
Palmer Amaranth (Amaranthus palmeri) Growth and Seed Production When in Competition with Peanut and Other Crops in North Carolina

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
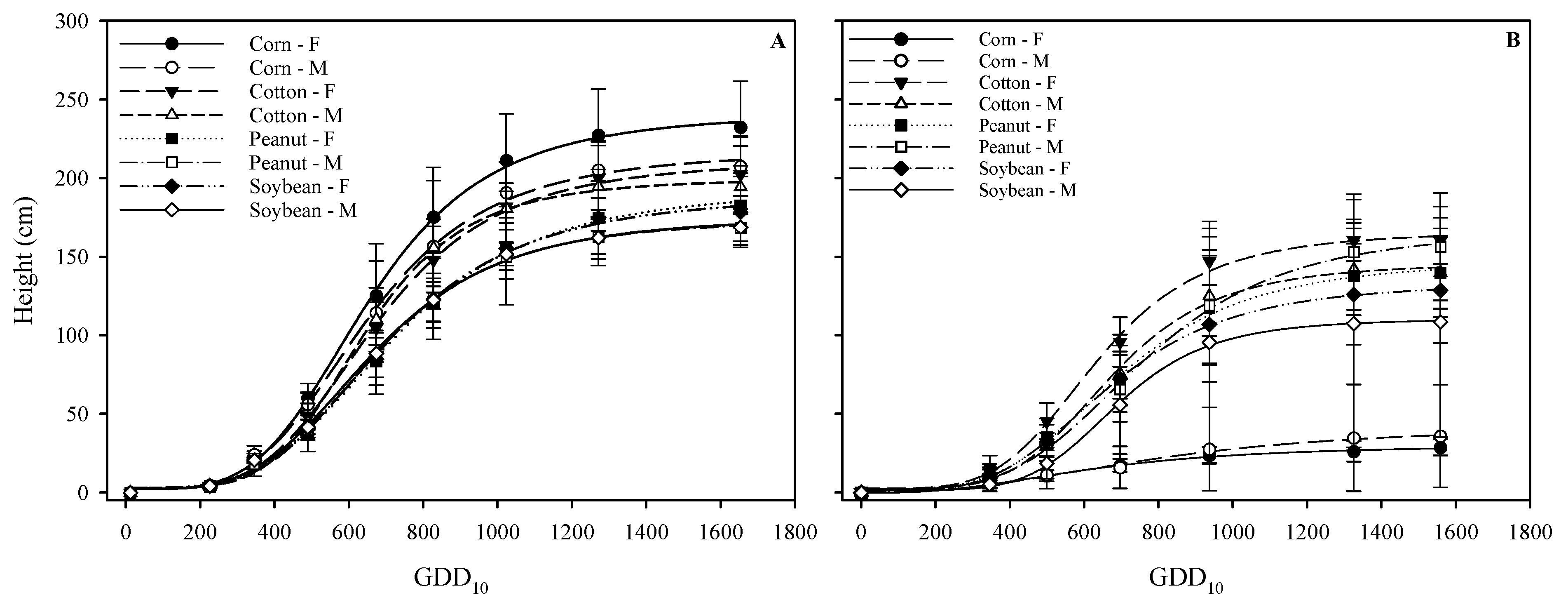
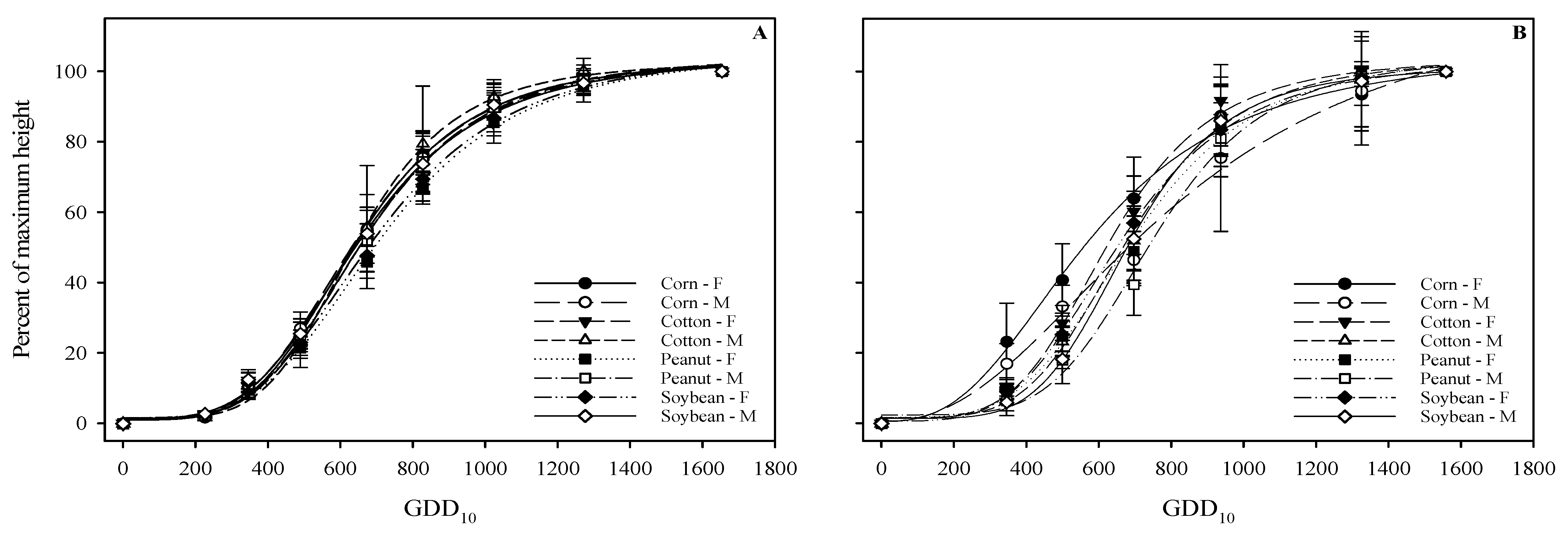
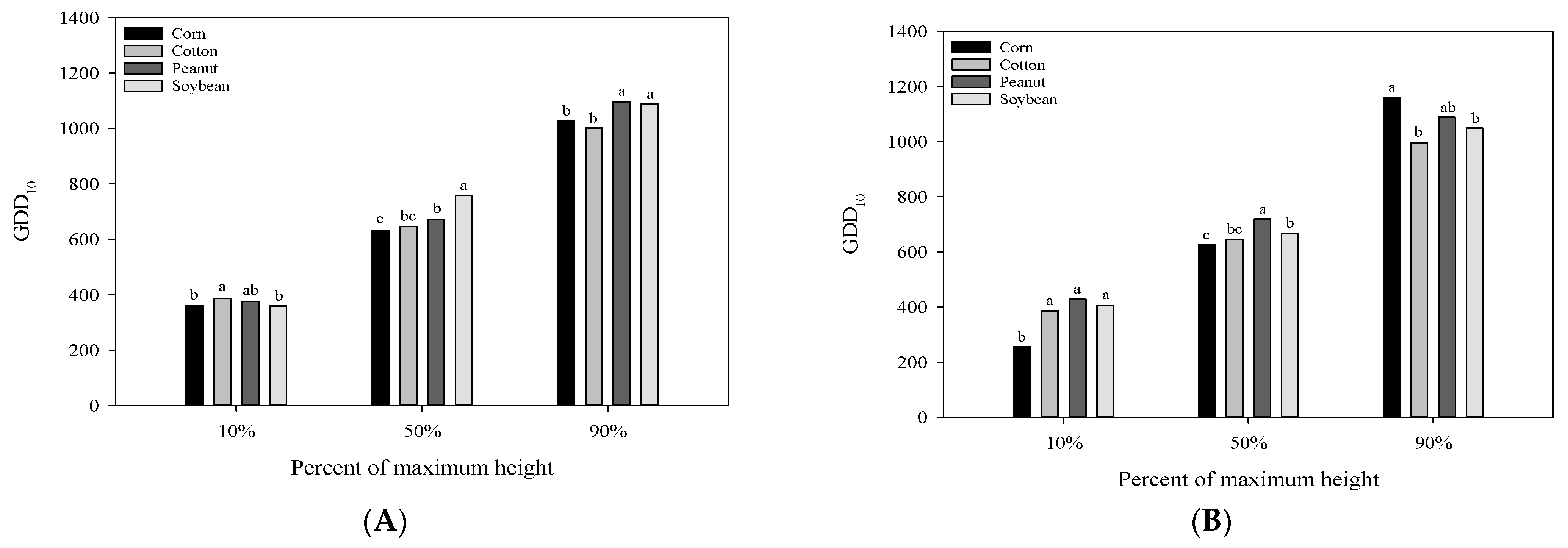
3.1. Plant Height
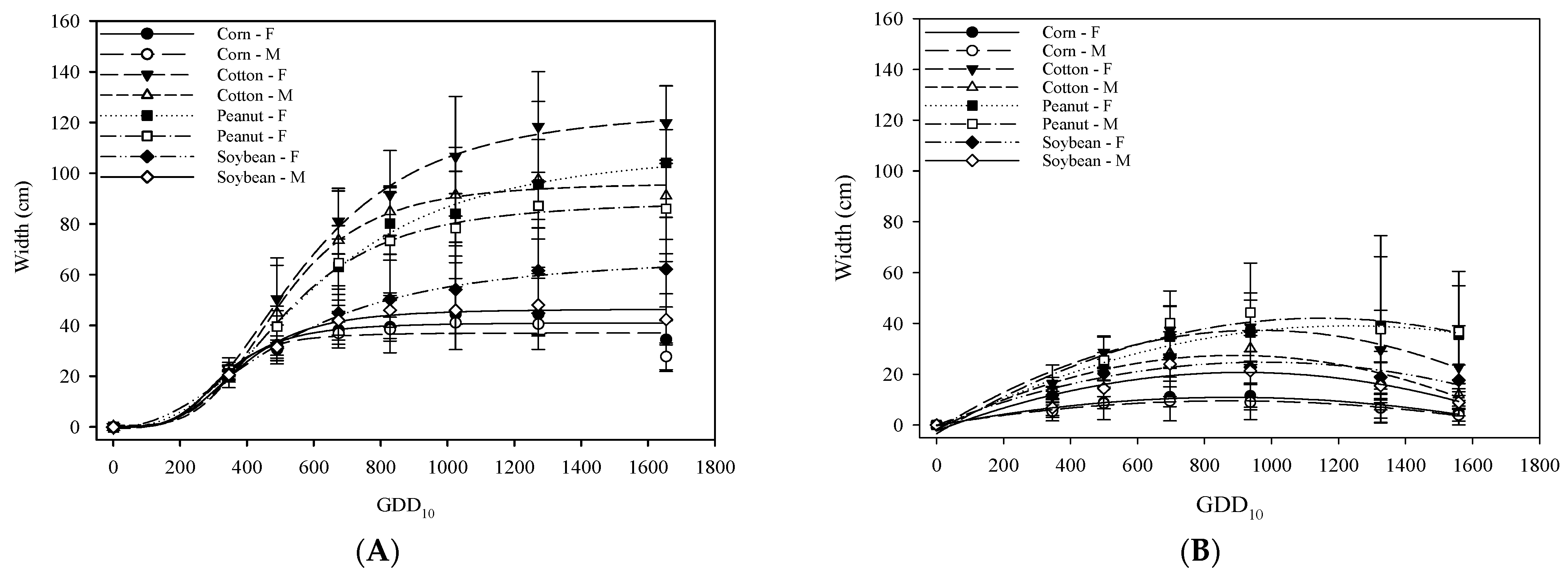
3.2. Canopy Wdth of A. palmeri
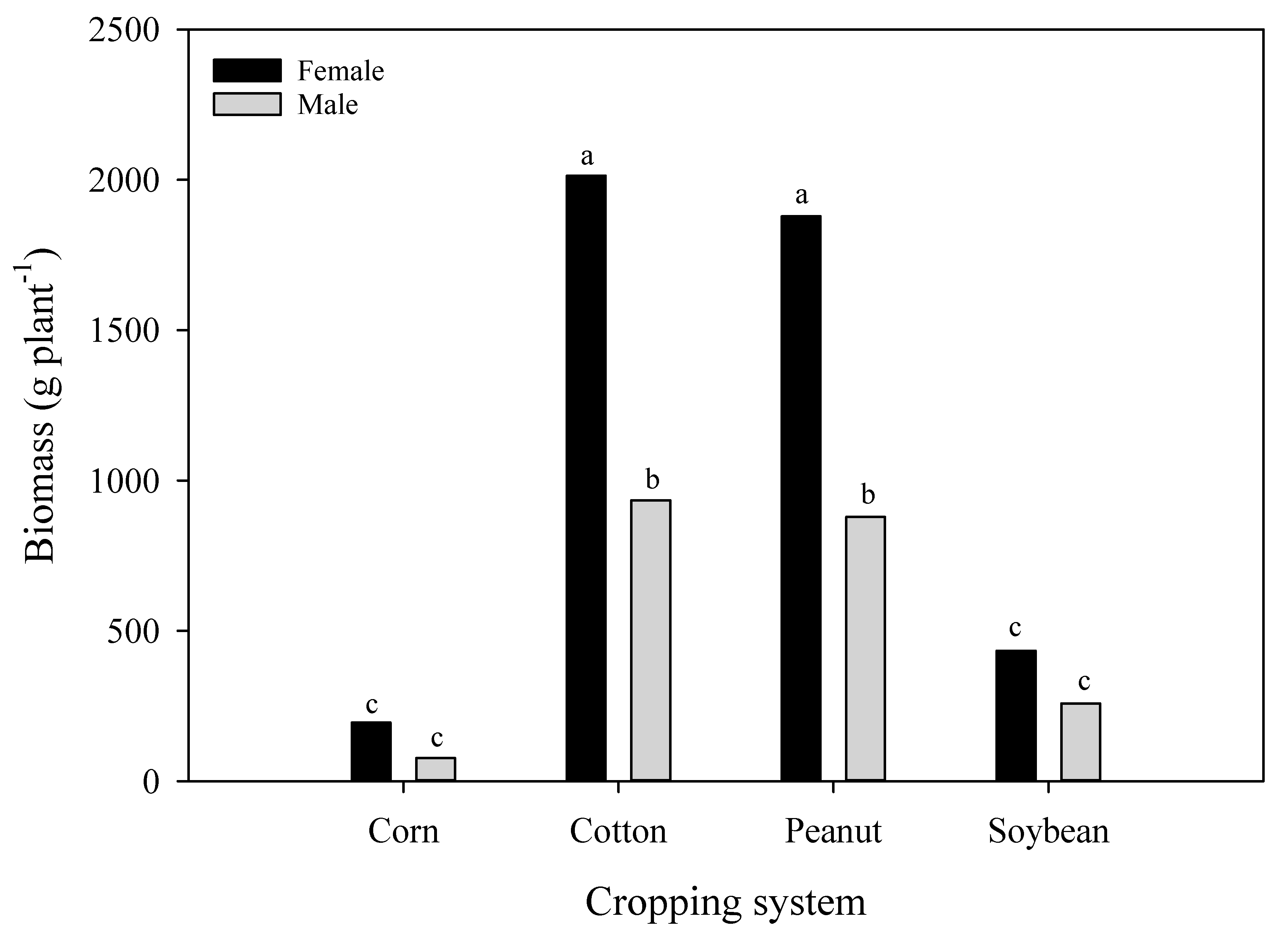
3.3. A. palmeri Soot Bomass
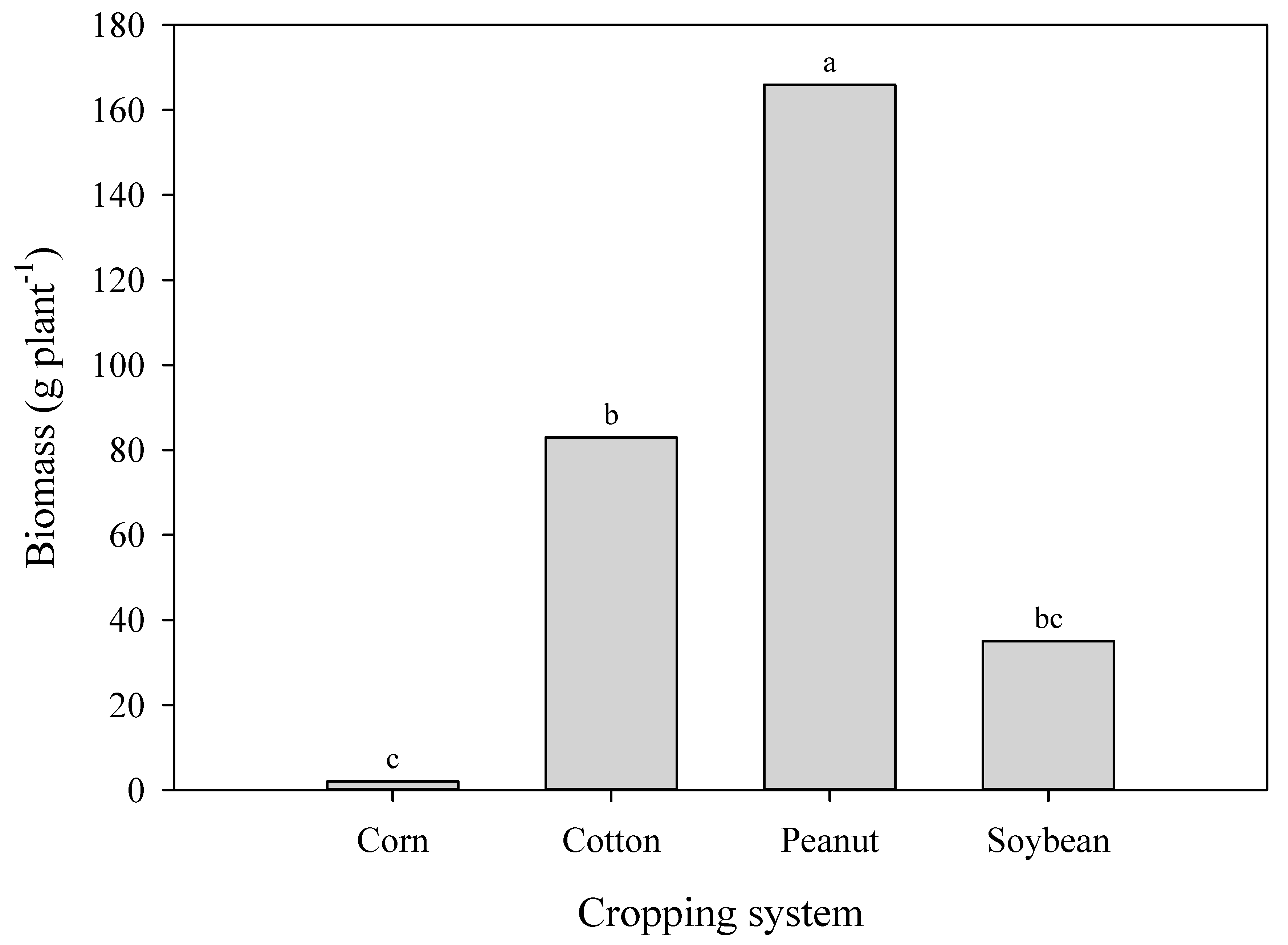
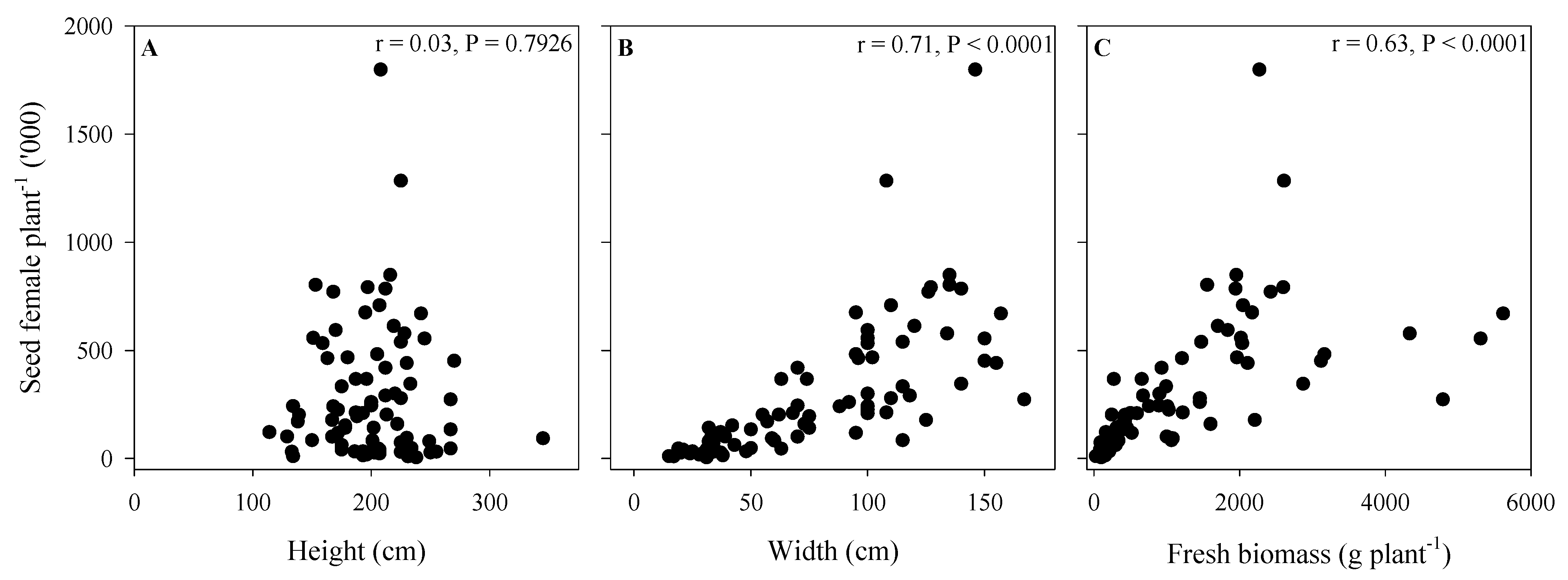
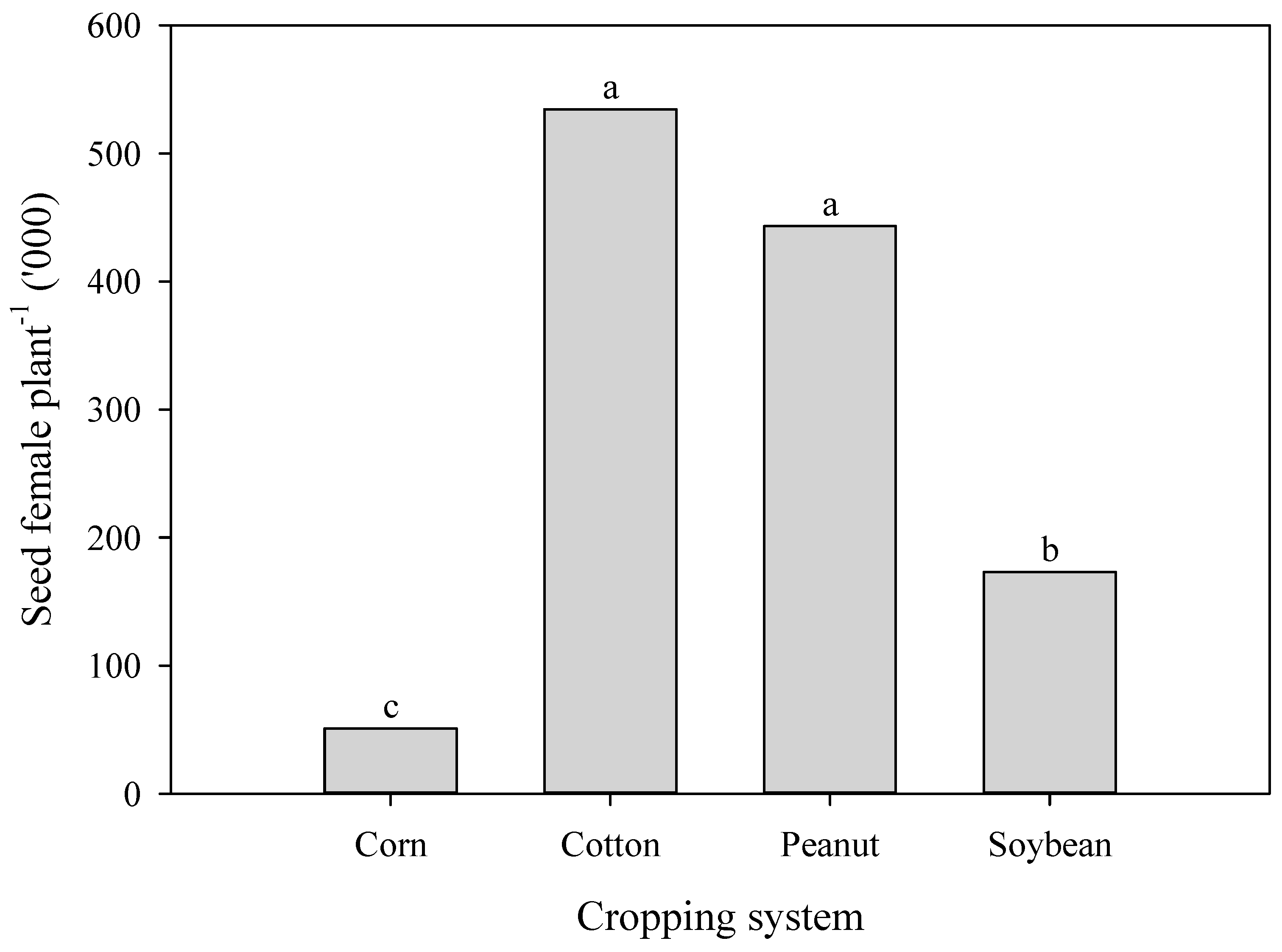
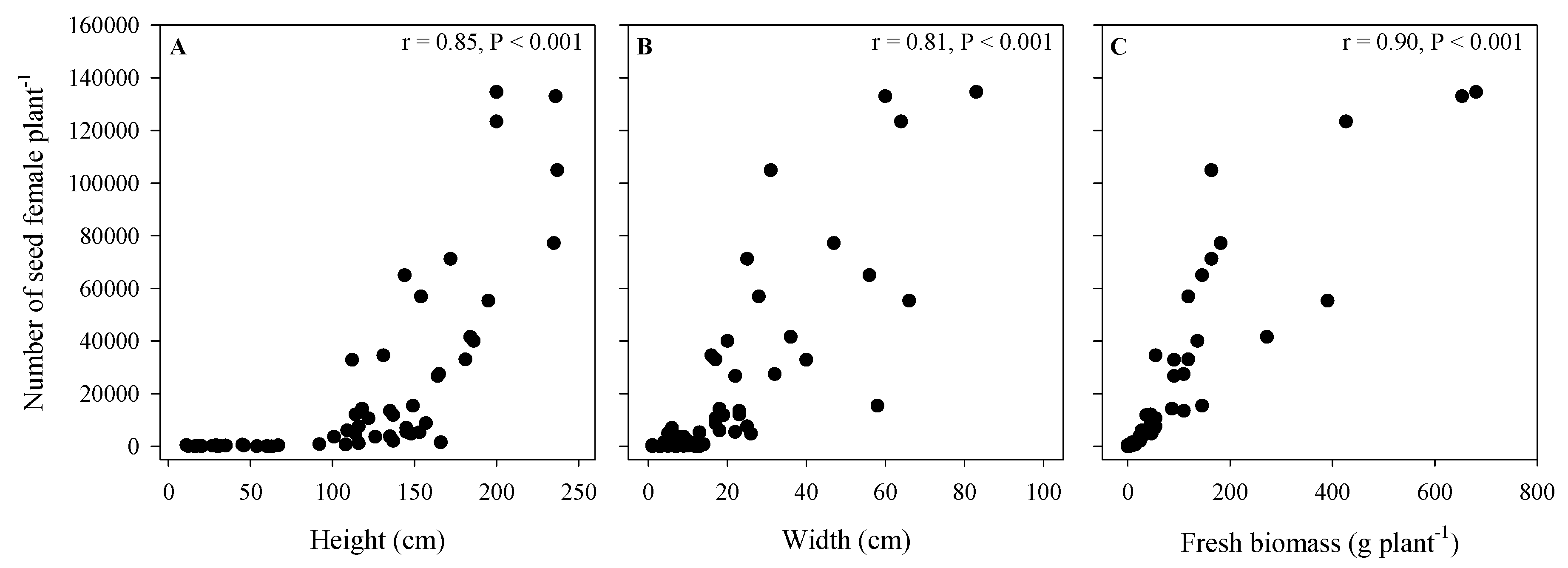
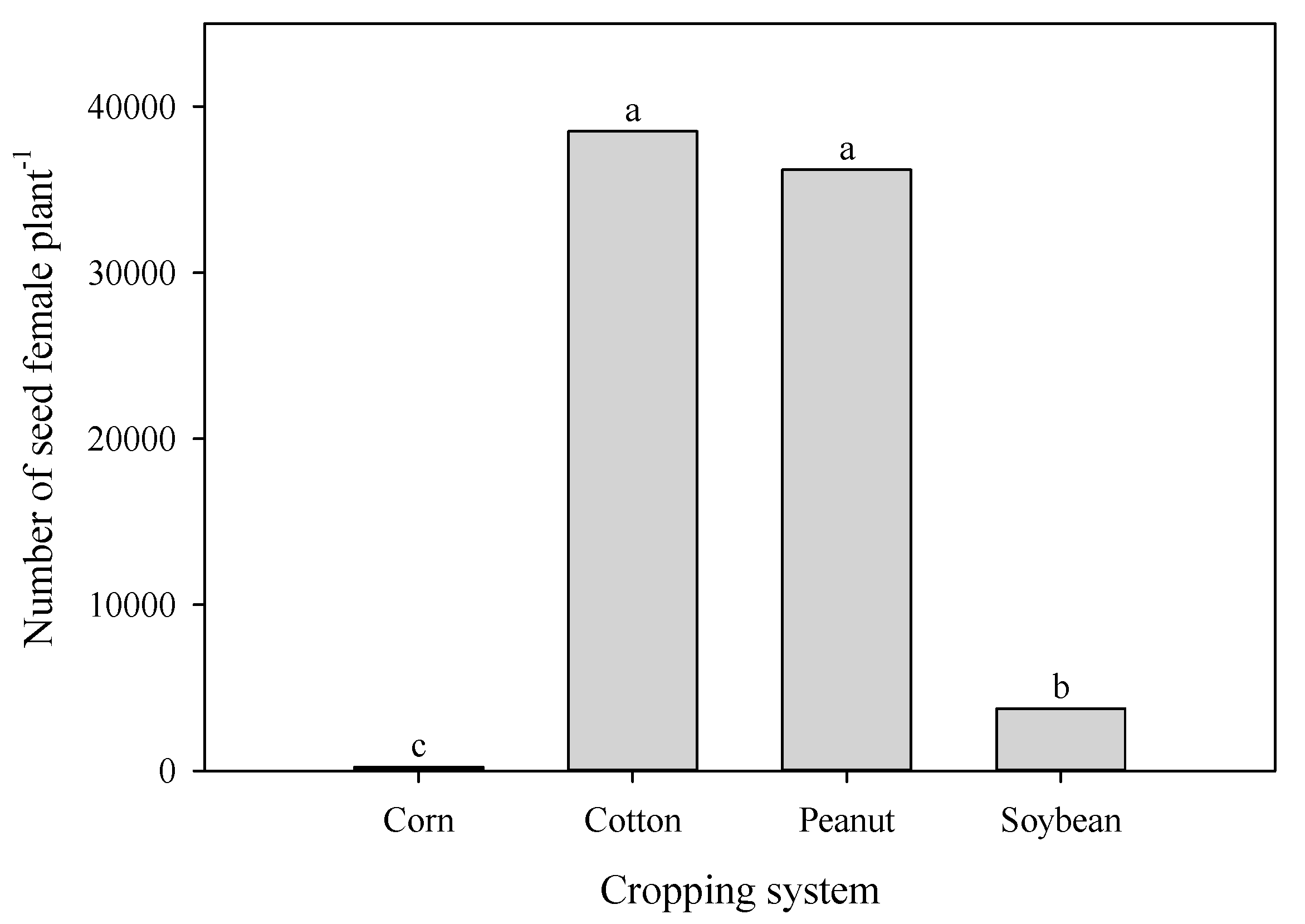
3.4. Seed Poduction of A. palmeri in Cop Cmpetition
4. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Wychen, L. WSSA Survey Ranks Palmer Amaranth as the Most Troublesome Weed in the US, Galium as the Most Troublesome in Canada. Weed Science Society of America. 2016. Available online: http://wssa.net/2016/04/wssa-survey-ranks-palmer-amaranth-as-the-most-troublesome-weed-in-the-u-s-galium-as-the-most-troublesome-in-canada/ (accessed on 12 December 2019).
- Keeley, P.E.; Carter, C.H.; Thullen, R.J. Influence of planting date on growth of Palmer amaranth (Amaranthus palmeri). Weed Sci. 1987, 35, 199–204. [Google Scholar] [CrossRef]
- Spaunhorst, D.J.; Devkota, P.; Johnson, W.G.; Smeda, R.J.; Meyer, C.J.; Norsworthy, J.K. Phenology of five Palmer amaranth (Amaranthus palmeri) populations grown in northern Indiana and Arkansas. Weed Sci. 2018, 66, 457–469. [Google Scholar] [CrossRef]
- Knight, A.M.; Everman, W.J.; Jordan, D.L.; Heiniger, R.W.; Smyth, T.J. Interactions of nitrogen source and rate and weed removal timing relative to nitrogen content in corn and weeds and corn grain yield. Int. Sch. Res. Not. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyers, S.L.; Jennings, K.M.; Schultheis, J.R.; Monks, D.W. Interference of Palmer amaranth (Amaranthus palmeri) in sweetpotato. Weed Sci. 2010, 58, 199–203. [Google Scholar] [CrossRef]
- Berger, S.T.; Ferrell, J.A.; Rowland, D.L.; Webster, T.W. Palmer amaranth (Amaranthus palmeri) competition for water in cotton. Weed Sci. 2015, 63, 928–935. [Google Scholar] [CrossRef]
- Chandi, A.; Milla-Lewis, S.R.; Jordan, D.L.; York, A.C.; Burton, J.D.; Zuleta, M.C.; Whitaker, J.R.; Culpepper, A.S. Use of AFLP markers to assess genetic diversity in Palmer amaranth (Amaranthus palmeri) populations from North Carolina and Georgia. Weed Sci. 2013, 61, 136–145. [Google Scholar] [CrossRef]
- Sosnoskie, L.M.; Webster, T.M.; Kichler, J.M.; MacRae, A.W.; Grey, T.L.; Culpepper, A.S. Pollen mediated dispersal of glyphosate-resistance in Palmer amaranth under field conditions. Weed Sci. 2012, 60, 366–373. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2021. Available online: www.weedscience.org (accessed on 12 December 2019).
- Ward, S.M.; Webster, T.M.; Steckel, L.E. Palmer amaranth (Amaranthus palmeri): A review. Weed Technol. 2013, 27, 12–27. [Google Scholar] [CrossRef] [Green Version]
- Burke, I.C.; Schroeder, M.; Thomas, W.E.; Wilcut, J.W. Palmer amaranth interference and seed production in peanut. Weed Technol. 2007, 21, 367–371. [Google Scholar] [CrossRef]
- Webster, T.M.; Grey, T.L. Glyphosate-resistant Palmer amaranth (Amaranthus palmeri) morphology, growth, and seed production in Georgia. Weed Sci. 2015, 63, 264–272. [Google Scholar] [CrossRef]
- Bagavathiannan, M.V.; Davis, A.S. An ecological perspective on managing weeds during the great selection for herbicide resistance. Pest. Manag. Sci. 2018, 74, 2277–2286. [Google Scholar] [CrossRef]
- Horak, M.J.; Loughin, T.M. Growth analysis of four Amaranthus species. Weed Sci. 2000, 48, 347–355. [Google Scholar] [CrossRef]
- Sellers, B.A.; Smeda, R.J.; Johnson, W.G.; Kendig, J.A.; Ellersieck, M.R. Comparative growth of six Amaranthus species in Missouri. Weed Sci. 2003, 51, 329–333. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Schrage, B.W.; Barber, T.L.; Schwartz, L.M. Emergence date influences growth and fecundity of Palmer amaranth in cotton. J. Cotton Sci. 2016, 20, 263–270. [Google Scholar]
- Van Wychen, L. Scientists Advocate a Community-Based Approach to Herbicide Resistance Management. Weed Science Society of America. 2015. Available online: http://wssa.net/2015/06/scientists-advocate-a-community-based-approach-to-herbicide-resistance-management/ (accessed on 12 December 2019).
- Poirier, A.H.; York, A.C.; Jordan, D.L.; Chandi, A.; Everman, W.J.; Whitaker, J.R. Distribution of glyphosate- and thifensulfuron-resistant Palmer amaranth (Amaranthus palmeri) in North Carolina. Int. J. Agron. 2014. [Google Scholar] [CrossRef]
- Inman, M.D.; Jordan, D.L.; York, A.C.; Jennings, K.M.; Monks, D.W.; Everman, W.J.; Bollman, S.T.; Fowler, J.T.; Cole, R.M.; Soteres, J.K. Long-term management of Palmer amaranth in dicamba-tolerant cotton. Weed Sci. 2016, 64, 161–169. [Google Scholar] [CrossRef]
- Isleib, T.G.; Milla-Lewis, S.R.; Pattee, H.E.; Copeland, S.C.; Zuleta, M.C.; Shew, B.B.; Hollowell, J.E.; Sanders, T.H.; Dean, L.O.; Hendrix, K.W.; et al. Registration of ‘Bailey’ peanut. J. Plant. Regist. 2011, 5, 27–39. [Google Scholar] [CrossRef]
- Edmisten, K.; Collins, G.; Crozier, C.; York, A.; Hardy, D.; Reisig, D.; Bullen, G.; Thiessen, L.; Vann, R.; Cahoon, C.; et al. North Carolina Cotton Production Guide; North Carolina Cooperative Extension Service Pub AG-417: Raleigh, NC, USA, 2019. [Google Scholar]
- Heiniger, R.; Spears, J.; Bowman, D.; Carson, M.L.; Crozier, C.; Dunphy, J.; Koenning, S.; Marra, M.; Naderman, G.C.; Van Duyn, J.; et al. Corn Production Guide; North Carolina Cooperative Extension Service Pub AG-590: Raleigh, NC, USA, 2019. [Google Scholar]
- Jordan, D.L.; Brandenburg, R.; Brown, B.; Bullen, G.; Roberson, G.; Shew, B. 2019 Peanut Information; North Carolina Cooperative Extension Service Pub. AG-331: Raleigh, NC, USA, 2019. [Google Scholar]
- Stowe, K.D.; Crozier, C.; Bullen, G.; Dunphy, E.J.; Everman, W.; Hardy, D.; Osmond, D.; Piggot, N.; Randa, S.; Reisig, D.; et al. North Carolina Soybean Production Guide; North Carolina Cooperative Extension Service Pub AG-835: Raleigh, NC, USA, 2017. [Google Scholar]
- Sosnoskie, L.M.; Webster, T.M.; Grey, T.L.; Culpepper, A.S. Severed stems of Amaranthus palmeri are capable of regrowth and seed production in Gossypium hirsutum. Ann. Appl. Biol. 2014, 165, 147–154. [Google Scholar] [CrossRef]
- Archontoulis, S.V.; Miguez, F.E. Nonlinear regression models and applications in agricultural research. Agron. J. 2015, 107, 786–798. [Google Scholar] [CrossRef] [Green Version]
- USDA-NASS. 2020 State Agriculture Overview—North Carolina: Crops—Planted, Harvested, Yield, Production, Price (MYA), Value of Production. Available online: https://www.nass.usda.gov/Quick_Stats/Ag_Overview/ (accessed on 20 May 2021).
- Schwartz, L.M.; Norsworthy, J.K.; Young, B.G.; Bradley, K.W.; Kruger, G.R.; Davis, V.M.; Steckel, L.E.; Walsh, M.J. Tall waterhemp (Amaranthus tuberculatus) and Palmer amaranth (Amaranthus palmeri) seed production and retention at soybean maturity. Weed Technol. 2016, 30, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Bensch, C.N.; Horak, M.J.; Peterson, D. Interference of redroot pigweed (Amaranthus retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A. rudis) in soybean. Weed Sci. 2003, 51, 37–43. [Google Scholar] [CrossRef]
- Massinga, R.A.; Currie, R.S.; Horak, M.J.; Boyer, J. Interference of Palmer amaranth in corn. Weed Sci. 2001, 49, 202–208. [Google Scholar] [CrossRef]
- Bravo, W.; Leon, R.G.; Ferrell, J.A.; Mulvaney, M.J.; Wood, C.W. Differentiation of life-history traits among Palmer amaranth populations (Amaranthus palmeri) and its relation to cropping systems and glyphosate sensitivity. Weed Sci. 2017, 65, 339–349. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Griffith, G.; Griffin, T.; Bagavathiannan, M.; Gbur, E.E. In-field movement of glyphosate-resistant Palmer amaranth (Amaranthus palmeri) and its impact on cotton lint yield: Evidence supporting a zero-tolerance threshold strategy. Weed Sci. 2014, 62, 237–249. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop | Gender | bc | cc | dc | GDD50 | RMSE d | MEF e |
|---|---|---|---|---|---|---|---|
| ____________cm____________ | |||||||
| _____________________________________Cohort 1_____________________________________ | |||||||
| Corn | Female | 4.0 ± 0.3 | 2.3 ± 4.4 | 241.6 ± 6.5 | 653.4 ± 16.8 | 11.4 | 0.98 |
| Male | 3.9 ± 0.3 | 2.3 ± 3.1 | 217.4 ± 4.7 | 648.7 ± 13.4 | 8.0 | 0.99 | |
| Cotton | Female | 4.1 ± 0.3 | 2.3 ± 3.2 | 211.6 ± 5.0 | 672.5 ± 14.6 | 8.5 | 0.99 |
| Male | 4.7 ± 0.7 | 3.0 ± 5.8 | 199.9 ± 7.6 | 642.9 ± 23.5 | 16.1 | 0.96 | |
| Peanut | Female | 3.7 ± 0.3 | 2.4 ± 3.1 | 193.8 ± 6.0 | 721.0 ± 19.2 | 8.2 | 0.99 |
| Male | 4.1 ± 0.2 | 2.5 ± 1.5 | 174.4 ± 2.4 | 671.8 ± 8.4 | 4.0 | 1.00 | |
| Soybean | Female | 3.8 ± 0.5 | 3.2 ± 4.4 | 189.7 ± 8.0 | 705.8 ± 26.0 | 11.5 | 0.97 |
| Male | 3.8 ± 0.2 | 2.5 ± 2.0 | 176.1 ± 3.2 | 668.3 ± 11.5 | 5.2 | 0.99 | |
| _____________________________________Cohort 2_____________________________________ | |||||||
| Corn | Female | 2.7 ± 0.5 | 0.1 ± 1.3 | 30.8 ± 2.5 | 628.8 ± 56.3 | 2.1 | 0.95 |
| Male | 2.5 ± 2.2 | 0.4 ± 6.1 | 43.4 ± 21.4 | 798.4 ± 396.3 | 10.3 | 0.56 | |
| Cotton | Female | 4.4 ± 0.4 | 2.8 ± 3.6 | 166.4 ± 4.0 | 637.4 ± 16.0 | 6.7 | 0.99 |
| Male | 4.7 ± 0.8 | 2.5 ± 5.2 | 146.2 ± 6.0 | 674.2 ± 27.0 | 9.9 | 0.96 | |
| Peanut | Female | 3.9 ± 0.8 | 2.3 ± 6.3 | 148.2 ± 9.2 | 699.8 ± 40.6 | 11.5 | 0.95 |
| Male | 4.0 ± 0.9 | 2.4 ± 6.8 | 168.0 ± 12.4 | 765.8 ± 47.5 | 13.0 | 0.95 | |
| Soybean | Female | 3.9 ± 0.8 | 0.9 ± 5.7 | 134.1 ± 7.5 | 668.1 ± 36.7 | 10.2 | 0.95 |
| Male | 5.5 ± 3.2 | 1.7 ± 12.0 | 110.7 ± 13.4 | 690.0 ± 77.4 | 24.8 | 0.72 | |
| Crop | Gender | Bc | Cc | Dc | GDD50 | RMSE d | MEF e |
|---|---|---|---|---|---|---|---|
| ____________cm____________ | |||||||
| Corn | Female | 3.9 ± 1.7 | 0.1 ± 3.3 | 41.1 ± 2.1 | 335.8 ± 32.1 | 5.8 | 0.83 |
| Male | 4.4 ± 1.9 | 0.1 ± 2.4 | 37.2 ± 1.4 | 314.7 ± 26.6 | 4.2 | 0.88 | |
| Cotton | Female | 3.0 ± 0.4 | −0.5 ± 4.6 | 125.7 ± 6.0 | 564.9 ± 31.3 | 8.3 | 0.96 |
| Male | 3.9 ± 0.7 | 0.7 ± 4.8 | 96.4 ± 4.0 | 498.8 ± 27.9 | 8.6 | 0.93 | |
| Peanut | Female | 2.6 ± 0.5 | −0.1 ± 5.3 | 110.3 ± 9.2 | 602.2 ± 56.8 | 9.4 | 0.92 |
| Male | 3.3 ± 0.5 | 0.2 ± 3.9 | 89.1 ± 4.0 | 514.3 ± 29.7 | 7.0 | 0.94 | |
| Soybean | Female | 2.2 ± 0.6 | 0.1 ± 3.8 | 67.7 ± 7.0 | 512.9 ± 67.3 | 6.8 | 0.89 |
| Male | 3.6 ± 0.8 | 0.2 ± 2.4 | 46.6 ± 1.7 | 377.9 ± 22.0 | 4.2 | 0.92 | |
| Crop | Gender | y0 b | a b | B b | RMSE c | MEF d |
|---|---|---|---|---|---|---|
| ____________________________cm____________________________ | ||||||
| Corn | Female | −0.28 ± 0.990 | 0.03 ± 0.002 | −1.45 × 10−5 ± 1.71 × 10−6 | 1.8 | 0.76 |
| Male | −0.02 ± 1.432 | 0.02 ± 0.004 | −1.25 × 10−5 ± 2.48 × 10−6 | 2.6 | 0.53 | |
| Cotton | Female | −1.77 ± 3.171 | 0.08 ± 0.009 | −4.23 × 10−5 ± 5.49 × 10−6 | 5.8 | 0.78 |
| Male | −1.77 ± 1.799 | 0.07 ± 0.005 | −3.70 × 10−5 ± 3.1 × 10−6 | 3.3 | 0.86 | |
| Peanut | Female | −1.97 ± 4.119 | 0.07 ± 0.012 | −2.70 × 10−5 ± 7.13 × 10−6 | 7.5 | 0.71 |
| Male | −3.40 ± 4.278 | 0.08 ± 0.012 | −3.45 × 10−5 ± 7.41 × 10−6 | 5.7 | 0.90 | |
| Soybean | Female | −0.12 ± 1.933 | 0.05 ± 0.006 | −2.62 × 10−5 ± 3.35 × 10−6 | 3.5 | 0.79 |
| Male | −2.29 ± 2.790 | 0.05 ± 0.008 | −2.76 × 10−5 ± 4.83 × 10−6 | 5.1 | 0.61 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahoney, D.J.; Jordan, D.L.; Hare, A.T.; Leon, R.G.; Roma-Burgos, N.; Vann, M.C.; Jennings, K.M.; Everman, W.J.; Cahoon, C.W. Palmer Amaranth (Amaranthus palmeri) Growth and Seed Production When in Competition with Peanut and Other Crops in North Carolina. Agronomy 2021, 11, 1734. https://doi.org/10.3390/agronomy11091734
Mahoney DJ, Jordan DL, Hare AT, Leon RG, Roma-Burgos N, Vann MC, Jennings KM, Everman WJ, Cahoon CW. Palmer Amaranth (Amaranthus palmeri) Growth and Seed Production When in Competition with Peanut and Other Crops in North Carolina. Agronomy. 2021; 11(9):1734. https://doi.org/10.3390/agronomy11091734
Chicago/Turabian StyleMahoney, Denis J., David L. Jordan, Andrew T. Hare, Ramon G. Leon, Nilda Roma-Burgos, Matthew C. Vann, Katherine M. Jennings, Wesley J. Everman, and Charles W. Cahoon. 2021. "Palmer Amaranth (Amaranthus palmeri) Growth and Seed Production When in Competition with Peanut and Other Crops in North Carolina" Agronomy 11, no. 9: 1734. https://doi.org/10.3390/agronomy11091734