
Root Zone Management for Improving Seedling Quality of Organically Produced Horticultural Crops

 ,
,  , , , , and
, , , , and 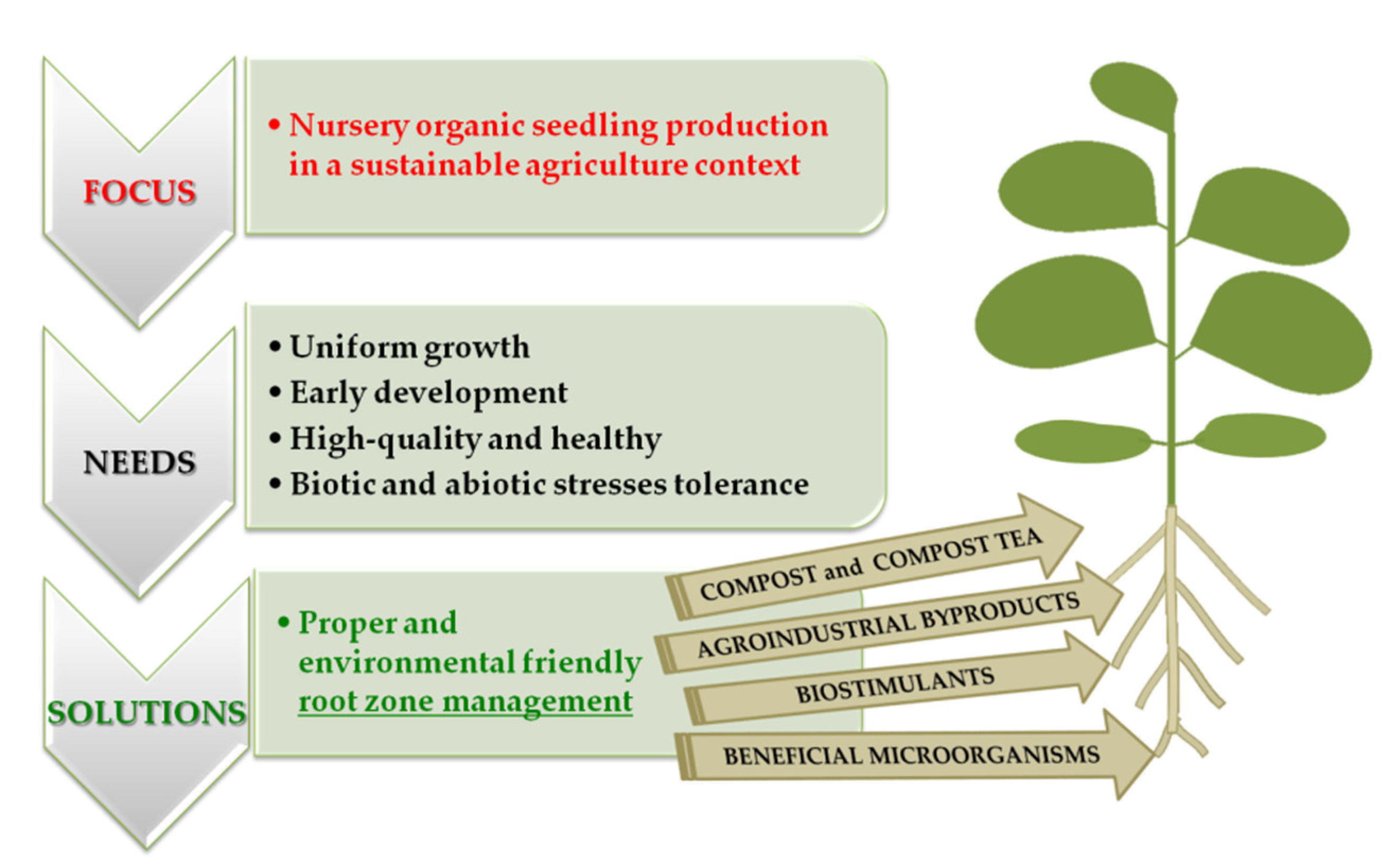{kind=link}
Abstract
:1. Introduction
2. Compost and Compost Tea to Produce Seedling in Nursery
3. Valorization of Agroindustrial Byproducts in Innovative Organic Fertilizers and Growing Media Suitable for Seedling Production
4. Implications of Microbial and Non-Microbial Biostimulants for Seedling Quality
5. Beneficial Microorganisms for Biological Control of Seedling Diseases
6. Conclusions
7. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Greene, C.; Ferreira, G.; Carlson, A.; Cooke, B.; Hitaj, C. Growing organic demand provides high-value opportunities for many types of producers. In Amber Waves; United States Department of Agriculture Economic Research Service: Washington, DC, USA, 2017. Available online: https://www.ers.usda.gov/amber-waves/2017/januaryfebruary/growing-organic-demand-provides-high-value-opportunities-for-many-types-of-producers/ (accessed on 27 January 2021).
- Narayan, R.; Narayan, S. Precision farming in vegetables. In Precision Farming: A New Approach; Ram, T., Lohan, S.K., Singh, R., Singh, P., Eds.; Daya Publishing House: Delhi, India, 2014; p. 383. [Google Scholar]
- Izquierdo, J. Biotechnology can help crop production to feed an increasing world population-positive and negative aspects need to be balanced: A perspective from FAO. In Plant Genetic Engineering: Towards the Third Millennium, Proceedings of the International Symposium on Plant Genetic Engineering, Havana, Cuba, 6–8 May 2020; FAO Regional Office for Latin America and the Caribbean: Santiago, Chile, 2020; Volumes 6–10, pp. 13–26. [Google Scholar]
- Grossnickle, S.C.; MacDonald, J.E. Why seedlings grow: Influence of plant attributes. New For. 2018, 49, 1–34. [Google Scholar] [CrossRef]
- Minami, K. High quality of seedling in vegetable production. In Proceedings of the IX International Symposium on Timing of Field Production in Vegetable Crops, Piracicaba, Sao Paulo, Brazil, 20–24 May 2001. [Google Scholar]
- Grossnickle, S.C. Importance of root growth in overcoming planting stress. New For. 2005, 30, 273–294. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; Kiiskila, S.B.; Haase, D.L. Seedling ecophysiology: Five questions to explore in the nursery for optimizing subsequent field success. Tree Plant Notes 2020, 63, 112–127. [Google Scholar]
- Landis, T.D. Root culturing in bareroot nurseries. For. Nurs. Notes 2008, 30, 9–15. [Google Scholar]
- Bayoumi, Y.A.; El-Henawy, A.S.; Abdelaal, K.A.A.; Elhawat, N. Grape fruit waste compost as a nursery substrate ingredient for high-quality cucumber (Cucumis sativus L.) seedlings production. Compost Sci. Util. 2019, 27, 205–216. [Google Scholar] [CrossRef]
- Pascual, J.A.; Ceglie, F.; Tuzel, Y.; Koller, M.; Koren, A.; Hitchings, R.; Tittarelli, F. Organic substrate for transplant production in organic nurseries. A review. Agron. Sustain. Dev. 2018, 38, 35. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Dai, J.; Zhang, Y.; Wang, X.; Zhu, W.; Yuan, X.; Yuan, H.; Cui, Z. Composted biogas residue and spent mushroom substrate as a growth medium for tomato and pepper seedlings. J. Environ. Manag. 2018, 216, 62–69. [Google Scholar] [CrossRef]
- Ronga, D.; Pane, C.; Zaccardelli, M.; Pecchioni, N. Use of spent coffee ground compost in peat-based growing media for the production of basil and tomato potting plants. Commun. Soil Sci. Plant. 2016, 47, 356–368. [Google Scholar] [CrossRef]
- Stewart-Wade, S.M. Efficacy of organic amendments used in containerized plant production: Part 1—Compost-based amendments. Sci. Hortic. 2020, 266, 108856. [Google Scholar] [CrossRef]
- Carmona, E.; Moreno, M.T.; Avilés, M.; Ordovás, J. Use of grape marc compost as substrate for vegetable seedlings. Sci. Hortic. 2012, 137, 69–74. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Stamatakis, A.; Moustakas, K.; Prasad, M.; Tzortzakis, N. Evaluation of municipal solid waste compost and/or fertigation as peat substituent for pepper seedlings production. Waste Biomass Valorization 2018, 9, 2285–2294. [Google Scholar] [CrossRef]
- Owen, W.G.; Jackson, B.E.; Whipker, B.E.; Fonteno, W.C.; Benson, M.D. Assessing the severity of damping-off caused by Pythium ultimum and Rhizoctonia solani in peat-based greenhouse substrates amended with pine wood chip aggregates. Acta Hortic. 2019, 1266, 27–34. [Google Scholar] [CrossRef]
- Kanaan, H.; Hadar, Y.; Medina, S.; Krasnovsky, A.; Mordechai-Lebiush, S.; Tsror, L. Effect of compost properties on progress rate of Verticillium dahliae attack on eggplant (Solanum melongena L.). Compost Sci. Util. 2018, 26, 71–78. [Google Scholar] [CrossRef]
- Shi, L.; Du, N.; Yuan, Y.; Shu, S.; Sun, J.; Guo, S. Vinegar residue compost as a growth substrate enhances cucumber resistance against the Fusarium wilt pathogen Fusarium oxysporum by regulating physiological and biochemical responses. Environ. Sci. Pollut. Res. 2016, 23, 18277–18287. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.B.; Ros, M.; Ayuso, L.M.; Bustamante, M.; de Los, A.; Moral, R.; Pascual, J.A. Agroindustrial composts to reduce the use of peat and fungicides in the cultivation of muskmelon seedlings. J. Sci. Food Agric. 2017, 97, 875–881. [Google Scholar] [CrossRef]
- Pane, C.; Piccolo, A.; Spaccini, R.; Celano, G.; Villecco, D.; Zaccardelli, M. Agricultural waste-based composts exhibiting suppressivity to diseases caused by the phytopathogenic soil-borne fungi Rhizoctonia solani and Sclerotinia minor. Appl. Soil Ecol. 2013, 65, 43–51. [Google Scholar] [CrossRef]
- Diab, H.G.; Hu, S.; Benson, D.M. Suppression of Rhizoctonia solani on impatiens by enhanced microbial activity in composted swine waste-amended potting mixes. Phytopathology 2003, 93, 1115–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pane, C.; Spaccini, R.; Piccolo, A.; Scala, F.; Bonanomi, G. Compost amendments enhance peat suppressiveness to Pythium ultimum, Rhizoctonia solani and Sclerotinia minor. Biol. Control 2011, 56, 115–124. [Google Scholar] [CrossRef]
- Segarra, G.; Santpere, G.; Elena, G.; Trillas, I. Enhanced Botrytis cinerea resistance of Arabidopsis plants grown in compost may be explained by increased expression of defense-related genes, as revealed by microarray analysis. PLoS ONE 2013, 8, e56075. [Google Scholar] [CrossRef] [Green Version]
- Blaya, J.; Lloret, E.; Ros, M.; Pascual, J.A. Identification of predictor parameters to determine agro-industrial compost suppressiveness against Fusarium oxysporum and Phytophthora capsici diseases in muskmelon and pepper seedlings. J. Sci. Food Agric. 2015, 95, 1482–1490. [Google Scholar] [CrossRef]
- Pane, C.; Sorrentino, R.; Scotti, R.; Molisso, M.; Di Matteo, A.; Celano, G.; Zaccardelli, M. Alpha and beta-diversity of microbial communities associated to plant disease suppressive functions of on-farm green composts. Agriculture 2020, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- Scotti, R.; Mitchell, A.L.; Pane, C.; Finn, R.D.; Zaccardelli, M. Microbiota characterization of agricultural green waste-based suppressive composts using omics and classic approaches. Agriculture 2020, 10, 61. [Google Scholar] [CrossRef] [Green Version]
- De Corato, U.; Salimbeni, R.; De Pretis, A. Suppression of soil-borne pathogens in container media amended with on-farm composted agro-bioenergy wastes and residues under glasshouse condition. J. Plant Dis. Prot. 2018, 125, 213–226. [Google Scholar] [CrossRef]
- Pascual, J.A.; Bernal-Vicente, A.; Martinez-Medina, A.; Ros, M.; Sánchez, C. Biostimulant and suppressive effect of Trichoderma harzianum enriched compost for melon cultivation from greenhouse nursery to field production. Acta Hortic. 2017, 1164, 225–232. [Google Scholar] [CrossRef]
- Oka, Y.; Yermiyahu, U. Suppressive effects of composts against the root-knot nematode Meloidogyne javanica on tomato. Nematology 2002, 4, 891–898. [Google Scholar] [CrossRef]
- Pane, C.; Spaccini, R.; Piccolo, A.; Celano, G.; Zaccardelli, M. Disease suppressiveness of agricultural greenwaste composts as related to chemical and bio-based properties shaped by different on-farm composting methods. Biol. Control 2019, 137, 104026. [Google Scholar] [CrossRef]
- Markakis, E.A.; Fountoulakis, M.S.; Daskalakis, G.C.; Kokkinis, M.; Ligoxigakis, E.K. The suppressive effect of compost amendments on Fusarium oxysporum f. sp. radicis-cucumerinum in cucumber and Verticillium dahliae in eggplant. Crop Prot. 2016, 79, 70–79. [Google Scholar]
- Abouelenien, F.; Fujiwara, W.; Namba, Y.; Kosseva, M.; Nishio, N.; Nakashimada, Y. Improved methane fermentation of chicken manure via ammonia removal by biogas recycle. Bioresour. Technol. 2010, 101, 6368–6373. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Jacobi, H.F.; Strach, K.; Xu, C.; Zhou, H.; Liebetrau, J. Mono-fermentation of chicken manure: Ammonia inhibition and recirculation of the digestate. Bioresour. Technol. 2015, 178, 238–246. [Google Scholar] [CrossRef]
- Bortolini, S.; Macavei, L.I.; Saadoun, J.H.; Foca, G.; Ulrici, A.; Bernini, F.; Malferrari, D.; Setti, L.; Ronga, D.; Maistrello, L. Hermetia illucens (L.) larvae as chicken manure management tool for circular economy. J. Clean. Prod. 2020, 262, 121289. [Google Scholar] [CrossRef]
- Setti, L.; Francia, E.; Pulvirenti, A.; Gigliano, S.; Zaccardelli, M.; Pane, C.; Caradonia, F.; Bortolini, S.; Maistrello, L.; Ronga, D. Use of black soldier fly (Hermetia illucens (L.), Diptera: Stratiomyidae) larvae processing residue in peat-based growing media. Waste Manag. 2019, 95, 278–288. [Google Scholar] [CrossRef]
- Ronga, D.; Mantovi, P.; Pacchioli, M.T.; Pulvirenti, A.; Bigi, F.; Allesina, G.; Pedrazzi, S.; Dal Prà, A. Combined effects of dewatering, composting and pelleting to valorize and delocalize livestock manure, improving agricultural sustainability. Agronomy 2020, 10, 661. [Google Scholar] [CrossRef]
- Pane, C.; Celano, G.; Villecco, D.; Zaccardelli, M. Control of Botrytis cinerea, Alternaria alternata and Pyrenochaeta lycopersici on tomato with whey compost-tea application. Crop Prot. 2012, 38, 80–86. [Google Scholar] [CrossRef]
- Lanthier, M. Compost tea and its impact on plant diseases. BC Org. Grow. 2007, 10, 7–11. [Google Scholar]
- Scheuerell, S.J.; Mahaffee, W.F. Compost tea: Principles and prospect for plant disease control. Compost Sci. Util. 2002, 10, 313–338. [Google Scholar] [CrossRef]
- Ingham, E.R. What is compost tea? Part 1. BioCycle 1999, 40, 74–75. [Google Scholar]
- Mengesha, W.K.; Powel, S.M.; Evans, K.J.; Barry, K.M. Diverse microbial communities in non-aerated compost teas suppress bacterial wilt. World J. Microbiol. Biotechnol. 2017, 33, 49. [Google Scholar] [CrossRef]
- Pane, C.; Celano, G.; Zaccardelli, M. Metabolic patterns of bacterial communities in aerobic compost teas associated with potential biocontrol of soilborne plant diseases. Phytopathol. Mediterr. 2014, 53, 277–286. [Google Scholar]
- Pane, C.; Palese, A.M.; Celano, G.; Zaccardelli, M. Effects of compost tea treatments on productivity of lettuce and kohlrabi systems under organic cropping management. Ital. J. Agron. 2014, 9, 596. [Google Scholar] [CrossRef] [Green Version]
- Pane, C.; Palese, A.M.; Spaccini, R.; Piccolo, A.; Celano, G.; Zaccardelli, M. Enhancing sustainability of a processing tomato cultivation system by using bioactive compost teas. Sci. Hortic. 2016, 202, 117–124. [Google Scholar] [CrossRef]
- Scheuerell, S.J.; Mahaffee, W.F. Compost tea as a container medium drench for suppressing seedling damping-off by Pythium ultimum. Phytopathology 2004, 94, 1156–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Mughrabi, K.I.; Berthélémé, C.; Livingston, T.; Burgoyne, A.; Poirier, R.; Vikram, A. Aerobic compost tea, compost and a combination of both reduce the severity of common scab (Streptomyces scabiei) on potato tubers. J. Plant Sci. 2008, 3, 168–175. [Google Scholar] [CrossRef]
- El-Masry, M.H.; Khalil, A.I.; Hassouna, M.S.; Ibrahim, H.A.H. In situ and in vitro suppressive effect of agricultural composts and their water extracts on some phytopathogenic fungi. World J. Microbiol. Biotechnol. 2002, 18, 551–558. [Google Scholar] [CrossRef]
- Zhang, W.; Han, D.J.; Dick, W.A.; Davis, K.R.; Hoitink, H.A.J. Compost and compost water extract—Induced systemic acquired in cucumber and Arabidopsis. Phytopathology 1998, 88, 450–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koné, S.B.; Dionne, A.; Tweddel, R.J.; Antoun, H.; Avis, T.J. Suppressive effect of non-aerated compost teas on foliar fungal pathogens of tomato. Biol. Control 2010, 52, 167–173. [Google Scholar] [CrossRef]
- Al-Dahmani, J.H.; Abbassi, P.A.; Miller, S.A.; Hoitink, H.A.J. Suppression of bacterial spot of tomato with foliar sprays of compost extracts under greenhouse and field conditions. Plant Dis. 2003, 87, 913–919. [Google Scholar] [CrossRef] [Green Version]
- Edwards, C.A.; Arancon, N.Q.; Greytak, S. Effects of vermicompost teas on plant growth and disease. Biocycle 2006, 47, 28–31. [Google Scholar]
- Elad, Y.; Shtienberg, D. Effect of compost water extracts on grey mould (Botrytis cinerea). Crop Prot. 1994, 13, 109–114. [Google Scholar] [CrossRef]
- Liguori, L.; Pane, C.; Albanese, D.; Celano, G.; Zaccardelli, M.; Di Matteo, M. Compost and compost tea management of mini watermelon cultivations affect the chemical, physical and sensory assessment of the fruits. Agric. Sci. 2015, 6, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Pant, A.; Radovich, T.J.K.; Hue, N.V.; Talcott, S.T.; Krenek, K.A. Vermicompost extracts influence growth, mineral nutrients, phytonutrients and antioxidant activity in Pak choi (Brassica rapa cv. Bonsai Chinensis group) grown under vermicompost and chemical fertilizer. J. Sci. Food Agric. 2009, 89, 2383–2392. [Google Scholar] [CrossRef]
- Pant, A.; Radovich, T.J.K.; Hue, N.V.; Arancon, N.Q. Effects of vermicompost tea (aqueous extract) on pak-choi yield, quality, and on soil biological properties. Compost Sci. Util. 2011, 19, 279–292. [Google Scholar] [CrossRef]
- Ronga, D.; Caradonia, F.; Setti, L.; Hagassou, D.; Giaretta Azevedo, C.V.; Milc, J.; Pedrazzi, S.; Allesina, G.; Arru, L.; Francia, E. Effects of innovative biofertilisers on yield of processing tomato cultivated in organic cropping systems in northern Italy. Acta Hortic. 2019, 1233, 129–136. [Google Scholar] [CrossRef]
- Welke, S.E. The effect of compost extract on the yield of strawberries and the severity of Botrytis cinerea. J. Sustain. Agric. 2005, 25, 57–68. [Google Scholar] [CrossRef]
- Zaccardelli, M.; Pane, C.; Villecco, D.; Palese, A.M.; Celano, G. Compost tea spraying increases yield performance of pepper (Capsicum annuum L.) grown in greenhouse under organic farming system. Ital. J. Agron. 2018, 13, 229–234. [Google Scholar] [CrossRef]
- Bernal-Vincente, A.; Ros, M.; Tittarelli, F.; Intrigliolo, F.; Pascual, J.A. Citrus compost and its water extract for cultivation of melon plants in green house nurseries. Evaluation of nutriactive and biocontrol effects. Bioresour. Technol. 2008, 99, 8722–8728. [Google Scholar] [CrossRef]
- Siddiqui, Y.; Sariah, M.; Razi, I.; Mawardi, R.; Asgar, A. Bio-efficiency of compost extracts on the wet rot incidence, morphological and physiological growth of okra (Abelmoschus esculentus [(L.) Moench]). Sci. Hortic. 2008, 117, 9–14. [Google Scholar] [CrossRef]
- Siddiqui, Y.; Sariah, M.; Razi, I.; Mawardi, R. Bio-potential of compost tea from agro-waste to suppress Choanephora cucurbitarum L. the causal pathogen of wet rok of okra. Biol. Control 2009, 49, 38–44. [Google Scholar] [CrossRef]
- Ekabote, S.D.; Divyajyothi, U.; Narayanaswamy, P.; Ravindra, H. Evaluation of propamocarb 530 + fosetyl 310–840 SL for the management of damping off of vegetables. J. Pharmacogn. Phytochem. 2019, 8, 1616–1618. [Google Scholar]
- Gilardi, G.; Demarchi, S.; Gullino, M.L.; Garibaldi, A. Nursery treatments with non-conventional products against crown and root rot, caused by Phytophthora capsici, on zucchini. Phytoparasitica 2015, 43, 501–508. [Google Scholar] [CrossRef]
- Villecco, D.; Pane, C.; Ronga, D.; Zaccardelli, M. Enhancing sustainability of tomato, pepper and melon nursery production systems by using compost tea spray applications. Agronomy 2020, 10, 1336. [Google Scholar] [CrossRef]
- Guo, Z.; Han, J.; Li, J.; Xu, Y.; Wang, X. Effects of long-term fertilization on soil organic carbon mineralization and microbial community structure. PLoS ONE 2019, 14, e0211163. [Google Scholar]
- Chen, J.; Lü, S.; Zhang, Z.; Zhao, X.; Li, X.; Ning, P.; Liu, M. Environmentally friendly fertilizers: A review of materials used and their effects on the environment. Sci. Total Environ. 2018, 613–614, 829–839. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Communication from The Commission to the European Parliament, The Council, The European Economic and Social Committee and The Committee of the Regions Closing the Loop—An EU Action Plan for the Circular Economy. 2015. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A52015DC0614 (accessed on 21 January 2021).
- European Commission. A Sustainable Bioeconomy for Europe: Strengthening the Connection between Economy, Society and the Environment. Updated Bioeconomy Strategy. 2018. Available online: https://op.europa.eu:443/en/publication-detail/-/publication/edace3e3-e189-11e8-b690-01aa75ed71a1/language-en (accessed on 21 January 2021).
- Regulation (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 Laying Down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003) Regulations. Available online: https://eur-lex.europa.eu/eli/reg/2019/1009/oj (accessed on 21 January 2021).
- Abbasi, T.; Tauseef, S.M.; Abbasi, S.A. Anaerobic digestion for global warming control and energy generation—An overview. Renew. Sustain. Energy Rev. 2012, 16, 3228–3242. [Google Scholar] [CrossRef]
- Monnet, F. An Introduction to Anaerobic Digestion of Organic Wastes. A Report. Remade Scotland. 2003. Available online: https://www.cti2000.it/Bionett/BioG-2003-002%20IntroAnaerobicDigestion.pdf (accessed on 27 January 2021).
- Nkoa, R. Agricultural benefits and environmental risks of soil fertilization with anaerobic digestates: A review. Agron. Sustain. Dev. 2014, 34, 473–492. [Google Scholar] [CrossRef] [Green Version]
- Aso, S.N. Digestate: The Coproduct of Biofuel Production in a Circular Economy, and New Results for Cassava Peeling Residue Digestate; Intech Open: London, UK, 2020; Available online: https://www.intechopen.com/online-first/digestate-the-coproduct-of-biofuel-production-in-a-circular-economy-and-new-results-for-cassava-peel (accessed on 27 January 2021).
- Jorobekova, S.; Kydralieva, K. Plant growth biostimulants from byproducts of anaerobic digestion of organic substances. In Organic Fertilizers—History, Production and Application; Larramendy, M., Solonesli, S., Eds.; Intech Open: London, UK, 2019; Available online: https://www.intechopen.com/books/organic-fertilizers-history-production-and-applications/plant-growth-biostimulants-from-by-products-of-anaerobic-digestion-of-organic-substances (accessed on 27 January 2021).
- Yagüe, M.R.; Lobo, M.C. Liquid digestate from organic residues as fertilizer: Carbon fractions, phytotoxicity and microbiological analysis. SJSS 2020, 10, 248–256. [Google Scholar]
- Lencioni, G.; Imperiale, D.; Cavirani, N.; Marmiroli, N.; Marmiroli, M. Environmental application and phytotoxicity of anaerobic digestate from pig farming by in vitro and in vivo trials. Int. J. Environ. Sci. Technol. 2016, 13, 2549–2560. [Google Scholar] [CrossRef]
- Ronga, D.; Setti, L.; Salvarani, C.; De Leo, R.; Bedin, E.; Pulvirenti, A.; Milc, J.; Pecchioni, N.; Francia, E. Effects of solid and liquid digestate for hydroponic baby leaf lettuce (Lactuca sativa L.) cultivation. Sci. Hortic. 2019, 244, 172–181. [Google Scholar] [CrossRef]
- Ronga, D.; Pellati, F.; Brighenti, V.; Laudicella, K.; Laviano, L.; Fedailaine, M.; Benvenuti, S.; Pecchioni, N.; Francia, E. Testing the influence of digestate from biogas on growth and volatile compounds of basil (Ocimum basilicum L.) and peppermint (Mentha x piperita L.) in hydroponics. J. Appl. Res. Med. Aroma 2018, 11, 18–26. [Google Scholar] [CrossRef]
- Greco, C.; Comparetti, A.; Fascella, G.; Febo, P.; La Placa, G.; Saiano, F.; Mammano, M.M.; Orlando, S.; Laudicina, V.A. Effects of vermicompost, compost and digestate as commercial alternative peat-based substrates on qualitative parameters of Salvia officinalis. Agronomy 2021, 11, 98. [Google Scholar] [CrossRef]
- Zeng, Y.; De Guardia, A.; Dabert, P. Improving composting as a post-treatment of anaerobic digestate. Bioresour. Technol. 2016, 201, 293–303. [Google Scholar] [CrossRef]
- Vitti, A.; Elshafie, H.S.; Logozzo, G.; Marzario, S.; Scopa, A.; Camele, I.; Nuzzaci, M. Physico-chemical characterization and biological activities of a digestate and a more stabilized digestate-derived compost from agro-waste. Plants 2021, 10, 386. [Google Scholar] [CrossRef]
- Ahuja, I.; Dauksas, E.; Remme, J.F.; Richardsen, R.; Løes, A.-K. Fish and fish waste-based fertilizers in organic farming—With status in Norway: A review. Waste Manag. 2020, 115, 95–112. [Google Scholar] [CrossRef]
- Busato, J.G.; de Carvalho, C.M.; Zandonadi, D.B.; Sodré, F.F.; Mol, A.R.; de Oliveira, A.L.; Navarro, R.D. Recycling of wastes from fish beneficiation by composting: Chemical characteristics of the compost and efficiency of their humic acids instimulating the growth of lettuce. Environ. Sci. Pollut. Res. 2018, 25, 35811–35820. [Google Scholar] [CrossRef] [PubMed]
- Billa, S.F.; Angwafo, T.E.; Ngome, A.F. Agro-environmental characterization of biochar issued from crop wastes in the humid forest zone of Cameroon. Int. J. Recycl. 2019, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Monterumici, C.; Rosso, D.; Montoneri, E.; Ginepro, M.; Baglieri, A.; Novotny, E.E.; Kwapinski, W.; Negre, M. Processed vs non-processed biowastes for agriculture: Effects of post-harvest tomato plants and biochar on radish growth, chlorophyll content and protein production. Int. J. Mol. Sci. 2015, 16, 8826–8843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalus, K.; Koziel, J.A.; Opaliński, S. A review of biochar properties and their utilization in crop agriculture and livestock production. Appl. Sci. 2019, 9, 3494. [Google Scholar] [CrossRef] [Green Version]
- Kaudal, B.B.; Chen, D.; Madhavan, D.B.; Downie, A.; Weatherley, A. An examination of physical and chemical properties of urban biochar for use as growing media substrate. Biomass Bioenergy 2016, 84, 49–58. [Google Scholar] [CrossRef]
- Kaur, T. Vermicomposting: An effective option for recycling organic wastes. In Organic Agriculture; Das, S.K., Ed.; IntechOpen: London, UK, 2020; p. 17. Available online: https://www.intechopen.com/books/organic-agriculture/vermicomposting-an-effective-option-for-recycling-organic-wastes (accessed on 27 January 2021).
- Atiyeh, R.M.; Subler, S.; Edwards, C.A.; Bachman, G.; Metzger, J.D. Effects of vermicomposts and composts on plant growth in horticultural container media and soil. Pedobiologia 2000, 44, 579–590. [Google Scholar] [CrossRef] [Green Version]
- Andreola, F.; Borghi, A.; Pedrazzi, S.; Allesina, G.; Tartarini, P.; Lancellotti, I.; Barbieri, L. Spent coffee grounds in the production of lightweight clay ceramic aggregates in view of urban and agricultural sustainable development. Materials 2019, 12, 3581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronga, D.; Parisi, M.; Barbieri, L.; Lancellotti, I.; Andreola, F.; Bignami, C. Valorization of spent coffee grounds, biochar and other residues to produce lightweight clay ceramic aggregates suitable for nursery grapevine production. Horticulturae 2020, 6, 58. [Google Scholar] [CrossRef]
- Coppens, J.; Grunert, O.; Van Den Hende, S.; Vanhoutte, I.; Boon, N.; Haesaert, G.; De Gelder, L. The use of microalgae as a high-value organic slow-release fertiliser results in tomatoes with increased carotenoid and sugar levels. J. Appl. Phycol. 2016, 28, 2367–2377. [Google Scholar] [CrossRef]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal biostimulants and biofertilisers in crop productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Faheed, F.A.; Fattah, Z.A. Effect of Chlorella vulgaris as biofertiliser on growth parameters and metabolic aspects of lettuce plant. J. Agric. Soc. Sci. 2008, 4, 165–169. [Google Scholar]
- Lv, J.; Liu, S.; Feng, J.; Liu, Q.; Guo, J.; Wang, L.; Xiaoyan, J.; Xie, S. Effects of microalgal biomass as biofertilizer on the growth of cucumber and microbial communities in the cucumber rhizosphere. Turk. J. Bot. 2020, 44, 167–177. [Google Scholar]
- Bumandalai, O.; Tserennadmid, R. Effect of Chlorella vulgaris as a biofertilizer on germination of tomato and cucumber seeds. Int. J. Aquat. Biol. 2019, 7, 95–99. [Google Scholar]
- Ciavatta, C.; Govi, M.; Sitti, L.; Gessa, C. Influence of blood meal organic fertilizer on soil organic matter: A laboratory study. J. Plant Nutr. 1997, 20, 1573–1591. [Google Scholar] [CrossRef]
- Bae, E.J.; Kim, B.W.; Kim, M.J.; Kwon, S.H.; Choi, J.H.; Na, H. Growth response of some vegetables seedlings according to blood fertilizer. J. Korean Soc. People Plant Environ. 2015, 18, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Spíchal, L.; Panzarová, K.; Casa, R.; Colla, G. High-throughput plant phenotyping for developing novel biostimulants: From lab to field or from field to lab? Front. Plant Sci. 2018, 9, 1197. [Google Scholar] [CrossRef]
- Rouphael, Y.; Lucini, L.; Miras-Moreno, B.; Colla, G.; Bonini, P.; Cardarelli, M. Metabolomic responses of maize shoots and roots elicited by combinatorial seed treatments with microbial and non-microbial biostimulants. Front. Microbiol. 2020, 11, 664. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Carillo, P.; Colla, G.; Fiorentino, N.; Sabatino, L.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cirillo, V.; Shabani, E.; et al. Appraisal of combined applications of Trichoderma virens and a biopolymer-based biostimulant on lettuce agronomical, physiological, and qualitative properties under variable N regimes. Agronomy 2020, 10, 196. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [Green Version]
- Goswami, M.; Deka, S. Plant growth-promoting rhizobacteria alleviators of abiotic stresses in soil—A review. Pedosphere 2020, 30, 40–61. [Google Scholar] [CrossRef]
- Woo, S.L.; Pepe, O. Microbial Consortia: Promising probiotics as plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1801. [Google Scholar] [CrossRef]
- Caradonia, F.; Ronga, D.; Flore, A.; Barbieri, R.; Moulin, L.; Terzi, V.; Francia, E. Biostimulants and cherry rootstock increased tomato fruit yield and quality in sustainable farming systems. Ital. J. Agron. 2020, 15, 1553. [Google Scholar]
- Bonini, P.; Cirino, V.; Reynaud, H.; Rouphael, Y.; Cardarelli, M.; Colla, G. Part 3: Innovation and practical applications—Designing and formulating microbial and non-microbial biostimulants. In Biostimulants for Sustainable Crop Production; Rouphael, Y., du Jardin, P., Brown, P., De Pascale, S., Colla, G., Eds.; Burleigh Dodds Science Publishing: Sawston, UK, 2019. [Google Scholar]
- Cardarelli, M.; Rouphael, Y.; Coppa, E.; Hoagland, L.; Colla, G. Using microgranular-based biostimulant in vegetable transplant production to enhance growth and nitrogen uptake. Agronomy 2020, 10, 842. [Google Scholar] [CrossRef]
- Ren, L.; Wang, B.; Yue, C.; Zhou, S.; Zhang, S.; Huo, H.; Xu, G. Mechanism of application nursery cultivation arbuscular mycorrhizal seedling in watermelon in the field. Ann. Appl. Biol. 2019, 174, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Bettoni, M.M.; Mogor, A.F.; Pauletti, V.; Goicoechea, N. Growth and metabolism of onion seedlings as affected by the application of humic substances, mycorrhizal inoculation and elevated CO2. Sci. Hortic. 2014, 180, 227–235. [Google Scholar] [CrossRef]
- Mohamed, H.A.; Barry, K.M.; Measham, P.F. The role of arbuscular mycorrhizal fungi in establishment and water balance of tomato seedlings and sweet cherry cuttings in low phosphorous soil. Acta Hortic. 2016, 1112, 109–115. [Google Scholar] [CrossRef]
- Ortas, I.; Sari, N.; Akpinar, C.; Yetsir, H. Screening mycorrhizae species for increased growth and P and Zn uptake in eggplant (Solanum melongena L.) grown under greenhouse conditions. Eur. J. Hortic. Sci. 2011, 76, 116–123. [Google Scholar]
- Sensoy, S.; Ocak, E.; Demir, S.; Tufenkci, S. Effects of humic acid, whey and arbuscular mycorrhizal fungi (AMF) applications on seedling growth and Fusarium wilt in Zucchini (Cucurbita pepo L.). J. Anim. Plant Sci. 2013, 23, 507–513. [Google Scholar]
- Ronga, D.; Caradonia, F.; Francia, E.; Morcia, C.; Rizza, F.; Badeck, F.R.; Ghizzoni, R.; Terzi, V. Interaction of tomato genotypes and arbuscular mycorrhizal fungi under reduced irrigation. Horticulturae 2019, 5, 79. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Chen, S.; Wang, M.; Liu, D.; Chang, R.; Wang, Z.; Lin, X.; Bai, B.; Ahammed, G.J. Arbuscular mycorrhizal fungus alleviates chilling stress by boosting redox poise and antioxidant potential of tomato seedlings. J. Plant Growth Regul. 2016, 35, 109–120. [Google Scholar] [CrossRef]
- Caradonia, F.; Francia, E.; Morcia, C.; Ghizzoni, R.; Moulin, L.; Terzi, V.; Ronga, D. Arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria avoid processing tomato leaf damage during chilling stress. Agronomy 2019, 9, 299. [Google Scholar] [CrossRef] [Green Version]
- Bidabadi, S.S.; Mehralian, M. Arbuscular mycorrhizal fungi inoculation to enhance chilling stress tolerance of watermelon. Gesunde Pflanz. 2020, 72, 171–179. [Google Scholar] [CrossRef]
- Turkmen, O.; Sensoy, S.; Demir, S.; Erdinc, C. Effects of two different AMF species on growth and nutrient content of pepper seedlings grown under moderate salt stress. Afr. J. Biotechnol. 2008, 7, 392–396. [Google Scholar]
- Ge, H.; Zhonghua, L. Characteristics of Azotobacter sp. strain AC11 and their effects on the growth of tomato seedlings under salt stress. Emir. J. Food Agric. 2019, 31, 520–525. [Google Scholar] [CrossRef]
- Mangmang, J.S.; Deaker, R.; Rogers, G. Early seedling growth response of lettuce, tomato and cucumber to Azospirillum brasilense inoculated by soaking and drenching. Hortic. Sci. 2015, 42, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Fasciglione, G.; Casanovas, E.M.; Yommi, A.; Sueldo, R.J.; Barassi, C.A. Azospirillum improves lettuce growth and transplant under saline conditions. Sci. Food Agric. 2012, 92, 2518–2523. [Google Scholar] [CrossRef]
- Cortés-Jiménez, D.; Gómez-Guzmán, A.; Iturriaga, G.; Suárez, R.; Alpírez, G.M.; Escalante, F.M.E. Microorganisms associated to tomato seedlings growing in saline culture act as osmoprotectant. Braz. J. Microbiol. 2014, 45, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Noel, T.C.; Sheng, C.; Yost, C.K.; Pharis, R.P.; Hynes, M.F. Rhizobium leguminosarum as a plant growth-promoting rhizobacterium: Direct growth promotion of canola and lettuce. Can. J. Microbiol. 1996, 42, 279–283. [Google Scholar] [CrossRef]
- Ferreira, L.D.V.M.; de Carvalho, F.; Andrade, J.F.C.; Oliveira, D.P.; De Medeiros, F.H.V.; Moreira, F.M.D.S. Co-inoculation of selected nodule endophytic rhizobacterial strains with Rhizobium tropici promotes plant growth and controls damping off in common bean. Pedosphere 2020, 30, 98–108. [Google Scholar] [CrossRef]
- Piccolo, A.; Celano, G.; Pietramellara, G. Effects of fractions of coal-derived humic substances on seed germination and growth of seedlings (Lactuga sativa and Lycopersicum esculentum). Biol. Fert. Soils 1993, 16, 11–15. [Google Scholar] [CrossRef]
- Lulakis, M.D.; Petsas, S.I. Effect of humic substances from vine-canes mature compost on tomato seedling growth. Bioresour. Technol. 1995, 54, 179–182. [Google Scholar] [CrossRef]
- Young, C.C.; Chen, L.F. Polyamines in humic acid and their effect on radical growth of lettuce seedlings. Plant Soil 1997, 195, 143–149. [Google Scholar] [CrossRef]
- Hartwigsen, J.A.; Evans, M.R. Humic acid seed and substrate treatments promote seedling root development. HortScience 2000, 35, 1231–1233. [Google Scholar] [CrossRef] [Green Version]
- Dursun, A.; Guvenx, I.; Turan, M. Effects of different levels of humic acids on seedling growth and macro and micronutrient contents of of tomato and eggplant. Acta Bot. 2002, 56, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Qin, K.; Leskovar, D.I. Humic substances improve vegetable seedling quality and post-transplant yield performance under stress conditions. Agriculture 2020, 10, 254. [Google Scholar] [CrossRef]
- Mattner, S.W.; Wite, D.; Riches, D.A.; Porter, I.J.; Arioli, T. The effect of kelp extract on seedling establishment of broccoli on contrasting soil types in southern Victoria, Australia. Biol. Agric. Hortic. 2013, 29, 258–270. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 2014, 26, 619–628. [Google Scholar] [CrossRef]
- Puglisi, I.; La Bella, E.; Rovetto, E.I.; Lo Piero, L.A.; Baglieri, A. Biostimulant effect and biochemical response in lettuce seedlings treated with a Scenedesmus quadricauda extract. Plants 2020, 9, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pascale, S.; Rouphael, Y.; Colla, G. Plant biostimulants: Innovative tool for enhancing plant nutrition in organic farming. Eur. J. Hortic. Sci. 2017, 82, 277–285. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G. A vegetal biopolymer-based biostimulant promoted root growth in melon while triggering brassinosteroids and stress-related compounds. Front. Plant Sci. 2018, 9, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Ku, K.M.; Choi, S.; Cardarelli, M. Vegetal-derived biostimulant enhances adventitious rooting in cuttings of basil, tomato, and Chrysanthemum via brassinosteroid-mediated processes. Agronomy 2019, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- Colman, S.L.; Salcedo, M.F.; Mansilla, A.Y.; Iglesias, M.J.; Fiol, D.F.; Martin-Saldana, S.; Alvarez, V.A.; Chevalier, A.A.; Casalongué, C.A. Chitosan microparticles improve tomato seedling biomass and modulate hormonal, redox and defense pathways. Plant Physiol. Biochem. 2019, 143, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Bu, R.; Xie, J.; Yu, J.; Liao, W.; Xiao, X.; Lv, J.; Wang, C.; Ye, J.; Calderón-Urrea, A. Autotoxicity in cucumber (Cucumis sativus L.) seedlings is alleviated by silicon through an increase in the activity of antioxidant enzymes and by mitigating lipid peroxidation. J. Plant Biol. 2016, 59, 247–259. [Google Scholar] [CrossRef]
- Aleandri, M.P.; Chilosi, G.; Bruni, N.; Tomassini, A.; Vettraino, A.M.; Vannini, A. Use of nursery potting mixes amended with local Trichoderma strains with multiple complementary mechanisms to control soil-borne disease. Crop Prot. 2015, 67, 269–278. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Graber, E.R.; Elad, Y.; Frenkel, O. Biochar as a management tool for soilborne diseases affecting early stage nursery seedling production. Crop Prot. 2019, 120, 34–42. [Google Scholar] [CrossRef]
- Gilardi, G.; Gullino, M.L.; Garibaldi, A. Preventative treatments in nursery with different biocontrol agents and potassium phosphite salt to control Fusarium wilt agents of lettuce and rocket. Acta Hortic. 2020, 1270, 295–300. [Google Scholar] [CrossRef]
- González, V.; Armijos, E.; Garcés-Claver, A. Fungal endophytes as biocontrol agents against the main soil-borne diseases of melon and watermelon in Spain. Agronomy 2020, 10, 820. [Google Scholar] [CrossRef]
- Tjosvold, S.A.; Chambers, D.L.; Koike, S.T.; Mori, S.R. Disease on nursery stock as affected by environmental factors and seasonal inoculum levels of Phytophthora ramorum in stream water used for irrigation. Plant Dis. 2008, 92, 1566–1573. [Google Scholar] [CrossRef]
- Parke, J.L.; Grünwald, N.J. A systems approach for management of pests and pathogens of nursery crops. Plant Dis. 2012, 96, 1236–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anith, K.N.; Radhakrishnan, N.V.; Manomohandas, T.P. Screening of antagonistic bacteria for biologicalcontrol of nursery wilt of black pepper (Piper nigrum L.). Microbiol. Res. 2003, 158, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhuo, T.; Hu, X.; Fan, X.; Zou, H. Identification of a Pseudomonas putida as biocontrol agent for tomato bacterial wilt disease. Biol. Control 2017, 114, 45–50. [Google Scholar] [CrossRef]
- O’Brien, P.A. Biological control of plant diseases. Australas. Plant Pathol. 2017, 46, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Verma, C.; Gupta, B.K.; Singh, A.; Kujur, A.; Sahu, R.; Prajapati, P. Biological control agents in the management of bell pepper nursery diseases: A review. J. Pharmacogn. Phytoch. 2020, 9, 462–468. [Google Scholar]
- Pal, K.K.; McSpadden Gardener, B. Biological control of plant pathogens. Plant Health Instr. 2006, 1117–1202. [Google Scholar] [CrossRef] [Green Version]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Lamichhane, J.R.; Dürr, C.; Schwanck, A.A.; Marie-Hélène, R.; Sarthou, J.P.; Cellier, V.; Messéan, A.; Aubertot, J.N. Integrated management of damping-off diseases. A review. Agron. Sustain. Dev. 2017, 37, 10. [Google Scholar] [CrossRef]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Tsahouridou, P.C.; Thanassoulopoulos, C.C. Proliferation of Trichoderma koningii in the tomato rhizosphere and the suppression of damping-off by Sclerotium rolfsii. Soil Biol. Biochem. 2002, 34, 767–776. [Google Scholar] [CrossRef]
- Nayak, B.K. Isolation and identification of phylloplane and endophytic fungi from one ornamental plant, Mangifera indica. Int. J. Chemtech. Res. 2015, 1, 188–192. [Google Scholar]
- Hoitink, H.A.; Musselman, C.A.; Moore, T.L.; Horst, L.; Krause, C.R.; Zondag, R.A.; Mathers, H. Biological suppression of foliar diseases of ornamental plants with composted manures, biosolids, and Trichoderma hamatum 382. Ext. Circ. 2003, 189, 50–56. [Google Scholar]
- Widmer, T.L.; Johnson-Brousseau, S.; Kosta, K.; Ghosh, S.; Schweigkofler, W.; Sharma, S.; Suslow, K. Remediation of Phytophthora ramorum-infested soil with Trichoderma asperellum isolate 04-22 under ornamental nursery conditions. Biol. Control 2018, 118, 67–73. [Google Scholar] [CrossRef]
- Lo, C.-T.; Nelson, E.B.; Hayes, C.K.; Harman, G.E. Ecological studies of transformed Trichoderma harzianum strain 1295-22 in the rhizosphere and on the phylloplane of creeping bentgrass. Phytopathology 1998, 88, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutler, H.G.; Himmelsbach, D.S.; Arrendale, R.F.; Cole, P.D.; Cox, R.H. Koninginin A: A novel plant growth regulator from Trichoderma koningii. Agr. Biol. Chem. 1989, 53, 2605–2611. [Google Scholar] [CrossRef] [Green Version]
- Benítez, T.; Rincon, A.M.; Limon, M.C.; Codon, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- Vitti, A.; Nuzzaci, M.; Scopa, A.; Sofo, A. Indirect and direct benefits of the use of Trichoderma harzianum strain T-22 in agronomic plants subjected to abiotic and biotic stresses. In Abiotic Stresses in Crop Plants; Chakraborty, U., Chakraborty, B., Eds.; CABI: Wallingford, UK, 2015; pp. 222–231. [Google Scholar]
- Vitti, A.; Pellegrini, E.; Nali, C.; Lovelli, S.; Sofo, A.; Valerio, M.; Scopa, A.; Nuzzaci, M. Trichoderma harzianum T-22 induces systemic resistance in tomato infected by cucumber mosaic virus. Front. Plant Sci. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, J.N.; Singh, A.K.; Kumar, M.; Kumai, A.; Kumar, R. Management of damping-off disease of seedling caused in solanaceous and cruciferous vegetable through integrated approach. J. Pharmacogn. Phytoch. 2018, 1, 3000–3003. [Google Scholar]
- Hewavitharana, N.; Kannangara, S.D.P. Evaluation of organic potting media enriched with Trichoderma spp. and their effect on growth performance of selected vegetables. Int. J. Sci. Basic Appl. 2019, 6, 13–25. [Google Scholar]
- Nahar, N.; Islam, R.; Uddin, M.M.; de Jong, P.; Struik, P.C.; Stomph, T.J. Reducing damping-off problems in eggplant (Solanum melongena L.): A participatory testing of nursery management in Bangladesh. Crop Prot. 2018, 112, 177–186. [Google Scholar] [CrossRef]
- Nahar, N.; Islam, R.; Uddin, M.M.; Jong, P.; Struik, P.C.; Stomph, T.J. Disease management in eggplant (Solanum melongena L.) nurseries also reduces wilt and fruit rot in subsequent plantings: A participatory testing in Bangladesh. Crop Prot. 2019, 120, 113–124. [Google Scholar] [CrossRef]
- Mastouri, F.; Bjoerkman, T.; Harman, G.E. Seed treatment with Trichoderma harzianum alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Medina, A.; Pascual, J.A.; Lloret, E.; Roldan, A. Interactions between arbuscular mycorrhizal fungi and Trichoderma harzianum and their effects on Fusarium wilt in melon plants grown in seedling nurseries. J. Sci. Food Agric. 2009, 89, 1843–1850. [Google Scholar] [CrossRef]
- de Vrije, T.; Antoine, N.; Buitelaar, R.; Bruckner, S.; Dissevelt, M.; Durand, A.; Gerlagh, M.; Jones, E.; Lüth, P.; Oostra, J.; et al. The fungal biocontrol agent Coniothyrium minitans: Production by solid-state fermentation, application and marketing. Appl. Microbiol. Biotechnol. 2001, 56, 58–68. [Google Scholar] [CrossRef]
- Patil, S.; Sriram, S. Biological control of Fusarium wilt in crop plants using non-pathogenic isolates of Fusarium species. Indian Phytopathol. 2020, 73, 11–19. [Google Scholar] [CrossRef]
- Punja, Z.K.; Utkhede, R.S. Using fungi and yeasts to manage vegetable crop diseases. Trends Biotechnol. 2003, 21, 400–407. [Google Scholar] [CrossRef]
- Gerbore, J.; Benhamou, N.; Vallance, J.; Le Floch, G.; Grizard, D.; Regnault-Roger, C.; Rey, P. Biological control of plant pathogens: Advantages and limitations seen through the case study of Pythium oligandrum. Environ. Sci. Pollut. Res. 2014, 21, 4847–4860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almoneafy, A.A.; Kakar, K.U.; Nawaz, Z.; Li, B.; Saand, M.A.; Chun-lan, Y.; Xie, G. Tomato plant growth promotion and antibacterial related-mechanisms of four Rhizobacterial bacillus strains against Ralstonia solanacearum. Symbiosis 2014, 63, 59–70. [Google Scholar] [CrossRef]
- Baysal, O.; Çalişkan, M.; Yeşilova, O. An inhibitory effect of a new Bacillus subtilis strain (EU07) against Fusarium oxysporum f. sp. radicis-lycopersici. Physiol. Mol. Plant Path 2008, 73, 25–32. [Google Scholar] [CrossRef]
- Buchenauer, H. Biological control of soil-borne diseases by rhizobacteria. J. Plant Dis. Prot. 1998, 105, 329–348. [Google Scholar]
- Heydari, A.; Pessarakli, M. A review on biological control of fungal plant pathogens using microbial antagonist. J. Biol. Sci. 2010, 10, 273–290. [Google Scholar] [CrossRef] [Green Version]
- Ouhaibi-Ben, A.N.; Vallance, J.; Gerbore, J.; Rey, P.; Daami-Remadi, M. Bio-suppression of Sclerotinia stem rot of tomato and biostimulation of plant growth using tomato-associated rhizobacteria. J. Plant Pathol. Microbiol. 2016, 7, 1000331. [Google Scholar]
- Weller, M. Biological control of soil borne plant pathogens in the rhizosphere with bacteria. Ann. Rev. Phytopathol. 1998, 26, 379–407. [Google Scholar] [CrossRef]
- Asaka, O.; Shoda, M. Biocontrol of rhizoctonia solani damping-off of tomato with Bacillus subtilis RB14. Appl. Environ. Microbiol. 1996, 62, 4081–4085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Zhang, N.; Yong, X.; Yang, X.; Shen, Q. Biocontrol of Rhizoctonia Solani damping-off disease in cucumber with Bacillus pumilus SQR-N43. Microbiol. Res. 2012, 167, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Kloepper, J.W.; Schroth, M.N.; Miller, T.D. Effects of rhizosphere colonizaton by plant growth-promoting rhizobacteria on potato plant development and yield. Phytopathology 1980, 70, 1078–1082. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Lifshitz, R.; Zablotowicz, R.M. Free-living bacterial inocula for enhancing crop productivity. Tibthech 1989, 7, 39–44. [Google Scholar] [CrossRef]
- Suslow, T.V.; Schroth, M.N. Rhizobacteria of sugar beets: Effects of seed application and root colonization on yield. Phytopathology 1982, 72, 199–206. [Google Scholar] [CrossRef]
- Tang, W.H. Yield increasing bacteria (YIB) and biocontrol of sheath blight in rice. In Improving Plant Productivity with Rhizosphere Bacteria; Ryder, M.H., Stephens, P.M., Bowen, G.D., Eds.; CSIRO Division of Soils: Adelaide, Australia, 1994; pp. 267–273. [Google Scholar]
- Agrios, G.N. Plant Pathology; Academic Press: San Diego, CA, USA, 2005. [Google Scholar]
- Bowers, J.H.; Park, J.L. Epidemiology of Pythium damping-off and Aphanomyces root-rot of peas after seed treatment with bacterial agents for biocontrol. Phytopathology 1993, 83, 1466–1473. [Google Scholar] [CrossRef]
- Dimkic, I.; Zivkovic, S.; Beric, T.; Ivanovic, Z.; Gavrilovic, V.; Stankovic, S.; Fira, D. Characterization and evaluation of two Bacillus strains, SS-12.6 and SS-13.1, as potential agents for the control of phytopathogenic bacteria and fungi. Biol. Control 2013, 65, 312–321. [Google Scholar] [CrossRef]
- Han, J.S.; Cheng, J.H.; Yoon, T.M.; Song, J.; Rajkarnikar, A.; Kim, W.G. Biological control agent of common scab disease by antagonistic strain Bacillus sp. sunhua. J. Appl. Microbiol. 2005, 99, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Khedher, S.B.; Kilani-Feki, O.; Dammak, M.; Jabnoun-Khiareddine, H.; Daami-Remadi, M.; Slim Tounsi, S. Efficacy of Bacillus subtilis V26 as a biological control agent against Rhizoctonia solani on potato. Comptes Rendus Biol. 2015, 338, 784–792. [Google Scholar] [CrossRef]
- Leben, S.D.; Wadi, J.A.; Easton, G.D. Effects of Pseudomonas fluorescens on potato plant growth and control of Verticillium dahliae. Phytopathology 1987, 77, 1592–1595. [Google Scholar] [CrossRef]
- Pane, C.; Villecco, D.; Campanile, F.; Zaccardelli, M. Novel strains of Bacillus, isolated from compost and compost-amended soils, as biological control agents against soil-borne phytopathogenic fungi. Biocontrol Sci. Technol. 2012, 22, 1373–1388. [Google Scholar] [CrossRef]
- Pane, C.; Zaccardelli, M. Evaluation of Bacillus strains isolated from Solanaceous phylloplane for biocontrol of Alternaria early blight of tomato. Biol. Control 2015, 84, 11–18. [Google Scholar] [CrossRef]
- Zaccardelli, M.; Sorrentino, R.; Caputo, M.; Scotti, R.; De Falco, E.; Pane, C. Stepwise-selected Bacillus amyloliquefaciens and B. subtilis strains from composted aromatic plant waste able to control soil-borne diseases. Agriculture 2020, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Costaccurta, A.; Vanderleyden, B. Synthesis of phythormones by plant associated bacteria. Crit. Rev. Microbiol. 1995, 21, 1–18. [Google Scholar] [CrossRef]
- D’Agostino, N.; Sorrentino, R.; Scotti, R.; Salzano, M.; Aurilia, V.; Zaccardelli, M. Draft genome sequence of the plant growth-promoting Rhizobacterium Pseudomonas fluorescens strain CREA C16 isolated from pea (Pisum sativum L.). Rhizosphere 2017, 5, e01456–e01516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Mañero, F.J.; Ramos-Solano, B.; Probanda, A.; Mehouachi, J.; Tadeo, F.R.; Talon, M. The plant growth-promoting rhizobacteria Bacillus pumilis and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiol. Plant 2001, 111, 206–211. [Google Scholar] [CrossRef]
- Kobayashi, M.; Suzuki, T.; Fruita, T.; Masuda, M.; Shinizu, S. Occurrence of enzymes ivolved in biosynthesis of indole-3-acetic acid from indole-3-acetonitrile in plant-associated bacteria, Agrobacteria and Rhizobium. Proc. Natl. Acad. Sci. USA 1995, 92, 714–718. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.E.; Schroth, M.N. Influence of bacterial sources of indole-3-acetic on root elongation of sugar beet. Phytopathology 1986, 76, 386–389. [Google Scholar]
- Cassan, F.; Diaz-Zorita, M. Azospirillum spp. in current agriculture: From the laboratory to the soil. Soil Biol. Biochem. 2016, 103, 117–130. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 65–270. [Google Scholar] [CrossRef]
- Becker, J.O.; Cook, E.J. Role of siderophores in suppression of Pythium species and production of increased growth response of wheat by fluorescent pseudomonads. Phytopathology 1988, 78, 778–782. [Google Scholar] [CrossRef]
- Darma, R.; Purnamasari, I.M.; Agustina, D.; Pramudito, T.E.; Sugiharti, M. A Strong antifungal-producing bacteria from bamboo powder for biocontrol of Sclerotium rolfsii in melon (Cucumis melo var. amanta). J. Plant Pathol. Microbiol. 2016, 7, 1000334. [Google Scholar] [CrossRef]
- Gong, M.; Wang, J.D.; Zhang, J.; Yang, H.; Lu, X.F.; Pey, Y.; EI Cheng, J.Q. Study of the antifungal ability of Bacillus subtilis strain PY-1 in vitro and identification of its antifungal substance (Iturin A). Acta Biochim. Biophys. Sin. 2006, 38, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Kim, P.I.; Ryu, J.; Kim, Y.H.; Chl, Y.T. Production of biosurfactant lipopeptides iturin A, fengycin and surfactin A from Bacillus subtilis CMB32 for control of Colletotrichum gloeosporioides. J. Microbiol. Biotechnol. 2010, 20, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, J.M.; Vlami, M.; de Souza, J.T. Antibiotic production by bacterial biocontrol agents. Antonie Leeuwenhoek 2020, 81, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Scher, F.M.; Baker, R. Effect of Pseudomonas putida and a synthetic iron chelator on induction of soil suppressiveness to Fusarium Wilt patogena. Phytopathol. 1982, 72, 1567–1573. [Google Scholar] [CrossRef]
- Tourè, Y.; Ongena, M.; Jacques, P.; Guiro, A.; Thonart, P. Role of lipopeptides produced by Bacillus subtilis GA1 in the reduction of grey mould disease caused by Botrytis cinerea on apple. J. Appl. Microbiol. 2004, 96, 1151–1160. [Google Scholar] [CrossRef]
- Zheng, M.; Shi, J.; Wang, Q.; Li, Y. Antimicrobial effects of volatiles produced by two antagonistic Bacillus strains on the anthracnose pathogen in postharvest mangos. Biol. Control 2013, 65, 200–206. [Google Scholar] [CrossRef]
- Bargabus, R.L.; Zidack, N.K.; Sherwood, J.E.; Jacobsen, B.J. Characterisation of systemic resistance in sugar beet elicited by a non-pathogenic, phyllosphere-colonizing Bacillus mycoides, biological control agent. Physiol. Mol. Plant Pathol. 2002, 61, 289–298. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. and plants with special reference to induced systemic resistance (ISR). Microbiol. Res. 2009, 164, 493–513. [Google Scholar] [CrossRef] [PubMed]
- Kloepper, J.W.; Ryu, C.M.; Zhang, S. Induce systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Wei, H.X.; Kloepper, J.W. Bacterial volatiles induce systemic resistance in Arabidopsis. Plant Physiol. 2004, 134, 1017–1026. [Google Scholar] [CrossRef] [Green Version]
- Scotti, R.; D’Agostino, N.; Zaccardelli, M. Gene expression profiling of tomato roots interacting with Pseudomonas fluorescens unravels the molecular reprogramming that occurs during the early phases of colonization. Symbiosis 2019. [Google Scholar] [CrossRef]
- Zaccardelli, M.; Carella, A.; Caiazzo, R.; Torre, R.; Tarantino, P.; Lahoz, E. Evaluation of two Bacillus pp. isolates inoculated on potato for antagonism against Rhizoctonia solani, plant growth promotion and provement of b-1,4-glycosidases, b-D-N-N’-diacetylchitobiosidase and peroxidases Activities. Plant Pathol. J. 2004, 6, 337. [Google Scholar]
- Nehl, D.B.; Allen, S.J.; Brown, J.F. Deleterious rhizobacteria: An integrating perspective. Appl. Soil Ecol. 1996, 5, 1–20. [Google Scholar] [CrossRef]
- Suslow, T.V.; Schroth, M.N. Role of deleterious rhizobacteria as minor pathogens in reducing crop growth. Phytopathology 1982, 72, 111–115. [Google Scholar] [CrossRef]
- Sahu, P.K.; Singh, S.; Gupta, A.; Singh, U.B.; Brahmaprakash, G.P.; Saxena, A.K. Antagonistic potential of bacterial endophytes and induction of systemic resistance against collar rot pathogen Sclerotium rolfsii in tomato. Biol. Control 2019, 137, 104014. [Google Scholar] [CrossRef]
- Ling, N.; Xue, C.; Huang, Q.; Yang, X.; Xu, Y.; Shen, Q. Development of a mode of application of bioorganic fertilizer for improving the biocontrol efficacy to Fusarium wilt. Biocontrol 2010, 55, 673–683. [Google Scholar] [CrossRef]
- Termorshuizen, A.J.; Jeger, M.J. Assessing inoculum of soilborne plant pathogens: Theory and practice in decision-making for soil disinfestation. Acta Hortic. 2014, 1044, 75–80. [Google Scholar] [CrossRef]
- Fravel, D.R. Commercialization and implementation of biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Roldán, A.; Pascual, J.A. Performance of a Trichoderma harzianum bentonite-vermiculite formulation against Fusarium wilt in seedling nursery melon plants. Hortic. Sci. 2009, 44, 2025–2027. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Vicente, A.; Ros, M.; Pascual, J.A. Increased effectiveness of the Trichoderma harzianum isolate T-78 against Fusarium wilt on melon plants under nursery conditions. J. Sci. Food Agric. 2009, 89, 827–833. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronga, D.; Vitti, A.; Zaccardelli, M.; Pane, C.; Caradonia, F.; Cardarelli, M.; Colla, G.; Rouphael, Y. Root Zone Management for Improving Seedling Quality of Organically Produced Horticultural Crops. Agronomy 2021, 11, 630. https://doi.org/10.3390/agronomy11040630
Ronga D, Vitti A, Zaccardelli M, Pane C, Caradonia F, Cardarelli M, Colla G, Rouphael Y. Root Zone Management for Improving Seedling Quality of Organically Produced Horticultural Crops. Agronomy. 2021; 11(4):630. https://doi.org/10.3390/agronomy11040630
Chicago/Turabian StyleRonga, Domenico, Antonella Vitti, Massimo Zaccardelli, Catello Pane, Federica Caradonia, Mariateresa Cardarelli, Giuseppe Colla, and Youssef Rouphael. 2021. "Root Zone Management for Improving Seedling Quality of Organically Produced Horticultural Crops" Agronomy 11, no. 4: 630. https://doi.org/10.3390/agronomy11040630