1. Introduction
Global climate change leads to an active expansion of habitats for phytopathogenic microorganisms [
1,
2]. In response, the use of pesticides in global agriculture is increasing [
3]. However, most pesticides, while being primarily effective against phytopathogens, could also be toxic and pollute the environment [
4] by penetrating soil and groundwater and accumulating in plants [
5]. They may have a negative impact on animals [
6,
7,
8,
9] and humans [
10,
11,
12]. In this regard, it is extremely important to search for new effective and environmentally friendly agents against phytopathogens [
13]. In this regard, various active bioagents of plant origin and biopesticides are being actively investigated now [
14,
15,
16,
17]. Biopesticides are natural compounds of biological origin that are used to control various agricultural pests that infect plants in forests, gardens, and agricultural lands [
16]. They are different depending on the origin: microbial, produced with the help of genetically modified organisms and biochemical pesticides (pheromones, plant extracts, and oils, insect growth regulators). Basically, biopesticides are aimed at regulating the number of insects. Thus, as an alternative to chemical pesticides, it was proposed to use essential oils, which are a complex mixture of hydrocarbons with traces of compounds based on sulfur and nitrogen extracted from plants. Key components of horticultural oils include paraffin and olefin. They are recommended to treat cultivated plants to protect them from harmful insects [
14]. In addition, insects and birds, extracts of various trees were used as an alternative to insecticides [
15]. Plant terpenoid compounds, flavonoids, alkaloids, polyphenols, cyanogenic glucosides, quinones, amides, aldehydes, thiophenes, amino acids, and saccharides are proposed to be used as attractants, nematicides, fungicides, repellents, insecticides, and insect growth regulators for pest control [
17].
Nanotechnologies are being actively introduced into various spheres of human economic activity [
18,
19,
20,
21,
22]. In particular, there are encouraging developments in the field of agrochemistry [
23,
24,
25,
26,
27,
28]. Innovative nanopesticides are nanomaterials developed for plant protection and characterized by the following properties: minimizing losses during the application, increasing leaf coverage, increasing stability, and reducing the amount of consumption of active substances. Nanopesticide preparations can be divided into self-organizing systems, such as liposomes, dendrimers, metallic and bimetallic nanoparticles (NPs), and active encapsulating ingredients, such as nanoemulsion, polymeric nanoparticles, lipid, nanoparticles, and nanotubes [
29]. Biopesticides can also be created on the basis of nanoparticles (NPs) [
13,
30], and we tested a few of them in this study. Nanobiopesticides are created by encapsulation of biomolecules extracted from plants, fungi, and bacteria, which have a prolonged action, or also with the help of numerous biomaterials synthesized by biogenic processes to obtain biocompatible and more effective biopesticides [
31]. Biologically derived nanopesticides can be made using any element, including metals such as Ag, Cu, SiO
2, and ZnO, with a wide spectrum of pest control efficiency [
32]. We have studied bionanopesticide selenium (Se) nanocomposites (NCs) based on natural polymer matrices and tested them in field conditions. Among them are Se NCs with different contents of Se NPs based on arabinogalactan (AG) [
33,
34,
35], starch (ST) [
36], and carrageenan (CAR) [
37], as well as silver NCs based on humic substances of various compositions (shales, coals, and pelloids) [
38,
39], manganese NCs based on AG and CAR, and sulfur NCs based on ST [
40]. It has been shown that these NCs reduce the viability of the phytopathogenic bacterium
Clavibacter sepedonicus and the phytopathogenic fungus
Phytophthora cactorum [
35]. Some of them stimulated the growth and development of potatoes in vitro, germination of soybean and pea seeds [
41], and increased biomass of potato tubers and quality of potato crop in field experiments. At the same time, the accumulation of NPs in plant tissues after their treatment with nanocomposites was not observed [
33,
34,
37]. It is also important to note that the studied NCs did not inhibit the viability of soil microorganisms
Acinetobacter guillouiae,
Rhodococcus erythropolis, and
Pseudomonas oryzihabitans [
35]. Published laboratory data on the biological activity of NCs indicate the promise and relative safety of their use against phytopathogens for plant health control and improvement [
33,
34,
37]. These experiments were carried out under in vitro conditions. However, for a comprehensive assessment of the effect of NCs on potatoes, it is necessary to conduct, in addition to laboratory experiments also, trials in natural conditions of potato vegetation and storage. This paper presents new data on the effect of Se NCs based on natural polymer matrices on the biomass, storage, and sprouting of potato tubers in the field in order to develop an effective and environmentally friendly growth stimulant for agricultural plants. Our main objective in this study was not so much to assess the effect of pre-planting treatment on the productivity and structure of potatoes, which would require multiple-year observations, but to study its immediate effect on the storage of the crop and its productivity in the next generation as a part of a systemic effect of nanocomposites. In our previous in vitro studies, we have shown that the NCs presented here have the properties of pesticides that kill pathogenic microflora but do not adversely affect the soil microflora and the plants themselves, which are treated with NCs [
33,
34,
35,
36,
37,
40,
42]. In this study, we studied the effect of NC on the productivity, germination, and storage of uninfected potatoes in the field conditions. At the same time, some growth-stimulating effects were found.
4. Discussion
Reactive oxygen species (ROS) are involved in the regulation of the most important physiological and biochemical cell processes both under and without stress [
49,
50,
51]. Tuber dormancy and germination in potatoes can also be controlled by manipulating the content of ROS, especially the amount of hydrogen peroxide (H
2O
2), through the inhibition of catalase (CAT) activity. In contrast to the hormonal regulation of potato dormancy, little attention has been paid to the role of ROS. The participation of ROS such as superoxide anion radical (O
2), H
2O
2, and hydroxyl radicals (OH·) in the regulation of dormancy and germination has been demonstrated in several plant species, particularly in grapes [
52]. The role of this process during potato germination is discussed in [
53,
54,
55].
When the concentration of H
2O
2 is excessive, the mitochondria remain dormant, and germination decreases. Indeed, high concentrations of H
2O
2 are toxic to plants, activates LPO, and causes damage to the membrane wall and a decrease in membrane integrity [
52]. Lipid peroxidation is a process in which free radicals (oxyl, peroxyl, hydroxyl) remove electrons from lipids and subsequently produce reactive intermediates that can enter into further reactions [
56]. As a result of this process, primary products are formed, hydroperoxides, also called diene conjugates (DC), which are considered the primary product of LPO [
57]. Therefore, as a biochemical marker of the effect of Se NCs on plant growth and development, we determined the content of primary LPO products, namely DC, in root and shoot tissues. Our results showed a decrease in the level of DC under the influence of Se/AG NC. The remaining Se NCs did not affect the level of DC (
Figure 2A). This result indicates that the treatment of tubers with Se NCs does not cause a stress response in the tuber cells. The decrease in the level of DC under the influence of Se/AG NC may be associated with the antioxidant activity of Se NP. It was shown that green synthesized Se NPs (25–200 μg/mL) reduced the amount of a free radical 2,2-diphenyl-1-picrylhydrazyl (DPPH) by 61% [
58]. Selenium NPs decorated with a different molecular weight of chitosan (1.5 kDa, 48 kDa, and 510 kDa) showed the strongest antioxidant activities. These NPs inactivated DPPH, ABTS+, and O
2− radicals [
59]. The antioxidant activity of Se NCs has been proven repeatedly in animals, including the use of LPO products as an indicator. The antioxidant activity of Se NCs in animal tissues is mainly associated with the regulation of the activity of Se-containing enzymes that is the family of glutathione peroxidases (GPX) and thioredoxin reductase (TR), which detoxify a wide range of peroxides, such as H
2O
2, phospholipid hydroperoxides, fatty acid hydroperoxides, and thymine hydroperoxyl groups [
60]. It has been shown in chickens that the addition of Se NP at doses of 0, 0.1, and 0.2 mg/kg to their diet increased the levels of GPX and superoxide dismutase (SOD) and reduced the level of the secondary LPO product, malondialdehyde (MDA) [
61]. The antioxidant and hepatoprotective effect of Se NP was found in the study of MDA content, total antioxidant capacity, GPX, SOD, glutathione (GSH), and CAT activity in rat liver tissue [
62].
The level of LPO in the body is controlled with the participation of antioxidant enzymes [
51], including the important role of GPX, which is involved in the reduction of organic peroxides formed during LPO. It was previously shown that treatment of tubers with Se NCs leads to a significant increase in GPX activity in tomato tissues 50 days after treatment [
63]. However, we did not reveal any changes in GPX activity in potatoes treated by any of the Se NCs tested in our experiments. Probably, stimulation of sprouting under the influence of Se/CAR NC is associated with other antioxidant enzymes. In addition, the results obtained on the content of DC and the levels of GPX activity indicate the absence of stress on potato tuber cells under the influence of NCs.
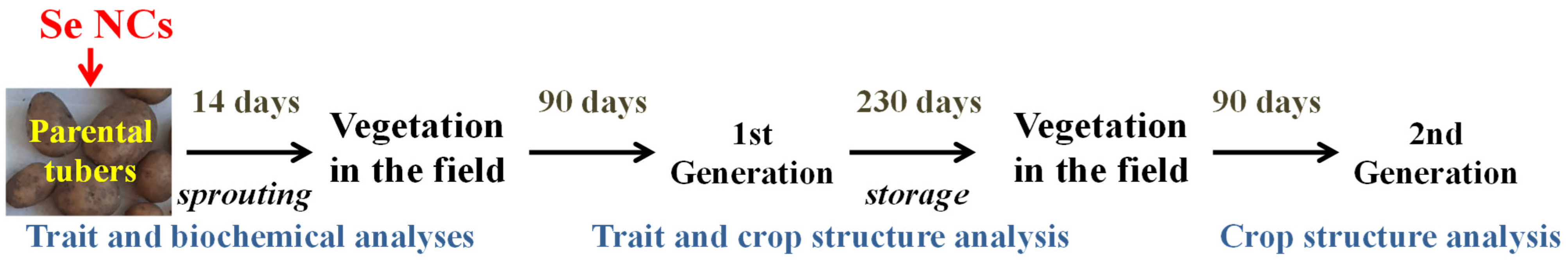
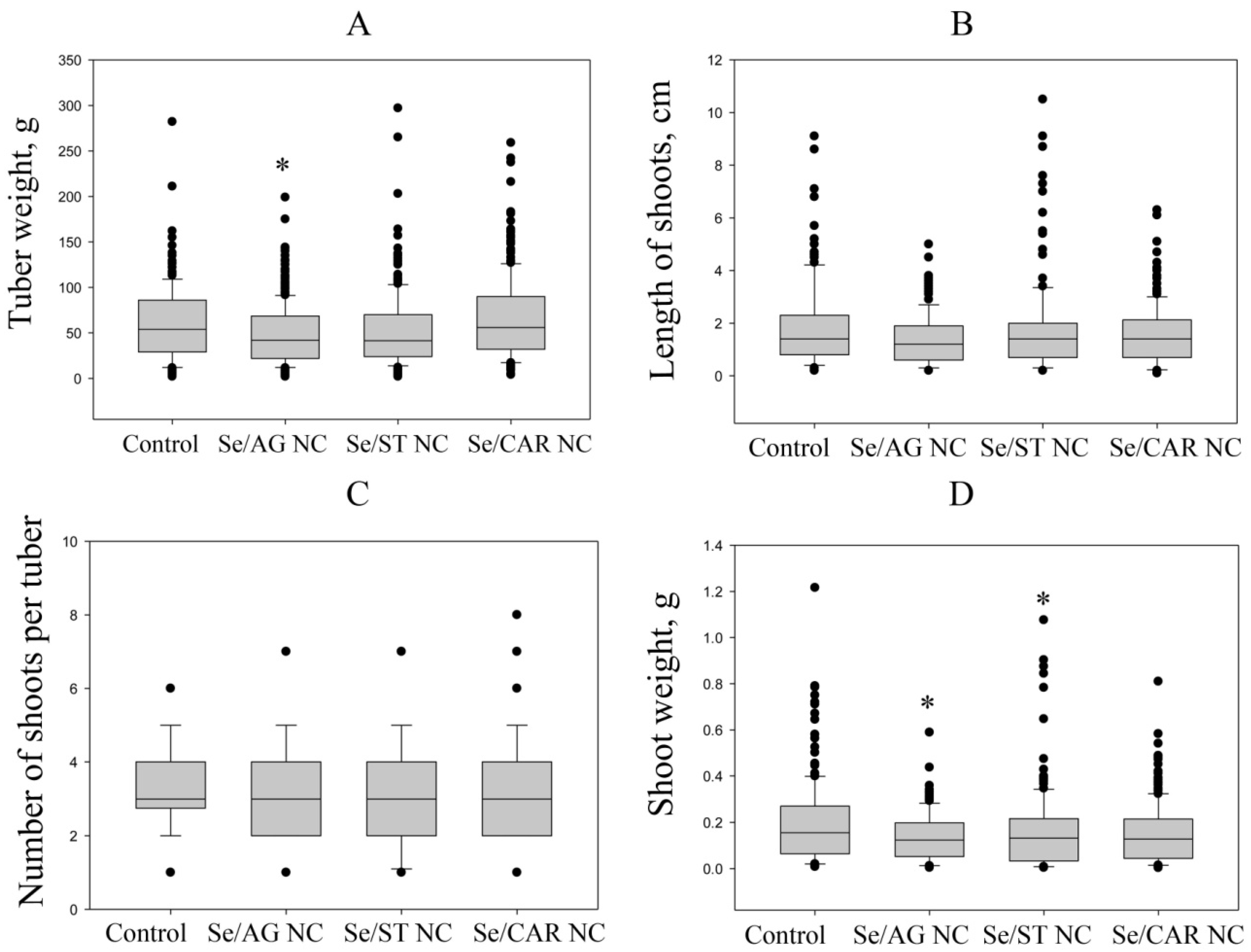
The data obtained in this study are summarized in
Table 6. It was found that only treatment with Se/CAR NC had a stimulating effect on the length and weight of shoots sprouted from the treated parental tubers. Tuber sprouting is always accompanied by activation of oxidative processes [
53].
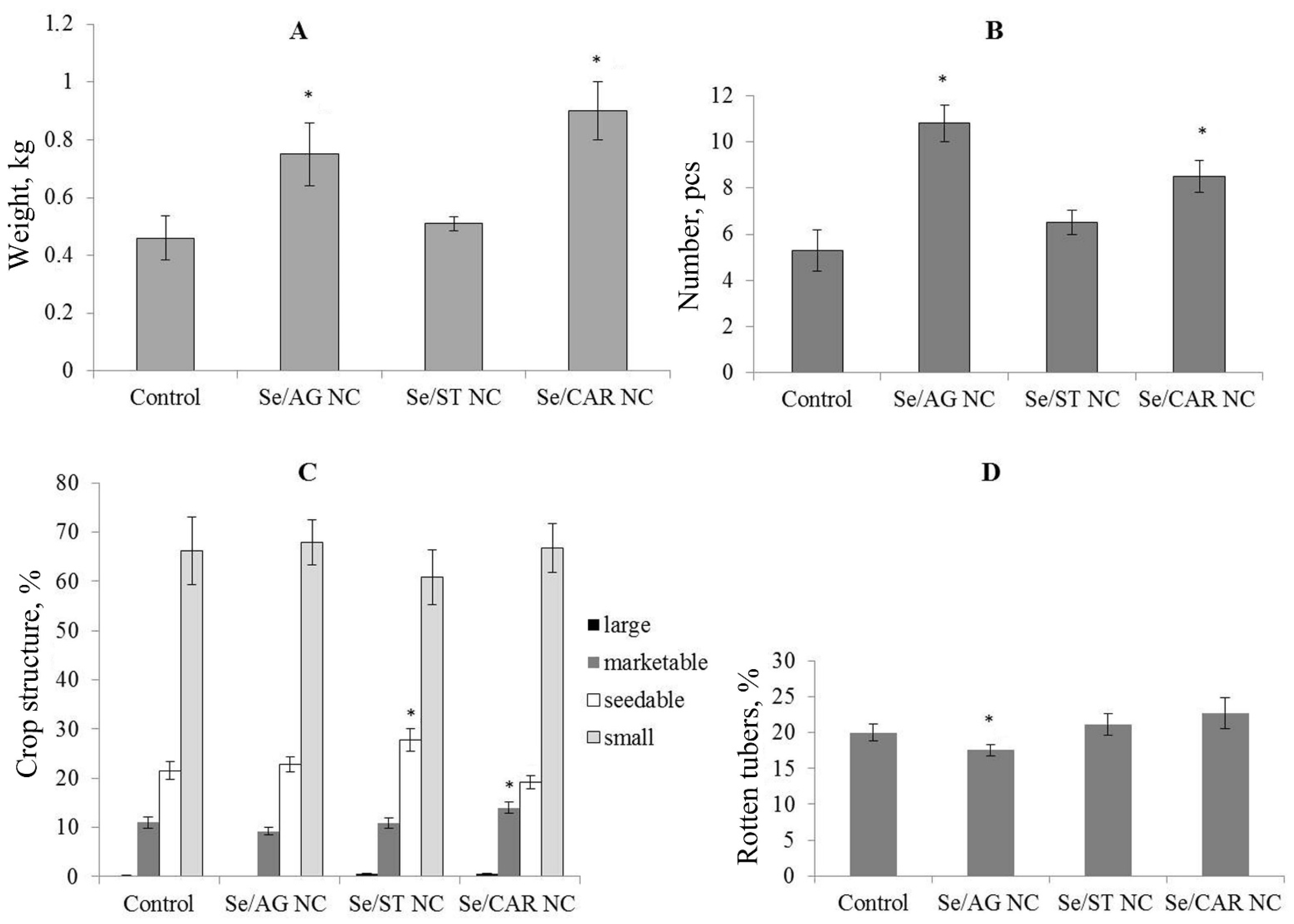
The results of the field experiment demonstrated that treatment of tubers with Se/AG and Se/CAR NCs stimulated the biomass of tubers in the first generation by increasing both the mass and number of tubers. It is in agreement with our previous studies, which showed that Se/AG NCs stimulated the growth and development of potato plants as well as their root formation in vitro [
34]. This effect may be due to the biological activity of not only Se NCs but also arabinogalactan, which stimulates the growth and development of plants [
64]. Experiments carried out on plants in vitro showed a stimulating effect of Se/CAR NC on biometric traits and a decrease in the negative effect of potato infection with a phytopathogenic bacterium [
37]. The different effects of Se NCs observed in our results can be associated with the nature of the matrix-different polysaccharides arabinogalactan [
65], carrageenan [
66], and starch [
67], which have rich individual biological activity. Most likely, the specific polysaccharide shell of antimicrobial Se NCs, which is trophic for microorganisms, can be captured with varying degrees of preference by phytopathogens (a kind of “Trojan horse” principle implemented for targeted microbial delivery of Se NCs) potentially both outside the cell [
68] and intracellularly [
69]. In the case of the observed positive effect of Se/CAR NC on the length and weight of seedlings after treatment with parental tubers, the weight and number of tubers in the 1st generation and an increase in the weight of tubers in the second generation, the growth-stimulating effect of sulfated carrageenan polysaccharide may also take place [
70,
71]. However, these hypotheses require further detailed verification. Selenium/AG and Se/ST NCs reduced premature tuber sprouting during storage. All three Se NCs increased the number of healthy tubers in the first generation tubers after their storage for 230 days. This effect can be explained by the fungicidal [
33] and antibacterial effects of Se NCs [
33,
34,
35,
36,
37]. The present data showed an increase in plant resistance to infections under the influence of treatments with Se NCs.
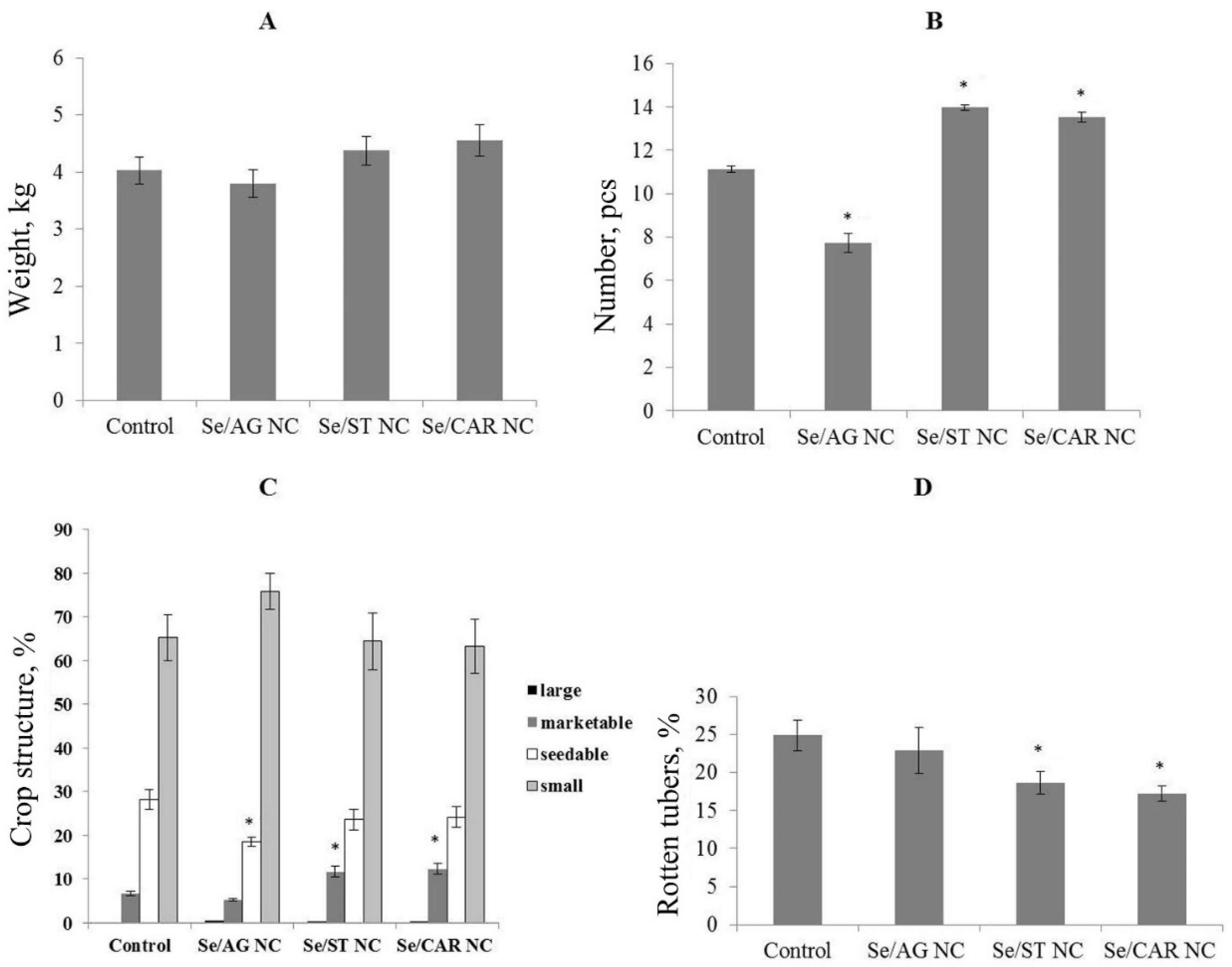
In the second generation, Se/AG NC did not affect the biomass and number of the tubers per plant (
Figure 6A,B). In addition, there was a significant decrease in the number of seed tubers in this treatment compared to the control (
Figure 6C). Selenium/ST and Se/CAR NCs increased the total biomass of tubers, the number of tubers per plant, and the number of marketable tubers in the crop structure (
Figure 6C;
Table 4). Most of which were healthy, likely indicating the stimulation of induced potato resistance under the influence of Se/ST and Se/CAR NCs, which had a prolonged effect that lasted even in the second generation (
Figure 6D). These transgenerational effects could be explained by stable epigenetic modifications caused by Se NCs and are worth to be studied further.
Over the past 5 years, many studies have been carried out to study the effect of Se NCs on the growth and development of plants, as well as their ability to resist stresses of various nature [
72,
73]. Most of the available literature data on the action of Se NCs indicated its positive effect on plants. For example, it has been shown that exogenous spraying by Se NCs increased the antioxidant potential of basil (
Ocimum basilicum L.) [
74] and enhanced the growth of tobacco (
Nicotiana tabacum L.) [
75] and peanut (
Arachis hypogaea L.) [
76]. The effect of selenium nanoparticles on seed germination was studied in
Hordeum vulgare L. [
77]. Nano-selenium dioxide increased the yield and intensity of plant growth and enhanced salt tolerance in
Phaseolus vulgaris L. growing in a field experiment on saline soils. It was suggested that an increase in the growth of higher plants treated by Se NCs occurs due to an increase in the productivity of photosynthesis [
78]. A change in the fatty acid profile of lipids in plant cells was also shown under the influence of Se NCs [
77]. It has been shown that spraying pear, grape, and peach plants during vegetation with a solution containing Se NPs increased the photosynthesis rate [
79]. In addition, it was found that Se NCs affect the activity of antioxidant enzymes in various plant organs, such as nitrate reductase in leaves and peroxidase in roots [
80].
It was also shown that Se NCs could function as stimulators of plant development, improving their antioxidant defense system and, consequently, their ability to tolerate stress [
75]. Se NCs affect cellular processes and, for example, regulate the activity of antioxidant enzymes and affect the photosynthetic apparatus. It was shown that Se NCs significantly reduced the content of heavy metals in rice grains grown on the polluted soil [
81]. Spraying plants with a solution of Se NCs improved the growth and increased the yield of rice, radish, and corn, and accelerated the growth of lettuce plants. It was found that Se NCs not only enhance the resistance of tomato plants to salt [
82] and biotic stress caused by a nematode (
Alternaria solani) but also increase their yield [
83]. The increased resistance of tomatoes to stress can be explained by the induction of some enzymes, such as SOD, ascorbate peroxidase, GP
X, phenylalanine ammonia lyase in leaves, and GP
X in fruits [
84]. In addition, the content of chlorophylls
a and
b was increased in the leaves, and the amount of vitamin C, glutathione, phenols, and flavonoids increased in the fruits [
85]. Salt stress tolerance and increased yields have been observed when strawberry plants were sprayed with
Fragaria ananassa and treated with Se NCs [
86]. The resulting effect was explained by a decrease in the level of LPO, an increase in the activity of antioxidant enzymes-SOD and peroxidase, as well as an increase in the content of proline in plant tissues [
85,
86]. In addition, an increase in the quality and nutritional properties of strawberries was noted due to an increase in the content of organic acids (for example, malic, citric, and succinic acids) and sugars (for example, glucose, fructose, and sucrose) in the berries of plants treated with Se NCs [
87].


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}