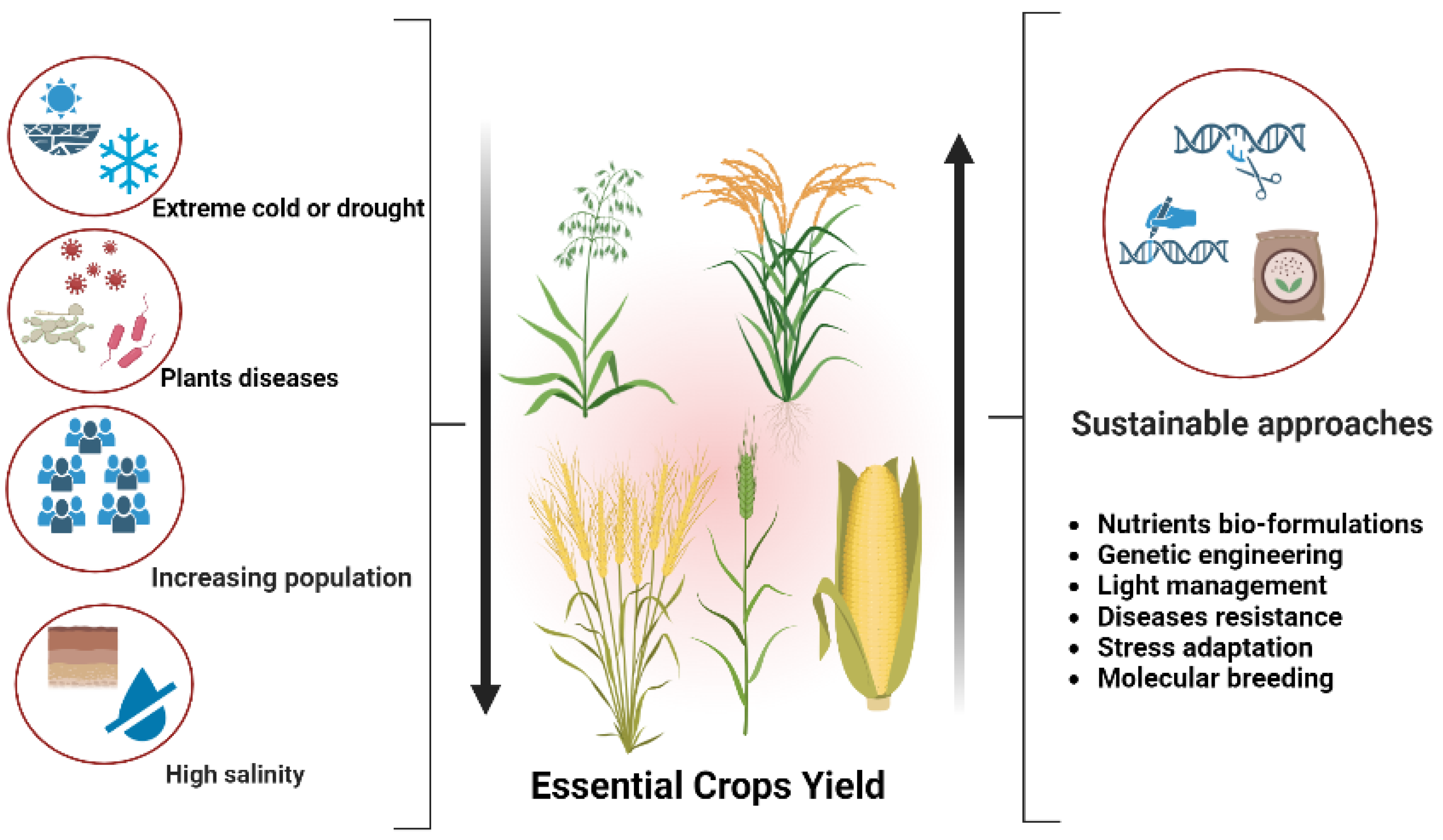{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methodology of the Review
3. Wheat
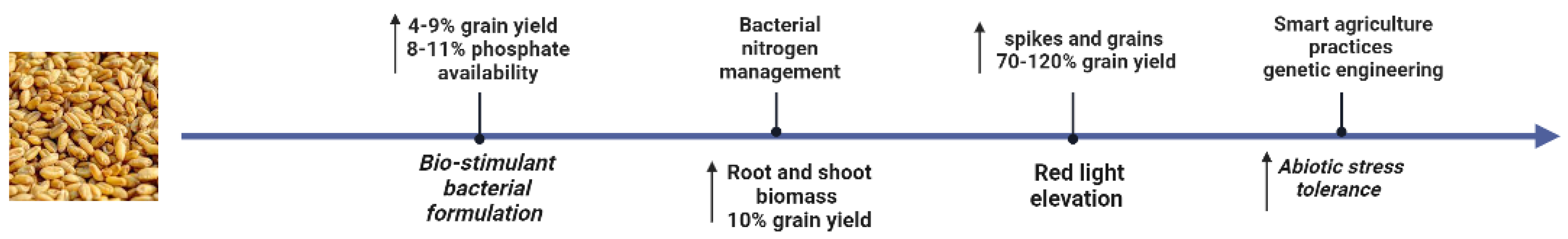
3.1. Synergistic Bio-Stimulant of Wheat Production by Autochthonous Phosphate_Solubilizing Bacteria Formulation
3.2. Improving Sustainable Field-Grown Wheat Production by Azospirillumbrasilense Nitrogen Management
3.3. Maximizing Wheat Yield by Increasing Exposure to Red to Far-Red Light during Specific Stages of Wheat Development
3.4. Negotiating Smart Agricultural Practices to Increase Wheat Yield
3.5. Improving Wheat Adaptation to Abiotic Stresses through Biotechnical and Genetic Approaches
3.6. Utilizing Genetic Rust Resistance in Wheat and Integrated Rust Management Techniques to Create More Resilient Cultivars
4. Rice
4.1. Varying Fertilizer Combinations and Planting Techniques Result in Different Rice Yields
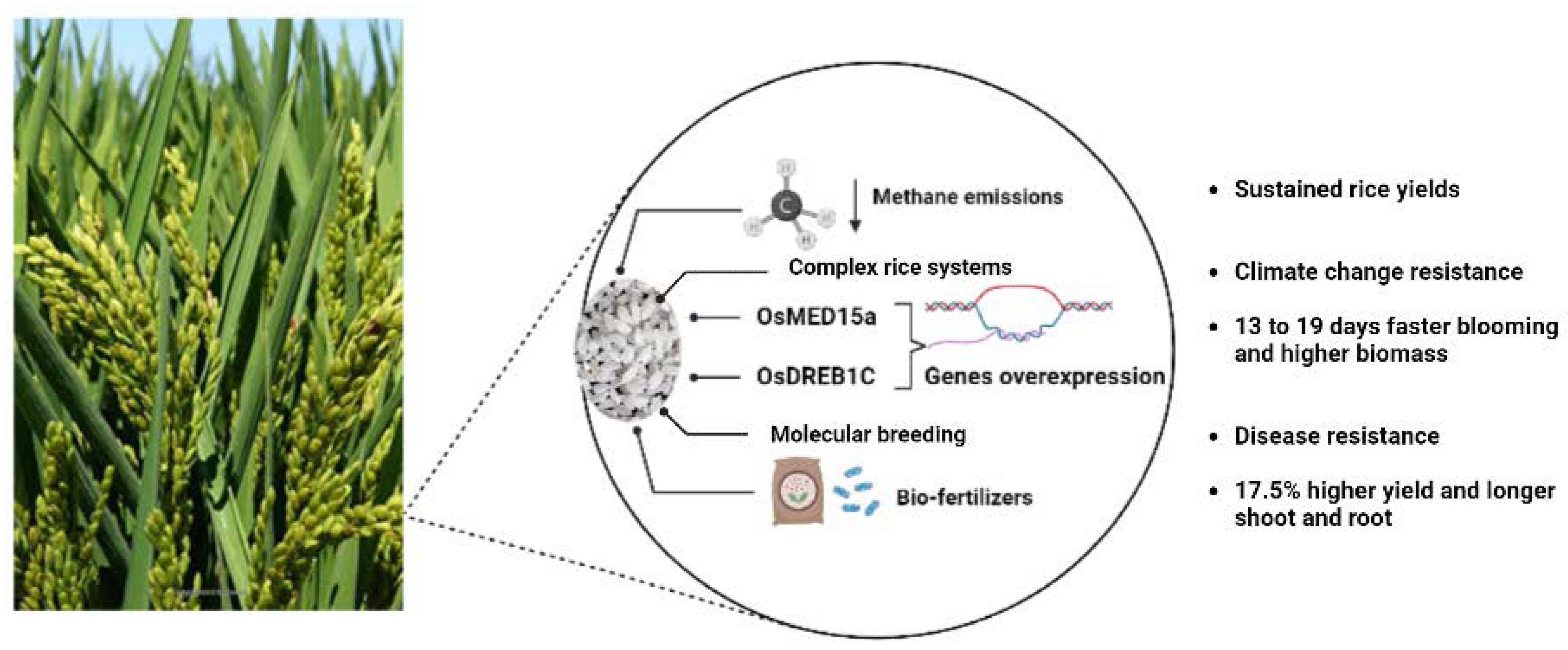
4.2. Maintaining Sustainable Rice Yield through Methane Emissions Reduction from Rice Cropping Systems
4.3. Increasing Rice Yield and Yield Stability Despite Changing Weather Conditions by Complex Rice Systems
4.4. Improving Rice Crop Yield through Integrated Molecular Genetics Approach
4.5. Enhancing Rice Crop Productivity by Sustainable Perennial Rice Approach
4.6. Increasing Rice Blast Disease Resistance through a Molecular Breeding Approach
5. Corn
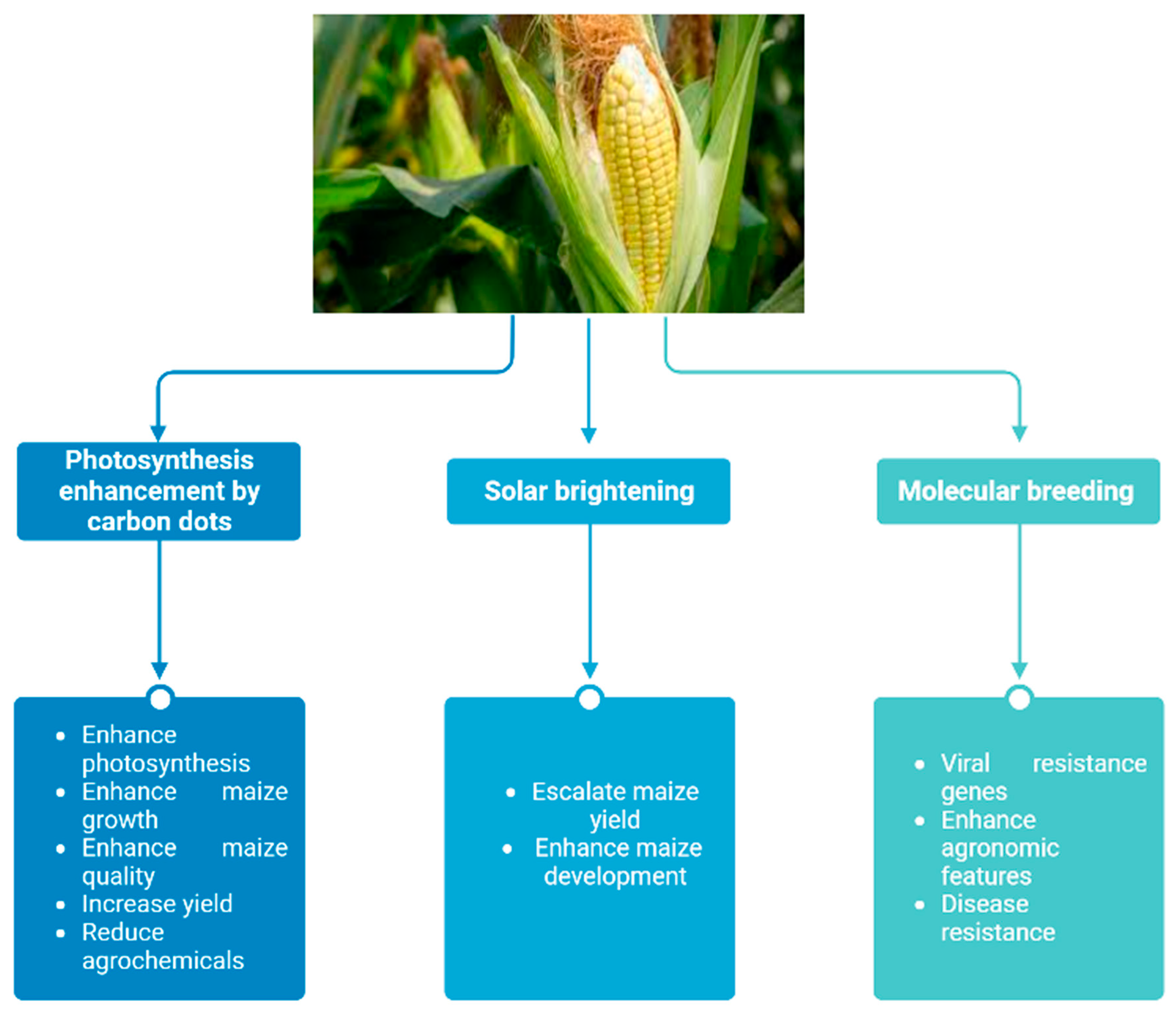
5.1. Enhancing Corn Photosystem’s Light Conversion Efficiency and Electron Supply Using Nitrogen-Doped Carbon Dots
5.2. Improving Maize Yield by Solar Brightening
5.3. Increasing Maize Disease Resistance through Molecular Breeding Genetic Analysis
6. Barley
7. Oats
8. Grain Varietal Improvement
9. Potential Challenges and Future Perspectives
10. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saeed, F.; Chaudhry, U.K.; Raza, A.; Charagh, S.; Bakhsh, A.; Bohra, A.; Ali, S.; Chitikineni, A.; Saeed, Y.; Visser, R.G.F.; et al. Developing Future Heat-Resilient Vegetable Crops. Funct. Integr. Genom. 2023, 23, 47. [Google Scholar] [CrossRef]
- Thudi, M.; Palakurthi, R.; Schnable, J.C.; Chitikineni, A.; Dreisigacker, S.; Mace, E.; Srivastava, R.K.; Satyavathi, C.T.; Odeny, D.; Tiwari, V.K.; et al. Genomic Resources in Plant Breeding for Sustainable Agriculture. J. Plant Physiol. 2021, 257, 153351. [Google Scholar] [CrossRef]
- United Nations Office for the Coordination of Humanitarian Affairs. Global Humanitarian Overview 2022; United Nations: New York, NY, USA, 2021; ISBN 978-92-1-001242-3. [Google Scholar]
- Parry, M.A.J.; Hawkesford, M.J. Food Security: Increasing Yield and Improving Resource Use Efficiency. Proc. Nutr. Soc. 2010, 69, 592–600. [Google Scholar] [CrossRef] [Green Version]
- Igrejas, G.; Branlard, G. The Importance of Wheat. In Wheat Quality for Improving Processing and Human Health; Igrejas, G., Ikeda, T.M., Guzmán, C., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–7. ISBN 978-3-030-34162-6. [Google Scholar]
- Shewry, P.R.; Hey, S.J. The Contribution of Wheat to Human Diet and Health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Kim, Y.; Chung, Y.S.; Lee, E.; Tripathi, P.; Heo, S.; Kim, K.-H. Root Response to Drought Stress in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2020, 21, 1513. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, D.B.; Brotman, Y.; Fernie, A.R. The Utility of Metabolomics as a Tool to Inform Maize Biology. Plant Commun. 2021, 2, 100187. [Google Scholar] [CrossRef]
- Salvador-Reyes, R.; Clerici, M.T.P.S. Peruvian Andean Maize: General Characteristics, Nutritional Properties, Bioactive Compounds, and Culinary Uses. Food Res. Int. 2020, 130, 108934. [Google Scholar] [CrossRef]
- Nuss, E.T.; Tanumihardjo, S.A. Maize: A Paramount Staple Crop in the Context of Global Nutrition. Compr. Rev. Food Sci. Food Saf. 2010, 9, 417–436. [Google Scholar] [CrossRef]
- Sullivan, P.; Arendt, E.; Gallagher, E. The Increasing Use of Barley and Barley By-Products in the Production of Healthier Baked Goods. Trends Food Sci. Technol. 2013, 29, 124–134. [Google Scholar] [CrossRef]
- Badr, A.; Müller, K.; Schäfer-Pregl, R.; Rabey, H.E.; Effgen, S.; Ibrahim, H.H.; Pozzi, C.; Rohde, W.; Salamini, F. On the Origin and Domestication History of Barley (Hordeum vulgare). Mol. Biol. Evol. 2000, 17, 499–510. [Google Scholar] [CrossRef] [Green Version]
- Giraldo, P.; Benavente, E.; Manzano-Agugliaro, F.; Gimenez, E. Worldwide Research Trends on Wheat and Barley: A Bibliometric Comparative Analysis. Agronomy 2019, 9, 352. [Google Scholar] [CrossRef] [Green Version]
- Paudel, D.; Dhungana, B.; Caffe, M.; Krishnan, P. A Review of Health-Beneficial Properties of Oats. Foods 2021, 10, 2591. [Google Scholar] [CrossRef]
- United Nations Department of Economic and Social Affairs Population Division. World Population Prospects 2019: Highlights; ST/ESA/SER.A/423; United Nations: New York, NY, USA, 2019. [Google Scholar]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield Trends Are Insufficient to Double Global Crop Production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Razzaq, A.; Mehmood, S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, J.R.; Kossmann, J. Improving Crops for a Changing World. Front. Plant Sci. 2021, 12, 728328. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, Z.; Cheng, M.; Chen, J.; Zhu, T.; Wang, R.; Liu, Y.; Qi, P.; Chen, G.; Jiang, Q.; et al. Uncovering the Dispersion History, Adaptive Evolution and Selection of Wheat in China. Plant Biotechnol. J. 2018, 16, 280–291. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.; Islam, S.; Yu, Z.; She, M.; Nevo, E.; Ma, W. Current Progress in Understanding and Recovering the Wheat Genes Lost in Evolution and Domestication. Int. J. Mol. Sci. 2020, 21, 5836. [Google Scholar] [CrossRef]
- Lu, M.; Cao, X.; Pan, J.; Gurajala, H.K.; He, Z.; Yang, X.; Khan, M.B. Genotypic Variations in Zinc Accumulation and Bioaccessibility among Wheat (Triticum aestivum L.) Genotypes under Two Different Field Conditions. J. Cereal Sci. 2020, 93, 102953. [Google Scholar] [CrossRef]
- International Wheat Genome Sequencing Consortium (IWGSC). Shifting the Limits in Wheat Research and Breeding Using a Fully Annotated Reference Genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [Green Version]
- Langridge, P. Wheat Genomics and the Ambitious Targets for Future Wheat Production. Genome 2013, 56, 545–547. [Google Scholar] [CrossRef]
- Genetic Improvement of Wheat Early Vigor Promote Weed-Competitiveness under Mediterranean Climate. Plant Sci. 2021, 303, 110785. [CrossRef]
- Würschum, T.; Langer, S.M.; Longin, C.F.H.; Tucker, M.R.; Leiser, W.L. A Modern Green Revolution Gene for Reduced Height in Wheat. Plant J. 2017, 92, 892–903. [Google Scholar] [CrossRef] [Green Version]
- Singh Bakala, H.; Singh Mandahal, K.; Ankita; Kaur Sarao, L.; Srivastava, P. Breeding Wheat for Biotic Stress Resistance: Achievements, Challenges and Prospects. In Current Trends in Wheat Research; Ansari, M.-R., Ed.; IntechOpen: London, UK, 2022; ISBN 978-1-83968-593-4. [Google Scholar]
- Hovmøller, M.S.; Sørensen, C.K.; Walter, S.; Justesen, A.F. Diversity of Puccinia Striiformis on Cereals and Grasses. Annu. Rev. Phytopathol. 2011, 49, 197–217. [Google Scholar] [CrossRef]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Bhavani, S.; Njau, P.; Herrera-Foessel, S.; Singh, P.K.; Singh, S.; Govindan, V. The Emergence of Ug99 Races of the Stem Rust Fungus Is a Threat to World Wheat Production. Annu. Rev. Phytopathol. 2011, 49, 465–481. [Google Scholar] [CrossRef] [Green Version]
- Chandran, H.; Meena, M.; Swapnil, P. Plant Growth-Promoting Rhizobacteria as a Green Alternative for Sustainable Agriculture. Sustainability 2021, 13, 10986. [Google Scholar] [CrossRef]
- Mubeen, K.; Wasaya, A.; ur Rehman, H.; Yasir, T.A.; Farooq, O.; Imran, M.; Ikram, R.M.; Nazeer, R.; Zahoor, F.; Yonas, M.W.; et al. Integrated Phosphorus Nutrient Sources Improve Wheat Yield and Phosphorus Use Efficiency under Sub Humid Conditions. PLoS ONE 2021, 16, e0255043. [Google Scholar] [CrossRef]
- Yahya, M.; Rasul, M.; Sarwar, Y.; Suleman, M.; Tariq, M.; Hussain, S.Z.; Sajid, Z.I.; Imran, A.; Amin, I.; Reitz, T.; et al. Designing Synergistic Biostimulants Formulation Containing Autochthonous Phosphate-Solubilizing Bacteria for Sustainable Wheat Production. Front. Microbiol. 2022, 13, 889073. [Google Scholar] [CrossRef]
- Mandic, V.; Krnjaja, V.; Tomic, Z.; Bijelic, Z.; Simic, A.; Ruzic Muslic, D.; Gogic, M. Nitrogen Fertilizer Influence on Wheat Yield and Use Efficiency under Different Environmental Conditions. Chil. J. Agric. Res. 2015, 75, 92–97. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Cui, S.; Zhang, Q.; Xu, G.; Feng, Q.; Chen, C.; Li, Y. Optimizing Wheat Yield, Water, and Nitrogen Use Efficiency with Water and Nitrogen Inputs in China: A Synthesis and Life Cycle Assessment. Front. Plant Sci. 2022, 13, 930484. [Google Scholar] [CrossRef]
- Galindo, F.S.; Pagliari, P.H.; Fernandes, G.C.; Rodrigues, W.L.; Boleta, E.H.M.; Jalal, A.; Céu, E.G.O.; de Lima, B.H.; Lavres, J.; Teixeira Filho, M.C.M. Improving Sustainable Field-Grown Wheat Production with Azospirillum Brasilense Under Tropical Conditions: A Potential Tool for Improving Nitrogen Management. Front. Environ. Sci. 2022, 10, 821628. [Google Scholar] [CrossRef]
- Kasajima, S.; Inoue, N.; Mahmud, R.; Fujita, K.; Kato, M. Effect of Light Quality on Developmental Rate of Wheat under Continuous Light at a Constant Temperature. Plant Prod. Sci. 2007, 10, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Dreccer, M.F.; Zwart, A.B.; Schmidt, R.; Condon, A.G.; Awasi, M.A.; Grant, T.J.; Galle, A.; Bourot, S.; Frohberg, C. Wheat Yield Potential Can Be Maximized by Increasing Red to Far-red Light Conditions at Critical Developmental Stages. Plant Cell Environ. 2022, 45, 2652–2670. [Google Scholar] [CrossRef]
- Wheeler, T.; von Braun, J. Climate Change Impacts on Global Food Security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef]
- Tian, S.; Ning, T.; Zhao, H.; Wang, B.; Li, N.; Han, H.; Li, Z.; Chi, S. Response of CH4 and N2O Emissions and Wheat Yields to Tillage Method Changes in the North China Plain. PLoS ONE 2012, 7, e51206. [Google Scholar] [CrossRef] [Green Version]
- Lipper, L.; Thornton, P.; Campbell, B.M.; Baedeker, T.; Braimoh, A.; Bwalya, M.; Caron, P.; Cattaneo, A.; Garrity, D.; Henry, K.; et al. Climate-Smart Agriculture for Food Security. Nat. Clim. Chang. 2014, 4, 1068–1072. [Google Scholar] [CrossRef]
- Ishtiaq, M.; Waqas Mazhar, M.; Maqbool, M.; Alataway, A.; Dewidar, A.Z.; Elansary, H.O.; Yessoufou, K. Application of Smart Agricultural Practices in Wheat Crop to Increase Yield and Mitigate Emission of Greenhouse Gases for Sustainable Ecofriendly Environment. Sustainability 2022, 14, 10453. [Google Scholar] [CrossRef]
- Chinese Team Finds New Ways to Enhance Wheat Yield. Available online: https://www.isaaa.org/kc/cropbiotechupdate/article/default.asp?ID=19643#:~:text=Scientists%20from%20the%20Chinese%20Academy,are%20featured%20in%20Nature%20Plants (accessed on 23 June 2023).
- Trono, D.; Pecchioni, N. Candidate Genes Associated with Abiotic Stress Response in Plants as Tools to Engineer Tolerance to Drought, Salinity and Extreme Temperatures in Wheat: An Overview. Plants 2022, 11, 3358. [Google Scholar] [CrossRef]
- Shiqing, G.; Huijun, X.; Xianguo, C.; Ming, C.; Zhaoshi, X.; Liancheng, L.; Xingguo, Y.; Lipu, D.; Xiaoyan, H.; Youzhi, M. Improvement of Wheat Drought and Salt Tolerance by Expression of a Stress-Inducible Transcription FactorGmDREB of Soybean (Glycine max). Chin. Sci. Bull. 2005, 50, 2714–2723. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, M.; Guo, J.; Wang, Y.; Min, D.; Jiang, Q.; Ji, H.; Huang, C.; Wei, W.; Xu, H.; et al. Overexpression of Soybean DREB1 Enhances Drought Stress Tolerance of Transgenic Wheat in the Field. J. Exp. Bot. 2020, 71, 1842–1857. [Google Scholar] [CrossRef]
- Noor, S.; Ali, S.; Rahman, H.-; Farhatullah, F.; Ali, G.M. Comparative Study of Transgenic (DREB1A) and Non-Transgenic Wheat Lines on Relative Water Content, Sugar, Proline and Chlorophyll under Drought and Salt Stresses. Sarhad J. Agric. 2018, 34, 986–993. [Google Scholar] [CrossRef]
- Saint Pierre, C.; Crossa, J.L.; Bonnett, D.; Yamaguchi-Shinozaki, K.; Reynolds, M.P. Phenotyping Transgenic Wheat for Drought Resistance. J. Exp. Bot. 2012, 63, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.-Q.; Chen, M.; Xia, L.-Q.; Xiu, H.-J.; Xu, Z.-S.; Li, L.-C.; Zhao, C.-P.; Cheng, X.-G.; Ma, Y.-Z. A Cotton (Gossypium hirsutum) DRE-Binding Transcription Factor Gene, GhDREB, Confers Enhanced Tolerance to Drought, High Salt, and Freezing Stresses in Transgenic Wheat. Plant Cell Rep. 2009, 28, 301–311. [Google Scholar] [CrossRef]
- Saad, A.S.I.; Li, X.; Li, H.-P.; Huang, T.; Gao, C.-S.; Guo, M.-W.; Cheng, W.; Zhao, G.-Y.; Liao, Y.-C. A Rice Stress-Responsive NAC Gene Enhances Tolerance of Transgenic Wheat to Drought and Salt Stresses. Plant Sci. 2013, 203–204, 33–40. [Google Scholar] [CrossRef]
- Rong, W.; Qi, L.; Wang, A.; Ye, X.; Du, L.; Liang, H.; Xin, Z.; Zhang, Z. The ERF Transcription Factor TaERF3 Promotes Tolerance to Salt and Drought Stresses in Wheat. Plant Biotechnol. J. 2014, 12, 468–479. [Google Scholar] [CrossRef]
- Qiu, D.; Hu, W.; Zhou, Y.; Xiao, J.; Hu, R.; Wei, Q.; Zhang, Y.; Feng, J.; Sun, F.; Sun, J.; et al. TaASR1-D Confers Abiotic Stress Resistance by Affecting ROS Accumulation and ABA Signalling in Transgenic Wheat. Plant Biotechnol. J. 2021, 19, 1588–1601. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Y.; Li, T.; Ni, C.; Bai, X.; Lin, R.; Xiao, K. TaNF-YA7-5B, a Gene Encoding Nuclear Factor Y (NF–Y) Subunit A in Triticum aestivum, Confers Plant Tolerance to PEG-Inducing Dehydration Simulating Drought through Modulating Osmotic Stress-Associated Physiological Processes. Plant Physiol. Biochem. 2022, 188, 81–96. [Google Scholar] [CrossRef]
- Wei, J.; Fang, Y.; Jiang, H.; Wu, X.; Zuo, J.; Xia, X.; Li, J.; Stich, B.; Cao, H.; Liu, Y. Combining QTL Mapping and Gene Co-Expression Network Analysis for Prediction of Candidate Genes and Molecular Network Related to Yield in Wheat. BMC Plant Biol. 2022, 22, 288. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology: Top 10 Fungal Pathogens. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Hafeez, A.N.; Arora, S.; Ghosh, S.; Gilbert, D.; Bowden, R.L.; Wulff, B.B.H. Creation and Judicious Application of a Wheat Resistance Gene Atlas. Mol. Plant 2021, 14, 1053–1070. [Google Scholar] [CrossRef]
- Mapuranga, J.; Zhang, L.; Zhang, N.; Yang, W. The Haustorium: The Root of Biotrophic Fungal Pathogens. Front. Plant Sci. 2022, 13, 963705. [Google Scholar] [CrossRef]
- Figlan, S.; Ntushelo, K.; Mwadzingeni, L.; Terefe, T.; Tsilo, T.J.; Shimelis, H. Breeding Wheat for Durable Leaf Rust Resistance in Southern Africa: Variability, Distribution, Current Control Strategies, Challenges and Future Prospects. Front. Plant Sci. 2020, 11, 549. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, P.; Dodds, P.; Lagudah, E. How Target-Sequence Enrichment and Sequencing (TEnSeq) Pipelines Have Catalyzed Resistance Gene Cloning in the Wheat-Rust Pathosystem. Front. Plant Sci. 2020, 11, 678. [Google Scholar] [CrossRef]
- Prasad, P.; Bhardwaj, S.C.; Thakur, R.K.; Adhikari, S.; Gangwar, O.P.; Lata, C.; Kumar, S. Prospects of Climate Change Effects on Wheat Diseases. J. Cereal Res. 2021, 13, 2. [Google Scholar] [CrossRef]
- Jeger, M.; Beresford, R.; Bock, C.; Brown, N.; Fox, A.; Newton, A.; Vicent, A.; Xu, X.; Yuen, J. Global Challenges Facing Plant Pathology: Multidisciplinary Approaches to Meet the Food Security and Environmental Challenges in the Mid-Twenty-First Century. CABI Agric. Biosci. 2021, 2, 20. [Google Scholar] [CrossRef]
- Predicting wheat stripe rust epidemics according to influential climatic variables. J. Plant Prot. Res. 2023, 59, 519–528. [CrossRef]
- Naseri, B.; Sasani, S. Cultivar, Planting Date and Weather Linked to Wheat Leaf Rust Development. Cereal Res. Commun. 2020, 48, 203–210. [Google Scholar] [CrossRef]
- Dmochowska-Boguta, M.; Kloc, Y.; Zielezinski, A.; Werecki, P.; Nadolska-Orczyk, A.; Karlowski, W.M.; Orczyk, W. TaWAK6 Encoding Wall-Associated Kinase Is Involved in Wheat Resistance to Leaf Rust Similar to Adult Plant Resistance. PLoS ONE 2020, 15, e0227713. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Gao, L.; Dong, S.; Sun, Y.; Shen, Q.; Guo, S. Role of Silicon on Plant–Pathogen Interactions. Front. Plant Sci. 2017, 8, 701. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Tian, W.; Tao, F.; Wang, J.; Shang, H.; Chen, X.; Xu, X.; Hu, X. TaRPM1 Positively Regulates Wheat High-Temperature Seedling-Plant Resistance to Puccinia striiformis f. sp. tritici. Front. Plant Sci. 2020, 10, 1679. [Google Scholar] [CrossRef] [Green Version]
- Naseri, B.; Marefat, A. Wheat Stripe Rust Epidemics in Interaction with Climate, Genotype and Planting Date. Eur. J. Plant. Pathol. 2019, 154, 1077–1089. [Google Scholar] [CrossRef]
- Naseri, B. Sowing, Field Size, and Soil Characteristics Affect Bean-Fusarium-Wilt Pathosystems. J. Plant Dis. Prot. 2014, 121, 171–176. [Google Scholar] [CrossRef]
- Kaur, B.; Ranawana, V.; Henry, J. The Glycemic Index of Rice and Rice Products: A Review, and Table of GI Values. Crit. Rev. Food Sci. Nutr. 2016, 56, 215–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Kurata, N.; Wei, X.; Wang, Z.-X.; Wang, A.; Zhao, Q.; Zhao, Y.; Liu, K.; Lu, H.; Li, W.; et al. A Map of Rice Genome Variation Reveals the Origin of Cultivated Rice. Nature 2012, 490, 497–501. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhao, Y.; Jiang, C.; Wang, L.; Chen, L.; Li, F.; Zhang, Y.; Pan, Y.; Zhang, T. Evolution of Different Rice Ecotypes and Genetic Basis of Flooding Adaptability in Deepwater Rice by GWAS. BMC Plant. Biol. 2022, 22, 526. [Google Scholar] [CrossRef]
- Bin Rahman, A.N.M.R.; Zhang, J. Trends in Rice Research: 2030 and Beyond. Food Energy Secur. 2023, 12, e390. [Google Scholar] [CrossRef]
- Mohd Hanafiah, N.; Mispan, M.S.; Lim, P.E.; Baisakh, N.; Cheng, A. The 21st Century Agriculture: When Rice Research Draws Attention to Climate Variability and How Weedy Rice and Underutilized Grains Come in Handy. Plants 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Fornasiero, A.; Wing, R.A.; Ronald, P. Rice Domestication. Curr. Biol. 2022, 32, R20–R24. [Google Scholar] [CrossRef]
- Chen, R.; Deng, Y.; Ding, Y.; Guo, J.; Qiu, J.; Wang, B.; Wang, C.; Xie, Y.; Zhang, Z.; Chen, J.; et al. Rice Functional Genomics: Decades’ Efforts and Roads Ahead. Sci. China Life Sci. 2022, 65, 33–92. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Hu, T.; Mahmoud, A.; Li, J.; Zhu, R.; Jiao, X.; Jing, P. A Quantitative Review of the Effects of Biochar Application on Rice Yield and Nitrogen Use Efficiency in Paddy Fields: A Meta-Analysis. Sci. Total Environ. 2022, 830, 154792. [Google Scholar] [CrossRef]
- Aung, A.; Youn, W.B.; Seo, J.M.; Dao, H.T.T.; Han, S.H.; Cho, M.S.; Park, B.B. Effects of Three Biomaterials Mixed with Growing Media on Seedling Quality of Prunus Sargentii. For. Sci. Technol. 2019, 15, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Gao, J.; Li, X.; Zhang, S.; Wang, H. Nitrate Accumulation and Leaching in Surface and Ground Water Based on Simulated Rainfall Experiments. PLoS ONE 2015, 10, e0136274. [Google Scholar] [CrossRef] [Green Version]
- Jadon, P.; Selladurai, R.; Yadav, S.S.; Coumar, M.V.; Dotaniya, M.L.; Singh, A.K.; Bhadouriya, J.; Kundu, S. Volatilization and Leaching Losses of Nitrogen from Different Coated Urea Fertilizers. J. Soil Sci. Plant Nutr. 2018, 18, 1036–1047. [Google Scholar] [CrossRef] [Green Version]
- Hindersah, R.; Kalay, A.M.; Talahaturuson, A. Rice Yield Grown in Different Fertilizer Combination and Planting Methods: Case Study in Buru Island, Indonesia. Open Agric. 2022, 7, 871–881. [Google Scholar] [CrossRef]
- Misheck, C.; Fanuel, C. Effect of Ethephon and Planting Density on Lodged Plant Percentage and Crop Yield in Maize (Zea mays L.). Afr. J. Plant Sci. 2014, 8, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, Y.; Ye, D.; Wang, W.; Qiu, X.; Duan, L.; Li, Z.; Zhang, M. Ethephon Improved Stalk Strength of Maize (Zea mays L.) Mainly through Altering Internode Morphological Traits to Modulate Mechanical Properties under Field Conditions. Agronomy 2019, 9, 186. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, J.; Wang, A.; Chen, J. Chemical Inhibitors of Methanogenesis and Putative Applications. Appl. Microbiol. Biotechnol. 2011, 89, 1333–1340. [Google Scholar] [CrossRef]
- Cho, S.R.; Verma, P.P.; Das, S.; Kim, G.W.; Lim, J.Y.; Kim, P.J. A New Approach to Suppress Methane Emissions from Rice Cropping Systems Using Ethephon. Sci. Total Environ. 2022, 804, 150159. [Google Scholar] [CrossRef]
- Hou, P.; Yu, Y.; Xue, L.; Petropoulos, E.; He, S.; Zhang, Y.; Pandey, A.; Xue, L.; Yang, L.; Chen, D. Effect of Long Term Fertilization Management Strategies on Methane Emissions and Rice Yield. Sci. Total Environ. 2020, 725, 138261. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, H.; Wang, J.; Zhang, J.; Liu, G.; Zhou, S. Effect of Moisture Gradient on Rice Yields and Greenhouse Gas Emissions from Rice Paddies. Environ. Sci. Pollut. Res. 2019, 26, 33416–33426. [Google Scholar] [CrossRef]
- Iizumi, T.; Luo, J.-J.; Challinor, A.J.; Sakurai, G.; Yokozawa, M.; Sakuma, H.; Brown, M.E.; Yamagata, T. Impacts of El Niño Southern Oscillation on the Global Yields of Major Crops. Nat. Commun. 2014, 5, 3712. [Google Scholar] [CrossRef] [Green Version]
- Challinor, A.J.; Watson, J.; Lobell, D.B.; Howden, S.M.; Smith, D.R.; Chhetri, N. A Meta-Analysis of Crop Yield under Climate Change and Adaptation. Nat. Clim. Chang. 2014, 4, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Mie, A.; Andersen, H.R.; Gunnarsson, S.; Kahl, J.; Kesse-Guyot, E.; Rembiałkowska, E.; Quaglio, G.; Grandjean, P. Human Health Implications of Organic Food and Organic Agriculture: A Comprehensive Review. Environ. Health 2017, 16, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural Sustainability and Intensive Production Practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Seufert, V.; Ramankutty, N.; Foley, J.A. Comparing the Yields of Organic and Conventional Agriculture. Nature 2012, 485, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Ponisio, L.C.; M’Gonigle, L.K.; Mace, K.C.; Palomino, J.; de Valpine, P.; Kremen, C. Diversification Practices Reduce Organic to Conventional Yield Gap. Proc. R. Soc. B 2015, 282, 20141396. [Google Scholar] [CrossRef] [Green Version]
- Khumairoh, U.; Groot, J.C.J.; Lantinga, E.A. Complex Agro-Ecosystems for Food Security in a Changing Climate: Complex Agro-Ecosystems. Ecol. Evol. 2012, 2, 1696–1704. [Google Scholar] [CrossRef]
- Ramanathan, K.; Sangeeviraman, V.; Chandrahasan, P.; Chaudhary, B.N.; Ramachandra, S.S. Integration of Fish Culture and Poultry Rearing in Transplanted Rice for Nutritional Security in Smallholder Farms. Sci. Rep. 2020, 10, 10566. [Google Scholar] [CrossRef]
- Elahi, E.; Khalid, Z.; Tauni, M.Z.; Zhang, H.; Lirong, X. Extreme Weather Events Risk to Crop-Production and the Adaptation of Innovative Management Strategies to Mitigate the Risk: A Retrospective Survey of Rural Punjab, Pakistan. Technovation 2022, 117, 102255. [Google Scholar] [CrossRef]
- Khumairoh, U.; Lantinga, E.A.; Schulte, R.P.O.; Suprayogo, D.; Groot, J.C.J. Complex Rice Systems to Improve Rice Yield and Yield Stability in the Face of Variable Weather Conditions. Sci. Rep. 2018, 8, 14746. [Google Scholar] [CrossRef] [Green Version]
- Zuo, J.; Li, J. Molecular Genetic Dissection of Quantitative Trait Loci Regulating Rice Grain Size. Annu. Rev. Genet. 2014, 48, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Thakur, J.K.; Agarwal, P.; Parida, S.; Bajaj, D.; Pasrija, R. Sequence and Expression Analyses of KIX Domain Proteins Suggest Their Importance in Seed Development and Determination of Seed Size in Rice, and Genome Stability in Arabidopsis. Mol. Genet. Genom. 2013, 288, 329–346. [Google Scholar] [CrossRef]
- Dwivedi, N.; Maji, S.; Waseem, M.; Thakur, P.; Kumar, V.; Parida, S.K.; Thakur, J.K. The Mediator Subunit OsMED15a Is a Transcriptional Co-Regulator of Seed Size/Weight–Modulating Genes in Rice. Biochim. Biophys. Acta 2019, 1862, 194432. [Google Scholar] [CrossRef]
- Wei, S.; Li, X.; Lu, Z.; Zhang, H.; Ye, X.; Zhou, Y.; Li, J.; Yan, Y.; Pei, H.; Duan, F.; et al. A Transcriptional Regulator That Boosts Grain Yields and Shortens the Growth Duration of Rice. Science 2022, 377, eabi8455. [Google Scholar] [CrossRef]
- Van Der Pol, L.K.; Nester, B.; Schlautman, B.; Crews, T.E.; Cotrufo, M.F. Perennial Grain Kernza® Fields Have Higher Particulate Organic Carbon at Depth than Annual Grain Fields. Can. J. Soil. Sci. 2022, 102, 1005–1009. [Google Scholar] [CrossRef]
- Crews, T.E.; Carton, W.; Olsson, L. Is the Future of Agriculture Perennial? Imperatives and Opportunities to Reinvent Agriculture by Shifting from Annual Monocultures to Perennial Polycultures. Glob. Sustain. 2018, 1, e11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Huang, G.; Zhang, Y.; Lv, X.; Wan, K.; Liang, J.; Feng, Y.; Dao, J.; Wu, S.; Zhang, L.; et al. Sustained Productivity and Agronomic Potential of Perennial Rice. Nat. Sustain. 2022, 6, 28–38. [Google Scholar] [CrossRef]
- Helliwell, E.E.; Yang, Y. Molecular Strategies to Improve Rice Disease Resistance. In Rice Protocols; Yang, Y., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2013; Volume 956, pp. 285–309. ISBN 978-1-62703-193-6. [Google Scholar]
- Helliwell, E.E.; Wang, Q.; Yang, Y. Transgenic Rice with Inducible Ethylene Production Exhibits Broad-Spectrum Disease Resistance to the Fungal Pathogens Magnaporthe oryzae and Rhizoctonia solani. Plant Biotechnol. J. 2013, 11, 33–42. [Google Scholar] [CrossRef]
- Ashkani, S.; Yusop, M.R.; Shabanimofrad, M.; Azady, A.; Ghasemzadeh, A.; Azizi, P.; Latif, M.A. Allele Mining Strategies: Principles and Utilisation for Blast Resistance Genes in Rice (Oryza sativa L.). Curr. Issues Mol. Biol. 2015, 17, 57–73. [Google Scholar]
- Ashkani, S.; Rafii, M.Y.; Shabanimofrad, M.; Miah, G.; Sahebi, M.; Azizi, P.; Tanweer, F.A.; Akhtar, M.S.; Nasehi, A. Molecular Breeding Strategy and Challenges Towards Improvement of Blast Disease Resistance in Rice Crop. Front. Plant Sci. 2015, 6, 886. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Das, A.; Devanna, B.N.; Thakur, S.; Singh, P.K.; Singh, N.K.; Sharma, T.R. Mining of Rice Blast Resistance Gene Pi54 Shows Effect of Single Nucleotide Polymorphisms on Phenotypic Expression of the Alleles. Eur. J. Plant Pathol. 2013, 137, 55–65. [Google Scholar] [CrossRef]
- Ramkumar, G.; Srinivasarao, K.; Mohan, K.M.; Sudarshan, I.; Sivaranjani, A.K.P.; Gopalakrishna, K.; Neeraja, C.N.; Balachandran, S.M.; Sundaram, R.M.; Prasad, M.S.; et al. Development and Validation of Functional Marker Targeting an InDel in the Major Rice Blast Disease Resistance Gene Pi54 (Pik h ). Mol. Breed. 2011, 27, 129–135. [Google Scholar] [CrossRef]
- Assem, S.K. Maize, Tropical (Zea mays L.). In Agrobacterium Protocols; Wang, K., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2015; Volume 1223, pp. 119–134. ISBN 978-1-4939-1694-8. [Google Scholar]
- Gwirtz, J.A.; Garcia-Casal, M.N. Processing Maize Flour and Corn Meal Food Products. Ann. N. Y. Acad. Sci. 2014, 1312, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Schnable, J.C. Genome Evolution in Maize: From Genomes Back to Genes. Annu. Rev. Plant Biol. 2015, 66, 329–343. [Google Scholar] [CrossRef]
- Wang, B.; Lin, Z.; Li, X.; Zhao, Y.; Zhao, B.; Wu, G.; Ma, X.; Wang, H.; Xie, Y.; Li, Q.; et al. Genome-Wide Selection and Genetic Improvement during Modern Maize Breeding. Nat. Genet. 2020, 52, 565–571. [Google Scholar] [CrossRef]
- Hou, H.; Ma, W.; Noor, M.A.; Tang, L.; Li, C.; Ding, Z.; Zhao, M. Quantitative Design of Yield Components to Simulate Yield Formation for Maize in China. J. Integr. Agric. 2020, 19, 668–679. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic Strategies for Improving Crop Yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Miao, P.; Han, K.; Tang, Y.; Wang, B.; Lin, T.; Cheng, W. Recent Advances in Carbon Nanodots: Synthesis, Properties and Biomedical Applications. Nanoscale 2015, 7, 1586–1595. [Google Scholar] [CrossRef]
- Lowry, G.V.; Avellan, A.; Gilbertson, L.M. Opportunities and Challenges for Nanotechnology in the Agri-Tech Revolution. Nat. Nanotechnol. 2019, 14, 517–522. [Google Scholar] [CrossRef]
- Gilbertson, L.M.; Pourzahedi, L.; Laughton, S.; Gao, X.; Zimmerman, J.B.; Theis, T.L.; Westerhoff, P.; Lowry, G.V. Guiding the Design Space for Nanotechnology to Advance Sustainable Crop Production. Nat. Nanotechnol. 2020, 15, 801–810. [Google Scholar] [CrossRef]
- Kah, M.; Kookana, R.S.; Gogos, A.; Bucheli, T.D. A Critical Evaluation of Nanopesticides and Nanofertilizers against Their Conventional Analogues. Nat. Nanotechnol. 2018, 13, 677–684. [Google Scholar] [CrossRef]
- Giraldo, J.P.; Wu, H.; Newkirk, G.M.; Kruss, S. Nanobiotechnology Approaches for Engineering Smart Plant Sensors. Nat. Nanotechnol. 2019, 14, 541–553. [Google Scholar] [CrossRef]
- Wang, C.; Yang, H.; Chen, F.; Yue, L.; Wang, Z.; Xing, B. Nitrogen-Doped Carbon Dots Increased Light Conversion and Electron Supply to Improve the Corn Photosystem and Yield. Environ. Sci. Technol. 2021, 55, 12317–12325. [Google Scholar] [CrossRef]
- Wild, M. Global Dimming and Brightening: A Review. J. Geophys. Res. 2009, 114, D00D16. [Google Scholar] [CrossRef] [Green Version]
- Hinkelman, L.M.; Stackhouse, P.W.; Wielicki, B.A.; Zhang, T.; Wilson, S.R. Surface Insolation Trends from Satellite and Ground Measurements: Comparisons and Challenges. J. Geophys. Res. 2009, 114, D00D20. [Google Scholar] [CrossRef] [Green Version]
- Wild, M.; Gilgen, H.; Roesch, A.; Ohmura, A.; Long, C.N.; Dutton, E.G.; Forgan, B.; Kallis, A.; Russak, V.; Tsvetkov, A. From Dimming to Brightening: Decadal Changes in Solar Radiation at Earth’s Surface. Science 2005, 308, 847–850. [Google Scholar] [CrossRef] [Green Version]
- Tollenaar, M.; Fridgen, J.; Tyagi, P.; Stackhouse Jr, P.W.; Kumudini, S. The Contribution of Solar Brightening to the US Maize Yield Trend. Nat. Clim. Chang. 2017, 7, 275–278. [Google Scholar] [CrossRef]
- Sun, X.; Qi, X.; Wang, W.; Liu, X.; Zhao, H.; Wu, C.; Chang, X.; Zhang, M.; Chen, H.; Gong, G. Etiology and Symptoms of Maize Leaf Spot Caused by Bipolaris Spp. in Sichuan, China. Pathogens 2020, 9, 229. [Google Scholar] [CrossRef] [Green Version]
- Azra, A.; Hussain, S. Screening of Maize Genotypes against Southern Corn Leaf Blight (Bipolaris maydis) under Artificial Epiphytotic Conditions. Sarhad J. Agric. 2019, 35, 1020–1356. [Google Scholar] [CrossRef]
- Boddupalli, P.; Suresh, L.M.; Mwatuni, F.; Beyene, Y.; Makumbi, D.; Gowda, M.; Olsen, M.; Hodson, D.; Worku, M.; Mezzalama, M.; et al. Maize Lethal Necrosis (MLN): Efforts toward Containing the Spread and Impact of a Devastating Transboundary Disease in Sub-Saharan Africa. Virus Res. 2020, 282, 197943. [Google Scholar] [CrossRef]
- Yang, Q.; Balint-Kurti, P.; Xu, M. Quantitative Disease Resistance: Dissection and Adoption in Maize. Mol. Plant 2017, 10, 402–413. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liu, H.; Gong, Y.; Tao, Y.; Jiang, L.; Zuo, W.; Yang, Q.; Ye, J.; Lai, J.; Wu, J.; et al. An Atypical Thioredoxin Imparts Early Resistance to Sugarcane Mosaic Virus in Maize. Mol. Plant 2017, 10, 483–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Deng, S.; Liu, B.; Tao, Y.; Ai, H.; Liu, J.; Zhang, Y.; Zhao, Y.; Xu, M. A Helitron-Induced RabGDIα Variant Causes Quantitative Recessive Resistance to Maize Rough Dwarf Disease. Nat. Commun. 2020, 11, 495. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Jiang, L.; Liu, Q.; Zhang, Y.; Zhang, R.; Ingvardsen, C.R.; Frei, U.K.; Wang, B.; Lai, J.; Lübberstedt, T.; et al. Combined Linkage and Association Mapping Reveals Candidates for Scmv1, a Major Locus Involved in Resistance to Sugarcane Mosaic Virus (SCMV) in Maize. BMC Plant Biol. 2013, 13, 162. [Google Scholar] [CrossRef] [Green Version]
- Leng, P.; Ji, Q.; Asp, T.; Frei, U.K.; Ingvardsen, C.R.; Xing, Y.; Studer, B.; Redinbaugh, M.; Jones, M.; Gajjar, P.; et al. Auxin Binding Protein 1 Reinforces Resistance to Sugarcane Mosaic Virus in Maize. Mol. Plant 2017, 10, 1357–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Hou, Q.; Wang, L.; Zhang, T.; Zhao, W.; Yan, T.; Zhao, L.; Li, J.; Wan, X. Genome-Wide Identification and Characterization of Polygalacturonase Gene Family in Maize (Zea mays L.). Int. J. Mol. Sci. 2021, 22, 10722. [Google Scholar] [CrossRef]
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. A Chromosome Conformation Capture Ordered Sequence of the Barley Genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Sato, K. History and Future Perspectives of Barley Genomics. DNA Res. 2020, 27, dsaa023. [Google Scholar] [CrossRef]
- Cammarano, D.; Ronga, D.; Francia, E.; Akar, T.; Al-Yassin, A.; Benbelkacem, A.; Grando, S.; Romagosa, I.; Stanca, A.M.; Pecchioni, N. Genetic and Management Effects on Barley Yield and Phenology in the Mediterranean Basin. Front. Plant Sci. 2021, 12, 655406. [Google Scholar] [CrossRef]
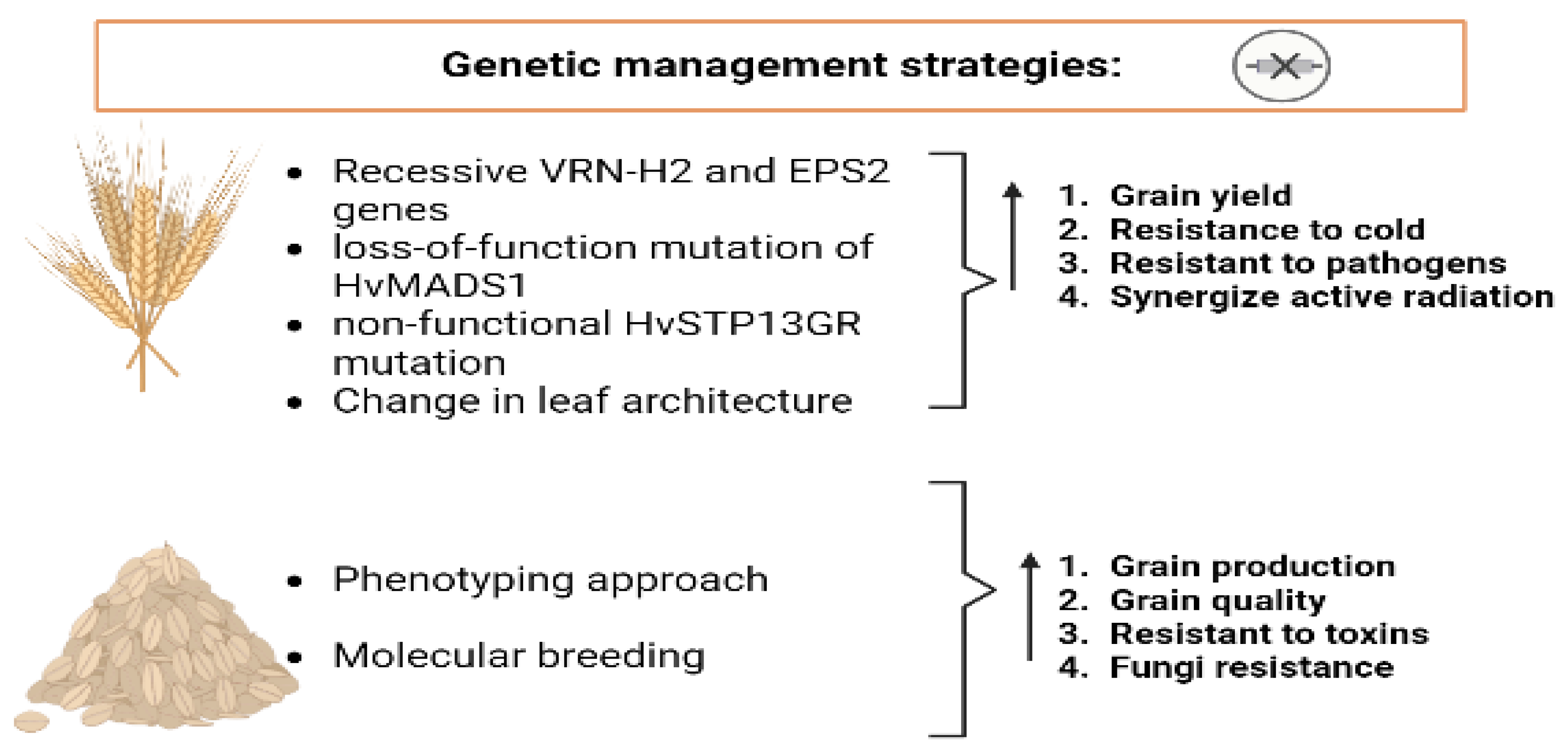
- Li, G.; Kuijer, H.N.J.; Yang, X.; Liu, H.; Shen, C.; Shi, J.; Betts, N.; Tucker, M.R.; Liang, W.; Waugh, R.; et al. MADS1 Maintains Barley Spike Morphology at High Ambient Temperatures. Nat. Plants 2021, 7, 1093–1107. [Google Scholar] [CrossRef]
- Rodrigues, O.; Minella, E.; Costenaro, E.R.; Scariotto, S.; Marchese, J.A. Improvement in Brazilian Barley Breeding: Changes in Developmental Phases and Ecophysiological Traits. Front. Plant Sci. 2022, 13, 1032243. [Google Scholar] [CrossRef]
- Wiegmann, M.; Maurer, A.; Pham, A.; March, T.J.; Al-Abdallat, A.; Thomas, W.T.B.; Bull, H.J.; Shahid, M.; Eglinton, J.; Baum, M.; et al. Barley Yield Formation under Abiotic Stress Depends on the Interplay between Flowering Time Genes and Environmental Cues. Sci. Rep. 2019, 9, 6397. [Google Scholar] [CrossRef] [Green Version]
- Skoppek, C.I.; Punt, W.; Heinrichs, M.; Ordon, F.; Wehner, G.; Boch, J.; Streubel, J. The Barley HvSTP13GR Mutant Triggers Resistance against Biotrophic Fungi. Mol. Plant Pathol. 2022, 23, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Barati, M.; Majidi, M.M.; Mostafavi, F.; Mirlohi, A.; Safari, M.; Karami, Z. Evaluation of Wild Barley Species as Possible Sources of Drought Tolerance for Arid Environments. Plant Genet. Resour. 2018, 16, 209–217. [Google Scholar] [CrossRef]
- Serna-Saldivar, S.O. Cereal Grains: Properties, Processing, and Nutritional Attributes; CRC Press: Boca Raton, FL, USA, 2016; ISBN 978-0-429-11211-9. [Google Scholar]
- Harwood, W.A. An Introduction to Barley: The Crop and the Model. In Barley; Harwood, W.A., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; Volume 1900, pp. 1–5. ISBN 978-1-4939-8942-3. [Google Scholar]
- Chen, G.; Liu, Y.; Ma, J.; Zheng, Z.; Wei, Y.; McIntyre, C.L.; Zheng, Y.-L.; Liu, C. A Novel and Major Quantitative Trait Locus for Fusarium Crown Rot Resistance in a Genotype of Wild Barley (Hordeum spontaneum L.). PLoS ONE 2013, 8, e58040. [Google Scholar] [CrossRef] [Green Version]
- Menon, R.; Gonzalez, T.; Ferruzzi, M.; Jackson, E.; Winderl, D.; Watson, J. Oats—From Farm to Fork. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2016; Volume 77, pp. 1–55. ISBN 978-0-12-804772-9. [Google Scholar]
- Chen, O.; Mah, E.; Dioum, E.; Marwaha, A.; Shanmugam, S.; Malleshi, N.; Sudha, V.; Gayathri, R.; Unnikrishnan, R.; Anjana, R.M.; et al. The Role of Oat Nutrients in the Immune System: A Narrative Review. Nutrients 2021, 13, 1048. [Google Scholar] [CrossRef]
- Smulders, M.J.M.; van de Wiel, C.C.M.; van den Broeck, H.C.; van der Meer, I.M.; Israel-Hoevelaken, T.P.M.; Timmer, R.D.; van Dinter, B.-J.; Braun, S.; Gilissen, L.J.W.J. Oats in Healthy Gluten-Free and Regular Diets: A Perspective. Food Res. Int. 2018, 110, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Tomaszewska, P.; Schwarzacher, T.; Heslop-Harrison, J.S. (Pat) Oat Chromosome and Genome Evolution Defined by Widespread Terminal Intergenomic Translocations in Polyploids. Front. Plant Sci. 2022, 13, 1026364. [Google Scholar] [CrossRef]
- Kamal, N.; Tsardakas Renhuldt, N.; Bentzer, J.; Gundlach, H.; Haberer, G.; Juhász, A.; Lux, T.; Bose, U.; Tye-Din, J.A.; Lang, D.; et al. The Mosaic Oat Genome Gives Insights into a Uniquely Healthy Cereal Crop. Nature 2022, 606, 113–119. [Google Scholar] [CrossRef]
- Allwood, J.W.; Martinez-Martin, P.; Xu, Y.; Cowan, A.; Pont, S.; Griffiths, I.; Sungurtas, J.; Clarke, S.; Goodacre, R.; Marshall, A.; et al. Assessing the Impact of Nitrogen Supplementation in Oats across Multiple Growth Locations and Years with Targeted Phenotyping and High-Resolution Metabolite Profiling Approaches. Food Chem. 2021, 355, 129585. [Google Scholar] [CrossRef]
- Martínez-López, J.A.; López-Urrea, R.; Martínez-Romero, Á.; Pardo, J.J.; Montoya, F.; Domínguez, A. Improving the Sustainability and Profitability of Oat and Garlic Crops in a Mediterranean Agro-Ecosystem under Water-Scarce Conditions. Agronomy 2022, 12, 1950. [Google Scholar] [CrossRef]
- Gavrilova, O.P.; Gagkaeva, T.Y.; Orina, A.S.; Markova, A.S.; Kabashov, A.D.; Loskutov, I.G. Resistance of Oat Breeding Lines to Grain Contamination with Fusarium Langsethiae and T-2/HT-2 Toxins. Vavilov J. Genet. Breed. 2021, 25, 732–739. [Google Scholar] [CrossRef]
- IMPROVED CROP VARIETIES CAN HELP MITIGATE RISING GRAIN PRICES. Available online: https://www.icarda.org/media/news/new-plan-meet-wheat-demand-egypt (accessed on 23 June 2023).
- Campanhola, C.; Pandey, S. (Eds.) Crop Variety Improvements. In Sustainable Food and Agriculture; Elsevier: Amsterdam, The Netherlands, 2019; pp. 205–207. ISBN 978-0-12-812134-4. [Google Scholar]
- IMPROVED VARIETIES OF WHEAT, PADDY, MUSTARD AND PIGEON PEA. Available online: https://nif.org.in/innovation/improved_varieties/16 (accessed on 23 June 2023).
- Ulzen, O.O.; Buri, M.M.; Sekyi-Annan, E.; Devkota, K.P.; Dossou-Yovo, E.R.; Essel Ayamba, B.; Adjei, E.O. Yield Potentials of Improved Rice Varieties for Increased Lowland Rice Production within the Mankran Watershed in Ghana. Plant Prod. Sci. 2023, 26, 17–27. [Google Scholar] [CrossRef]
- Cairns, J.E.; Prasanna, B. Developing and Deploying Climate-Resilient Maize Varieties in the Developing World. Curr. Opin. Plant Biol. 2018, 45, 226–230. [Google Scholar] [CrossRef]
- Andersen, E.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M. Disease Resistance Mechanisms in Plants. Genes 2018, 9, 339. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albahri, G.; Alyamani, A.A.; Badran, A.; Hijazi, A.; Nasser, M.; Maresca, M.; Baydoun, E. Enhancing Essential Grains Yield for Sustainable Food Security and Bio-Safe Agriculture through Latest Innovative Approaches. Agronomy 2023, 13, 1709. https://doi.org/10.3390/agronomy13071709
Albahri G, Alyamani AA, Badran A, Hijazi A, Nasser M, Maresca M, Baydoun E. Enhancing Essential Grains Yield for Sustainable Food Security and Bio-Safe Agriculture through Latest Innovative Approaches. Agronomy. 2023; 13(7):1709. https://doi.org/10.3390/agronomy13071709
Chicago/Turabian StyleAlbahri, Ghosoon, Amal A. Alyamani, Adnan Badran, Akram Hijazi, Mohamad Nasser, Marc Maresca, and Elias Baydoun. 2023. "Enhancing Essential Grains Yield for Sustainable Food Security and Bio-Safe Agriculture through Latest Innovative Approaches" Agronomy 13, no. 7: 1709. https://doi.org/10.3390/agronomy13071709