
New Brittle Stars (Echinodermata, Ophiuroidea) from the Oligocene of the Mainz Basin, Germany

1
Natural History Museum Luxembourg, Department of Palaeontology, 25 Rue Münster, L-2160 Luxembourg, Luxembourg
2
Independent Researcher, Kettenheimer Strasse 16, 55232 Alzey, Germany
*
Author to whom correspondence should be addressed.
Taxonomy 2022, 2(2), 196-207; https://doi.org/10.3390/taxonomy2020015
Submission received: 4 April 2022
/
Revised: 15 April 2022
/
Accepted: 20 April 2022
/
Published: 27 April 2022
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The fossil record of the Ophiuroidea is still patchy, especially in the Cenozoic. Only four species have been described from the entire Oligocene, which is in stark contrast to the present-day diversity counting more than 2000 species. Here, we describe two new species of ophiuroid, Ophiura tankardi sp. nov. and Ophiodoris niersteinensis sp. nov., from the Lower Oligocene of the Mainz Basin. The species are based on microfossils extracted from the sieving residues of bulk sediment samples from a flush drill in Nierstein, Rhineland-Palatinate. The new species belong to extant genera and add to the poor Oligocene fossil record of the class. Based on present-day distributions, the occurrence of Ophiodoris suggests deep sublittoral to shallow bathyal palaeodepths for the Nierstein area of the Mainz Basin.
1. Introduction
Ophiuroids, or brittle stars, are the slender-armed cousins of the starfish and live in every corner of the world seas, from the poles to the tropics and from the intertidal to the deep sea [1]. Their skeleton is composed of a multitude of high-magnesium calcite ossicles connected with soft tissue. Due to the rapid post-mortem decay of the soft tissue, ophiuroid skeletons rarely fossilize intact. Disarticulated ossicles, in contrast, have a very high fossilization potential and can be found in great numbers in marine rocks [2,3,4]. They can be extracted from bulk sediment samples using standard microfossil processing techniques, in particular screen-washing and picking under dissecting microscopes [5].
Although the systematic study of the ophiuroid microfossil record started several decades ago [2,3], the number of new species descriptions based on fossil dissociated lateral arm plates has skyrocketed thanks to the recent progress in understanding the complex micromorphology of the ophiuroid skeleton [5,6]. It is now well established that specific types of ossicles, in particular the spine-bearing lateral arm plates, yield a sufficient amount of morphological information for identifications to species level [5] and for inclusion in phylogenetic estimates [7,8]. During the last few years, the ophiuroid fossil record has been explored systematically [4]. Nevertheless, some stratigraphic intervals are still poorly known. The Oligocene fossil record is particularly patchy, with only four species from that epoch [4,9,10,11].
Here, we describe two new species of ophiuroids from the lower Oligocene of the Mainz Basin, based on dissociated lateral arm plates and other skeletal components. The study was incited by preliminary reports on ophiuroid microfossils from sieving residues of Oligocene sediments from the Mainz Basin [12]. The species described in the present paper significantly add to the generally poor Cenozoic fossil record of the Ophiuroidea.
2. Materials and Methods
The fossils described in the present paper consist in microfossils retrieved from the sieving residues of bulk sediment samples. The material originates from samples taken in 2005 during a geothermal drilling at the marketplace in Nierstein (WGS84 49°52′29.5″ N/8°20′11″ E), in the Mainz Basin (Figure 1), by the employees of the drilling company. After drying, the samples were processed with water and washed over a 63 µm sieve and the residues were picked under the microscope.
The stratigraphic interpretation of the samples was based on sedimentology and microfauna. Due to the sample collecting method (flush drilling), contamination of the samples by other material cannot always be excluded with certainty, which explains, for example, the apparently high thickness of the Rosenberg Subformation (see below). At a depth of 94 meters, the drilling hit Rotliegend rocks from the Permian. The sedimentary- rocks between 92 and 26 meters belong to the Bodenheim Formation, with the Wallau Subformation at 92 meters depth, the Hochberg Subformation from 90 to 54 meters, and the Rosenberg Subformation from 52 to 26 meters. The top of the drilled section belongs to the Pfadberg Subformation of the Stadecken Formation. Ophiuroid remains were found in several samples of the Rosenberg Subformation.
During the Oligocene, the Mainz Basin was part of an arm of the sea extending southwards from the then North Sea, from the Kassel Bay to the Upper Rhine Graben. Whether there was a connection to the Paratethys in the south, at least temporarily, is still debated [13]. Nierstein is located at the eastern edge of the Mainz Basin at the border to the Upper Rhine Graben, in the area of the Alzey-Nierstein horst consisting of Upper Carboniferous to Lower Permian rocks (Figure 2). At the time of deposition of the Rosenberg Subformation, Nierstein was located relatively far from the coast at a greater depth, but not at maximum water depth due to its location in the area of the Alzey-Niersteiner Horst (Figure 2).
The Bodenheim Formation was deposited in the Oligocene during the second Rupelian transgression in a fully marine setting. It is subdivided into the Wallau, Hochberg and Rosenberg Subformations. The Rosenberg Subformation overlies not only the Hochberg Subformation, but also the older coastal sediments and the pre-Cenozoic basement, which documents an increase in water depth and consequently an extension of the depositional space [13]. Based on sedimentological evidence and the composition of benthic communities, maximum water depth probably reached 150 meters, although some authors argue for a shallower water depth [14,15]. For the Hochberg Subformation, Grimm [16] used stable isotope analyses to reconstruct a surface water temperature of 11–22 °C and bottom water temperature of 8–12 °C, corresponding to a subtropical-Mediterranean climate. Based on ostracod evidence, the depositional setting of the Rosenberg Subformation was a euryhaline, quiet area of the middle to deep sublittoral [17].
The Rosenberg Subformation consists of grey to grey-green clayey marls and reaches a thickness of up to 20 meters [14]. It belongs to nannoplankton zone NP 24 and to dinoflagellate zones D 14/D 15 [13] and, thus, to the Rupelian. The subformation yields a rich microfauna dominated by foraminifera. Ostracods, sea urchin remains, and fish bones and teeth occur more rarely, but regularly, while brittle star microfossils are only found at a few sites [18].
The brittle-star microfossils described in the present paper were retrieved from 4 samples, taken at depths of 34, 42, 46, and 50 meters, respectively. They consist of fully dissociated skeletal plates, including lateral, ventral, and dorsal arm plates; vertebrae; arm spines; oral plates; and genital plates, as well as a few articulated, distal arm segments. Selected plates and arm segments were mounted on aluminum stubs and silver-coated for scanning electron microscopy using a JEOL Neoscope JMC-5000. All figured specimens are housed at the Natural History Museum Mainz/State Collection of Natural History of Rhineland-Palatinate (acronym NHMMZ).
Terminology follows Stöhr et al. [1] and Thuy and Stöhr [5]. We use the classification by O’Hara et al., (2017, 2018).
The electronic version of this article in Portable Document Format (PDF) represents a published work according to the International Commission on Zoological Nomenclature (ICZ), and, hence, the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org:pub:3D4DDF80-AD07-43F4-91A9-D934CF7647D5.
3. Systematic Palaeontology
Class Ophiuroidea Gray, 1840
Subclass Myophiuroida Matsumoto, 1915
Infraclass Metophiurida Matsumoto, 1913
Superorder Euryophiurida O’Hara, Hugall, Thuy, Stöhr & Martynov, 2017
Order Ophiurida Müller & Troschel, 1840 sensu O’Hara et al., 2017
Suborder Ophiurina Müller & Troschel, 1840 sensu O’Hara et al., 2017
Family Ophiuridae Müller & Troschel, 1840
Genus Ophiura Lamarck, 1801
Diagnosis (emended and specifically focusing on lateral arm plate morphology): lateral arm plates of rectangular to rounded rectangular outline, with a deep, well-defined, distalwards-pointing tentacle notch in the middle of the distal edge or slightly more ventrally, resulting in nearly equal-sized dorsal and ventral portions of the lateral arm plate; outer surface with a fine tuberculation sometimes merged into a fine vertical striation; outer distal edge often with one or two small, pointed, poorly defined spurs; generally three arm spine articulations on the same level as the outer surface stereom, with the muscle opening proximally bordered by a thin, rugose, vertical ridge; and inner side of lateral arm plate with a thin C-shaped vertebral articular ridge, often with the ventral and dorsal tips enlarged.
Ophiura tankardi sp. nov. (Figure 3)
urn:lsid:zoobank.org:act:5E4A429C-160F-489D-BCEB-70D7E3BF5BDE
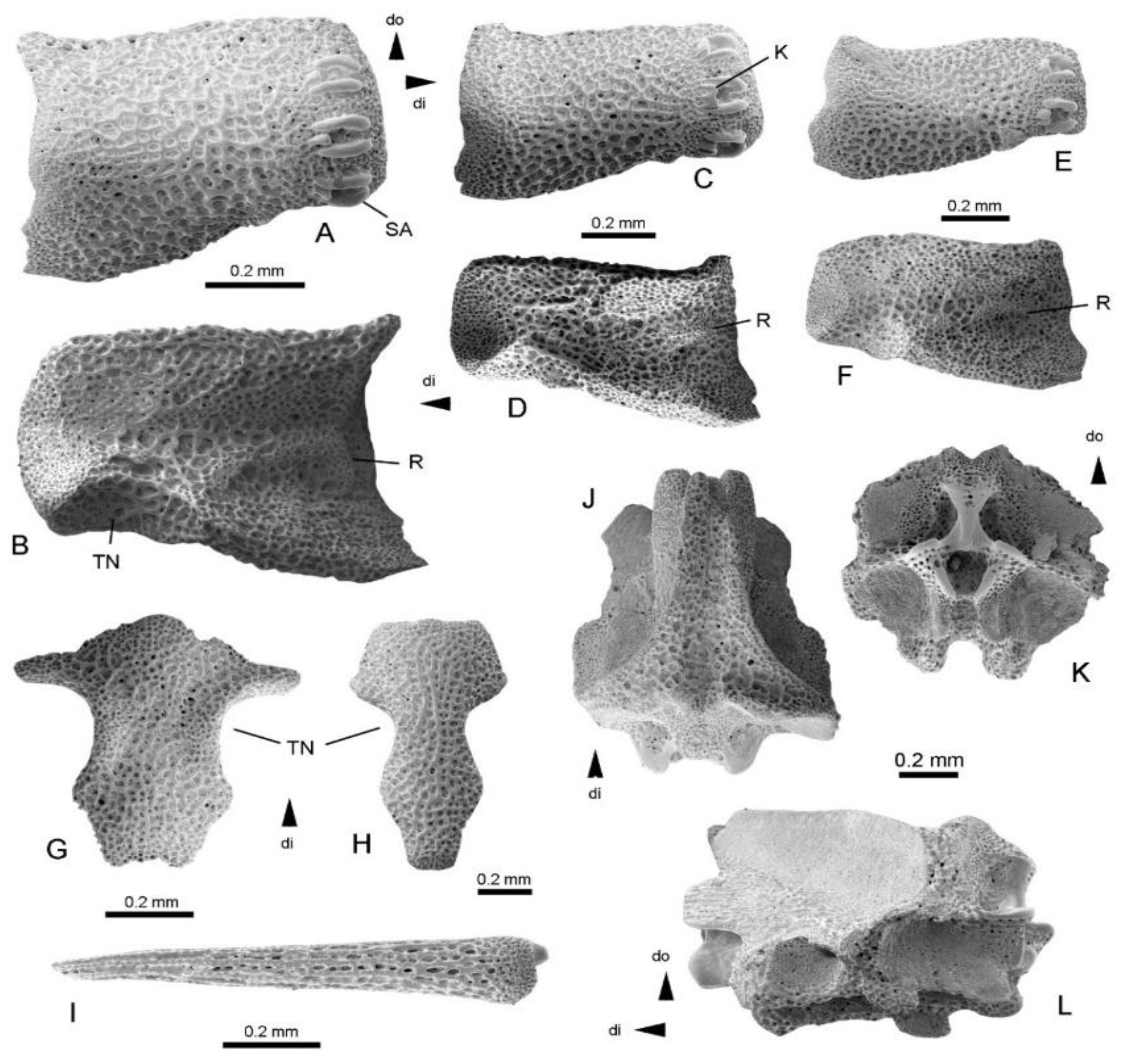
Figure 3.
Ophiura tankardi sp. nov. from the Rupelian, Oligocene (Rosenheim Subformation, Bodenheim Formation, nannoplankton zone NP 24 and to dinoflagellate zones D 14/D 15) of a flush drill made at Nierstein, Rhineland-Palatinate, Germany. Holotype (NHMMZ PWL 2022/5560.1-LS), dissociated proximal lateral arm plate in external (A) and internal (B) views; paratype (NHMMZ PWL 2022/5560.2-LS), dissociated median lateral arm plate in external (C) and internal (D) views; paratype (NHMMZ PWL 2022/5560.3-LS), dissociated distal lateral arm plate in external (E) and internal (F) views; paratype (NHMMZ PWL 2022/5560.4-LS), dissociated proximal vertebra in lateral (G), ventral (H) and distal (I) views; paratype (NHMMZ PWL 2022/5560.5-LS), dissociated proximal dorsal arm plate (J) in external view; paratype (NHMMZ PWL 2022/5560.6-LS), dissociated oral plate in adradial (K) and adradial (L) views; paratype (NHMMZ PWL 2022/5560.7-LS), distal arm fragment in lateral view (M) and with detail of arm spines (N). Abbreviations: AdMA: adradial muscle attachment area; AS: arm spine; di: distal; do: dorsal; PB: podial basin; R: vertebral articular ridge; SA: spine articulation; TN: tentacle notch.
Figure 3.
Ophiura tankardi sp. nov. from the Rupelian, Oligocene (Rosenheim Subformation, Bodenheim Formation, nannoplankton zone NP 24 and to dinoflagellate zones D 14/D 15) of a flush drill made at Nierstein, Rhineland-Palatinate, Germany. Holotype (NHMMZ PWL 2022/5560.1-LS), dissociated proximal lateral arm plate in external (A) and internal (B) views; paratype (NHMMZ PWL 2022/5560.2-LS), dissociated median lateral arm plate in external (C) and internal (D) views; paratype (NHMMZ PWL 2022/5560.3-LS), dissociated distal lateral arm plate in external (E) and internal (F) views; paratype (NHMMZ PWL 2022/5560.4-LS), dissociated proximal vertebra in lateral (G), ventral (H) and distal (I) views; paratype (NHMMZ PWL 2022/5560.5-LS), dissociated proximal dorsal arm plate (J) in external view; paratype (NHMMZ PWL 2022/5560.6-LS), dissociated oral plate in adradial (K) and adradial (L) views; paratype (NHMMZ PWL 2022/5560.7-LS), distal arm fragment in lateral view (M) and with detail of arm spines (N). Abbreviations: AdMA: adradial muscle attachment area; AS: arm spine; di: distal; do: dorsal; PB: podial basin; R: vertebral articular ridge; SA: spine articulation; TN: tentacle notch.

Holotype: NHMMZ PWL 2022/5560.1-LS.
Paratypes: NHMMZ PWL 2022/5560.2-LS—NHMMZ PWL 2022/5560.7-LS
Other material: 23 dissociated ossicles (not registered).
Type horizon and locality: Rosenheim Subformation, Bodenheim Formation, nannoplankton zone NP 24 and to dinoflagellate zones D 14/D 15, Rupelian, Oligocene, Cenozoic (33.9–28.1 million years before present); sample taken at 46 meters depth from flush drill made at Nierstein marketplace (WGS84 49°52′29.5” N/8°2 0′11” E), Rhineland Palatinate, Mainz Basin, Germany.
Etymology: Species named in honor of German thrash metal band Tankard, to celebrate their loyal and consistent connection to their fan base.
Diagnosis: Small species of Ophiura with fragile lateral arm plates of oblique trapezoidal outline, with a well-developed, fine vertical striation on the outer surface, three small spine articulations on the same level as the outer surface stereom, with a distalwards-pointing tentacle notch in proximal to distal arm segments, and with a vertebral articular ridge on the inner side composed of two well-defined knobs connected with a thin, poorly defined vertical ridge.
Description: Holotype specimen (NHMMZ PWL 2022/5560.1-LS), dissociated proximal lateral arm plate (Figure 3A,B), small, fragile, approximately as high as long, of trapezoidal outline, with a straight, oblique dorsal edge, a straight, horizontal ventral edge with a ventralwards-pointing protrusion close to the ventro-proximal tip, a deeply incised distal edge, and an irregularly wavy proximal edge; outer surface with a well-developed, fine, vertical striation replaced by fine tubercles on the ventral portion of the outer surface and close to the proximal edge; narrow band of finely meshed stereom along the outer proximal edge of the lateral arm plate, devoid of spurs, but with a small, poorly defined area of horizontally striated stereom; three small spine articulations in the dorsal half of the outer distal edge, at the same level as outer surface stereom and separated from the distal edge by a wide ledge of coarsely-meshed stereom; spine articulations vertically elongate, composed of large muscle opening proximally bordered by a thin, crenulated, strongly bent ridge, and distally separated from a smaller nerve opening by a thick, smooth, lip-shaped vertical ridge; spine articulations nearly equal-sized and equidistant; and very deep, conspicuous, distalwards -pointing tentacle notch, ventrally bordered by smoother stereom. Inner side of lateral arm plate with a vertical vertebral articular ridge composed of a very thin, poorly defined central part, a large, well-defined, prominent ventral knob composed of more finely-meshed stereom, and a similar but much smaller dorsal knob; stereom on inner side coarsely meshed except for overlap area along the inner distal edge, and along the proximal edge.
Paratype supplements and variation: dissociated median lateral arm plate (NHMMZ PWL 2022/5560.2-LS) (Figure 3C,D), longer than high, of rectangular outline, dorsal edge straight, ventral edge slightly concave and with a pointed ventralward protrusion close to the ventro-proximal tip; proximal edge lined by a narrow band of more finely-meshed stereom, with well-developed, fine horizontal striation and a very weak, poorly-defined, slightly protruding central spur; outer surface striation as in holotype; three spine articulations similar to those of holotype but smaller and in a slightly more ventral position; tentacle notch on distal edge smaller and shallower than in holotype; inner side of lateral arm plate with a single, triangular, sharply-defined vertebral articular structure composed of more finely-meshed stereom, with a poorly defined, dorsalward extension; stereom on inner side coarsely meshed except for band of more finely-meshed stereom in overlap area along the inner distal edge; and well-developed dorsal and ventral contact surfaces with opposite lateral arm plate.
Dissociated distal lateral arm plate (NHMMZ PWL 2022/5560.3-LS) (Figure 3E,F), approximately 2.5 times longer than high, with concave dorsal and ventral edges; proximal edge lined by a very narrow band with a fine horizontal striation, and with a well-developed, prominent and protruding spur; outer surface striation as in holotype; two spine articulations on outer distal edge of lateral arm plate, similar to those of holotype but in a much more ventral position; inner side of lateral arm plate with two horizontally elongate knobs, well defined, prominent, composed of more finely meshed stereom and arranged in a horizontal line; stereom on inner side of lateral arm plate coarsely meshed, except for vertically striated dorsal and ventral edges and band of more finely meshed stereom along inner distal edge; and tentacle notch deep, sharply defined, pointing distalwards; well-developed dorsal contact surface with opposite lateral arm plate.
Dissociated vertebra (NHMMZ PWL 2022/5560.4-LS), very slender, with a deeply incised middle saddle (Figure 3G), and with conspicuous, enlarged, fragile dorso-distal muscle fossae; small, well-defined, laterally pointing tentacle basins (Figure 3G,H); distal articular facets with small, slender, dorsalwards converging zygocondyles and a large, lozenge-shaped zygosphene (Figure 3I); and ventro-distal muscle fossae round and fragile.
Dissociated dorsal arm plate (NHMMZ PWL 2022/5560.5-LS) fragile, fan-shaped, with a convex distal edge, a pointed proximal edge, and a finely tuberculated outer surface (Figure 3J).
Dissociated oral plate (NHMMZ PWL 2022/5560.6-LS) split in two halves (Figure 3K,L), originally longer than high, devoid of lateral wings, with adradial muscle attachment area ventrally lining the adradial articulation area (Figure 3K).
Distal arm fragment (NHMMZ PWL 2022/5560.7-LS) composed of two articulated arm segments (Figure 3M), strongly elongate, with arm spines in place (Figure 3N), adpressed against arm segment, short, pointed, with a rugose, irregular, longitudinal striation.
Remarks: The material described above originates from a sample taken at 46 meters depth from the Nierstein marketplace flush drill, yielding only lateral arm plates of the same species. We, therefore, assume that all the associated skeletal plates, in particular the vertebrae, dorsal arm plates, and oral plates, belong to the same species. Similarities in relative size, fragile overall morphology, outer surface ornamentation (dorsal arm plate), and lateral articular structures (vertebrae) corroborate that the lateral arm plates and all the associated skeletal parts belong to the same species. Additional material not figured in the present paper was found in samples taken at 34, 42, and 50 m depth in the flush drill.
The general outline of the lateral arm plates, in particular with respect to the relative size of the ventral portion, the outer surface ornamentation, the shape and position of the spine articulations, the shape of the vertebral articular ridges and knobs on the inner side, and the shape of the distal arm spines suggests assignment to the genus Ophiura within the family Ophiuridae. Ishida et al. [19] recently revised the fossil record of Ophiura, summarizing the species once assigned to Ophiura that have since been transferred to other genera, and revising the remaining fossil species of Ophiura. Apart from fossil reports of extant species, e.g., Ophiura sarsii sarsii Lütken, 1855 from the middle Miocene to the Pleistocene of Japan [20,21,22,23,24,25,26,27,28,29], only the following extinct species have been retained as probably belonging to Ophiura: Ophiura achatae Rasmussen, 1972 from the upper Danian (lower Paleocene) Agatdal Formation of Nugssuaq (Greenland) [30], Ophiura wetherelli Forbes, 1852 from the Lower Eocene (Ypresian) London Clay of Highgate (London, UK) [31], Ophiura furiae Rasmussen, 1972 from the Lower Eocene (Ypresian) Mo-Clay Formation of the Island of Fur (Denmark) [30], Ophiura bognoriensis Rasmussen, 1972 from the Lower Eocene (Ypresian) Astarte Bed and Starfish Bed (Lower Clay) of Bognor Regis (Sussex, England) [30], Ophiura hendleri Blake and Aronson, 1998 from the Upper Eocene La Meseta Formation of Seymour Island (Antarctic Peninsula) [32], Ophiura bartonensis Rasmussen, 1972 from the Upper Eocene (Bartonian) Lower Barton Beds of Hampshire (England) [30], Ophiura? sternbergica Kutscher, 1980 from the Oligocene ‘Sternberger Gestein’ of Magdeburg (eastern Germany) [11], Ophiura marylandica Berry, 1934 from the Miocene Mary’s Member of Maryland (USA) [33], Ophiura paucilepis Stöhr, Jagt and Klompmaker, 2011 from the lower Zanclean to mid-Piacenzian (Pliocene) Oosterhout Formation of Mill-Langenboom (Noord-Brabant, the Netherlands) [34], and Ophiura pohangensis Ishida et al., 2022 from the Middle Miocene Duho Formation (Yeonil Group) of Pohang City, (Korea) [19]. None of these species have lateral arm plates comparable to those of the species described in the present paper, especially with respect to the small size, the oblique dorsal edge, the fragile architecture, and the fine vertical striation.
Among the recent species of Ophiura, the closest similarities are shared with the small species showing fragile, elongated arm segments and vertically striated lateral arm plates, in particular Ophiura ooplax (H.L. Clark, 1911), Ophiura trimeni Bell, 1905, Ophiura ljungmani (Lyman, 1878), and Ophiura grubei Heller, 1863. The former two differ from the material described in the present paper in having larger spine articulations with a dorsalward increase in size, and in having tentacle openings transformed into perforations at least from median arm segments onwards. Ophiura ljungmani and O. grubei differ from the Oligocene specimens in having a much weaker outer surface striation and a differently shaped vertebral articular structure on the inner side. In the light of these differences, we assign the material described herein to the new species Ophiura tankardi. The small size, the elongate arm segments and the elongate, divided oral plate suggest that Ophiura tankardi is a paedomorphic species.
Superorder Ophintegrida O’Hara, Hugall, Thuy, Stöhr & Martynov, 2017
Order Amphilepida O’Hara, Hugall, Thuy, Stöhr & Martynov, 2017
Suborder Ophionereidina O’Hara, Hugall, Thuy, Stöhr & Martynov, 2017
Superfamily Ophionereidoidea Ljungman, 1867
Family Ophionereididae Ljungman, 1867
Genus Ophiodoris Koehler, 1904
Diagnosis (emended and specifically focusing on lateral arm plate morphology): small, relatively elongate lateral arm plates with a coarsely meshed outer surface stereom devoid of tubercles or otherwise modified trabecular intersections; up to three large spine articulations with massive dorsal and ventral lobes proximally separated by single or multiple wedge-shaped knobs; and inner side with a thin, J-shaped, almost horizontal vertebral articular ridge.
Ophiodoris niersteinensis sp. nov. (Figure 4)
urn:lsid:zoobank.org:act:0E2FF12D-6D61-451D-B0BC-BD42FD597E05
Holotype: NHMMZ PWL 2022/5561.1-LS.
Paratypes: NHMMZ PWL 2022/5561.2-LS—NHMMZ PWL 2022/5561.7-LS
Other material: 8 dissociated ossicles (not registered).
Type horizon and locality: Rosenheim Subformation, Bodenheim Formation, nannoplankton zone NP 24 and to dinoflagellate zones D 14/D 15, Rupelian, Oligocene, Cenozoic (33.9–28.1 million years before present); sample taken at 42 meters depth from flush drill made at Nierstein marketplace (WGS84 49°52′29.5″ N/8°2 0′11″ E), Rhineland Palatinate, Mainz Basin, Germany.
Etymology: Species named after the type locality Nierstein.
Diagnosis: Small species of Ophiodoris with relatively elongate lateral arm plates, showing a coarsely meshed outer surface stereom devoid of tubercles; up to three spine articulations composed of smooth, imperforate dorsal and ventral lobes proximally separated by single, double or triple wedge-shaped knobs; and J-shaped vertebral articular ridge on the inner side.
Description: Holotype specimen (NHMMZ PWL 2022/5561.1-LS) is a small, dissociated proximal lateral arm plate (Figure 4A,B), slightly longer than high, with a weakly concave dorsal edge, an evenly convex distal edge, an oblique, ventro-proximalwards protruding ventral portion with a large but shallow and poorly defined tentacle notch, and a wavy proximal edge lined by a narrow, poorly-defined band of more finely meshed stereom and comprising a slightly swollen, protruding central part; outer surface composed of coarsely meshed stereom devoid of tubercles; three large, freestanding spine articulations on distal edge, composed of smooth, imperforate, nearly parallel dorsal and lobes proximally separated by a single, wedge-shaped knob; dorsal-most spine articulation slightly smaller than the other two. Inner side of lateral arm plate with a conspicuous J-shaped vertebral articular structure, moderately well defined, composed of slightly more finely meshed stereom and almost horizontal; no perforation discernible; inner distal edge tapering, composed of more finely meshed stereom, devoid of spurs; and inner side of tentacle notch shallow but large, conspicuous, bordered by a thickened part of the ventral edge.
Figure 4.
Ophiodoris niersteinensis sp. nov. from the Rupelian, Oligocene (Rosenheim Subformation, Bodenheim Formation, nannoplankton zone NP 24 and to dinoflagellate zones D 14/D 15) of a flush drill made at Nierstein, Rhineland-Palatinate, Germany. Holotype (NHMMZ PWL 2022/5561.1-LS), dissociated proximal lateral arm plate in external (A) and internal (B) views; paratype (NHMMZ PWL 2022/5561.2-LS), dissociated median lateral arm plate in external (C) and internal (D) views; paratype (NHMMZ PWL 2022/5561.3-LS), dissociated distal lateral arm plate in external (E) and internal (F) views; paratype (NHMMZ PWL 2022/5561.4-LS), dissociated proximal ventral arm plate (G); paratype (NHMMZ PWL 2022/5561.5-LS), dissociated median to distal ventral arm plate (H); paratype (NHMMZ PWL 2022/5561.6-LS), dissociated arm spine (I); paratype (NHMMZ PWL 2022/5561.7-LS), dissociated proximal vertebra in dorsal (J), proximal (K) and lateral (L) views. Abbreviations: di: distal; do: dorsal; K: wedge shaped knob; R: vertebral articular ridge; SA: spine articulation; TN: tentacle notch.
Figure 4.
Ophiodoris niersteinensis sp. nov. from the Rupelian, Oligocene (Rosenheim Subformation, Bodenheim Formation, nannoplankton zone NP 24 and to dinoflagellate zones D 14/D 15) of a flush drill made at Nierstein, Rhineland-Palatinate, Germany. Holotype (NHMMZ PWL 2022/5561.1-LS), dissociated proximal lateral arm plate in external (A) and internal (B) views; paratype (NHMMZ PWL 2022/5561.2-LS), dissociated median lateral arm plate in external (C) and internal (D) views; paratype (NHMMZ PWL 2022/5561.3-LS), dissociated distal lateral arm plate in external (E) and internal (F) views; paratype (NHMMZ PWL 2022/5561.4-LS), dissociated proximal ventral arm plate (G); paratype (NHMMZ PWL 2022/5561.5-LS), dissociated median to distal ventral arm plate (H); paratype (NHMMZ PWL 2022/5561.6-LS), dissociated arm spine (I); paratype (NHMMZ PWL 2022/5561.7-LS), dissociated proximal vertebra in dorsal (J), proximal (K) and lateral (L) views. Abbreviations: di: distal; do: dorsal; K: wedge shaped knob; R: vertebral articular ridge; SA: spine articulation; TN: tentacle notch.

Paratype supplements and variation: dissociated median lateral arm plate (NHMMZ PWL 2022/5561.2-LS) (Figure 4C,D), 1.5 times longer than high, similar to holotype, except for spine articulations showing a double or triple knob proximally separating the dorsal and ventral lobes; inner side as in holotype but with very poorly defined vertebral articular ridge and smaller tentacle notch showing a ventralward pointing protrusion.
Dissociated distal lateral arm plate (NHMMZ PWL 2022/5561.3-LS) (Figure 4E,F), almost two times longer than high, with a strongly concave dorsal edge and a wavy distal edge with a narrow band of more finely meshed stereom; outer surface stereom as in holotype; two spine articulations with dorsal and ventral lobes proximally separated by a double knob; and inner side as in paratype specimen.
Dissociated proximal ventral arm plate (NHMMZ PWL 2022/5561.4-LS), approximately as long as wide, of bullhead-shaped outline, with an obtuse distal angle, conspicuous pointed lateral protrusions, deep lateral tentacle notches, and a straight proximal edge; and outer surface with coarsely meshed stereom, thickened around the tentacle notches (Figure 4G).
Dissociated median to distal ventral arm plate (NHMMZ PWL 2022/5561.5-LS), almost two times longer than wide, with a straight distal edge, pointed lateral edges, deep lateral tentacle notches and a straight proximal edge; and outer surface stereom coarsely meshed (Figure 4H).
Dissociated proximal vertebra (NHMMZ PWL 2022/5561.7-LS), relatively compact except for very large dorso-distal muscle fossae, forming a distalwards pointing, almost keel-like protrusion (Figure 4J); ventro-proximal muscle fossae deep, large; ventro-distal and dorso-proximal muscle fossae smaller (Figure 4K,L); lateral articular structure J-shaped, poorly defined; ventral furrow large and deep; dorsal furrow large, with lateral extensions paralleling the edge of the dorso-distal muscle fossae, devoid of dorso-proximal depression; vertebral articulation zygospondylous; and podial basins well-defined, ventro-distalwards pointing.
Dissociated arm spine (NHMMZ PWL 2022/5561.6-LS) long, cylindrical, pointed, with a coarsely meshed, irregular longitudinal striation beset with minute thorns (Figure 4I).
Remarks: The lateral arm plates described above belong to the family Ophionereididae on account of their outline with the large, ventro-distalwards protruding ventral portion, and the large spine articulations composed of parallel dorsal and ventral lobes proximally separated by single or multiple knobs. Comparison with the lateral arm plates of recent ophionereids suggests that similarities are greatest with the lateral arm plates of Ophiodoris owing to the plate outline, the shape of the dorsal and ventral lobes, and the shape of the vertebral articular ridge on the inner side of the lateral arm plate [35,36,37].
The fossil record of Ophiodoris is very sparse so far, with Ophiodoris reconciliator Thuy, 2015 and Ophiodoris francojurassicus (Hess, 1975) from the Jurassic of France and Switzerland, recpectively [35,38], Ophiodoris holterhoffi Thuy, Gale, Stöhr & Wiese, 2014 from the Cretaceous of Texas, USA [35], and an unnamed species from the Pliocene of the Mediterranean [37] as the only known records so far. Unambiguous fossil occurrences of the closely related genera Ophioplax Lyman, 1875 and Ophiochiton Lyman, 1878 are rare too [39]. Comparison with these fossil reports and with recent species of Ophiodoris shows that the material described above stands out in having a coarsely meshed stereom devoid of tubercles on the outer surface, and in having an almost horizontal, conspicuously J-shaped vertebral articular ridge on the inner side. We, therefore, introduce the new species Ophiodoris niersteinensis sp. nov. to accommodate the material in question.
4. Discussion
The material described in the present paper confirms the considerable potential of microfossils to study the fossil record of the Ophiuroidea. Furthermore, it shows that even well-studied areas, such as the Mainz Basin, can yield unknown species. With the material described in the present paper, we add two new species to the poorly known Oligocene fossil record of the ophiuroids. Both species described herein belong to extant genera. In the case of Ophiodoris, the new Oligocene species bridges a considerable stratigraphic gap between the Early Cretaceous and the Plio-Pleistocene records of the genus [35,37].
Ophiura has a wide geographical and bathymetric distribution in the present-day oceans [40]. Ophiodoris, in contrast, is mostly restricted to deep sublittoral to shallow bathyal depths [37,40]. Assuming that the distribution preferences of the genus can be extrapolated to the Oligocene, the occurrence of Ophiodoris in the deeper offshore part of the Mainz Basin corroborates deep sublittoral to shallow bathyal paleodepths for the concerned area.
The present paper describes the ophiuroid species found in a particular part of the Mainz Basin. It seems very likely that other parts of the Basin yield additional ophiuroid species. Ongoing investigations have confirmed the presence of yet unnamed amphiurid ophiuroids in shallower parts of the Basin [12]. We aim to systematically describe these additional ophiuroids in forthcoming works.
Author Contributions
Conceptualization, B.T. and L.D.N.-T.; methodology, B.T. and L.D.N.-T.; validation, B.T., L.D.N.-T. and K.N.; formal analysis, B.T.; investigation, B.T. and K.N.; writing—original draft preparation, B.T. and K.N.; writing—review and editing, B.T., L.D.N.-T. and K.N.; visualization, B.T., L.D.N.-T. and K.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Therra Therm Erdwärme GmbH for taking the flush drill samples and donating them to the Nierstein Museum, and the late Arnulf Stapf of Nierstein Museum for making the samples available for study. We also thank Przemyslaw Gorzelak and an anonymous reviewer whose comments greatly improved an earlier version of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stöhr, S.; O’Hara, T.D.; Thuy, B. Global diversity of brittle stars (Echinodermata: Ophiuroidea). PLoS ONE 2012, 7, e31940. [Google Scholar] [CrossRef]
- Hess, H. Mikropaläontologische Untersuchungen an Ophiuren. I. Einleitung. Eclogae Geol. Helv. 1962, 55, 595–608. [Google Scholar]
- Hess, H. Mikropaläontologische Untersuchungen an Ophiuren. II. Die Ophiuren aus dem Lias (Pliensbachien-Toarcien) von Seewen (Kt. Solothurn). Eclogae Geol. Helv. 1962, 55, 609–656. [Google Scholar]
- Thuy, B. Temporary expansion to shelf depths rather than an onshore-offshore trend: The shallow-water rise and demise of the modern deep-sea brittle star family Ophiacanthidae (Echinodermata: Ophiuroidea). Eur. J. Taxon. 2013, 48, 1–242. [Google Scholar] [CrossRef] [Green Version]
- Thuy, B.; Stöhr, S. Lateral arm plate morphology in brittle stars (Echinodermata: Ophiuroidea): New perspectives for ophiuroid micropalaeontology and classification. Zootaxa 2011, 3013, 1–47. [Google Scholar] [CrossRef] [Green Version]
- Martynov, A. Reassessment of the classification of the Ophiuroidea (Echinodermata), based on morphological characters. I. General character evaluation and delineation of the families Ophiomyxidae and Ophiacanthidae. Zootaxa 2010, 2697, 1–154. [Google Scholar] [CrossRef] [Green Version]
- Thuy, B.; Stöhr, S. A new morphological phylogeny of the Ophiuroidea (Echinodermata) accords with molecular evidence and renders microfossils accessible for cladistics. PLoS ONE 2016, 11, e0156140. [Google Scholar] [CrossRef] [PubMed]
- Thuy, B.; Stöhr, S. Unravelling the origin of the basket stars and their allies (Echinodermata, Ophiuroidea, Euryalida). Sci. Rep. 2018, 8, 8493. [Google Scholar] [CrossRef]
- Berry, C.T. An ophiuran from the Byram Marl (Oligocene) of Mississippi. J. Paleontol. 1937, 11, 235–240. [Google Scholar]
- Ishida, Y. Fossil ophiuroids from the Oligocene Asagai Formation of Iwaki, Fukushima, Japan. Bull. Nat. Sci. Mus. Ser. C 1992, 18, 65–78. [Google Scholar]
- Kutscher, M. Die Echinodermen des Oberoligozäns von Sternberg. Z. Geol. Wiss. 1981, 9, 221–239. [Google Scholar]
- Nungesser, K.; Talosi, B. Der Pfadberg bei Stadecken—Neue Funde von einer klassischen Lokalität, 1. Teil. Steinkern 2010, 4, 48–57. [Google Scholar]
- Schäfer, P. Mainzer Becken–Stratigraphie–Paläontologie–Exkursionen, 2nd ed.; Gebr. Borntraeger: Stuttgart, Germany, 2012; pp. 1–333. [Google Scholar]
- Grimm, K.I.; Grimm, M.C. Geologischer Führer durch das Mainzer Tertärbecken. In Die Fossilen Wirbellosen des Mainzer Tertiärbeckens, Teil 1-1; Grimm, K.I., Grimm, M.C., Eds.; Beiheft 26; Mainzer Naturwissenschaftliches Archiv: Mainz, Germany, 2003; pp. 1–158. [Google Scholar]
- Eck, K. Die Taphozönose des Fischschiefers (Rupelium/Oligozän) der Paläogen-Scholle “Wiesloch-Rot” (Baden-Württemberg/Deutschland)—Taxonomie, Taphonomie und Paläoökologie. Ph.D. Thesis, University of Heidelberg, Heidelberg, Germany, 2018. [Google Scholar]
- Grimm, K.I. Paläoökologie, Palaeogeographie und Stratigraphie im Mainzer Becken, im Oberrheingraben, in der Hessischen Senke und in der Leipziger Bucht während des Mittleren Rupeltons (Fischschiefer/Rupelium/Unteroligozän). Mitt. Pollichia 1994, 81, 7–193. [Google Scholar]
- Kammerer, T. Ostracoden aus dem Oberen Rupelton, Schleichsand und Cyrenenmergel (Zwischenschichten, Rupelium Bis Unteres Chattium) des Mainzer Beckens. Ph.D. Thesis, University of Mainz, Mainz, Germany, 1993. [Google Scholar]
- Nungesser, K. Das Tertiär des Mainzer Beckens—Erdgeschichte und Fossilien in Rheinhessen und Umgebung; Steinkern-Verlag: Bielefeld, Germany, 2021; pp. 1–224. [Google Scholar]
- Ishida, Y.; Thuy, B.; Nam, G.S.; Martynov, A.; Fujita, T.; Kim, J.-H. A new species of Ophiura (Echinodermata, Ophiuroidea) from Miocene deep-sea deposits in the Pohang Basin, Korea. Paleontol. Res. 2022, 26, 18–30. [Google Scholar] [CrossRef]
- Ishida, Y.; Takakuwa, Y.; Kamimura, H. Ophiura sarsii sarsii (Echinodermata, Ophiuroidea) from the latest middle Miocene to earliest late Miocene Itahana Formation, Gunma Prefecture, central Japan. Bull. Gunma Mus. Nat. Hist. 2004, 8, 21–26. [Google Scholar]
- Ishida, Y.; Nagamori, H.; Narita, K. Ophiura sarsii sarsii (Echinodermata, Ophiuroidea) from the late Miocene Ogawa Formation, Shinshushinmachi, Nagano Prefecture, Central Japan. Res. Rep. Shinshushinmachi Fossil Mus. 1998, 1, 9–16. [Google Scholar]
- Ishida, Y.; Nagasawa, K.; Abe, H. Ophiura sarsii sarsii (Echinodermata, Ophiuroidea) from the early Pliocene Noguchi and Nakawatari Formations in Yamagata Prefectue, northern Japan. Res. Rep. Foss. Sperm Whale Mamurogawa-Machi Yamagata Pref. 2002, 1, 125–136. [Google Scholar]
- Ishida, Y.; Nagasawa, K.; Tokairin, H. Ophiura sarsii sarsii (Echinodermata, Ophiuroidea) from the late Miocene to early Pliocene formations of Yamagata Prefecture, northern Japan. Earth Sci. 1999, 53, 223–232. [Google Scholar]
- Ishida, Y. Escape behavior and postures of epifaunal ophiuroids from burial: Extant and late Miocene Ophiura sarsii sarsii as an example. Memoirs Geol. Soc. Jpn. 1999, 54, 161–173, (In Japanese with English abstract). [Google Scholar]
- Ishida, Y. Ophiura sarsii sarsii (Echinodermata, Ophiuroidea) from the upper Pliocene Higashigawa Formation in Niigata Prefecture, central Japan. Bull. Fossa Magna Mus. 2004, 3, 17–26. [Google Scholar]
- Ishida, Y.; Fujita, T. Ophiura sarsii sarsii (Echinodermata, Ophiuroidea) from the late middle Miocene to early Pliocene formations of North Hokkaido, Japan. J. Geol. Soc. Japan 2001, 107, 784–793. [Google Scholar] [CrossRef]
- Ishida, Y.; Inoue, K. Fossil Ophiuroids (Ophiura sarsii Lütken, 1854) from the Pleistocene Ichijuku Formation (Kazusa Group), Chiba Prefecture, Central Japan. Nat. Hist. Res. 1993, 2, 103–112. [Google Scholar]
- Ishida, Y.; Inoue, K. Mode of occurrence and paleoecology of Ophiura sarsii Lütken, 1854–Fossil ophiuroids from the Pleistocene Ichijuku Formation (Kazusa Group), Chiba Prefecture, Central Japan. Earth Sci. 1995, 49, 32–44. [Google Scholar]
- Ishida, Y.; Kurita, Y. Ophiura sarsii sarsii (Echinodermata, Ophiuroidea) from the late Pliocene Hachioji Formation in Niigata Prefecture, Central Japan. Paleontol. Res. 1998, 2, 137–144. [Google Scholar]
- Rasmussen, H.W. Lower Tertiary Crinoidea, Asteroidea and Ophiuroidea from northern Europe and Greenland. Biol. Skr. Dan. Vid. Selsk. 1972, 19, 1–123. [Google Scholar]
- Forbes, E. Monograph of the Echinodermata of the British Tertiaries. Palaeontogr. Soc. Monogr. 1852, 6, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Blake, D.B.; Aronson, R.B. Eocene stelleroids (Echinodermata) at Seymour Island, Antarctic Peninsula. J. Paleontol. 1998, 72, 339–353. [Google Scholar] [CrossRef]
- Berry, C.T. Miocene and Recent Ophiuran skeletons. JHU Stud. Geol. 1934, 11, 9–136. [Google Scholar]
- Stöhr, S.; Jagt, J.W.M.; Klompmaker, A.A. Ophiura paucilepis, a new species of brittlestar (Echinodermata, Ophiuroidea) from the Pliocene of the southern North Sea Basin. Swiss J. Palaeontol. 2011, 130, 113–121. [Google Scholar] [CrossRef]
- Thuy, B.; Gale, A.S.; Stöhr, S.; Wiese, F. Shallow-water brittle-star (Echinodermata: Ophiuroidea) assemblages from the Aptian (Early Cretaceous) of the North Atlantic: First insights into bathymetric distribution patterns. Göttingen Contr. Geosci. 2014, 77, 163–182. [Google Scholar]
- Thuy, B. A peri-reefal brittle-star (Echinodermata, Ophiuroidea) assemblage from the Middle Jurassic of the northeast Paris Basin. Ferrantia Trav. Sci. Mus. Nat. Hist. Nat. Lux. 2015, 71, 87–106. [Google Scholar]
- Numberger-Thuy, L.D.; Thuy, B. Pliocene deep-sea ophiuroids from the Mediterranean with Western Atlantic affinities. In Progress in Echinoderm Palaeobiology; Zamora, S., Rábano, I., Eds.; Cuadernos del Museo Geominero: Madrid, Spain, 2015; Volume 19, pp. 125–127. [Google Scholar]
- Thuy, B.; Marty, D.; Comment, G. A remarkable example of a Late Jurassic shallow-water ophiuroid assemblage from the Swiss Jura Mountains. Swiss J. Geosci. 2013, 106, 409–426. [Google Scholar] [CrossRef]
- Ewin, T.; Thuy, B. Brittle stars from the British Oxford Clay: Unexpected ophiuroid diversity on Jurassic sublittoral mud bottoms. J. Paleontol. 2017, 91, 781–798. [Google Scholar] [CrossRef] [Green Version]
- GBIF Secretariat: GBIF Backbone Taxonomy. Available online: https://www.gbif.org/species/2277142 (accessed on 21 March 2022).
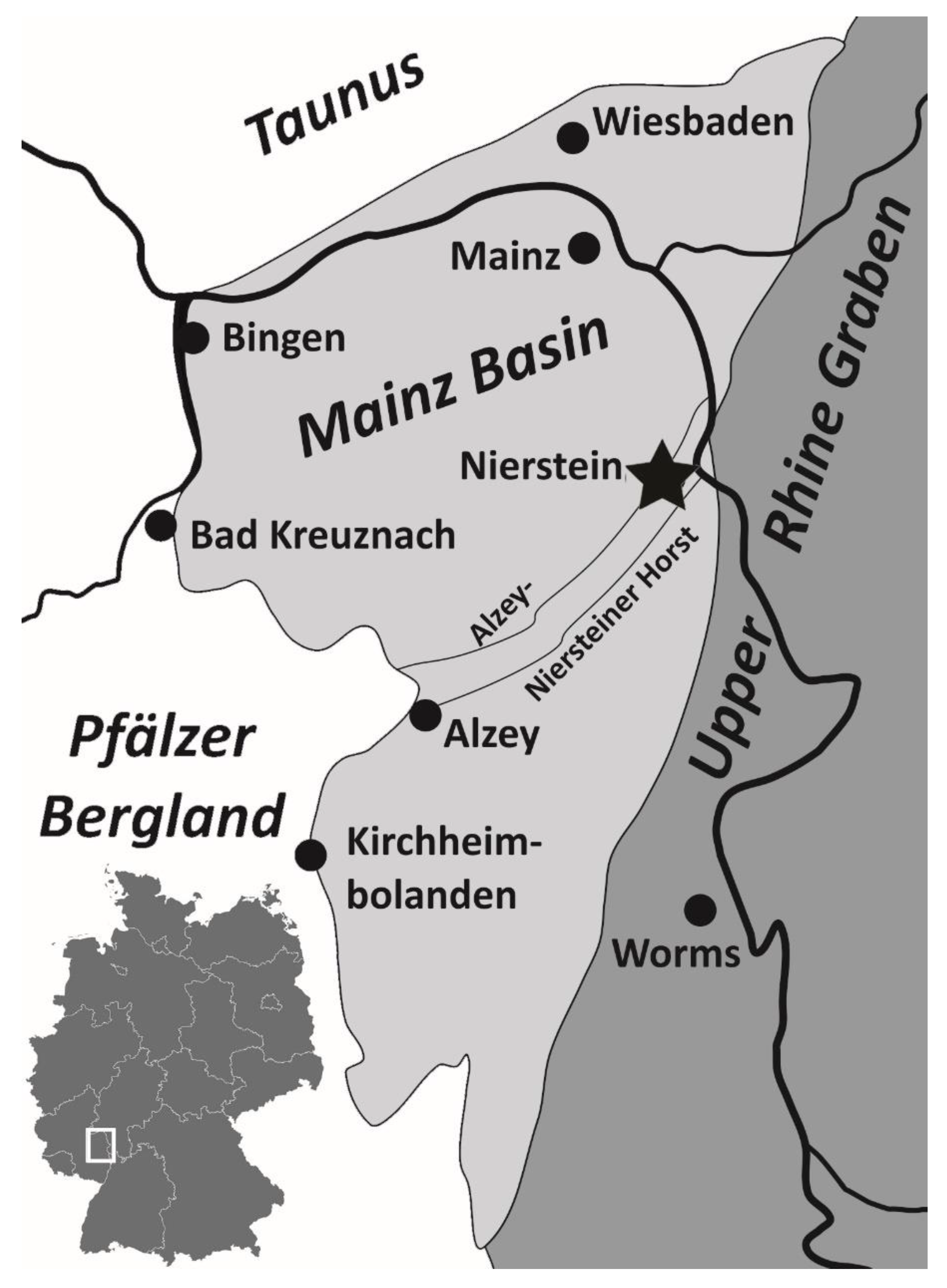
Figure 1.
Tectonic limits of the Mainz Basin, with sampling locality highlighted by a star, modified from Schäfer [13].
Figure 1.
Tectonic limits of the Mainz Basin, with sampling locality highlighted by a star, modified from Schäfer [13].

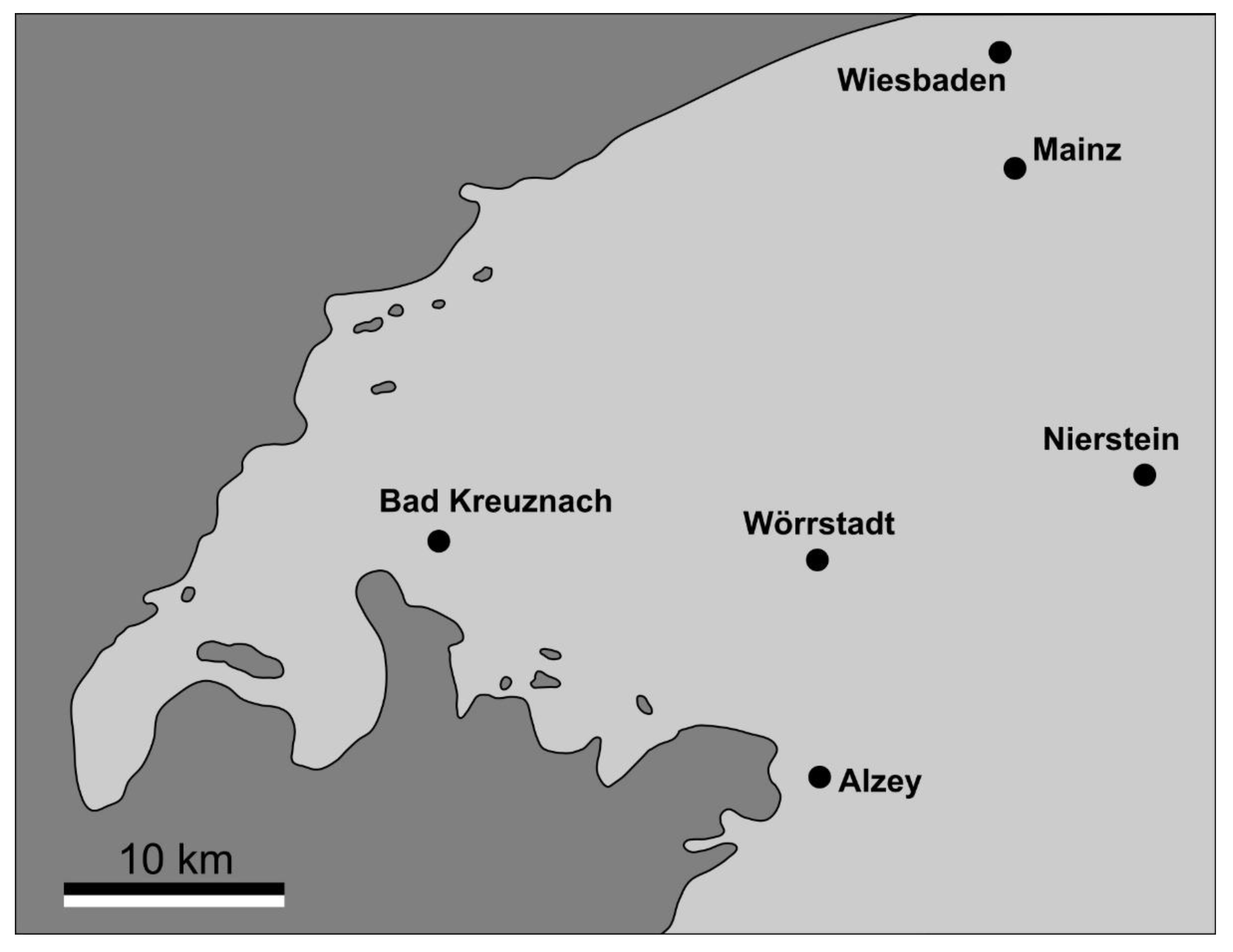
Figure 2.
Palaeogeography of the northern Mainz Basin at the time of deposition of the Rosenberg Subformation, with emerged land in dark grey and areas covered by sea in light grey, modified from Schäfer [13].
Figure 2.
Palaeogeography of the northern Mainz Basin at the time of deposition of the Rosenberg Subformation, with emerged land in dark grey and areas covered by sea in light grey, modified from Schäfer [13].

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Thuy, B.; Nungesser, K.; Numberger-Thuy, L.D. New Brittle Stars (Echinodermata, Ophiuroidea) from the Oligocene of the Mainz Basin, Germany. Taxonomy 2022, 2, 196-207. https://doi.org/10.3390/taxonomy2020015
AMA Style
Thuy B, Nungesser K, Numberger-Thuy LD. New Brittle Stars (Echinodermata, Ophiuroidea) from the Oligocene of the Mainz Basin, Germany. Taxonomy. 2022; 2(2):196-207. https://doi.org/10.3390/taxonomy2020015
Chicago/Turabian StyleThuy, Ben, Kai Nungesser, and Lea D. Numberger-Thuy. 2022. "New Brittle Stars (Echinodermata, Ophiuroidea) from the Oligocene of the Mainz Basin, Germany" Taxonomy 2, no. 2: 196-207. https://doi.org/10.3390/taxonomy2020015