
Mycorrhizal Fungi Inoculation Improves Capparis spinosa’s Yield, Nutrient Uptake and Photosynthetic Efficiency under Water Deficit

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Rhizospheric Soil Samples
2.3. Mycorrhizal Fungi Inoculum
2.4. Experimental Design and Treatments
2.5. Growth Measurements
2.6. Plant Mineral Analysis
2.7. Chlorophyll Fluorescence
2.8. Content of Photosynthetic Pigments
2.9. Symbiotic Development
2.10. Statistical Analysis
3. Results
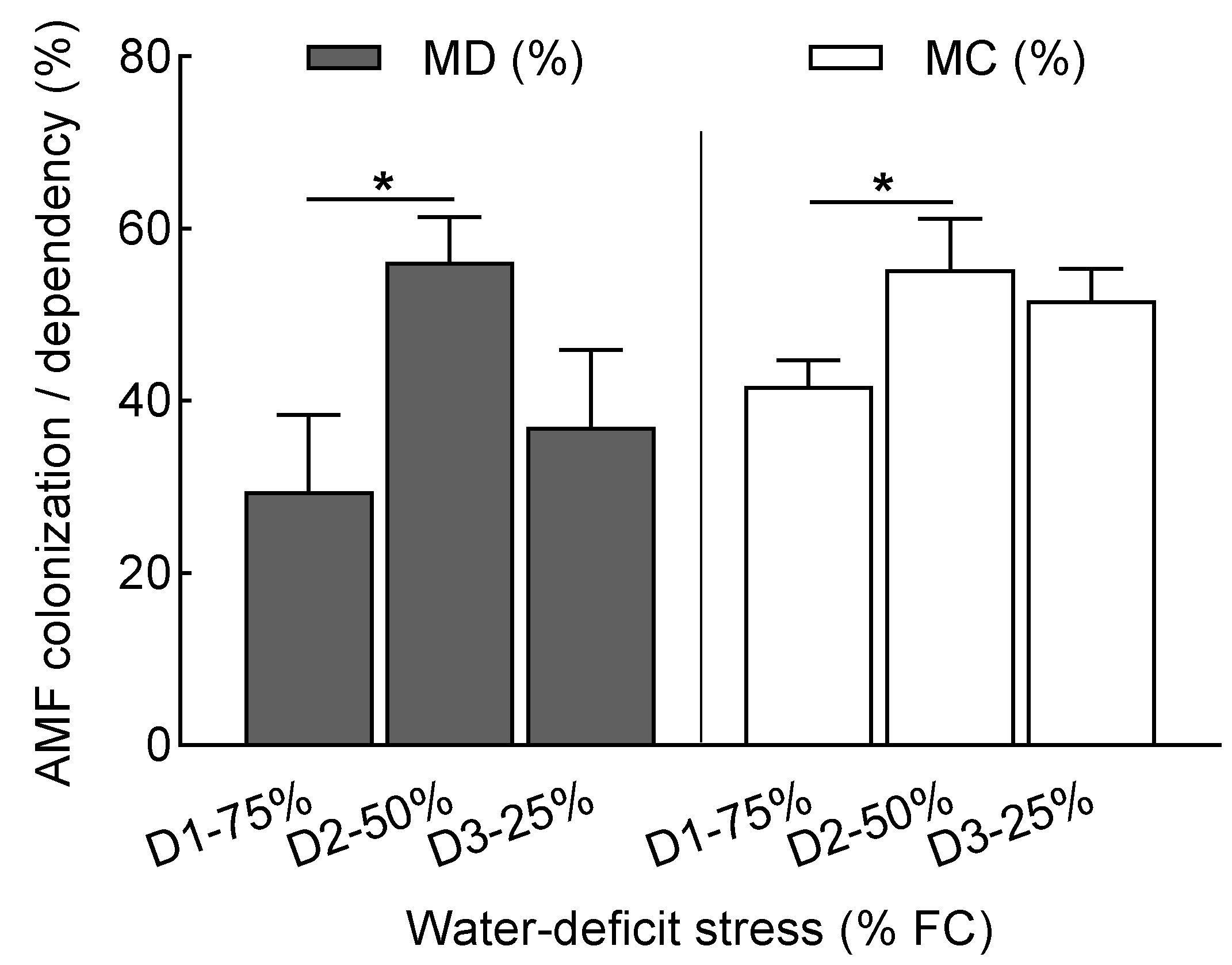
3.1. Mycorrhizal Colonization
3.2. Morphological Parameters
3.3. Biomass Production
3.4. Macro- and Micromineral Nutrient Contents
3.5. Chlorophyll Fluorescence
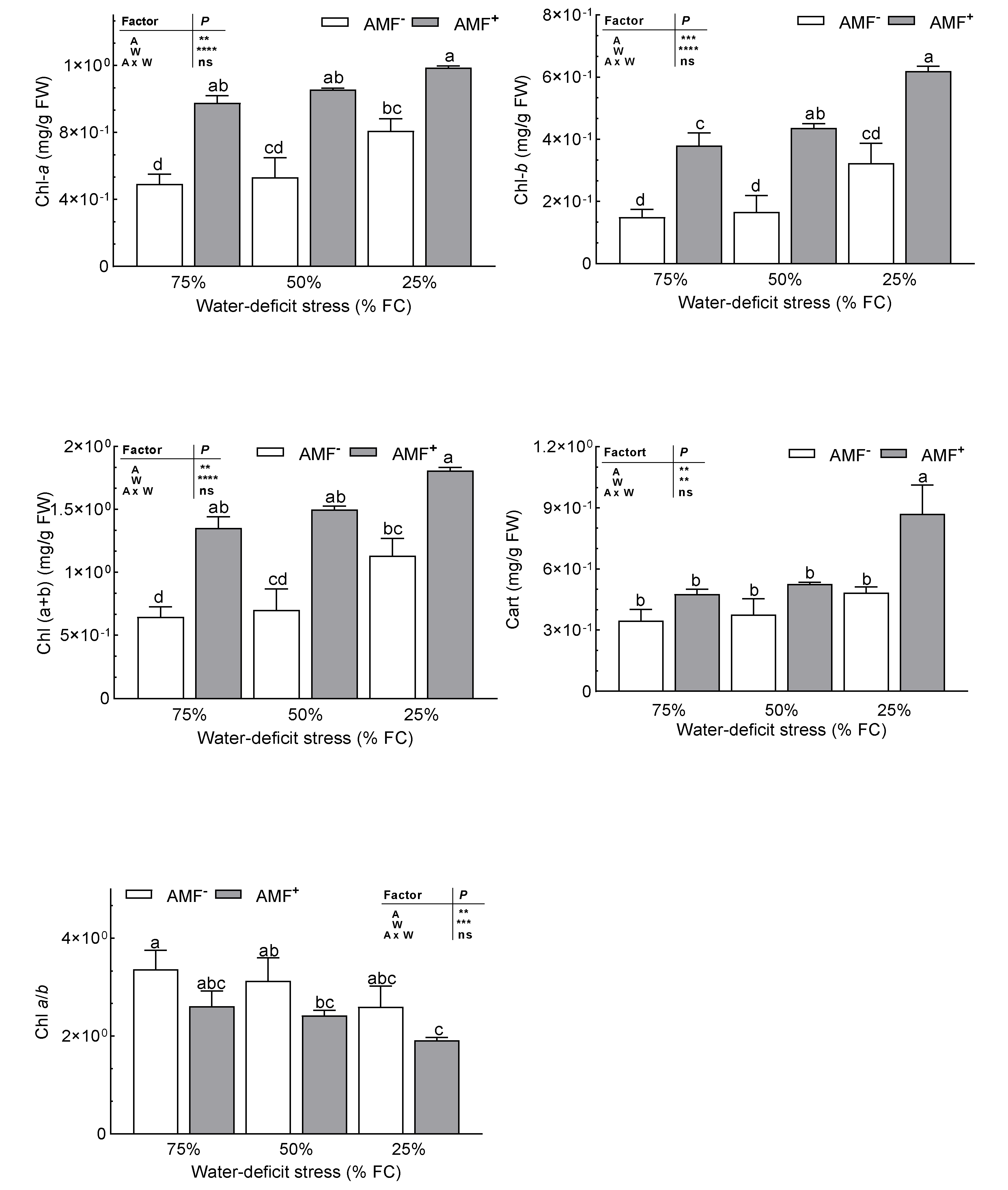
3.6. Photosynthetic Pigments
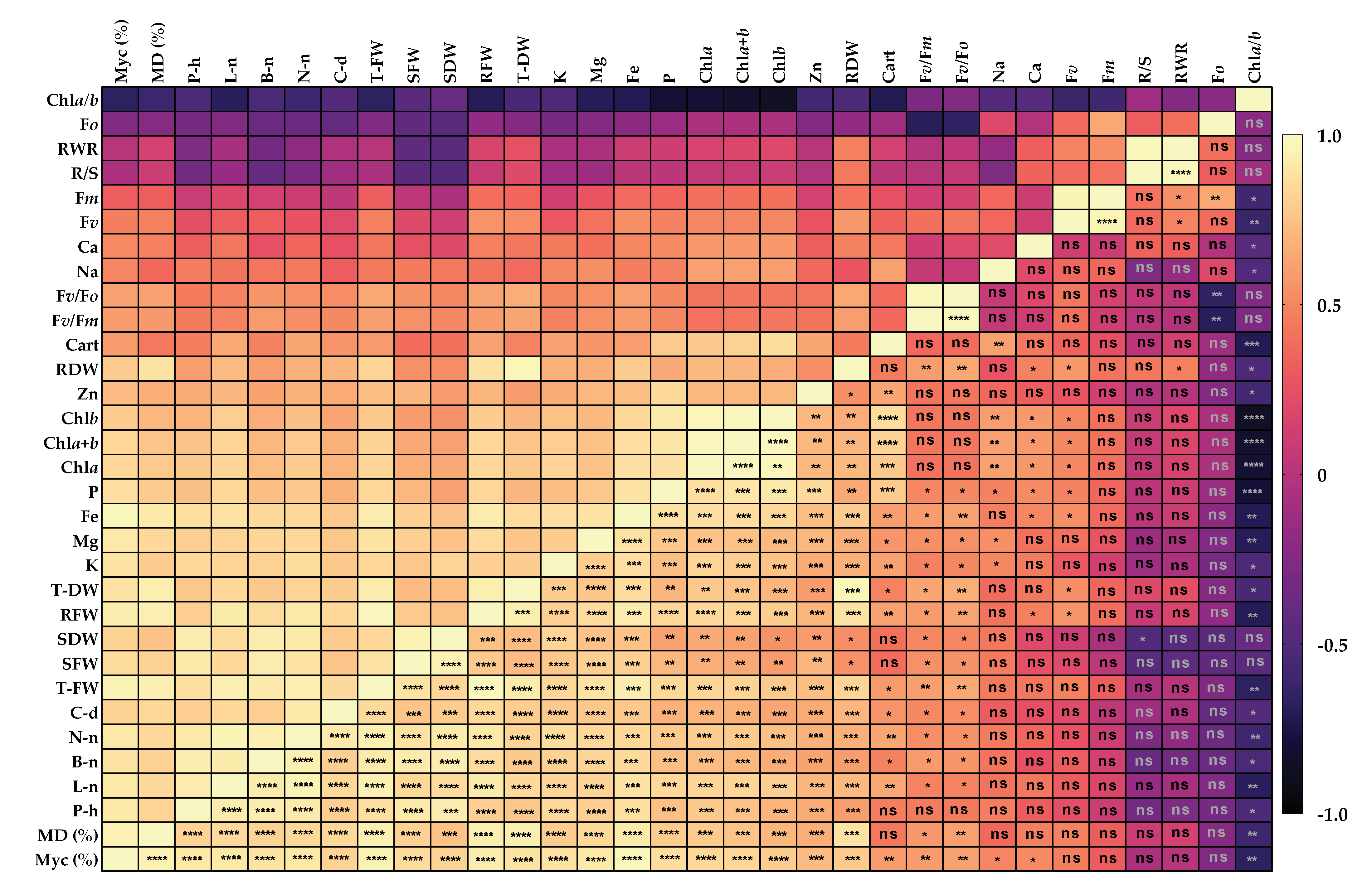
3.7. Relationship between Caper Seedlings Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chedraoui, S.; Abi-Rizk, A.; El-Beyrouthy, M.; Chalak, L.; Ouaini, N.; Rajjou, L. Capparis spinosa L. in A Systematic Review: A Xerophilous Species of Multi Values and Promising Potentialities for Agrosystems under the Threat of Global Warming. Front. Plant Sci. 2017, 8, 1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanucci, A.; Zengin, G.; Locatelli, M.; Macedonio, G.; Wang, C.-K.; Novellino, E.; Mahomoodally, M.; Mollica, A. Impact of different geographical locations on varying profile of bioactives and associated functionalities of caper (Capparis spinosa L.). Food Chem. Toxicol. 2018, 118, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Sozzi, G.O. Caper Bush: Botany and Horticulture. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons.: Hoboken, NJ, USA, 2001; Volume 27, pp. 125–188. ISBN 9780471387909. [Google Scholar]
- Saleem, H.; Khurshid, U.; Sarfraz, M.; Ahmad, I.; Alamri, A.; Anwar, S.; Alamri, A.S.; Locatelli, M.; Tartaglia, A.; Mahomoodally, M.F.; et al. Investigation into the biological properties, secondary metabolites composition, and toxicity of aerial and root parts of Capparis spinosa L.: An important medicinal food plant. Food Chem. Toxicol. 2021, 155, 112404. [Google Scholar] [CrossRef] [PubMed]
- Rajhi, I.; Hernandez-Ramos, F.; Abderrabba, M.; Ben Dhia, M.T.; Ayadi, S.; Labidi, J. Antioxidant, Antifungal and Phytochemical Investigations of Capparis spinosa L. Agriculture 2021, 11, 1025. [Google Scholar] [CrossRef]
- Tlili, N.; Nasri, N.; Saadaoui, E.; Khaldi, A.; Triki, S. Carotenoid and Tocopherol Composition of Leaves, Buds, and Flowers of Capparis spinosa Grown Wild in Tunisia. J. Agric. Food Chem. 2009, 57, 5381–5385. [Google Scholar] [CrossRef]
- La Bella, S.; Rossini, F.; Licata, M.; Virga, G.; Ruggeri, R.; Iacuzzi, N.; Leto, C.; Tuttolomondo, T. Four-Year Study on the Bio-Agronomic Response of Biotypes of Capparis spinosa L. on the Island of Linosa (Italy). Agriculture 2021, 11, 327. [Google Scholar] [CrossRef]
- Rhizopoulou, S. Development and Structure of Drought-tolerant Leaves of the Mediterranean Shrub Capparis spinosa L. Ann. Bot. 2003, 92, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Psaras, G.; Sofroniou, I. Wood Anatomy of Capparis Spinosa from an Ecological Perspective. IAWA J. 1999, 20, 419–429. [Google Scholar] [CrossRef]
- Levizou, E.; Drilias, P.; Kyparissis, A. Exceptional Photosynthetic Performance of Capparis spinosa L. Under Adverse Conditions of Mediterranean summer. Photosynthetica 2004, 42, 229–235. [Google Scholar] [CrossRef]
- Liu, J.; Ma, M. Anatomical Characteristics of Capparis spinosa L. in Tibet. Bull. Bot. Res. 2012, 32, 392–394. [Google Scholar] [CrossRef]
- Levizou, E.; Kyparissis, A. A novel pattern of leaf movement: The case of Capparis spinosa L. Tree Physiol. 2016, 36, 1117–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, W.; Ma, M.; Ma, Z.; Gao, R.; Guo, Y.; Jiang, W.; Liu, J.; Tian, L. Study of Photosynthetic Physiological Characteristics of Desert Plant Capparis spinosa L. J. Shihezi Univ. (Nat. Sci.) 2021, 30, 282–288. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-SHZN201203006.htm (accessed on 22 November 2021).
- Gan, L.; Zhang, C.; Yin, Y.; Lin, Z.; Huang, Y.; Xiang, J.; Fu, C.; Li, M. Anatomical adaptations of the xerophilous medicinal plant, Capparis spinosa, to drought conditions. Hortic. Environ. Biotechnol. 2013, 54, 156–161. [Google Scholar] [CrossRef]
- Mercati, F.; Fontana, I.; Gristina, A.S.; Martorana, A.; El Nagar, M.; De Michele, R.; Fici, S.; Carimi, F. Transcriptome analysis and codominant markers development in caper, a drought tolerant orphan crop with medicinal value. Sci. Rep. 2019, 9, 10411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inès, S.; Talbi, O.; Nasreddine, Y.; Rouached, A.; Gharred, J.; Jdey, A.; Hanana, M.; Abdelly, C. Drought tolerance traits in Medicago species: A review. Arid. Land Res. Manag. 2021, 1–17. [Google Scholar] [CrossRef]
- Sadeghi, H.; Rostami, L. Changes in biochemical characteristics and Na and K content of caper (Capparis spinosa L.) seedlings under water and salt stress. J. Agric. Rural Dev. Trop. Subtrop. 2017, 118, 199–206. [Google Scholar]
- Sadeghi, H.; Rostami, L. Evaluating the physiological and hormonal responses of caper plant (Capparis spinosa) subjected to drought and salinity. Desert (Biaban) 2016, 21, 49–55. [Google Scholar] [CrossRef]
- Bagherifard, A.; Hamidoghli, Y.; Biglouei, M.H.; Ghaedi, M. Effects of drought stress and superabsorbent polymer on morpho-physiological and biochemical traits of Caper (Capparis spinosa L.). Aust. J. Crop Sci. 2020, 14, 13–20. [Google Scholar] [CrossRef]
- Liu, J.; Ma, M. Effects of Arbuscular Mycorrhiza Fungi on Growth and Some Physiological Indices of Capparis spinosa. Acta Agric. Boreali-Occident. Sin. 2013, 22, 158–162. Available online: http://caod.oriprobe.com/articles/41173394/Effects_of_Arbuscular_Mycorrhiza_Fungi_on_Growth_and_Some_Physiologica.htm (accessed on 5 October 2021).
- Pugnaire, F.I.; Esteban, E. Nutritional adaptations of caper shrub (Capparis Ovata Desf.) to environmental stress. J. Plant Nutr. 1991, 14, 151–161. [Google Scholar] [CrossRef]
- Wang, W.; Shi, J.; Xie, Q.; Jiang, Y.; Yu, N.; Wang, E. Nutrient Exchange and Regulation in Arbuscular Mycorrhizal Symbiosis. Mol. Plant 2017, 10, 1147–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meddad-Hamza, A.; Beddiar, A.; Gollotte, A.; Lemoine, M.C.; Kuszala, C.; Gianinazzi, S. Arbuscular mycorrhizal fungi improve the growth of olive trees and their resistance to transplantation stress. Afr. J. Biotechnol. 2010, 9, 1159–1167. [Google Scholar] [CrossRef] [Green Version]
- Begum, N.; Ahanger, M.A.; Zhang, L. AMF inoculation and phosphorus supplementation alleviates drought induced growth and photosynthetic decline in Nicotiana tabacum by up-regulating antioxidant metabolism and osmolyte accumulation. Environ. Exp. Bot. 2020, 176, 104088. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; Xu, G.; Zhou, L.; Li, Y. Arbuscular mycorrhizal fungi improve the growth and drought tolerance of Zenia insignis seedlings under drought stress. New For. 2018, 50, 593–604. [Google Scholar] [CrossRef]
- Ouahmane, L.; Ndoye, I.; Morino, A.; Ferradous, A. Inoculation of Ceratonia siliqua L. with native arbuscular mycorrhizal fungi mixture improves seedling establishment under greenhouse conditions. Afr. J. Biotechnol. 2012, 11, 16422–16426. [Google Scholar] [CrossRef]
- Jadrane, I.; Al Feddy, M.N.; Dounas, H.; Kouisni, L.; Aziz, F.; Ouahmane, L. Inoculation with selected indigenous mycorrhizal complex improves Ceratonia siliqua’s growth and response to drought stress. Saudi J. Biol. Sci. 2020, 28, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [Green Version]
- Al-Karaki, G.N.; Clark, R.B. Growth, mineral acquisition, and water use by mycorrhizal wheat grown under water stress. J. Plant Nutr. 1998, 21, 263–276. [Google Scholar] [CrossRef]
- Asrar, A.-W.A.; Elhindi, K.M. Alleviation of drought stress of marigold (Tagetes erecta) plants by using arbuscular mycorrhizal fungi. Saudi J. Biol. Sci. 2011, 18, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Mickan, B.; Feng, G.; Chen, Y. Arbuscular mycorrhizal fungi improved plant growth and nutrient acquisition of desert ephemeral Plantago minuta under variable soil water conditions. J. Arid. Land 2014, 7, 414–420. [Google Scholar] [CrossRef]
- Mo, Y.; Wang, Y.; Yang, R.; Zheng, J.; Liu, C.; Li, H.; Ma, J.; Zhang, Y.; Wei, C.; Zhang, X. Regulation of Plant Growth, Photosynthesis, Antioxidation and Osmosis by an Arbuscular Mycorrhizal Fungus in Watermelon Seedlings under Well-Watered and Drought Conditions. Front. Plant Sci. 2016, 7, 644. [Google Scholar] [CrossRef] [Green Version]
- Basyal, B.; Emery, S.M. An arbuscular mycorrhizal fungus alters switchgrass growth, root architecture, and cell wall chemistry across a soil moisture gradient. Mycorrhiza 2020, 31, 251–258. [Google Scholar] [CrossRef]
- Youssef, S.; Lahcen, O.; Abdelaziz, A. Breaking seed dormancy in Cupressus atlantica Gaussen, an endemic and threatened coniferous tree in Morocco. J. For. Res. 2012, 23, 385–390. [Google Scholar] [CrossRef]
- Ouahmane, L.; Hafidi, M.; Thioulouse, J.; Ducousso, M.; Kisa, M.; Prin, Y.; Galiana, A.; Boumezzough, A.; Duponnois, R. Improvement of Cupressus atlantica Gaussen growth by inoculation with native arbuscular mycorrhizal fungi. J. Appl. Microbiol. 2007, 103, 683–690. [Google Scholar] [CrossRef]
- INVAM. International Culture Collection of Vesicular Arbuscular Mycorrhizal Fungi, Species Descriptions from Reference Cultures. 2019. Available online: http://fungi.invam.wvu.edu/the-fungi/species-descriptions.html (accessed on 10 November 2019).
- Uddin, A.H.; Khalid, R.S.; Alaama, M.; Abdualkader, A.M.; Kasmuri, A.; Abbas, S.A. Comparative study of three digestion methods for elemental analysis in traditional medicine products using atomic absorption spectrometry. J. Anal. Sci. Technol. 2016, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthodes d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae, Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, France, 1–5 July 1985; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA Press: Paris, France, 1986; pp. 217–221. [Google Scholar]
- Bethlenfalvay, G.J.; Brown, M.S.; Ames, R.N.; Thomas, R.S. Effects of drought on host and endophyte development in mycorrhizal soybeans in relation to water use and phosphate uptake. Physiol. Plant. 1988, 72, 565–571. [Google Scholar] [CrossRef]
- Baslam, M.; Goicoechea, N. Water deficit improved the capacity of arbuscular mycorrhizal fungi (AMF) for inducing the accumulation of antioxidant compounds in lettuce leaves. Mycorrhiza 2011, 22, 347–359. [Google Scholar] [CrossRef]
- Greipsson, S.; El-Mayas, H. Arbuscular Mycorrhizae of Leymus arenarius on Coastal Sands and Reclamation Sites in Iceland and Response to Inoculation. Restor. Ecol. 2000, 8, 144–150. [Google Scholar] [CrossRef]
- Caravaca, F.; Barea, J.M.; Palenzuela, J.; Figueroa, D.; Alguacil, M.D.M.; Roldan, A. Establishment of shrub species in a degraded semiarid site after inoculation with native or allochthonous arbuscular mycorrhizal fungi. Appl. Soil Ecol. 2003, 22, 103–111. [Google Scholar] [CrossRef]
- Pellegrino, E.; Bedini, S.; Avio, L.; Bonari, E.; Giovannetti, M. Field inoculation effectiveness of native and exotic arbuscular mycorrhizal fungi in a Mediterranean agricultural soil. Soil Biol. Biochem. 2011, 43, 367–376. [Google Scholar] [CrossRef]
- Ouledali, S.; Ennajeh, M.; Zrig, A.; Gianinazzi, S.; Khemira, H. Estimating the contribution of arbuscular mycorrhizal fungi to drought tolerance of potted olive trees (Olea europaea). Acta Physiol. Plant. 2018, 40, 81. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, P.; Zou, Y.-N.; Wu, Q.-S.; Kuča, K. Effects of mycorrhizal fungi on root-hair growth and hormone levels of taproot and lateral roots in trifoliate orange under drought stress. Arch. Agron. Soil Sci. 2019, 65, 1316–1330. [Google Scholar] [CrossRef]
- Leventis, G.; Tsiknia, M.; Feka, M.; Ladikou, E.; Papadakis, I.; Chatzipavlidis, I.; Papadopoulou, K.; Ehaliotis, C. Arbuscular mycorrhizal fungi enhance growth of tomato under normal and drought conditions, via different water regulation mechanisms. Rhizosphere 2021, 19, 100394. [Google Scholar] [CrossRef]
- Wu, M.; Yan, Y.; Wang, Y.; Mao, Q.; Fu, Y.; Peng, X.; Yang, Z.; Ren, J.; Liu, A.; Chen, S.; et al. Arbuscular mycorrhizal fungi for vegetable (VT) enhance resistance to Rhizoctonia solani in watermelon by alleviating oxidative stress. Biol. Control. 2020, 152, 104433. [Google Scholar] [CrossRef]
- Chen, W.; Meng, P.; Feng, H.; Wang, C. Effects of Arbuscular Mycorrhizal Fungi on Growth and Physiological Performance of Catalpa bungei C.A.Mey. under Drought Stress. Forests 2020, 11, 1117. [Google Scholar] [CrossRef]
- Abobatta, W.F. Drought adaptive mechanisms of plants—A review. Adv. Agric. Environ. Sci. 2019, 2, 62–65. [Google Scholar] [CrossRef]
- Rostami, L.; Sadeghi, H.; Hosseini, S. Response of Caper Plant to Drought and Different Ratios of Calcium and Sodium Chloride1. Planta Daninha 2016, 34, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Sozzi, G.O.; Peter, K.V.; Nirmal Babu, K.; Divakaran, M. Capers and caperberries. In Handbook of Herbs and Spices: Second Edition; Peter, K.V., Ed.; Woodhead: Abington Cambridge, UK, 2012; Volume 2, pp. 193–224. ISBN 9780857095688. [Google Scholar]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Zhang, W.; Zhang, B.; Xie, F. Effect of drought stress on sugar metabolism in leaves and roots of soybean seedlings. Plant Physiol. Biochem. 2019, 146, 1–12. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.; White, P.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- McCarthy, M.C.; Enquist, B. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation. Funct. Ecol. 2007, 21, 713–720. [Google Scholar] [CrossRef]
- Moustakas, M.; Bayçu, G.; Sperdouli, I.; Eroğlu, H.; Eleftheriou, E.P. Arbuscular Mycorrhizal Symbiosis Enhances Photosynthesis in the Medicinal Herb Salvia fruticosa by Improving Photosystem II Photochemistry. Plants 2020, 9, 962. [Google Scholar] [CrossRef] [PubMed]
- Lanfranco, L.; Fiorilli, V.; Gutjahr, C. Partner communication and role of nutrients in the arbuscular mycorrhizal symbiosis. New Phytol. 2018, 220, 1031–1046. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Smith, F.A. Fresh perspectives on the roles of arbuscular mycorrhizal fungi in plant nutrition and growth. Mycologia 2012, 104, 1–13. [Google Scholar] [CrossRef]
- Madrid-Delgado, G.; Orozco-Miranda, M.; Cruz-Osorio, M.; Hernández-Rodríguez, O.A.; Rodríguez-Heredia, R.; Roa-Huerta, M.; Avila-Quezada, G.D. Pathways of Phosphorus Absorption and Early Signaling between the Mycorrhizal Fungi and Plants. Phyton 2021, 90, 1321–1338. [Google Scholar] [CrossRef]
- Püschel, D.; Bitterlich, M.; Rydlová, J.; Jansa, J. Drought accentuates the role of mycorrhiza in phosphorus uptake. Soil Biol. Biochem. 2021, 157, 108243. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Schwarz, D.; Franken, P.; Colla, G. Plant responses to drought stress: From morphological to molecular features. In Plant Responses to Drought Stress: From Morphological to Molecular Features, 1st ed.; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 171–195. [Google Scholar]
- Kaba, J.S.; Abunyewa, A.A.; Kugbe, J.; Kwashie, G.K.; Ansah, E.O.; Andoh, H. Arbuscular mycorrhizal fungi and potassium fertilizer as plant biostimulants and alternative research for enhancing plants adaptation to drought stress: Opportunities for enhancing drought tolerance in cocoa (Theobroma cacao L.). Sustain. Environ. 2021, 7, 1963927. [Google Scholar] [CrossRef]
- Özcan, M. Mineral Composition of Different Parts of Capparis ovata Desf. var. canescens (Coss.) Heywood Growing Wild in Turkey. J. Med. Food 2005, 8, 405–407. [Google Scholar] [CrossRef]
- Amo, J.; Lara, A.; Martínez-Martínez, A.; Martínez, V.; Rubio, F.; Nieves-Cordones, M. The protein kinase SlCIPK23 boosts K + and Na + uptake in tomato plants. Plant Cell Environ. 2021, 44, 3819–3835. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. Plant of the Millennium, Caper (Capparis spinosa L.), chemical composition and medicinal uses. Bull. Natl. Res. Cent. 2021, 45, 131. [Google Scholar] [CrossRef]
- Aragüés, R.; Urdanoz, V.; Çetin, M.; Kirda, C.; Daghari, H.; Ltifi, W.; Lahlou, M.; Douaik, A. Soil salinity related to physical soil characteristics and irrigation management in four Mediterranean irrigation districts. Agric. Water Manag. 2011, 98, 959–966. [Google Scholar] [CrossRef] [Green Version]
- Ondrasek, G.; Rengel, Z. Environmental salinization processes: Detection, implications & solutions. Sci. Total. Environ. 2020, 754, 142432. [Google Scholar] [CrossRef]
- Wu, G.-Q.; Feng, R.-J.; Liang, N.; Yuan, H.-J.; Sun, W.-B. Sodium chloride stimulates growth and alleviates sorbitol-induced osmotic stress in sugar beet seedlings. Plant Growth Regul. 2014, 75, 307–316. [Google Scholar] [CrossRef]
- Huihui, Z.; Yuze, H.; Kaiwen, G.; Zisong, X.; Liu, S.; Wang, Q.; Wang, X.; Nan, X.; Wu, Y.; Guangyu, S. Na+ accumulation alleviates drought stress induced photosynthesis inhibition of PSII and PSI in leaves of Medicago sativa. J. Plant Interactions 2020, 16, 1–11. [Google Scholar] [CrossRef]
- Ma, Q.; Yue, L.-J.; Zhang, J.; Wu, G.-Q.; Bao, A.-K.; Wang, S.-M. Sodium chloride improves photosynthesis and water status in the succulent xerophyte Zygophyllum xanthoxylum. Tree Physiol. 2011, 32, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.-L.; Bao, A.-K.; Wang, S.-M.; Jin, H.-X. NaCl stimulates growth and alleviates drought stress in the salt-secreting xerophyte Reaumuria soongorica. Environ. Exp. Bot. 2019, 162, 433–443. [Google Scholar] [CrossRef]
- Guo, H.; Cui, Y.-N.; Pan, Y.-Q.; Wang, S.-M.; Bao, A.-K. Sodium chloride facilitates the secretohalophyte Atriplex canescens adaptation to drought stress. Plant Physiol. Biochem. 2020, 150, 99–108. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Xia, R.-X. Arbuscular mycorrhizal fungi influence growth, osmotic adjustment and photosynthesis of citrus under well-watered and water stress conditions. J. Plant Physiol. 2006, 163, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Kabir, A.H.; Debnath, T.; Das, U.; Prity, S.A.; Haque, A.; Rahman, M.; Parvez, S. Arbuscular mycorrhizal fungi alleviate Fe-deficiency symptoms in sunflower by increasing iron uptake and its availability along with antioxidant defense. Plant Physiol. Biochem. 2020, 150, 254–262. [Google Scholar] [CrossRef]
- Khushboo; Bhardwaj, K.; Singh, P.; Raina, M.; Sharma, V.; Kumar, D. Exogenous application of calcium chloride in wheat genotypes alleviates negative effect of drought stress by modulating antioxidant machinery and enhanced osmolyte accumulation. Vitr. Cell. Dev. Biol.-Anim. 2018, 54, 495–507. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Azcon, R. Effect of calcium application on the tolerance of mycorrhizal lettuce plants to polyethylene glycol-induced water stress. Symbiosis 1997, 23, 9–22. [Google Scholar]
- Saboor, A.; Ali, M.A.; Hussain, S.; El Enshasy, H.A.; Hussain, S.; Ahmed, N.; Gafur, A.; Sayyed, R.; Fahad, S.; Danish, S.; et al. Zinc nutrition and arbuscular mycorrhizal symbiosis effects on maize (Zea mays L.) growth and productivity. Saudi J. Biol. Sci. 2021, 28, 6339–6351. [Google Scholar] [CrossRef]
- Kim, S.A.; Guerinot, M.L. Mining iron: Iron uptake and transport in plants. FEBS Lett. 2007, 581, 2273–2280. [Google Scholar] [CrossRef] [Green Version]
- Al-Karaki, G.N.; Al-Raddad, A.; Clark, R.B. Water stress and mycorrhizal isolate effects on growth and nutrient acquisition of wheat. J. Plant Nutr. 1998, 21, 891–902. [Google Scholar] [CrossRef]
- Ohnishi, M.; Furutani, R.; Sohtome, T.; Suzuki, T.; Wada, S.; Tanaka, S.; Ifuku, K.; Ueno, D.; Miyake, C. Photosynthetic Parameters Show Specific Responses to Essential Mineral Deficiencies. Antioxidants 2021, 10, 996. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Xu, H.; Wang, Y.; Ye, X.; Lai, N.; Huang, Z.; Yang, L.; Li, Y.; Chen, L.-S.; Guo, J. Differences in morphological and physiological features of citrus seedlings are related to Mg transport from the parent to branch organs. BMC Plant Biol. 2021, 21, 239. [Google Scholar] [CrossRef] [PubMed]
- Ilhan, S.; Ozdemir, F.; Bor, M. Contribution of trehalose biosynthetic pathway to drought stress tolerance of Capparis ovata Desf. Plant Biol. 2014, 17, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Ozkur, O.; Ozdemir, F.; Bor, M.; Turkan, I. Physiochemical and antioxidant responses of the perennial xerophyte Capparis ovata Desf. to drought. Environ. Exp. Bot. 2009, 66, 487–492. [Google Scholar] [CrossRef]
- Augé, R.M.; Toler, H.D.; Saxton, A. Mycorrhizal Stimulation of Leaf Gas Exchange in Relation to Root Colonization, Shoot Size, Leaf Phosphorus and Nitrogen: A Quantitative Analysis of the Literature Using Meta-Regression. Front. Plant Sci. 2016, 7, 1084. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm plant−1) | Leaves (no. plant−1) | Branches (no. plant−1) | Nodes (no. plant−1) | Collar Diameter (mm plant−1) | |
|---|---|---|---|---|---|---|
| Water Stress (% FC) | Mycorrhizal Inoculation | |||||
| 75% FC | −AMF | 14.9 ± 0.7 b | 21 ± 2.0 b | 3 ± 1.0 b | 31.7 ± 5.0 b | 5.4 ± 0.4 bc |
| +AMF | 24.2 ± 0.3 a | 43 ± 5.0 a | 6 ± 0.0 a | 52.3 ± 5.1 a | 6.4 ± 0.7 ab | |
| 50% FC | −AMF | 13.7 ± 1.6 b | 18 ± 1.0 b | 1.3 ± 0.6 b | 25 ± 3.0 b | 5.6 ± 0.4 bc |
| +AMF | 22.7 ± 2.4 a | 49 ± 12.4 a | 6.3 ± 1.5 a | 58.7 ± 7.5 a | 7.0 ± 0.01 a | |
| 25% FC | −AMF | 13.4 ± 1.2 b | 21.7 ± 1.5 b | 1.3 ± 0.6 b | 27 ± 1.0 b | 5.1 ± 0.1 c |
| +AMF | 21.4 ± 0.6 a | 47.3 ± 7.2 a | 5.7 ± 0.6 a | 53.3 ± 12.5 a | 6.4 ± 0.7 ab | |
| Water stress (W) | ns | ns | ns | ns | ns | |
| AMF inoculation (A) | **** | **** | **** | **** | *** | |
| W × A | ns | ns | ns | ns | ns | |
| Treatments | SFW | RFW | SDW | RDW | TFW | TDW | Rdw/Sdw | RWR | |
|---|---|---|---|---|---|---|---|---|---|
| Water Stress (% FC) | Mycorrhizal Inoculation | (g plant−1) | (g g−1) | ||||||
| 75% FC | −AMF | 4.3 ± 0.3 b | 6.2 ± 0.8 c | 1.0 ± 0.2 bc | 1.6 ± 0.3 c | 10.4 ± 0.8 c | 2.6 ± 0.2 d | 1.6 ± 0.7 b | 0.60 ± 0.03 b |
| +AMF | 5.6 ± 0.4 a | 9.6 ±1.1 b | 1.5 ± 0.1 a | 2.1 ± 0.2 c | 15.2 ± 1.3 b | 3.6 ± 0.2 bc | 1.4 ± 0.2 b | 0.58 ± 0.04 b | |
| 50% FC | −AMF | 2.7 ± 0.3 c | 5.90 ± 0.4 c | 0.6 ± 0.1 c | 1.9 ± 0.4 c | 8.6 ±0.1 c | 2.6 ± 0.6 d | 3.0 ± 0.1 a | 0.75 ± 0.01 a |
| +AMF | 5.5 ± 0.2 a | 13.9 ± 0.6 a | 1.5 ± 0.3 a | 4.3 ± 0.4 a | 19.4 ± 0.7 a | 5.8 ± 0.3 a | 2.9 ±0.6 a | 0.74 ± 0.04 a | |
| 25% FC | −AMF | 3.1 ± 0.1 c | 6.9 ± 0.7 c | 0.7 ± 0.1 c | 2.1 ± 0.3 c | 10.0 ± 0.8 c | 2.8 ± 0.4 cd | 3.1 ± 0.2 a | 0.75 ± 0.02 a |
| +AMF | 5.1 ± 0.1 a | 12.2 ± 0.9 a | 1.3 ± 0.2 ab | 3.2 ± 0.2 b | 17.3 ± 1.0 ab | 4.5 ± 0.1 b | 2.6 ± 0.5 ab | 0.71 ± 0.04 a | |
| Water stress (W) | **** | ** | * | **** | ns | *** | *** | **** | |
| AMF inoculation (A) | **** | **** | **** | **** | **** | **** | ns | ns | |
| W × A | ** | *** | ns | *** | *** | *** | ns | ns | |
| Treatments | P | K+ | Na+ | Ca++ | Mg++ | Fe++ | Zn++ | |
|---|---|---|---|---|---|---|---|---|
| Water Stress (% FC) | Mycorrhizal Inoculation | (mg.g−1) | (mg.g−1) | (mg.g−1) | (mg.g−1) | (mg.g−1) | (mg.g−1) | (mg.g−1) |
| 75% FC | −AMF | 1.4 ± 0.1 c | 11.1 ± 1.3 c | 27.5 ± 1.3 a | 11.6 ± 1.5 a | 3.4 ± 0.7 b | 0.8 ± 0.1 c | 0.05 ± 0.02 b |
| +AMF | 2.0 ± 0.3 b | 15.9 ± 0.9 ab | 29.5 ± 1.3 a | 13.4 ± 2.0 a | 4.6 ± 0.4 a | 1.6 ± 0.1 b | 0.07 ± 0.02 ab | |
| 50% FC | −AMF | 1.4 ± 0.1 c | 11.3 ± 2.2 c | 27.0 ± 3.4 a | 12.1 ± 1.8 a | 3.4 ± 0.4 b | 0.9 ± 0.1 c | 0.04 ± 0.01 b |
| +AMF | 2.1 ± 0.1 b | 16.5 ± 1.4 a | 28.3 ± 1.6 a | 14.4 ± 2.3 a | 4.9 ± 0.3 a | 1.8 ± 0.1 a | 0.07 ± 0.01 ab | |
| 25%FC | −AMF | 1.6 ± 0.1 c | 11.8 ± 1.6 bc | 28.3 ± 1.6 a | 13.9 ± 1.7 a | 3.2 ± 0.2 b | 0.9 ± 0.1 c | 0.05 ± 0.01 ab |
| +AMF | 2.7 ± 0.1 a | 16.6 ± 1.9 a | 30.9 ± 0.2 a | 14.9 ± 0.6 a | 4.9 ± 0.2 a | 1.9 ± 0.1 a | 0.08 ± 0.02 a | |
| Water stress (W) | *** | ns | ns | ns | ns | * | ns | |
| AMF inoculation (A) | **** | **** | ns | ns | **** | **** | ** | |
| W × A | * | ns | ns | ns | ns | * | ns | |
| Treatments | Fo | Fv | Fm | Fv/Fm | Fv/Fo | |
|---|---|---|---|---|---|---|
| Water Stress (% FC) | Mycorrhizal Inoculation | |||||
| 75% | −AMF | 34.7 ± 10 b | 119 ± 29.5 a | 153.7 ± 39 a | 0.78 ± 0.02 abc | 3.5 ± 0.3 ab |
| +AMF | 43.7 ± 4.2 ab | 129.3 ± 5.9 a | 173 ± 9.5 a | 0.75 ± 0.01 abc | 3.0 ± 0.2 ab | |
| 50% | −AMF | 48 ± 7.9 ab | 132 ± 19.9 a | 180 ± 27.9 a | 0.73 ± 0.01 bc | 2.8 ± 0.1 b |
| +AMF | 40.7 ± 2.1 ab | 162.3 ± 22 a | 203 ± 23.1 a | 0.80 ± 0.02 a | 4.0 ± 0.6 a | |
| 25% | −AMF | 56.7 ± 9.5 a | 147.7 ± 10 a | 204.3 ± 6.0 a | 0.72 ± 0.05 c | 2.7 ± 0.6 b |
| +AMF | 42.3 ± 4.5 ab | 161 ± 14.2 a | 203.3 ± 18 a | 0.79 ± 0.01 ab | 3.8 ± 0.2 a | |
| Water stress (W) | ns | * | * | ns | ns | |
| AMF inoculation (A) | ns | ns | ns | ** | ** | |
| W × A | * | ns | ns | ** | ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouskout, M.; Bourhia, M.; Al Feddy, M.N.; Dounas, H.; Salamatullah, A.M.; Soufan, W.; Nafidi, H.-A.; Ouahmane, L. Mycorrhizal Fungi Inoculation Improves Capparis spinosa’s Yield, Nutrient Uptake and Photosynthetic Efficiency under Water Deficit. Agronomy 2022, 12, 149. https://doi.org/10.3390/agronomy12010149
Bouskout M, Bourhia M, Al Feddy MN, Dounas H, Salamatullah AM, Soufan W, Nafidi H-A, Ouahmane L. Mycorrhizal Fungi Inoculation Improves Capparis spinosa’s Yield, Nutrient Uptake and Photosynthetic Efficiency under Water Deficit. Agronomy. 2022; 12(1):149. https://doi.org/10.3390/agronomy12010149
Chicago/Turabian StyleBouskout, Mohammed, Mohammed Bourhia, Mohamed Najib Al Feddy, Hanane Dounas, Ahmad Mohammad Salamatullah, Walid Soufan, Hiba-Allah Nafidi, and Lahcen Ouahmane. 2022. "Mycorrhizal Fungi Inoculation Improves Capparis spinosa’s Yield, Nutrient Uptake and Photosynthetic Efficiency under Water Deficit" Agronomy 12, no. 1: 149. https://doi.org/10.3390/agronomy12010149