
Divergent Serpentoviruses in Free-Ranging Invasive Pythons and Native Colubrids in Southern Florida, United States
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
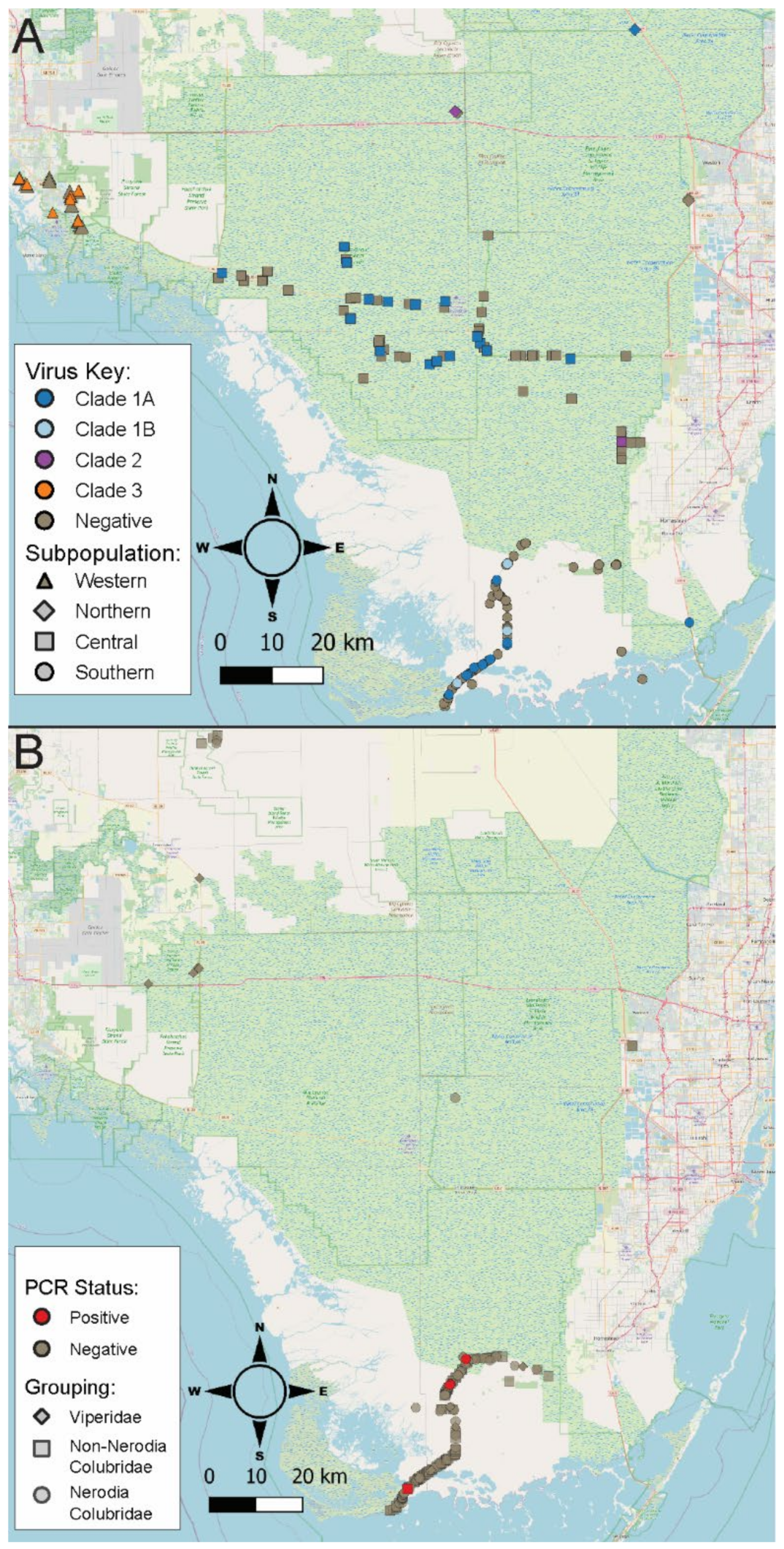
2.1. Burmese Python Population and Sampling Investigation
2.2. Longitudinal Sampling
2.3. Native Snake Population and Sampling Investigation
2.4. Postmortem Examinations
2.5. Serpentovirus rtPCR Screening
2.6. Illumina MiSeq Nextgen Sequencing
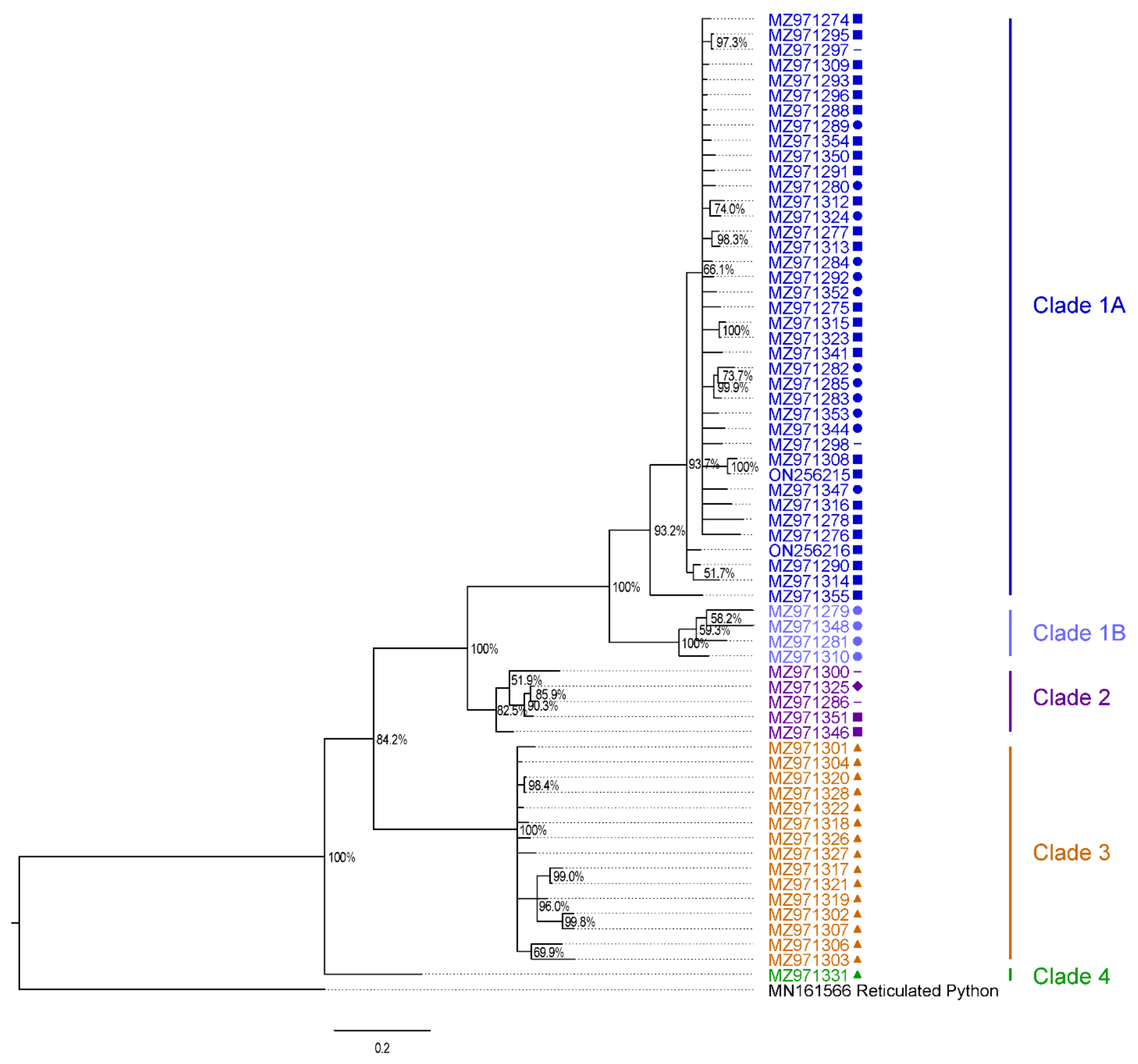
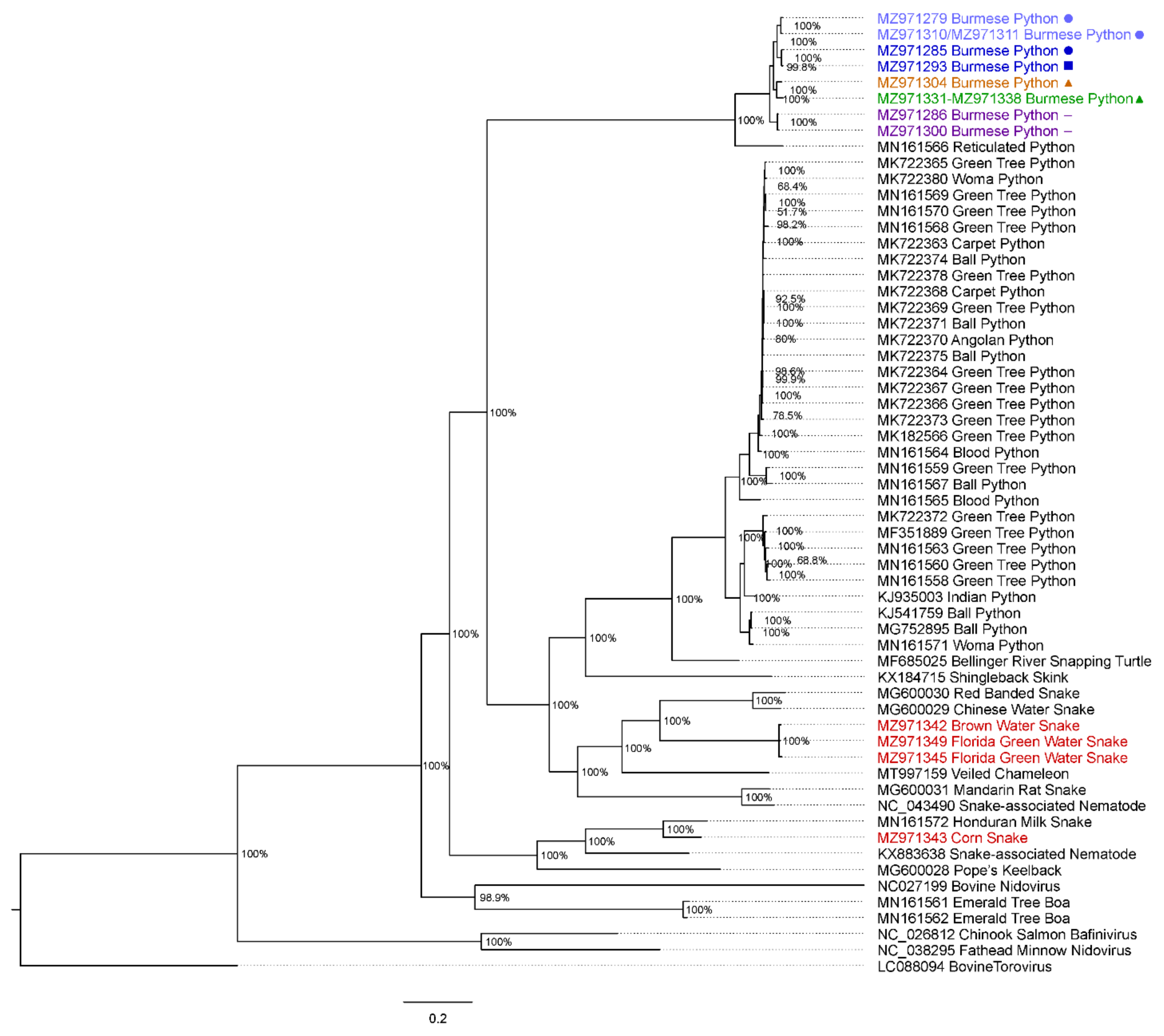
2.7. Phylogenetic Analysis and Preliminary Taxonomic Classification
2.8. Virus Isolation Attempts
3. Results
3.1. Burmese Python Population and Sampling Investigation
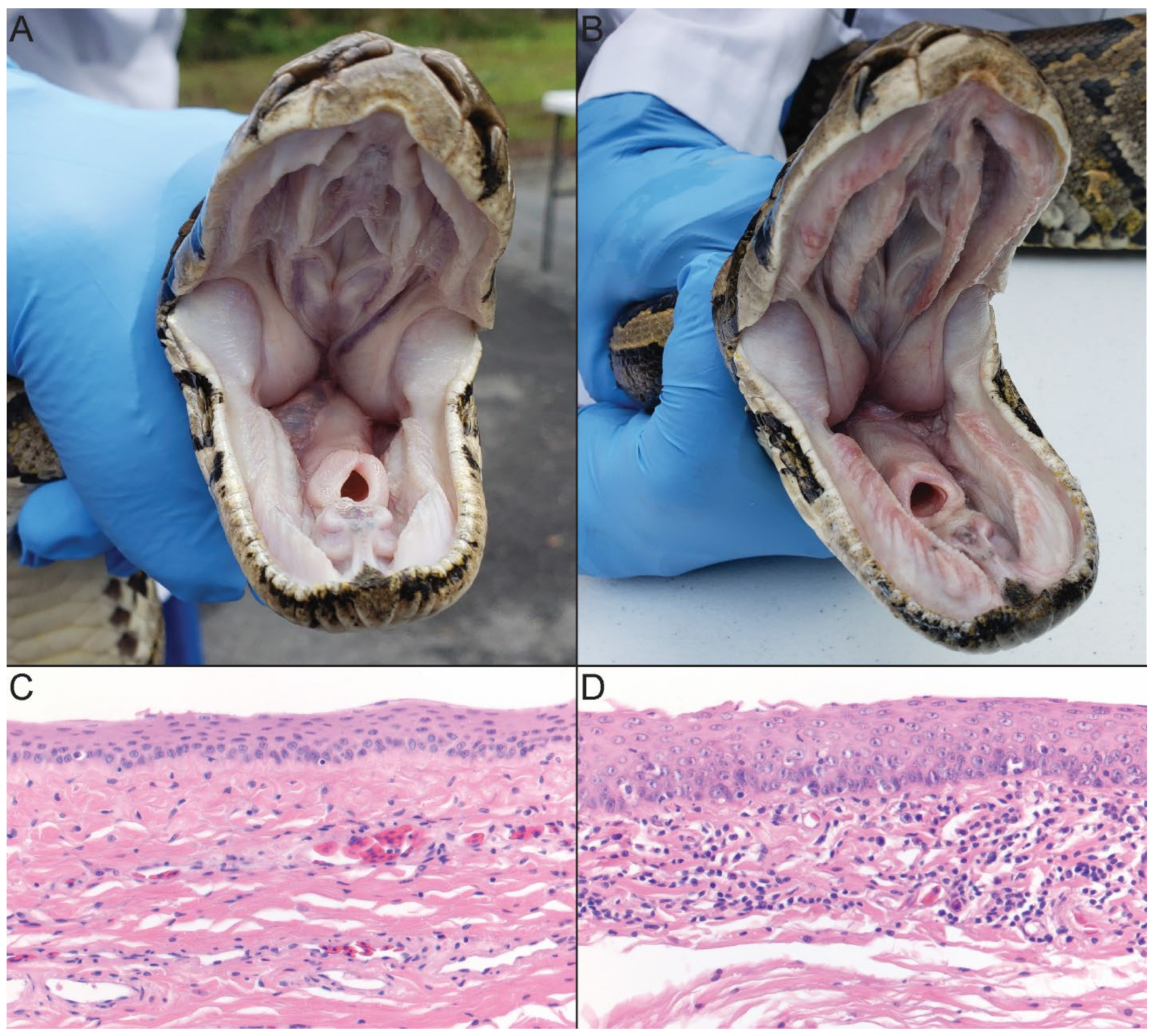
3.2. Clinical Signs of Infection
3.3. Postmortem Findings
3.4. Longitudinal Sampling
3.5. Native Snake Sampling
3.6. Virus Isolation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Willson, J.D.; Dorcas, M.E.; Snow, R.W. Identifying Plausible Scenarios for the Establishment of Invasive Burmese Pythons (Python Molurus) in Southern Florida. Biol. Invasions 2011, 13, 1493–1504. [Google Scholar] [CrossRef]
- Snow, R.; Krysko, K.; Enge, K.; Oberhofer, L.; Warren-Bradley, A.; Wilkins, L. Introduced Populations of Boa constrictor (Boidae) and Python molurus bivittatus (Pythonidae) in Southern Florida. In Biology of the Boas and Pythons; Henderson, R.W., Powell, R., Eds.; Eagle Mountain Publishing: Eagle Mountain, UT, USA, 2007; pp. 416–438. [Google Scholar]
- Dove, C.J.; Snow, R.W.; Rochford, M.R.; Mazzotti, F.J. Birds Consumed by the Invasive Burmese Python (Python molurus bivittatus) in Everglades National Park, Florida, USA. Wilson J. Ornithol. 2011, 123, 126–131. [Google Scholar] [CrossRef]
- Dorcas, M.E.; Willson, J.D.; Reed, R.N.; Snow, R.W.; Rochford, M.R.; Miller, M.A.; Meshaka, W.E.; Andreadis, P.T.; Mazzotti, F.J.; Romagosa, C.M.; et al. Severe Mammal Declines Coincide with Proliferation of Invasive Burmese Pythons in Everglades National Park. Proc. Natl. Acad. Sci. USA 2012, 109, 2418–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCleery, R.A.; Sovie, A.; Reed, R.N.; Cunningham, M.W.; Hunter, M.E.; Hart, K.M. Marsh Rabbit Mortalities Tie Pythons to the Precipitous Decline of Mammals in the Everglades. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150120. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Kinsella, J.M.; Snow, R.W.; Hayes, M.M.; Falk, B.G.; Reed, R.N.; Mazzotti, F.J.; Guyer, C.; Romagosa, C.M. Parasite Spillover: Indirect Effects of Invasive Burmese Pythons. Ecol. Evol. 2018, 8, 830–840. [Google Scholar] [CrossRef]
- Miller, M.A.; Kinsella, J.M.; Snow, R.W.; Falk, B.C.; Reed, R.N.; Goetz, S.M.; Mazzotti, F.J.; Guyer, C.; Romagosa, C.M. Highly Competent Native Snake Hosts Extend the Range of an Introduced Parasite beyond Its Invasive Burmese Python Host. Ecosphere 2020, 11, e03153. [Google Scholar] [CrossRef]
- Huegel, C.; Cook, D. Florida’s Nonvenomous Snakes. Florida Wildl. 2004, 45, 9–14. [Google Scholar]
- Krysko, K.L.; Enge, K.M.; Moler, P.E. Atlas of Amphibians and Reptiles in Florida; Florida Fish and Wildlife Conservation Commission: Tallahassee, FL, USA, 2011. [Google Scholar]
- Tuberville, T.D.; Bodie, J.R.; Jensen, J.B.; LaClaire, L.; Gibbons, J.W. Apparent decline of the southern hog-nosed snake, Heterodon simus. J. Elisha Mitchell Sci. Soc. 2000, 116, 19–40. [Google Scholar]
- Jansen, K.P.; Mushinsky, H.R.; Karl, S.A. Population Genetics of the Mangrove Salt Marsh Snake, Nerodia clarkii Compressicauda, in a Linear, Fragmented Habitat. Conserv. Genet. 2008, 9, 401–410. [Google Scholar] [CrossRef]
- Cheatwood, J.L.; Jacobson, E.R.; May, P.G.; Farrell, T.M.; Homer, B.L.; Samuelson, D.A.; Kimbrough, J.W. An Outbreak of Fungal Dermatitis and Stomatitis in a Free-Ranging Population of Pigmy Rattlesnakes (Sistrurus miliarius barbouri) in Florida. J. Wildl. Dis. 2003, 39, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Lind, C.M.; McCoy, C.M.; Farrell, T.M. Tracking Outcomes of Snake Fungal Disease in Free-Ranging Pygmy Rattlesnakes (Sistrurus miliarius). J. Wildl. Dis. 2018, 54, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Enjuanes, L.; Gorbalenya, A.E.; de Groot, R.J.; Cowley, J.A.; Ziebuhr, J.; Snijder, E.J. Nidovirales. In Encyclopedia of Virology; Academic Press: Cambridge, MA, USA, 2008; pp. 419–430. ISBN 9780123744104. [Google Scholar]
- Bartas, M.; Brázda, V.; Bohálová, N.; Cantara, A.; Volná, A.; Stachurová, T.; Malachová, K.; Jagelská, E.B.; Porubiaková, O.; Červeň, J.; et al. In-Depth Bioinformatic Analyses of Nidovirales Including Human SARS-CoV-2, SARS-CoV, MERS-CoV Viruses Suggest Important Roles of Non-Canonical Nucleic Acid Structures in Their Lifecycles. Front. Microbiol. 2020, 11, 1583. [Google Scholar] [CrossRef] [PubMed]
- Hoon-Hanks, L.L.; Layton, M.L.; Ossiboff, R.J.; Parker, J.S.L.; Dubovi, E.J.; Stenglein, M.D. Respiratory Disease in Ball Pythons (Python Regius) Experimentally Infected with Ball Python Nidovirus. Virology 2018, 517, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Lempp, C.; Schürch, A.C.; Habierski, A.; Hahn, K.; Lamers, M.; Von Dörnberg, K.; Wohlsein, P.; Drexler, J.F.; Haagmans, B.L.; et al. Novel Divergent Nidovirus in a Python with Pneumonia. J. Gen. Virol. 2014, 95, 2480–2485. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Finlaison, D.S.; Frost, M.J.; Gestier, S.; Gu, X.; Hall, J.; Jenkins, C.; Parrish, K.; Read, A.J.; Srivastava, M.; et al. Identification of a Novel Nidovirus as a Potential Cause of Large Scale Mortalities in the Endangered Bellinger River Snapping Turtle (Myuchelys Georgesi). PLoS ONE 2018, 13, e0205209. [Google Scholar] [CrossRef] [Green Version]
- Dervas, E.; Hepojoki, J.; Smura, T.; Prähauser, B.; Windbichler, K.; Blümich, S.; Ramis, A.; Hetzel, U.; Kipar, A. Python Nidoviruses, More than Respiratory Pathogens. J. Virol. 2020, 94, e00649-20. [Google Scholar] [CrossRef] [PubMed]
- Hoon-Hanks, L.L.; Ossiboff, R.J.; Bartolini, P.; Fogelson, S.B.; Perry, S.M.; Stöhr, A.C.; Cross, S.T.; Wellehan, J.F.X.; Jacobson, E.R.; Dubovi, E.J.; et al. Longitudinal and Cross-Sectional Sampling of Serpentovirus (Nidovirus) Infection in Captive Snakes Reveals High Prevalence, Persistent Infection, and Increased Mortality in Pythons and Divergent Serpentovirus Infection in Boas and Colubrids. Front. Vet. Sci. 2019, 6, 338. [Google Scholar] [CrossRef] [Green Version]
- Hoon-Hanks, L.L.; Stöhr, A.C.; Anderson, A.J.; Evans, D.E.; Nevarez, J.G.; Díaz, R.E.; Rodgers, C.P.; Cross, S.T.; Steiner, H.R.; Parker, R.R.; et al. Serpentovirus (Nidovirus) and Orthoreovirus Coinfection in Captive Veiled Chameleons (Chamaeleo calyptratus) with Respiratory Disease. Viruses 2020, 12, 1329. [Google Scholar] [CrossRef]
- O’Dea, M.A.; Jackson, B.; Jackson, C.; Xavier, P.; Warren, K. Discovery and Partial Genomic Characterisation of a Novel Nidovirus Associated with Respiratory Disease in Wild Shingleback Lizards (Tiliqua rugosa). PLoS ONE 2016, 11, e0165209. [Google Scholar] [CrossRef] [Green Version]
- Blahak, S.; Jenckel, M.; Höper, D.; Beer, M.; Hoffmann, B.; Schlottau, K. Investigations into the Presence of Nidoviruses in Pythons. Virol. J. 2020, 17, 6. [Google Scholar] [CrossRef] [Green Version]
- Leary, S.; Underwood, W.; Anthony, R.; Cartner, S.; Grandin, T.; Greenacre, C.; Gwaltney-brant, S.; Mccrackin, M.A.; Meyer, R.; Miller, D.; et al. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition; American Veterinary Medical Association: Schaumburg, IL, USA, 2020; ISBN 9781882691548. [Google Scholar]
- Katoh, K.; Toh, H. Improved Accuracy of Multiple NcRNA Alignment by Incorporating Structural Information into a MAFFT-Based Framework. BMC Bioinform. 2008, 9, 212. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Schwartz, T.; Pickett, B.E.; He, S.; Klem, E.B.; Scheuermann, R.H.; Passarotti, M.; Kaufman, S.; Oleary, M.A. A RESTful API for Access to Phylogenetic Tools via the CIPRES Science Gateway. Evol. Bioinform. 2015, 11, 43–48. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A Rapid Bootstrap Algorithm for the RAxML Web Servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Falk, B.G.; Snow, R.W.; Reed, R.N. Prospects and Limitations of Citizen Science in Invasive Species Management: A Case Study with Burmese Pythons in Everglades National Park. Southeast. Nat. 2016, 15, 89–102. [Google Scholar] [CrossRef]
- Gorbalenya, A.; Samborskiy, D.; Junglen, S.; Lauber, C.; Neuman, B.; Ziebuhr, J. Create 47 New Taxa in the Order, Ranging from Subfamilies to Species (Nidovirales). 2021. Available online: https://ictv.global/ICTV/proposals/2021.005S.R.Nidovirales.zip (accessed on 14 November 2022).
- Hunter, M.E.; Johnson, N.A.; Smith, B.J.; Davis, M.C.; Butterfield, J.S.S.; Snow, R.W.; Hart, K.M. Cytonuclear Discordance in the Florida Everglades Invasive Burmese Python (Python bivittatus) Population Reveals Possible Hybridization with the Indian Python (P. molurus). Ecol. Evol. 2018, 8, 9034–9047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartoszek, I. Natural History Note: Python bivittatus (Burmese Python). Aberrant Pattern. Herpetol. Rev. 2020, 51, 155–156. [Google Scholar]
- Smith, B.J.; Rochford, M.R.; Brien, M.; Cherkiss, M.S.; Mazzotti, F.J.; Snow, S.; Hart, K.M. Largest Breeding Aggregation of Burmese Pythons (Python bivittatus) Kuhl 1820 (Squamata: Pythonidae) and Implications for Potential Development of a Control Tool. IRCF Reptil. Amphib. 2015, 22, 16–19. [Google Scholar] [CrossRef]
- Bartoszek, I.A.; Smith, B.J.; Reed, R.N.; Hart, K.M. Spatial Ecology of Invasive Burmese Pythons in Southwestern Florida. Ecosphere 2021, 12, e03564. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Bushmaker, T.; Munster, V.J. Stability of Middle East Respiratory Syndrome Coronavirus (MERS-CoV) under Different Environmental Conditions. Eurosurveillance 2013, 18, 20590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uccellini, L.; Ossiboff, R.J.; De Matos, R.E.; Morrisey, J.K.; Petrosov, A.; Navarrete-Macias, I.; Jain, K.; Hicks, A.L.; Buckles, E.L.; Tokarz, R.; et al. Identification of a Novel Nidovirus in an Outbreak of Fatal Respiratory Disease in Ball Pythons (Python regius). Virol. J. 2014, 11, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenglein, M.D.; Jacobson, E.R.; Wozniak, E.J.; Wellehan, J.F.X.; Kincaid, A.; Gordon, M.; Porter, B.F.; Baumgartner, W.; Stahl, S.; Kelley, K.; et al. Ball Python Nidovirus: A Candidate Etiologic Agent for Severe Respiratory Disease in Python regius. MBio 2014, 5, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Dervas, E.; Hepojoki, J.; Laimbacher, A.; Romero-Palomo, F.; Jelinek, C.; Keller, S.; Smura, T.; Hepojoki, S.; Kipar, A.; Hetzel, U. Nidovirus-Associated Proliferative Pneumonia in the Green Tree Python (Morelia viridis). J. Virol. 2017, 91, e00718-17. [Google Scholar] [CrossRef] [PubMed]
- Claunch, N.M.; Bartoszek, I.A.; Tillis, S.; Stacy, N.I.; Ossiboff, R.J.; Oakey, S.; Schoenle, L.A.; Wellehan, J.F.X.; Romagosa, C.M. Physiological Effects of Capture and Short-Term Captivity in an Invasive Snake Species, the Burmese Python (Python bivittatus) in Florida. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2022, 267, 111162. [Google Scholar] [CrossRef] [PubMed]
- Krysko, K.L.; Enge, K.M.; King, F.W. The Veiled Chameleon, Chamaeleo calyptratus: A New Exotic Lizard Species. Fla. Sci. 2004, 67, 249–253. [Google Scholar]
- Tillis, S.B.; Josimovich, J.M.; Miller, M.A.; Hoon-Hanks, L.L.; Hartmann, A.M.; Claunch, N.M.; Iredale, M.E.; Logan, T.D.; Yackel Adams, A.A.; Bartoszek, I.A.; et al. Serpentoviruses in Free-Ranging Invasive Pythons and Native Colubrids in Southern Florida, United States, 2018–2020: US Geological Survey Data Release, 2022. Available online: https://doi.org/10.5066/P98AZ5AU (accessed on 14 November 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Common Name | Sampled | rtPCR Positive |
|---|---|---|---|---|
| Agkistrodon | A. conanti | Florida Cottonmouth | 5 (2.3%) | - |
| Cemophora | C. coccinea | Scarlet Snake | 20 (9.1%) | - |
| Coluber | C. constrictor | Southern Black Racer | 4 (1.8%) | - |
| Crotalus | C. adamanteus | Eastern Diamondback Rattlesnake | 6 (2.7%) | - |
| Diadophis | D. punctatus | Ringneck Snake | 8 (3.6%) | - |
| Drymarchon | D. couperi | Eastern Indigo | 1 (0.5%) | - |
| Lampropeltis | 6 (2.7%) | - | ||
| L. getula | Eastern Kingsnake | 4 (1.8%) | - | |
| L. elapsoides | Scarlet Kingsnake | 2 (0.9%) | - | |
| Nerodia | 102 (46.6%) | 4 (3.9%) | ||
| N. fasciata | Banded Watersnake | 31 (14.2%) | - | |
| N. taxispilota | Brown Watersnake | 27 (12.3%) | 2 (7.4%) | |
| N. floridana | Florida Green Watersnake | 18 (8.2%) | 2 (11.1%) | |
| N. clarkii | Salt Marsh Snake | 26 (11.9%) | - | |
| Pantherophis | 29 (13.2%) | 1 (3.4%) | ||
| P. guttatus | Corn Snake | 21 (9.6%) | 1 (4.8%) | |
| P. alleghaniensis | Eastern Ratsnake | 8 (3.7%) | - | |
| Liodytes | L. alleni | Striped Crayfish Snake | 9 (4.1%) | - |
| Storeria | S. dekayi | DeKay’s Brown Snake | 3 (1.4%) | - |
| Thamnophis | 26 (11.9%) | - | ||
| T. sirtalis | Garter Snake | 11 (5.0%) | - | |
| T. saurita | Ribbon Snake | 15 (6.8%) | - | |
| Total | 18 | 219 | 5 (2.3%) |
| Sense Primer | Antisense Primer | Annealing Temp (°C) | Target Length (Approx. nt) |
|---|---|---|---|
| BarniPVTF (5′-GAG GAC TCC ACA ARC CAG TCA C-3′) | BarniDYTR (5′-RCT RCG GTC GCA TTT CGT RTA RTC-3′) | 46 | 150 |
| BurmLHHF1 (5′-TCG AGG ACT TCA AAG CCG TC-3′) | BurmLHHR1 (5′-TGT TCG TCG TTG GGT GTT GA-3′) | 42 | 650 |
| BurmROF1 (5′-CTC ATG TCM GTC AAR CAA GAC GAC AT-3′) | BurmROR2 (5′-AAR CAA AAD GCW GCC ATC TC-3′) | 42 | 450 |
| BurmSTF1 (5′-CAA GGY CTC ATG TCA GTC AA-3′) | BurmSTR3 (5′-AAR GCW GTN GTY GCR TCY CCT GA-3′) | 46 | 550 |
| Variable | Category | Positive | Negative | p Value | df | Cramér’s V |
|---|---|---|---|---|---|---|
| Wild Prevalence | (95% CI) | 42 (24.4%) ± 6.5% | 130 (75.6%) ± 6.5% | - | - | - |
| Subpopulation 1 | Western | 8 (34.8%) | 15 (65.2%) | 4.3 × 10−1 | 3 | 0.117 |
| Northern | 2 (40.0%) | 3 (60.0%) | ||||
| Central | 17 (22.1%) | 60 (77.9%) | ||||
| Southern | 15 (22.4%) | 52 (77.6%) | ||||
| Sex 2 | Male | 35 (34.3%) | 67 (65.7%) | 3.9 × 10−3 * | 1 | 0.243 |
| Female | 7 (12.1%) | 51 (87.9%) | ||||
| Season 2 | Winter | 13 (38.2%) | 21 (61.8%) | 7.3 × 10−2 | 3 | 0.196 |
| Spring | 10 (24.4%) | 31 (75.6%) | ||||
| Summer | 15 (17.4%) | 71 (82.6%) | ||||
| Fall | 4 (36.4%) | 7 (63.6%) | ||||
| Morphometrics 3 | Mass (g) (95% CI) | 9955.8 ± 3324.3 | 7091.6 ± 1952.7 | 1.6 × 10−1 | 171 | 0.968 |
| SVL (cm) (95% CI) | 234.0 ± 17.5 | 184.7 ± 14.7 | 1.2 × 10−3 * | 171 | 0.943 | |
| Latitude 3 | Northing (95% CI) | 2,839,487 ± 12133 | 2,832,197 ± 5758 | 2.4 × 10−1 | 171 | 0.981 |
| SubPop | Virus | Snake | Oct | Nov | Dec | Jan | Feb | Mar | Apr | Jul | Aug | Sep | nt Identity between + Tests | Fragment Length (nt) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Central | 1A | 1 | − | + | |||||||||||||
| 2 | + | − | + | 100% | 133 | ||||||||||||
| 3 | + | − | |||||||||||||||
| 4 | − | + | − | ||||||||||||||
| 5 | + | − | |||||||||||||||
| 6 | + | + | − | − | 100% | 133 | |||||||||||
| 7 | + | − | |||||||||||||||
| 8 | + | + | 100% | 133 | |||||||||||||
| 9 | + | + | 100% | 133 | |||||||||||||
| 10 | + | + | 100% | 133 | |||||||||||||
| 11 | + | − | |||||||||||||||
| 12 | + | + | − | 99.5% | 411 | ||||||||||||
| 2 | 13 | + | − | ||||||||||||||
| NA | 14 | − | − | − | − | ||||||||||||
| 15 | − | − | − | ||||||||||||||
| 16 | − | − | − | − | |||||||||||||
| 17 | − | − | |||||||||||||||
| 18 | − | − | |||||||||||||||
| 19 | − | − | |||||||||||||||
| 20 | − | − | − | − | |||||||||||||
| 21 | − | − | |||||||||||||||
| 22 | − | − | |||||||||||||||
| 23 | − | − | − | − | − | ||||||||||||
| 24 | − | − | − | ||||||||||||||
| 25 | − | − | − | ||||||||||||||
| 26 | − | − | − | − | |||||||||||||
| 27 | − | − | |||||||||||||||
| Southern | 1A | 28 | + | − | − | − | + | − | 100% | 133 | |||||||
| 29 | + | + | + | 100% | 133 | ||||||||||||
| 30 | − | + | |||||||||||||||
| 31 | + | + | 100% | 133 | |||||||||||||
| 32 | + | + | 96.7% | 196 | |||||||||||||
| 1B | 33 | − | + | ||||||||||||||
| NA | 34 | − | − | − | − | − | |||||||||||
| 35 | − | − | |||||||||||||||
| 36 | − | − | |||||||||||||||
| 37 | − | − | |||||||||||||||
| Western | 3 | 38 | − | + | |||||||||||||
| 39 | − | + | |||||||||||||||
| 40 | − | + | |||||||||||||||
| 4 | 41 | − | + | ||||||||||||||
| NA | 42 | − | − | ||||||||||||||
| 43 | − | − | |||||||||||||||
| 44 | − | − | |||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tillis, S.B.; Josimovich, J.M.; Miller, M.A.; Hoon-Hanks, L.L.; Hartmann, A.M.; Claunch, N.M.; Iredale, M.E.; Logan, T.D.; Yackel Adams, A.A.; Bartoszek, I.A.; et al. Divergent Serpentoviruses in Free-Ranging Invasive Pythons and Native Colubrids in Southern Florida, United States. Viruses 2022, 14, 2726. https://doi.org/10.3390/v14122726
Tillis SB, Josimovich JM, Miller MA, Hoon-Hanks LL, Hartmann AM, Claunch NM, Iredale ME, Logan TD, Yackel Adams AA, Bartoszek IA, et al. Divergent Serpentoviruses in Free-Ranging Invasive Pythons and Native Colubrids in Southern Florida, United States. Viruses. 2022; 14(12):2726. https://doi.org/10.3390/v14122726
Chicago/Turabian StyleTillis, Steven B., Jillian M. Josimovich, Melissa A. Miller, Laura L. Hoon-Hanks, Arik M. Hartmann, Natalie M. Claunch, Marley E. Iredale, Tracey D. Logan, Amy A. Yackel Adams, Ian A. Bartoszek, and et al. 2022. "Divergent Serpentoviruses in Free-Ranging Invasive Pythons and Native Colubrids in Southern Florida, United States" Viruses 14, no. 12: 2726. https://doi.org/10.3390/v14122726