
How Does Nitrogen Application Rate Affect Plant Functional Traits and Crop Growth Rate of Perennial Ryegrass-Dominated Permanent Pastures?

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Conditions
2.2. Experimental Setup
2.3. Herbage Production, Nitrogen Content and Nitrogen Uptake
2.4. Plant Functional Trait Measurements
2.5. Statistical Analysis
3. Results
3.1. Climate and Weather
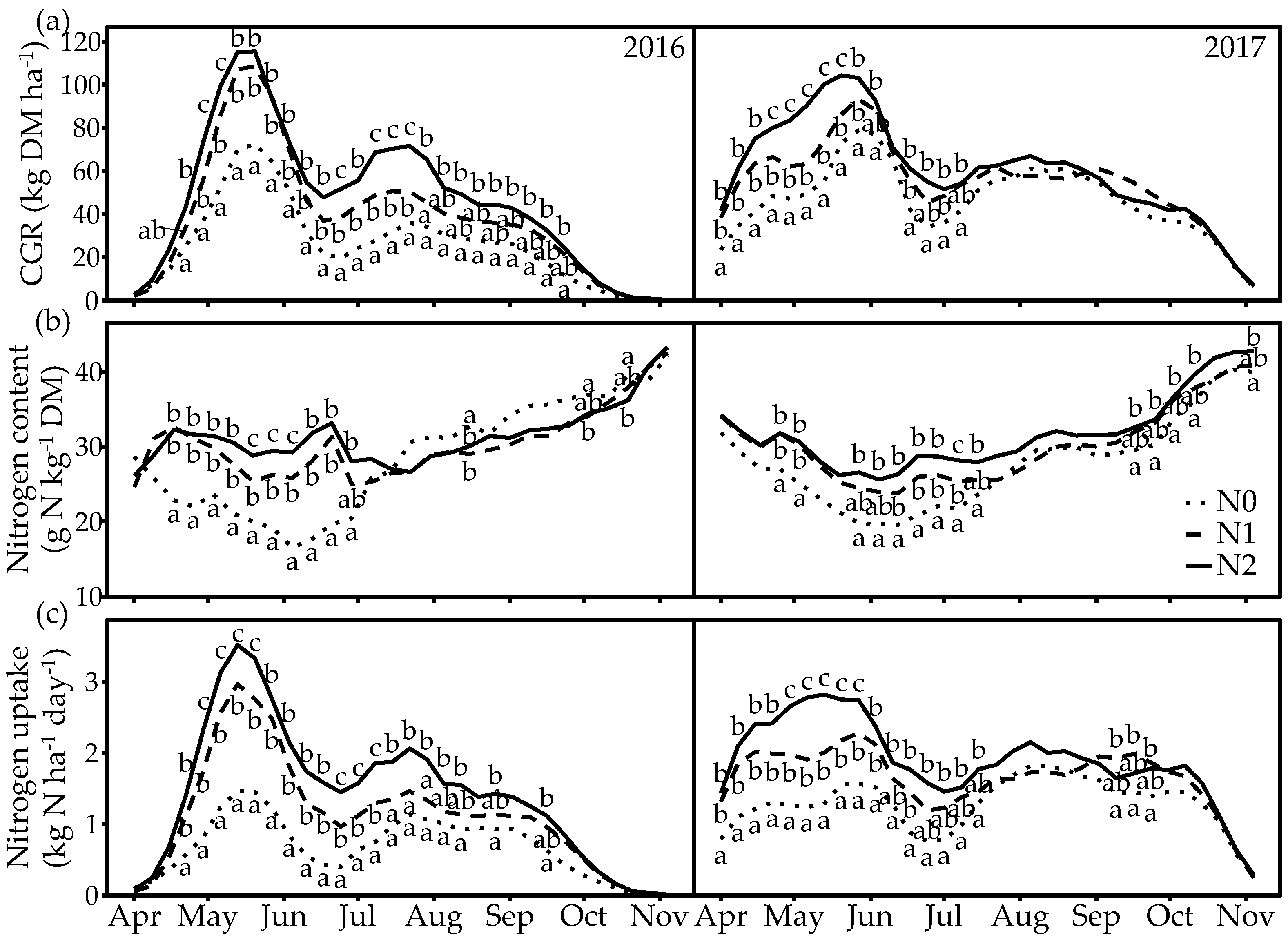
3.2. Herbage Production, Nitrogen Content and Nitrogen Uptake
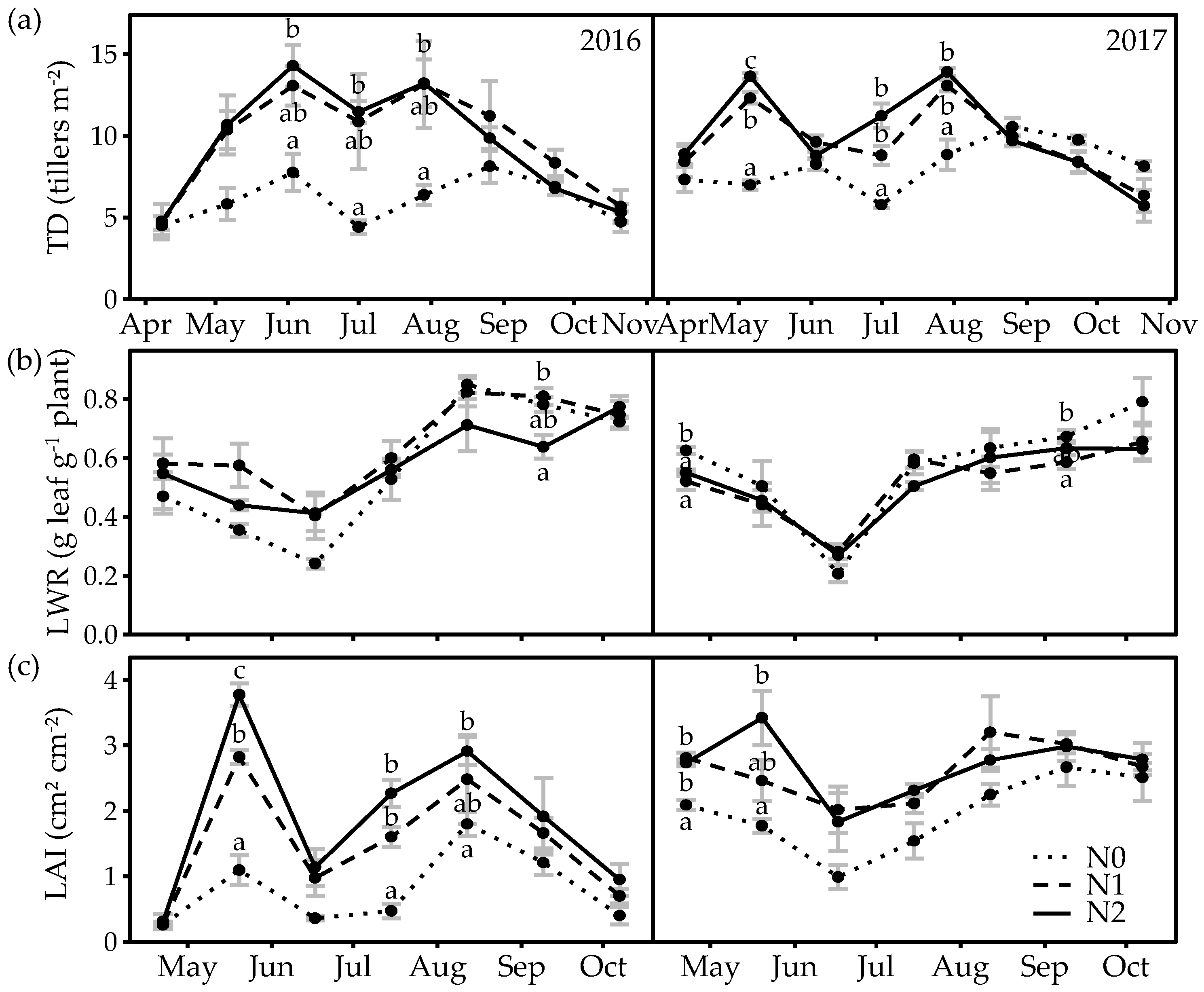
3.3. Phenology and Tiller Weight
3.4. Leaf- and Tiller-Related Plant Functional Traits
3.5. Relationships of Each PFT to CGR and Nitrogen Uptake
4. Discussion
4.1. Seasonal Dynamics in CGR, Nitrogen Uptake and PFTs
4.2. Effect of N Application Rate on Herbage Production and PFT
4.3. Relationships of Each PFT to CGR and Nitrogen Uptake
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Enriquez-Hidalgo, D.; Gilliland, T.J.; Hennessy, D. Herbage and nitrogen yields, fixation and transfer by white clover to companion grasses in grazed swards under different rates of nitrogen fertilization. Grass Forage Sci. 2016, 71, 559–574. [Google Scholar] [CrossRef]
- Bryant, R.H.; Gregorini, P.; Edwards, G.R. Effects of N fertilisation, leaf appearance and time of day on N fractionation and chemical composition of Lolium perenne cultivars in spring. Anim. Feed Sci. Technol. 2012, 173, 210–219. [Google Scholar] [CrossRef]
- Trott, H.; Wachendorf, M.; Ingwersen, B.; Taube, F. Performance and environmental effects of forage production on sandy soils I. Impact of defoliation system and nitrogen input on performance and N balance of grassland. Grass Forage Sci. 2004, 59, 41–55. [Google Scholar] [CrossRef]
- Burchill, W.; Lanigan, G.J.; Li, D.; Williams, M.; Humphreys, J. A system N balance for a pasture-based system of dairy production under moist maritime climatic conditions. Agric. Ecosyst. Environ. 2016, 220, 202–210. [Google Scholar] [CrossRef]
- Powell, J.M.; Rotz, C.A. Measures of nitrogen use efficiency and nitrogen loss from dairy production systems. J. Environ. Qual. 2015, 44, 336–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinsch, T.; Loza, C.; Malisch, C.S.; Vogeler, I.; Kluß, C.; Loges, R.; Taube, F. Toward specialized or integrated systems in Northwest Europe: On-farm eco-efficiency of dairy farming in Germany. Front. Sustain. Food Syst. 2021, 5, 4348. [Google Scholar] [CrossRef]
- Moir, J.L.; Edwards, G.R.; Berry, L.N. Nitrogen uptake and leaching loss of thirteen temperate grass species under high N loading. Grass Forage Sci. 2013, 68, 313–325. [Google Scholar] [CrossRef]
- Gastal, F.; Lemaire, G. N uptake and distribution in crops: An agronomical and ecophysiological perspective. J. Exp. Bot. 2002, 53, 789–799. [Google Scholar] [CrossRef] [Green Version]
- European Union. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 establishing a framework for Community action in the field of water policy. Off. J. Eur. Union 2000, L327. Available online: http://data.europa.eu/eli/dir/2000/60/oj (accessed on 2 August 2021).
- European Union. Council Directive of 12 December 1991 concerning the protection of waters against pollution caused by nitrates from agricultural sources (91/676/EEC); amended by regulation (EC) No 1137/2008 on 22 October 2008. Off. J. Eur. Union 2008, 4, 144–197. Available online: http://data.europa.eu/eli/dir/1991/676/2008-12-11 (accessed on 2 August 2021).
- Parsons, A.; Chapman, D. The principles of pasture growth and utilization. In Grass, Its Production and Utilization, 3rd ed.; Hopkins, A., Ed.; Blackwell Science: Oxford, UK, 2000; pp. 31–89. [Google Scholar]
- Gastal, F.; Lemaire, G. Defoliation, shoot plasticity, sward structure and herbage utilization in pasture: Review of the underlying ecophysiological processes. Agriculture 2015, 5, 1146–1171. [Google Scholar] [CrossRef] [Green Version]
- Gautier, H.; Varlet-Grancher, C.; Hazard, L. Tillering responses to the light environment and to defoliation in populations of perennial ryegrass (Lolium perenne L) selected for contrasting leaf length. Ann. Bot. 1999, 83, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Matthew, C.; Assuero, S.; Black, C.; Hamilton, N.S. Tiller dynamics of grazed swards. In Grassland Ecophysiology and Grazing Ecology; Lemaire, G., Hodgson, J., de Moraes, P.C., Nabinger, C., de Carvalho, P.C.F., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 127–150. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Laliberté, E.; Shipley, B.; Norton, D.A.; Scott, D. Which plant traits determine abundance under long-term shifts in soil resource availability and grazing intensity? J. Ecol. 2012, 100, 662–677. [Google Scholar] [CrossRef]
- Beecher, M.; Hennessy, D.; Boland, T.; McEvoy, M.; O’Donovan, M.; Lewis, E. The variation in morphology of perennial ryegrass cultivars throughout the grazing season and effects on organic matter digestibility. Grass Forage Sci. 2015, 70, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Grassein, F.; Lemauviel-Lavenant, S.; Lavorel, S.; Bahn, M.; Bardgett, R.D.; Desclos-Theveniau, M.; Laine, P. Relationships between functional traits and inorganic nitrogen acquisition among eight contrasting European grass species. Ann. Bot. 2015, 115, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Pontes, L.D.S.; Soussana, J.F.; Louault, F.; Andueza, D.; Carrère, P. Leaf traits affect the above-ground productivity and quality of pasture grasses. Funct. Ecol. 2007, 21, 844–853. [Google Scholar] [CrossRef]
- Schellberg, J.; Pontes, L.D.S. Plant functional traits and nutrient gradients on grassland. Grass Forage Sci. 2012, 67, 305–319. [Google Scholar] [CrossRef]
- Hanisch, M.; Schweiger, O.; Cord, A.F.; Volk, M.; Knapp, S. Plant functional traits shape multiple ecosystem services, their trade-offs and synergies in grasslands. J. Appl. Ecol. 2020, 57, 1535–1550. [Google Scholar] [CrossRef]
- Hendricks, S.; Donaghy, D.; Matthew, C.; Bretherton, M.; Sneddon, N.; Cosgrove, G.; Christensen, C.; Kaufononga, S.; Howes, J.; Osborne, M.; et al. Dry matter yield, nutritive value and tiller density of tall fescue and perennial ryegrass swards under grazing. J. N. Z. Grassl. 2016, 78, 149–156. [Google Scholar] [CrossRef]
- Komainda, M.; Isselstein, J. Effects of functional traits of perennial ryegrass cultivars on forage quality in mixtures and pure stands. J. Agric. Sci. 2020, 158, 173–184. [Google Scholar] [CrossRef]
- Van Arendonk, J.J.C.M.; Niemann, G.J.; Boon, J.J.; Lambers, H. Effects of nitrogen supply on the anatomy and chemical composition of leaves of four grass species belonging to the genus Poa, as determined by image-processing analysis and pyrolysis-mass spectrometry. Plant. Cell Environ. 1997, 20, 881–897. [Google Scholar] [CrossRef]
- An, H.; Shangguan, Z.P. Specific leaf area, leaf nitrogen content, and photosynthetic acclimation of Trifolium repens L seedlings grown at different irradiances and nitrogen concentrations. Photosynthetica 2008, 46, 143–147. [Google Scholar] [CrossRef]
- Corrall, A.J.; Fenlon, J.S. A comparative method for describing the seasonal distribution of production from grasses. J. Agric. Sci. 1978, 91, 61–67. [Google Scholar] [CrossRef]
- Düngung von Wiesen, D.L.G. Weiden und Feldfutter DLG-Merkblatt 433. 2018. Available online: https://www.dlg.org/fileadmin/downloads/landwirtschaft/themen/publikationen/merkblaetter/dlg-merkblatt_433.pdf (accessed on 3 May 2021).
- Whitehead, D.C. Nutrient Elements in Grassland: Soil-Plant.-Animal Relationships; CABI Publishing: Oxon, UK, 2000. [Google Scholar] [CrossRef] [Green Version]
- Baker, C.W.; Barnes, R. The application of Near Infra-Red Spectrometry to forage evaluation in the agricultural development and advisory service. In Feedstuff Evaluation; Wiseman, J., Cole, D.J.A., Eds.; Butterworth-Heinemann: Oxford, UK, 1990; pp. 337–351. [Google Scholar] [CrossRef]
- Trott, H.; Ingwersen, B.; Wachendorf, M.; Taube, F. Estimation of dry matter yield on permanent grassland by means of height assessment. Pflanzenbauwissenschaften 2002, 6, 78–83. [Google Scholar]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Farzadfar, S.; Williams, S.; Arcand, M.M. Nitrogen use efficiency definitions of today and tomorrow. Front. Plant. Sci. 2021, 12, 7108. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.J.; Moser, L.E.; Vogel, K.P.; Waller, S.S.; Johnson, B.E.; Pedersen, J.F. Describing and quantifying growth stages of perennial forage grasses. Agron. J. 1991, 83, 1073–1077. [Google Scholar] [CrossRef] [Green Version]
- Verbeke, G.; Molenberghs, G. Linear Mixed Models for Longitudinal Data; Springer: New York, NY, USA, 2013. [Google Scholar]
- Kozak, M.; Piepho, H.-P. What’s normal anyway? Residual plots are more telling than significance tests when checking ANOVA assumptions. J. Agron. Crop. Sci. 2018, 204, 86–98. [Google Scholar] [CrossRef]
- Hassink, J. Effects of soil texture and grassland management on soil organic C and N and rates of C and N mineralization. Soil Biol. Biochem. 1994, 26, 1221–1231. [Google Scholar] [CrossRef]
- Soussana, J.-F.; Lemaire, G. Coupling carbon and nitrogen cycles for environmentally sustainable intensification of grasslands and crop-livestock systems. Agric. Ecosyst. Environ. 2014, 190, 9–17. [Google Scholar] [CrossRef]
- Tilib, E.; Younger, S. The influence of fertilizer nitrogen, white clover content and environmental factors on the nitrate content of perennial ryegrass and ryegrass/white clover swards. Grass Forage Sci. 1999, 54, 275–285. [Google Scholar] [CrossRef]
- King, C.; McEniry, J.; Richardson, M.; O’Kiely, P. Yield and chemical composition of five common grassland species in response to nitrogen fertiliser application and phenological growth stage. Acta Agric. Scand. Sect. B Soil Plant. Sci. 2012, 62, 644–658. [Google Scholar] [CrossRef]
- Anslow, R.C.; Green, J.O. The seasonal growth of pasture grasses. J. Agric. Sci. 1967, 68, 109–122. [Google Scholar] [CrossRef]
- Lemaire, G.; Oosterom, E.v.; Sheehy, J.; Jeuffroy, M.H.; Massignam, A.; Rossato, L. Is crop N demand more closely related to dry matter accumulation or leaf area expansion during vegetative growth? Field Crops Res. 2007, 100, 91–106. [Google Scholar] [CrossRef]
- Nassiri, M.; Elgersma, A. Effects of nitrogen on leaves, dry matter allocation and regrowth dynamics in Trifolium repens L. and Lolium perenne L. in pure and mixed swards. Plant. Soil 2002, 246, 107–121. [Google Scholar] [CrossRef]
- Nassiri, M.; Elgersma, A. Competition in perennial ryegrass–white clover mixtures under cutting 2. Leaf characteristics, light interception and dry-matter production during regrowth. Grass Forage Sci. 1998, 53, 367–379. [Google Scholar] [CrossRef]
- Orr, R.J.; Parsons, A.J.; Treacher, T.T.; Penning, P.D. Seasonal patterns of grass production under cutting or continuous stocking managements. Grass Forage Sci. 1988, 43, 199–207. [Google Scholar] [CrossRef]
- Korte, C.J.; Watkin, B.R.; Harris, W. Tillering in ‘Grasslands Nui’ perennial ryegrass swards 1 Effect of cutting treatments on tiller appearance and longevity, relationship between tiller age and weight, and herbage production. N. Z. J. Agric. Res. 1985, 28, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Ong, C.K.; Marshall, C. The growth and survival of severely-shaded Tillers in Lolium perenne L. Ann. Bot. 1979, 43, 147–155. [Google Scholar] [CrossRef]
- Taube, F. Growth characteristics of contrasting varieties of perennial ryegrass (Lolium perenne L.). J. Agron. Crop. Sci. 1990, 165, 159–170. [Google Scholar] [CrossRef]
- Cruz, P.; Boval, M. Effect of nitrogen on some morphogenetic traits of temperate and tropical perennial forage grasses. In Grassland Ecophysiology and Grazing Ecology; Lemaire, G., Hodgson, J., de Moraes, P.C., Nabinger, C., de Carvalho, P.C.F., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 151–168. [Google Scholar] [CrossRef]
- Akmal, M.; Janssens, M.J.J. Productivity and light use efficiency of perennial ryegrass with contrasting water and nitrogen supplies. Field Crops Res. 2004, 88, 143–155. [Google Scholar] [CrossRef]
- Joy Pearse, P.; Wilman, D. Effects of applied nitrogen on grass leaf initiation, development and death in field swards. J. Agric. Sci. 1984, 103, 405–413. [Google Scholar] [CrossRef]
- Heide, O.M. Control of flowering and reproduction in temperate grasses. New Phytol. 1994, 128, 347–362. [Google Scholar] [CrossRef]
- Macadam, J.W.; Volenec, J.J.; Nelson, C.J. Effects of nitrogen on mesophyll cell division and epidermal cell elongation in tall fescue leaf blades. Plant. Physiol. 1989, 89, 549–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arredondo, J.T.; Schnyder, H. Components of leaf elongation rate and their relationship to specific leaf area in contrasting grasses. New Phytol. 2003, 158, 305–314. [Google Scholar] [CrossRef]
- Jamieson, N.; Barraclough, D.; Unkovich, M.; Monaghan, R. Soil N dynamics in a natural calcareous grassland under a changing climate. Biol. Fertil. Soils 1998, 27, 267–273. [Google Scholar] [CrossRef]
- Verloop, J.; Hilhorst, G.J.; Oenema, J.; van Keulen, H.; Sebek, L.B.J.; van Ittersum, M.K. Soil N mineralization in a dairy production system with grass and forage crops. Nutr. Cycl. Agroecosyst. 2014, 98, 267–280. [Google Scholar] [CrossRef]
- Garnier, E.; Cordonnier, P.; Guillerm, J.L.; Sonié, L. Specific leaf area and leaf nitrogen concentration in annual and perennial grass species growing in mediterranean old-fields. Oecologia 1997, 111, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Knops, J.M.; Reinhart, K. Specific leaf area along a nitrogen fertilization gradient. Am. Midl. Nat. 2000, 144, 265–272. [Google Scholar] [CrossRef]
- Matthew, C.; Hernández-Garay, A.; Hodgson, J. Making sense of the link between tiller density and pasture production. Proc. N. Z. Grassl. Assoc. 1996, 57, 83–87. [Google Scholar] [CrossRef]
- Garay, A.H.; Mathew, C.; Hodgson, J. Tiller size/density compensation in perennial ryegrass miniature swards subject to differing defoliation heights and a proposed productivity index. Grass Forage Sci. 1999, 54, 347–356. [Google Scholar] [CrossRef]
- Garay, A.H.; Matthew, C.; Hodgson, J. Effect of spring grazing management on perennial ryegrass and ryegrass-white clover pastures. N. Z. J. Agric. Res. 1997, 40, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Neuteboom, J.H.; Lantinga, E. Tillering potential and relationship between leaf and tiller production in perennial ryegrass. Ann. Bot. 1989, 63, 265–270. [Google Scholar] [CrossRef]
- Seefeldt, S.S.; Armstrong, L.M. Impact of perennial ryegrass seeding densities on weed emergence growth and development. N. Z. Plant. Prot. 2000, 53, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Klapp, E.; Stählin, A. Standorte, Pflanzengesellschaften und Leistung des Grünlandes: Am Beispiel Thüringischer Wiesen; Ulmer: Stuttgart, Germany, 1936. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | Y | VP |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean temperature (°C) | ||||||||||||||
| 2016 | 1.1 | 3.5 | 4.5 | 7.2 | 13.6 | 17.1 | 17.8 | 17.1 | 17.5 | 9.8 | 4.6 | 4.8 | 9.9 | 14.3 |
| 2017 | 1.3 | 2.8 | 6.5 | 7.2 | 13.3 | 16.3 | 16.7 | 16.9 | 13.9 | 12.0 | 6.3 | 4.2 | 9.8 | 13.8 |
| 1991–2020 | 1.7 | 2.2 | 4.3 | 8.1 | 12.0 | 15.3 | 17.8 | 17.6 | 14.3 | 10.0 | 5.8 | 2.9 | 9.3 | 13.6 |
| Growing degree days > 5 °C (°C) | ||||||||||||||
| 2016 | 17 | 16 | 21 | 74 | 267 | 364 | 396 | 375 | 375 | 150 | 32 | 33 | 2119 | 2000 |
| 2017 | 0 | 14 | 57 | 71 | 257 | 339 | 363 | 368 | 266 | 218 | 51 | 23 | 2027 | 1882 |
| 1991–2020 | 14 | 14 | 31 | 100 | 218 | 309 | 394 | 390 | 277 | 158 | 54 | 21 | 1981 | 1847 |
| Rainfall (mm) | ||||||||||||||
| 2016 | 82 | 85 | 28 | 64 | 27 | 94 | 87 | 54 | 41 | 79 | 32 | 43 | 716 | 446 |
| 2017 | 39 | 59 | 50 | 47 | 42 | 118 | 100 | 61 | 76 | 125 | 89 | 83 | 889 | 569 |
| 1991–2020 | 67 | 51 | 52 | 38 | 50 | 64 | 83 | 78 | 62 | 74 | 65 | 71 | 754 | 449 |
| Solar radiation (MJ m−2) | ||||||||||||||
| 2016 | 53 | 118 | 235 | 379 | 610 | 584 | 536 | 498 | 379 | 143 | 74 | 40 | 3647 | 2964 |
| 2017 | 60 | 95 | 269 | 414 | 613 | 585 | 528 | 477 | 309 | 162 | 70 | 41 | 3623 | 2958 |
| 1991–2020 | 60 | 114 | 259 | 432 | 575 | 586 | 588 | 497 | 320 | 180 | 75 | 44 | 3729 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peters, T.; Taube, F.; Kluß, C.; Reinsch, T.; Loges, R.; Fenger, F. How Does Nitrogen Application Rate Affect Plant Functional Traits and Crop Growth Rate of Perennial Ryegrass-Dominated Permanent Pastures? Agronomy 2021, 11, 2499. https://doi.org/10.3390/agronomy11122499
Peters T, Taube F, Kluß C, Reinsch T, Loges R, Fenger F. How Does Nitrogen Application Rate Affect Plant Functional Traits and Crop Growth Rate of Perennial Ryegrass-Dominated Permanent Pastures? Agronomy. 2021; 11(12):2499. https://doi.org/10.3390/agronomy11122499
Chicago/Turabian StylePeters, Tammo, Friedhelm Taube, Christof Kluß, Thorsten Reinsch, Ralf Loges, and Friederike Fenger. 2021. "How Does Nitrogen Application Rate Affect Plant Functional Traits and Crop Growth Rate of Perennial Ryegrass-Dominated Permanent Pastures?" Agronomy 11, no. 12: 2499. https://doi.org/10.3390/agronomy11122499