
Comparison of the Effect of Fertilization with Ash from Wood Chips on Bacterial Community in Podzolic and Chernozem Soils for the Cultivation of Winter Oilseed Rape: A Preliminary Study

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
- -
- Control—no K2O fertilization;
- -
- NPK K2O in mineral fertilizers (127 kg K2O ha−1);
- -
- 100 kg K2O ha−1 in ash (0.5 t ha−1 of ash in bulk weight);
- -
- 200 kg K2O ha−1 in ash (1.0 t ha−1 of ash in bulk weight);
- -
- 300 kg K2O ha−1 in ash (1.5 t ha−1 of ash in bulk weight);
- -
- 400 kg K2O ha−1 in ash (2.0 t ha−1 of ash in bulk weight);
- -
- 500 kg K2O ha−1 in ash (2.5 t ha−1 of ash in bulk weight).
2.1. Microbiological Analysis
2.1.1. Soil Preparation for Microbiological Analysis
2.1.2. Mass Spectrometry Identification of Isolates
3. Results and Discussion
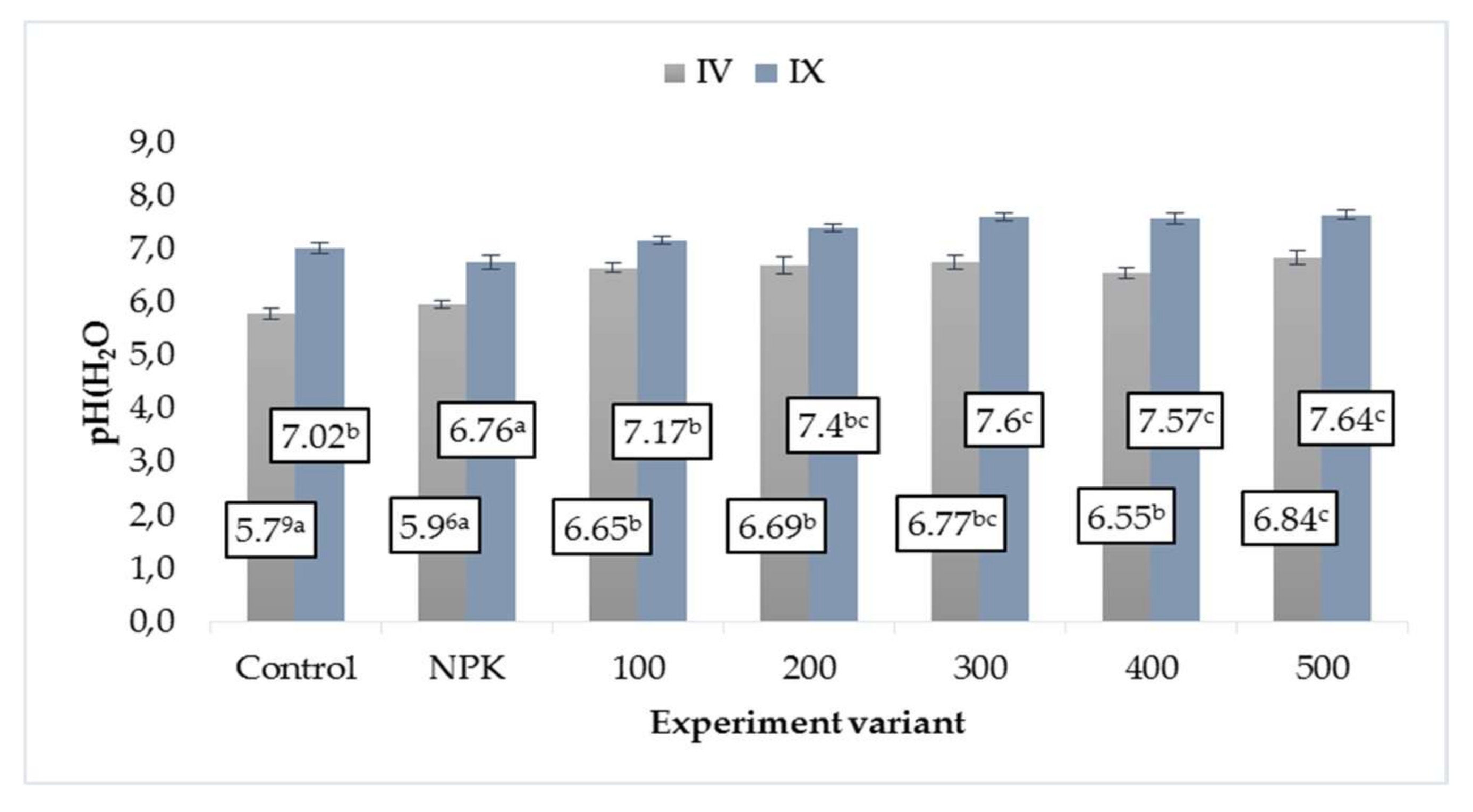
3.1. Soil pH
3.2. Salinity
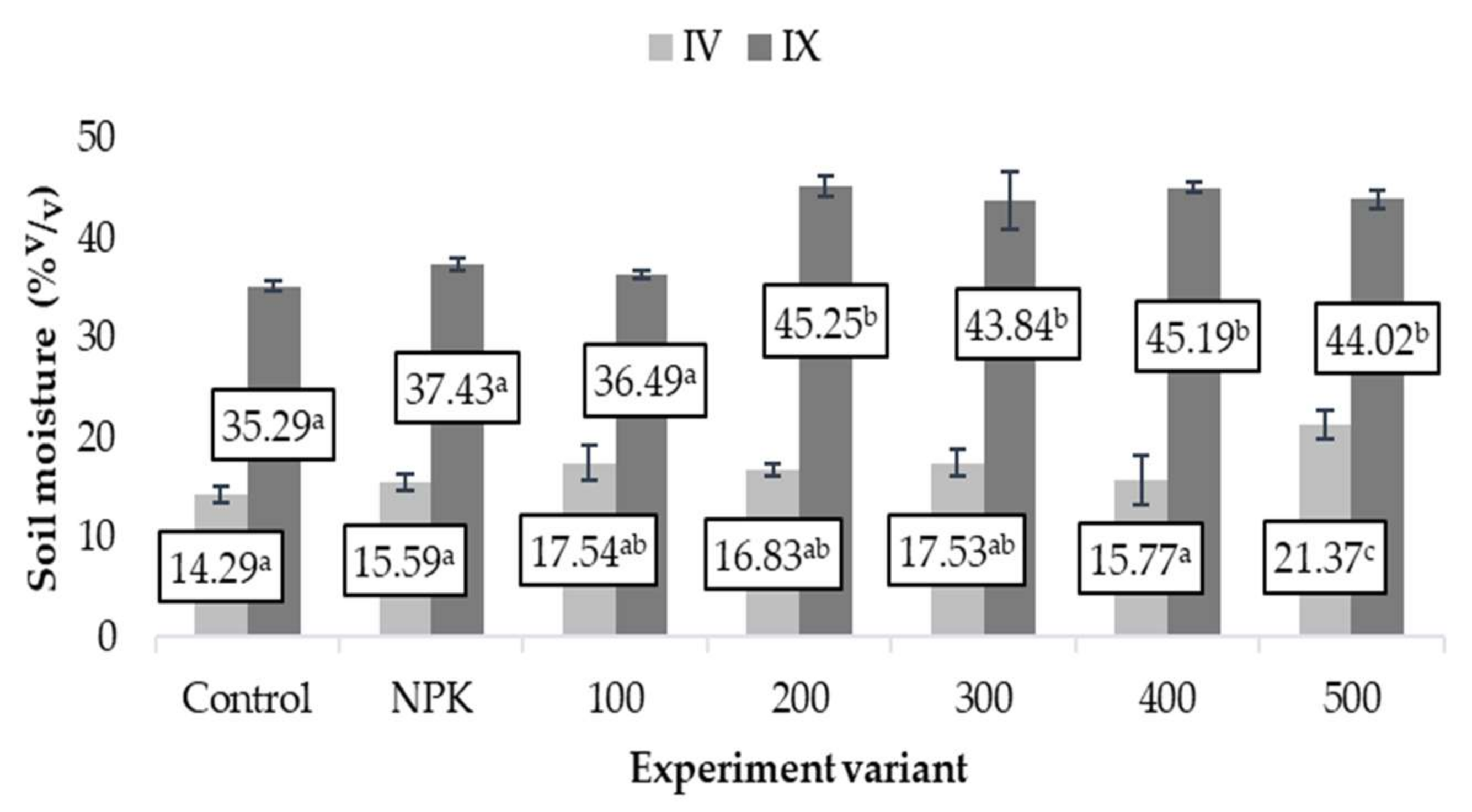
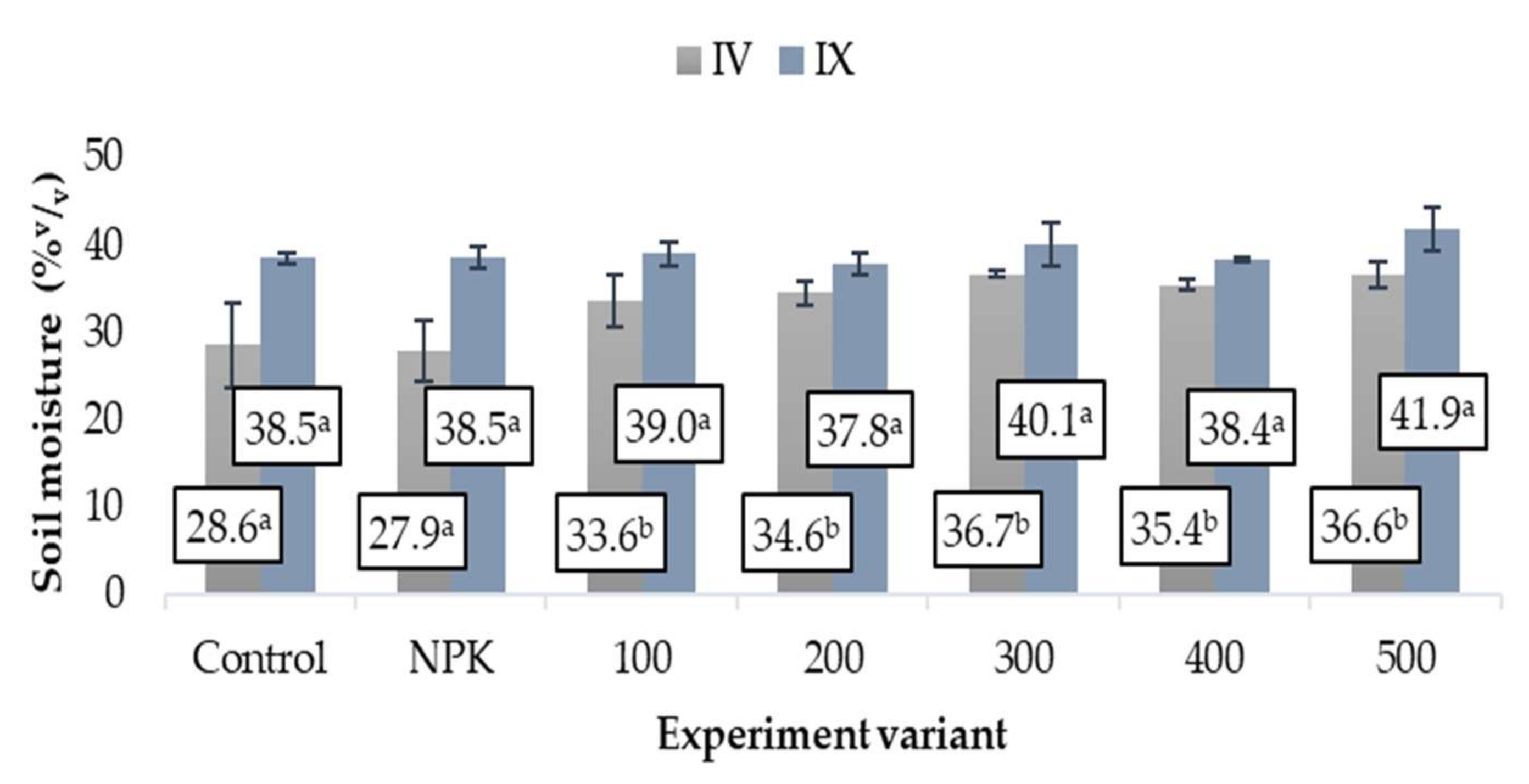
3.3. Soil Moisture
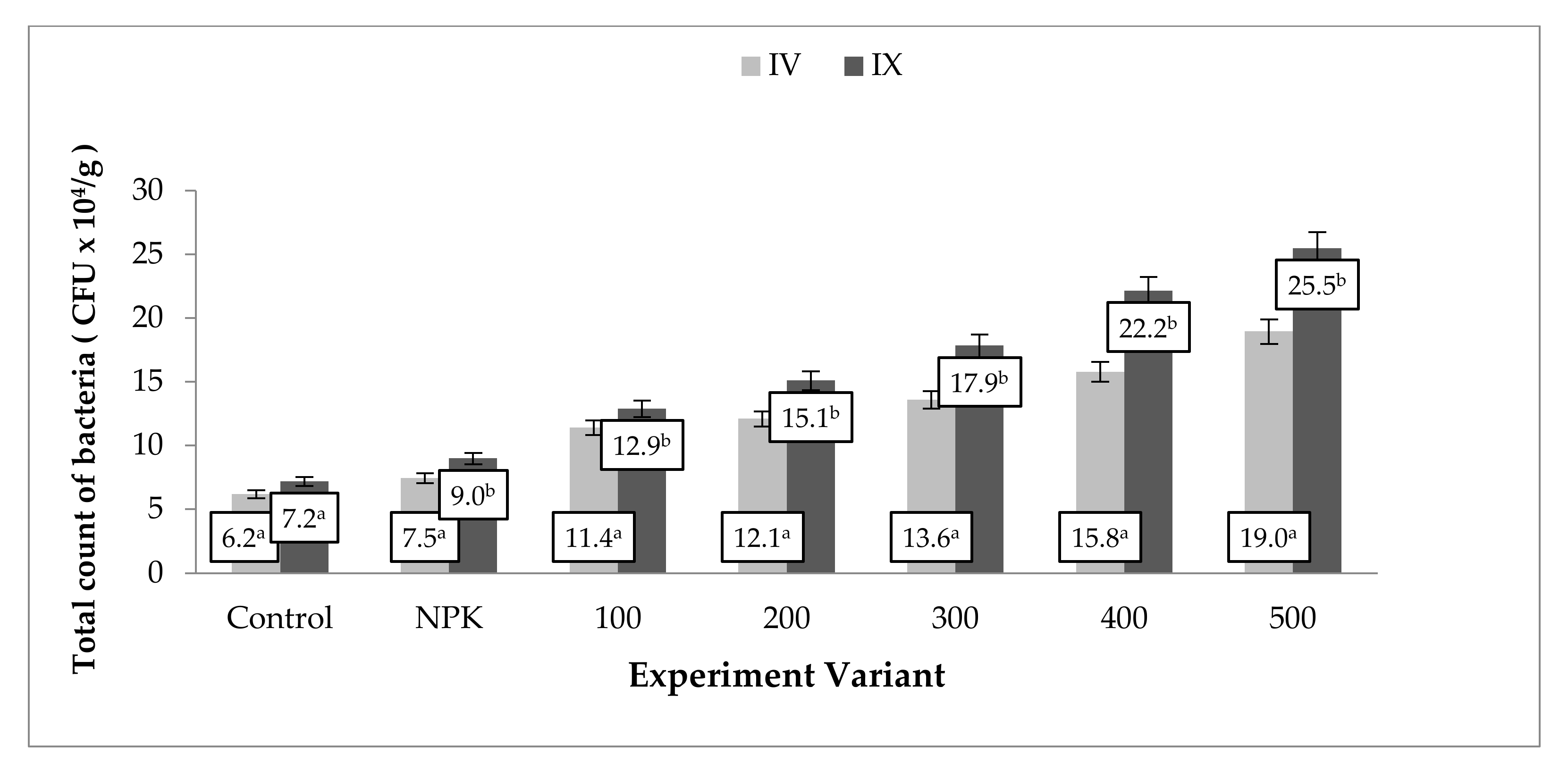
3.4. Microorganisms
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davison, J. Plant Beneficial Bacteria. Nat. Biotechnol. 1988, 6, 282–286. [Google Scholar] [CrossRef]
- Brierley, J.A. Use of microorganisms for mining metals. In Engineered Organisims in the Environment: Scientifc Issues; Halvorson, H.O., Pramer, D., Rogul, M., Eds.; ASM Press: Washington, DC, USA, 1985; pp. 141–146. [Google Scholar]
- Ehrlich, H.L. Geomicrobiology. Geomicrobiol. J. 1990, 2, 646. [Google Scholar]
- Middledrop, P.; Briglia, M.; Salkinoja-Salonen, M. Biodegradation of Pentachlorophenol in Natural Polluted Soil by Inoculated Rhodococcus chlorophenolicus. Microb. Ecol. 1990, 20, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Burd, G.I.; Dixon, D.G.; Glick, B.R. Plant Growth-Promoting Bacteria That Decrease Heavymetal Toxicity in Plants. Can. J. Microbiol. 2000, 46, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Chen, J.; Shim, H.; Bai, Z. New Advances in Plant Growth-Promoting Rhizobacteria for Bioremediation. Environ. Int. 2007, 33, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.; Usmani, S.; Singh, B.R.; Musarrat, J. Significance of Bacillus subtilis Strains SJ-101 as a Bioinoculant for Concurrent Plant Growth Promotion and Nickel Accumulation in Brassica juncea. Chemosphere 2008, 64, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil Beneficial Bacteria and Their Role in Plant Growth Promotion: A Review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Paul, E.A. Soil Microbiology, Ecology, and Biochemistry; Academic Press: Cambridge, MA, USA, 2014; p. 576. [Google Scholar]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going Back to the Roots: The Microbial Ecology of the Rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Wittebolle, L.; Marzorati, M.; Clement, L.; Balloi, A.; Daffonchio, D.; Heylen, K.; De Vos, P.; Verstraete, W.; Boon, N. Initial community evenness favours functionality under selective stress. Nature 2009, 458, 623–626. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial Diversity and Soil Functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Kunicki-Goldfinger, W.J.H. Życie Bakterii; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2008. [Google Scholar]
- Lee, S.-H.; Oh, B.-I.; Kim, J.-G. Effect of Various Amendments on Heavy Mineral Oil Bioremediation and Soil Microbial Activity. Bioresour. Technol. 2008, 99, 2578–2587. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.-R.; Doyle, A.; Holden, P.A.; Schimel, J.P. Drying and Rewetting Effects on C and N Mineralization and Microbial Activity in Surface and Subsurface California Grassland Soils. Soil Biol. Biochem. 2008, 40, 2281–2289. [Google Scholar] [CrossRef]
- Young, I.; Ritz, K. Tillage, Habitat Space Andfunction of Soil Microbes. Soil Tillage 2000, 53, 201–213. [Google Scholar] [CrossRef]
- Brzezińska, M. Aktywność Biologiczna Oraz Procesy Jej Towarzyszące w Glebach Organicznych Nawadnianych Oczyszczonymi Ściekami Miejskimi (Badania Polowe i Mode- Lowe). Acta Agrophys. 2006, 2, 131. [Google Scholar]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The Rhizosphere Microbiome: Significance of Plant Beneficial, Plant Pathogenic, and Human Pathogenic Microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Finlay, B.J.; Esteban, G.F. Oxygen Sensing Drives Predictable Migrations in a Microbial Community. Environ. Microbiol. 2009, 11, 81–85. [Google Scholar] [CrossRef]
- Gajda, A.M.; Czyż, E.A.; Ukalska-Jaruga, A. Comparison of the Effects of Different Crop Production Systems on Soil Physico-Chemical Properties and Microbial Activity under Winter Wheat. Agronomy 2020, 10, 1130. [Google Scholar] [CrossRef]
- Gajda, A.M.; Czyż, E.A.; Klimkowicz-Pawlas, A. Effects of Different Tillage Intensities on Physicochemical and Microbial Properties of a Eutric Fluvisol Soil. Agronomy 2021, 11, 1497. [Google Scholar] [CrossRef]
- Pett-Ridge, J.; Firestone, M.K. Redox Fluctuationstructures Microbial Communities in a Wet Tropical Soil. Appl. Environ. Microbiol. 2005, 71, 6998–7007. [Google Scholar] [CrossRef] [Green Version]
- Standing, D.; Killham, K. The Soil Environment. In Modern Soil Microbiology; Van Elsas, J.D., Jansson, J.K., Trevors, J.T., Eds.; CRC Press: New York, NY, USA, 2007; pp. 1–22. [Google Scholar]
- Dance, A. Soil Ecology: What Lies Beneath. Nature 2008, 455, 724–725. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.N.; Zhang, F.S.; Marschner, P.; Fan, F.L.; Gao, H.M.; Bao, X.G.; Sun, J.H.; Li, L. Effect of Intercropping on Crop Yield and Chemical and Microbiological Properties in Rhizosphere of Wheat (Triticum aestivum L.), Maize (Zea mays L.), and Faba Bean (Vicia faba L.). Biol. Fertil. Soils 2007, 43, 565–574. [Google Scholar] [CrossRef]
- Mauchline, T.H.; Malone, J.G. Life in Earth-the Root Microbiome to the Rescue? Curr. Curr. Opin. Microbiol. 2017, 37, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreasen, T.; Nielsen, J.T.; Voriskova, J.; Heise, J.; Rønn, R.; Kjøller, R.; Jacobsen, C.S. Wood Ash Induced PH Changes Strongly Affect Soil Bacterial Numbers and Community Composition. Front. Microbiol. 2017, 8, 1400. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Ver Loren van Themaat, E.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, S.; Frey, B. Soil Respiration and Microbial Properties in an Acid Forest Soil: Effects of Wood Ash. Soil Biol. Biochem. 2002, 34, 1727–1737. [Google Scholar] [CrossRef]
- Björk, R.G.; Ernfors, M.; Sikström, U.; Nilsson, M.B.; Andersson, M.X.; Rütting, T.; Klemedtsson, L. Contrasting Effects of Wood Ash Application on Microbial Community Structure, Biomass and Processes in Drained Forested Peatlands: Wood Ash Effects on Drained Forested Peatlands. FEMS Microbiol. Ecol. 2010, 73, 550–562. [Google Scholar] [CrossRef] [Green Version]
- Saarsalmi, A.; Smolander, A.; Kukkola, M.; Arola, M. Effect of Wood Ash and Nitrogen Fertilization on Soil Chemical Properties, Soil Microbial Processes, and Stand Growth in Two Coniferous Stands in Finland. Plant Soil 2010, 331, 329–340. [Google Scholar] [CrossRef]
- Moilanen, M.; Silfverberg, K.; Hokkanen, T.J. Effects of Wood-Ash on the Tree Growth, Vegetation and Substrate Quality of a Drained Mire: A Case Study. For. Ecol. Manag. 2002, 171, 321–338. [Google Scholar] [CrossRef]
- Bååth, E.; Arnebrant, K. Growth Rate and Response of Bacterial Communities to pH in Limed and Ash Treated Forest Soils. Soil Biol. Biochem. 1994, 26, 995–1001. [Google Scholar] [CrossRef]
- Fritze, H.; Perkiömäki, J.; Saarela, U.; Katainen, R.; Kim, T.P.; Karp, M.; Haimi, J.; Romantschuk, M. Effect of Cd-Containing Wood Ash on the Microflora of Coniferous Forest Humus. FEMS Microbiol. Ecol. 2000, 32, 43–51. [Google Scholar] [CrossRef]
- Ditzler, C.; Scheffe, K.; Monger, H.C.; Soil Science Division Staff. Soil Survey Manual; USDA Handbook 18; Government Printing Office: Washington, DC, USA, 2017; p. 603.
- Stanek-Tarkowska, J.; Szostek, M.; Rybak, M. Effect of Different Doses of Ash from Biomass Combustion on the Development of Diatom Assemblages on Podzolic Soil under Oilseed Rape Cultivation. Agronomy 2021, 11, 2422. [Google Scholar] [CrossRef]
- Cao, H.; Chen, R.; Wang, L.; Jiang, L.; Yang, F.; Zheng, S.; Wang, G.; Lin, X. Soil pH, Total Phosphorus, Climate and Distance Are the Major Factors Influencing Microbial Activity at a Regional Spatial Scale. Sci. Rep. 2016, 6, 25815. [Google Scholar] [CrossRef]
- Jala, S.; Goyal, D. Fly Ash as a Soil Ameliorant for Improving Crop Production—A Review. Bioresour. Technol. 2006, 97, 1136–1147. [Google Scholar] [CrossRef]
- Pandey, V.C.; Singh, N. Impact of Fly Ash Incorporation in Soil Systems. Agric. Ecosyst. Environ. 2010, 136, 16–27. [Google Scholar] [CrossRef]
- Marschner, C.P.H.; Yang, R.; Lieberei, D.E. Crowley Soil and Plant Specific Effects on Bacterial Community Composition in the Rhizosphere Soil Biol. Biol. Biochem. 2001, 33, 1437–1445. [Google Scholar] [CrossRef]
- Girvan, M.S.; Bullimore, J.; Pretty, J.N.; Osborn, A.M.; Ball, A.S. Soil Type Is the Primary Determinant of the Composition of the Total and Active Bacterial Communities in Arable Soils. Appl. Environ. Microbiol. 2003, 69, 1800–1809. [Google Scholar] [CrossRef] [Green Version]
- Miethling, R.; Wieland, G.; Backhaus, H.; Tebbe, C.C. Variation of Microbial Rhizosphere Communities in Response to Crop Species, Soil Origin, and Inoculation with Sinorhizobium meliloti L33. Microb. Ecol. 2000, 40, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Strejcek, M.; Smrhova, T.; Junkova, P.; Uhlik, O. Whole-Cell MALDI-TOF MS versus 16S RRNA Gene Analysis for Identification and Dereplication of Recurrent Bacterial Isolates. Front. Microbiol. 2018, 9, 1294. [Google Scholar] [CrossRef] [PubMed]
- Borowik, A.; Wyszkowska, J. Soil Moisture as a Factor Affecting the Microbiological and Biochemical Activity of Soil. Plant Soil Environ. 2016, 62, 250–255. [Google Scholar] [CrossRef]
- Perkiömäki, J.; Fritze, H. Does Simulated Acid Rain Increase the Leaching of Cadmium from Wood Ash to Toxic Levels to Coniferous Forest Humus Microbes? FEMS Microbiol. Ecol. 2003, 44, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Aronsson, K.A.; Ekelund, N.G.A. Biological Effects of Wood Ash Application to Forest and Aquatic Ecosystems. J. Environ. Qual. 2004, 33, 1595–1605. [Google Scholar] [CrossRef] [Green Version]
- Iovieno, P.; Bååth, E. Effect of Drying and Rewetting on Bacterial Growth Rates in Soil: Rewetting and Bacterial Growth in Soil. FEMS Microbiol. Ecol. 2008, 65, 400–407. [Google Scholar] [CrossRef] [Green Version]
- Demeyer, A.; Voundi Nkana, J.C.; Verloo, M.G. Characteristics of Wood Ash and Influence on Soil Properties and Nutrient Uptake: An Overview. Bioresour. Technol. 2001, 77, 287–295. [Google Scholar] [CrossRef]
- Dutta, S.; Podile, A.R. Plant Growth Promoting Rhizobacteria (PGPR): The Bugs to Debug the Root Zone. Crit. Rev. Microbiol. 2010, 36, 232–244. [Google Scholar] [CrossRef]
- Mayak, S.; Tirosh, T.; Glick, B.R. Effect of Wild-Type and Mutant Plant Growth-Promoting Rhizobacteria on the Rooting of Mung Bean Cuttings. J. Plant Growth Regul. 1999, 18, 49–53. [Google Scholar] [CrossRef]
- Glick, B.R. The Enhancement of Plant Growth by Free-Living Bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Babalola, O.O. Beneficial Bacteria of Agricultural Importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef]
- Vega-Celedón, P.; Bravo, G.; Velásquez, A.; Cid, F.P.; Valenzuela, M.; Ramírez, I.; Vasconez, I.-N.; Álvarez, I.; Jorquera, M.A.; Seeger, M. Microbial Diversity of Psychrotolerant Bacteria Isolated from Wild Flora of Andes Mountains and Patagonia of Chile towards the Selection of Plant Growth-Promoting Bacterial Consortia to Alleviate Cold Stress in Plants. Microorganisms 2021, 9, 538. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Pathma, J.; Kennedy, R.K.; Sakthivel, N. Mechanisms of Fluorescent Pseudomonads That Mediate Biological Control of Phytopathogens and Plant Growth Promotion of Crop Plants. In Bacteria in Agrobiology: Plant Growth Responses; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Jain, R.; Pandey, A. A Phenazine-1-Carboxylic Acid Producing Polyextremophilic Pseudomonas chlororaphis (MCC2693) Strain, Isolated from Mountain Ecosystem, Possesses Biocontrol and Plant Growth Promotion Abilities. Microbiol. Res. 2016, 190, 63–71. [Google Scholar] [CrossRef]
- Kumar, A.; Meena, R.; Meena, V.S.; Bisht, J.K.; Pattanayak, A. Towards the Stress Management and Environmental Sustainability. J. Clean. Prod. 2016, 137, 821–822. [Google Scholar] [CrossRef]
- Yasin, M.; Munir, I.; Faisal, M. Can Bacillus spp. enhance K+ uptake in crop species. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Meena, V.S., Maurya, B.R., Verma, J.P., Meena, R.S., Eds.; Springer: Delhi, India, 2016; pp. 163–170. [Google Scholar] [CrossRef]
- Srivastava, R. Antifungal Activity of P. fluorescens against Different Plant Pathogenic Fungi. EJEAFChe 2008, 7, 2789–2796. [Google Scholar]
- Davies, P.J. Plant Hormones: Biosynthesis, Signal Transduction, Action! Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Ligon, J.M.; Hill, D.S.; Hammer, P.E.; Torkewitz, N.R.; Hofmann, D.; Kempf, H.-J.; van Pée, K.-H. Natural Products with Antifungal Activity from Pseudomonas Biocontrol Bacteria. Pest Manag. Sci. 2000, 56, 688–695. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Vlami, M.; de Souza, J.T. Antibiotic Production by Bacterial Biocontrol Agents. Antonie Van Leeuwenhoek 2002, 81, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R.; Hashem, A.; Abd_Allah, E. Bacillus: A Biological Tool for Crop Improvement through Bio-Molecular Changes in Adverse Environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [PubMed]
- Glick, W.H.; Miller, C.C. Cardinal Making a Life in the Field of Organization Science. J. Organ. Behav. 2007, 28, 817–835. [Google Scholar] [CrossRef]
- Busse, H.-J. Review of the Taxonomy of the Genus Arthrobacter, Emendation of the Genus Arthrobacter sensu lato, Proposal to Reclassify Selected Species of the Genus Arthrobacter in the Novel Genera Glutamicibacter gen. nov., Paeniglutamicibacter gen. nov., Pseudoglutamicibacter gen. nov., Paenarthrobacter gen. nov. and Pseudarthrobacter gen. nov., and Emended Description of Arthrobacter roseus. Int. J. Syst. Evol. Microbiol. 2016, 66, 9–37. [Google Scholar] [CrossRef] [Green Version]
- Roy, P.; Kumar, A. Arthrobacter. In Beneficial Microbes in Agro-Ecology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–14. [Google Scholar]
- Krishnan, R.; Menon, R.R.; Tanaka, N.; Busse, H.-J.; Krishnamurthi, S.; Rameshkumar, N. Arthrobacter pokkalii sp nov, a Novel Plant Associated Actinobacterium with Plant Beneficial Properties, Isolated from Saline Tolerant Pokkali Rice, Kerala, India. PLoS ONE 2016, 11, e0150322. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Li, Y.; Zhu, X.; Cai, B. Isolation and Characterization of Atrazine Degrading Arthrobacter sp. AD26 and Use of This Strain in Bioremediation of Contaminated Soil. J. Environ. Sci. 2008, 20, 1226–1230. [Google Scholar] [CrossRef]
- Wang, Q.; Xie, S. Isolation and Characterization of a High-Efficiency Soil Atrazine-Degrading Arthrobacter sp. Strain. Int. Biodeterior. Biodegrad. 2012, 71, 61–66. [Google Scholar] [CrossRef]
- Xie, S.; Wan, R.; Wang, Z.; Wang, Q. Atrazine Biodegradation by Arthrobacter Strain DAT1: Effect of Glucose Supplementation and Change of the Soil Microbial Community. Environ. Sci. Pollut. Res. Int. 2013, 20, 4078–4084. [Google Scholar] [CrossRef]
- Parag, A.; Kanekar, P.P.; Dhakephalkar, P.K. Isolation and Characterization of Arthrobacter sp. Strain MCM B-436, an Atrazine-Degrading Bacterium, from Rhizospheric Soil. Int. Biodeterior. Biodegrad. 2007, 60, 273–278. [Google Scholar]
- Guo, Q.; Zhang, J.; Wan, R.; Xie, S. Impacts of Carbon Sources on Simazine Biodegradation by Arthrobacter Strain SD3-25 in Liquid Culture and Soil Microcosm. Int. Biodeterior. Biodegrad. 2014, 89, 1–6. [Google Scholar] [CrossRef]
- Xiao, W.; Ye, X.; Yang, X.; Zhu, Z.; Sun, C.; Zhang, Q.; Xu, P. Isolation and Characterization of Chromium (VI)-Reducing Bacillus sp. FY1 and Arthrobacter sp. WZ2 and Their Bioremediation Potential. Bioremed. J. 2017, 21, 100–108. [Google Scholar] [CrossRef]
- Singer, A.C.; Gilbert, E.S.; Luepromchai, E.; Crowley, D.E. Bioremediation of Polychlorinated Biphenylcontaminated Soil Using Carvone and Surfactant-Grown Bacteria. Appl. Microbiol. Biotechnol. 2000, 54, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, E.S.; Crowley, D.E. Plant Compounds That Induce Polychlorinated Biphenyl Biodegradation by Arthrobacter sp. Strain B1B. Appl. Environ. Microbiol. 1997, 63, 1933–1938. [Google Scholar] [CrossRef] [Green Version]
- Joshi, M.N.; Pandit, A.S.; Sharma, A.; Pandya, R.V.; Desai, S.M.; Saxena, A.K.; Bagatharia, S.B. Draft Genome Sequence of Arthrobacter crystallopoietes Strain BAB-32, Revealing Genes for Bioremediation. Genome Announc. 2013, 1, e00452-13. [Google Scholar] [CrossRef] [Green Version]
- Murugappan, R.M.; Benazir-Begun, S.; Usha, C.; Lok-Kirubahar, S.; Karthikeyan, M. Growth Promoting and Probiotic Potential of the Endophytic Bacterium Rhodococcus Globerulus Colonizing the Medicinal Plant Plectranthus amboinicus (Lour.) Spreng. Int. J. Curr. Res. Rev. 2017, 9, 7–13. [Google Scholar]
- McLeod, M.P.; Warren, R.L.; Hsiao, W.W.L.; Araki, N.; Myhre, M.; Fernandes, C.; Miyazawa, D.; Wong, W.; Lillquist, A.L.; Wang, D.; et al. The Complete Genome of Rhodococcus sp. RHA1 Provides Insights into a Catabolic Powerhouse. Proc. Natl. Acad. Sci. USA 2006, 103, 15582–15587. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fertilizer—Trade Name | Amount of Pure Component in 100 kg of the Fertilizer | Dose (kg/L per 1 ha) | Date of Application | |
|---|---|---|---|---|
| Fertilizer | Pure Component | |||
| Biomass combustion ash | 1.63% P (3.73 kg P), 19.4% K (23.37 kg K), 4.96% Mg (8.222 kg Mg) | Varied depending on the experimental variant | 30 August 2018 29 August 2019 25 August 2020 | |
| Monoammonium phosphate (MAP) NH4H2PO4 (12%N-NH4, 52% P2O5, 22.7% P) | 22.7 kg P | 150 | 34 | 30 August 2018 (all variants) 29 August 2019 (all variants) 25 August 2020 (all variants) |
| 12 kg N | 18 | |||
| Potassium salt (60%) | 60 kg K | 175 | 105 | 30 August 2018 (NPK variant only) 29 August 2019 (NPK variant only) 28 August 2020 (NPK variant only) |
| RSM ® 32% N (aqueous solution of urea-ammonium nitrate, density 1.32 kg/dcm3) | 42.2 kg N (32 × 1.32) | 150 | 63.3 | 4 March 2019 10 March 2020 15 March 2021 |
| pH H2O | EC µS·cm−1 | Ca (mg kg−1) | K (mg kg−1) | Na (mg kg−1) | P (mg kg−1) |
|---|---|---|---|---|---|
| 12.82 | 8.81 | 145.081 | 129.617 | 1452 | 9244 |
| Temperature in °C | Precipitation in mm | |||||||
|---|---|---|---|---|---|---|---|---|
| 2020 | ||||||||
| Month | Ten-Day Period | Mean | Ten-Day Period | Total | ||||
| I | II | III | I | II | III | |||
| I | 0.5 | 2.3 | 1.4 | 1.4 | 3.9 | 0.1 | 7.9 | 11.8 |
| II | 2.7 | 4.7 | 4.0 | 3.8 | 23.7 | 8.2 | 21.5 | 53.3 |
| III | 5.3 | 7.2 | 2.9 | 5.1 | 15.0 | 2.9 | 2.0 | 19.8 |
| IV | 7.9 | 8.5 | 11.2 | 9.2 | 0.0 | 4.7 | 5.3 | 10.0 |
| V | 11.1 | 11.1 | 11.7 | 11.3 | 25.3 | 24.4 | 33.6 | 83.3 |
| VI | 15.9 | 19.0 | 19.5 | 18.1 | 20.2 | 22.6 | 120.0 | 162.9 |
| VII | 19.9 | 17.1 | 19.3 | 18.8 | 10.2 | 8.5 | 0.2 | 18.9 |
| VIII | 20.2 | 19.5 | 20.1 | 19.9 | 0.2 | 0.1 | 7.0 | 7.3 |
| IX | 15.7 | 14.7 | 14.5 | 15.0 | 4.2 | 0.0 | 39.3 | 43.5 |
| X | 13.8 | 8.3 | 11.1 | 11.1 | 16.8 | 30.4 | 7.1 | 54.3 |
| XI | 12.9 | 8.5 | 10.6 | 10.6 | 17.5 | 29.8 | 8.1 | 55.4 |
| XII | 7.5 | 6.4 | 1.0 | 5.0 | 7.4 | 2.3 | 5.6 | 15.3 |
| Total | 535.8 | |||||||
| 2021 | ||||||||
| I | 2.2 | −4.4 | 1.2 | −0.3 | 14.8 | 8.8 | 25.2 | 48.8 |
| II | −2.3 | −5.7 | 4.4 | −1.2 | 31.3 | 14.9 | 1.9 | 48.1 |
| III | 2.2 | 1.9 | 5.4 | 3.2 | 7.2 | 1.8 | 8.5 | 17.5 |
| IV | 5.4 | 6.4 | 7.7 | 6.5 | 8.2 | 36.4 | 4.8 | 49.4 |
| V | 11.4 | 14.1 | 13.0 | 12.8 | 21.2 | 15.6 | 27.1 | 63.9 |
| VI | 15.7 | 18.6 | 22.2 | 18.8 | 9.0 | 6.8 | 31.5 | 47.3 |
| VII | 21.1 | 22.8 | 21.0 | 21.6 | 29.3 | 16.5 | 9.2 | 55.0 |
| VIII | 18.7 | 19.0 | 14.7 | 17.5 | 37.1 | 6.9 | 63.4 | 107.4 |
| IX | 13.3 | 14.0 | 11.9 | 13.1 | 5.6 | 61.6 | 18.6 | 85.8 |
| X | 10.1 | 7.8 | 9.5 | 9.1 | 0.9 | 1.1 | 0.5 | 2.5 |
| XI | 7.2 | 5.5 | 4.1 | 5.6 | 6.8 | 6.4 | 19.0 | 32.2 |
| XII | 0.2 | 1.2 | −3.7 | −0.8 | 7.6 | 8.5 | 13.5 | 29.6 |
| Total | 587.4 | |||||||
| Taxa | Podzolic Soil | Chernozem Soil | ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IV | IX | IV | IX | |||||||||||||||||||||||||
| Control | NPK | 100 | 200 | 300 | 400 | 500 | Control | NPK | 100 | 200 | 300 | 400 | 500 | Control | NPK | 100 | 200 | 300 | 400 | 500 | Control | NPK | 100 | 200 | 300 | 400 | 500 | |
| Bacillus cereus | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |||||||||
| Bacillus cytotoxicus | + | + | ||||||||||||||||||||||||||
| Bacillus megaterium | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | ||||||||
| Bacillus mycoides | + | + | + | + | + | + | + | + | + | + | + | + | + | + | ||||||||||||||
| Bacillus pseudomycoides | + | + | + | + | + | + | ||||||||||||||||||||||
| Bacillus simplex | + | + | + | + | ||||||||||||||||||||||||
| Bacillus thuringiensis | + | + | + | + | + | |||||||||||||||||||||||
| Bacillus weihenstephanensis | + | + | + | + | + | + | + | + | + | + | + | + | ||||||||||||||||
| Paenarthrobacter aurescens | + | + | + | + | + | |||||||||||||||||||||||
| Paenarthrobacter histidinolovorans | + | + | + | + | + | |||||||||||||||||||||||
| Paenarthrobacter ilicis | + | + | + | + | ||||||||||||||||||||||||
| Paenarthrobacter nicotinovorans | + | + | + | |||||||||||||||||||||||||
| Pseudarthrobacter chlorophenolicus | + | + | + | |||||||||||||||||||||||||
| Pseudarthrobacter oxydans | + | + | + | + | + | + | + | |||||||||||||||||||||
| Pseudarthrobacter polychromogenes | + | + | + | |||||||||||||||||||||||||
| Pseudomonas agarici | + | + | + | |||||||||||||||||||||||||
| Pseudomonas antarctica | + | + | + | + | ||||||||||||||||||||||||
| Pseudomonas azotoformans | + | + | ||||||||||||||||||||||||||
| Pseudomonas brassicacearum | + | + | + | + | + | + | + | + | + | + | + | |||||||||||||||||
| Pseudomonas brenneri | + | + | ||||||||||||||||||||||||||
| Pseudomonas cedrina ssp. cedrina | + | + | ||||||||||||||||||||||||||
| Pseudomonas chlororaphis | + | + | + | + | ||||||||||||||||||||||||
| Pseudomonas chlororaphis ssp. aurantiaca | + | + | + | + | + | + | + | + | + | + | + | + | ||||||||||||||||
| Pseudomonas chlororaphis ssp. chlororaphis | + | + | + | + | + | + | ||||||||||||||||||||||
| Pseudomonas corrugata | + | + | + | + | + | + | + | + | + | |||||||||||||||||||
| Pseudomonas extremorientalis | + | + | + | + | + | |||||||||||||||||||||||
| Pseudomonas fluorescens | + | + | + | |||||||||||||||||||||||||
| Pseudomonas frederiksbergensis | + | + | + | |||||||||||||||||||||||||
| Pseudomonas grimontii | + | |||||||||||||||||||||||||||
| Pseudomonas graminis | + | + | + | |||||||||||||||||||||||||
| Pseudomonas gessardii | + | + | + | + | + | + | + | |||||||||||||||||||||
| Pseudomonas jessenii | + | + | + | + | + | + | + | + | ||||||||||||||||||||
| Pseudomonas kilonensis | + | + | + | + | + | + | ||||||||||||||||||||||
| Pseudomonas libanensis | + | + | ||||||||||||||||||||||||||
| Pseudomonas migulae | + | + | + | + | + | + | + | |||||||||||||||||||||
| Pseudomonas oleovorans | + | + | + | |||||||||||||||||||||||||
| Pseudomonas poae | + | + | + | |||||||||||||||||||||||||
| Pseudomonas protegens | + | + | + | |||||||||||||||||||||||||
| Pseudomonas putida | + | + | + | + | + | + | + | |||||||||||||||||||||
| Pseudomonas rhodesiae | + | + | + | + | ||||||||||||||||||||||||
| Pseudomonas thivervalensis | + | + | + | + | + | + | + | + | + | + | ||||||||||||||||||
| Pseudomonas trivialis | + | + | ||||||||||||||||||||||||||
| Pseudomonas vancouverensis | + | + | + | + | + | + | ||||||||||||||||||||||
| Pseudomonas veronii | + | + | + | |||||||||||||||||||||||||
| Rhodococcus globerulus | + | + | + | |||||||||||||||||||||||||
| Total | 1 | 1 | 10 | 5 | 6 | 5 | 13 | 0 | 2 | 11 | 8 | 11 | 12 | 25 | 2 | 1 | 14 | 5 | 14 | 9 | 16 | 2 | 1 | 15 | 8 | 20 | 20 | 22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanek-Tarkowska, J.; Pastuszczak, M.; Szpunar-Krok, E.; Kačániová, M.; Kluz, M.I.; Czyż, E.A.; Pieniążek, R.; Skrobacz, K.; Pietrzyk, K. Comparison of the Effect of Fertilization with Ash from Wood Chips on Bacterial Community in Podzolic and Chernozem Soils for the Cultivation of Winter Oilseed Rape: A Preliminary Study. Agronomy 2022, 12, 576. https://doi.org/10.3390/agronomy12030576
Stanek-Tarkowska J, Pastuszczak M, Szpunar-Krok E, Kačániová M, Kluz MI, Czyż EA, Pieniążek R, Skrobacz K, Pietrzyk K. Comparison of the Effect of Fertilization with Ash from Wood Chips on Bacterial Community in Podzolic and Chernozem Soils for the Cultivation of Winter Oilseed Rape: A Preliminary Study. Agronomy. 2022; 12(3):576. https://doi.org/10.3390/agronomy12030576
Chicago/Turabian StyleStanek-Tarkowska, Jadwiga, Miłosz Pastuszczak, Ewa Szpunar-Krok, Miroslava Kačániová, Maciej Ireneusz Kluz, Ewa Antonina Czyż, Rafał Pieniążek, Karol Skrobacz, and Karol Pietrzyk. 2022. "Comparison of the Effect of Fertilization with Ash from Wood Chips on Bacterial Community in Podzolic and Chernozem Soils for the Cultivation of Winter Oilseed Rape: A Preliminary Study" Agronomy 12, no. 3: 576. https://doi.org/10.3390/agronomy12030576