The Effects of Biofertilizers on Growth, Soil Fertility, and Nutrients Uptake of Oil Palm (Elaeis Guineensis) under Greenhouse Conditions

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Experimental Design
2.2. Analysis of Soil Chemical Properties
2.3. Measurement of Oil Palm Seedlings Growth
2.4. Plants Analysis
2.5. Data Analysis
3. Results
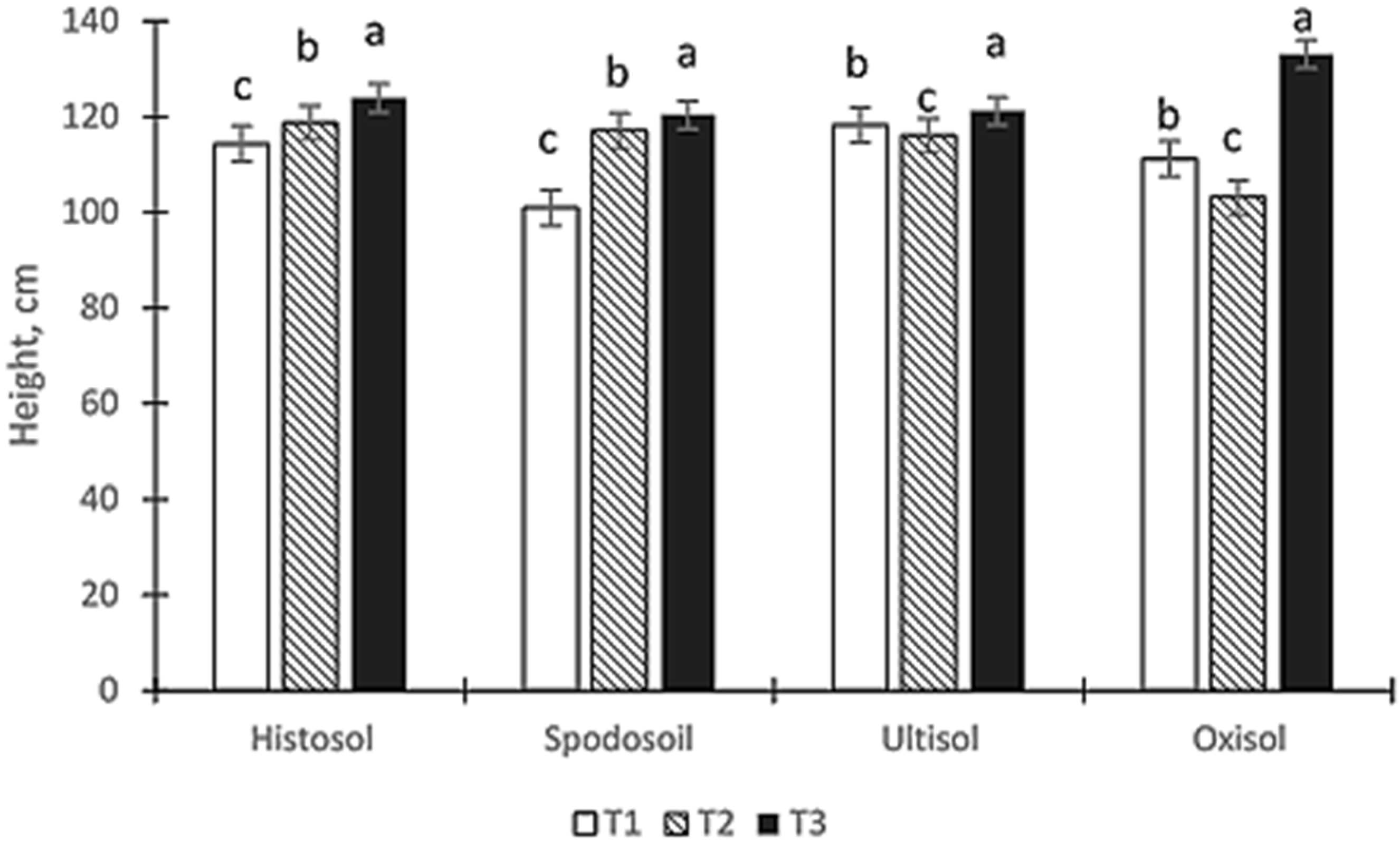
3.1. Growth Performance of Fronds
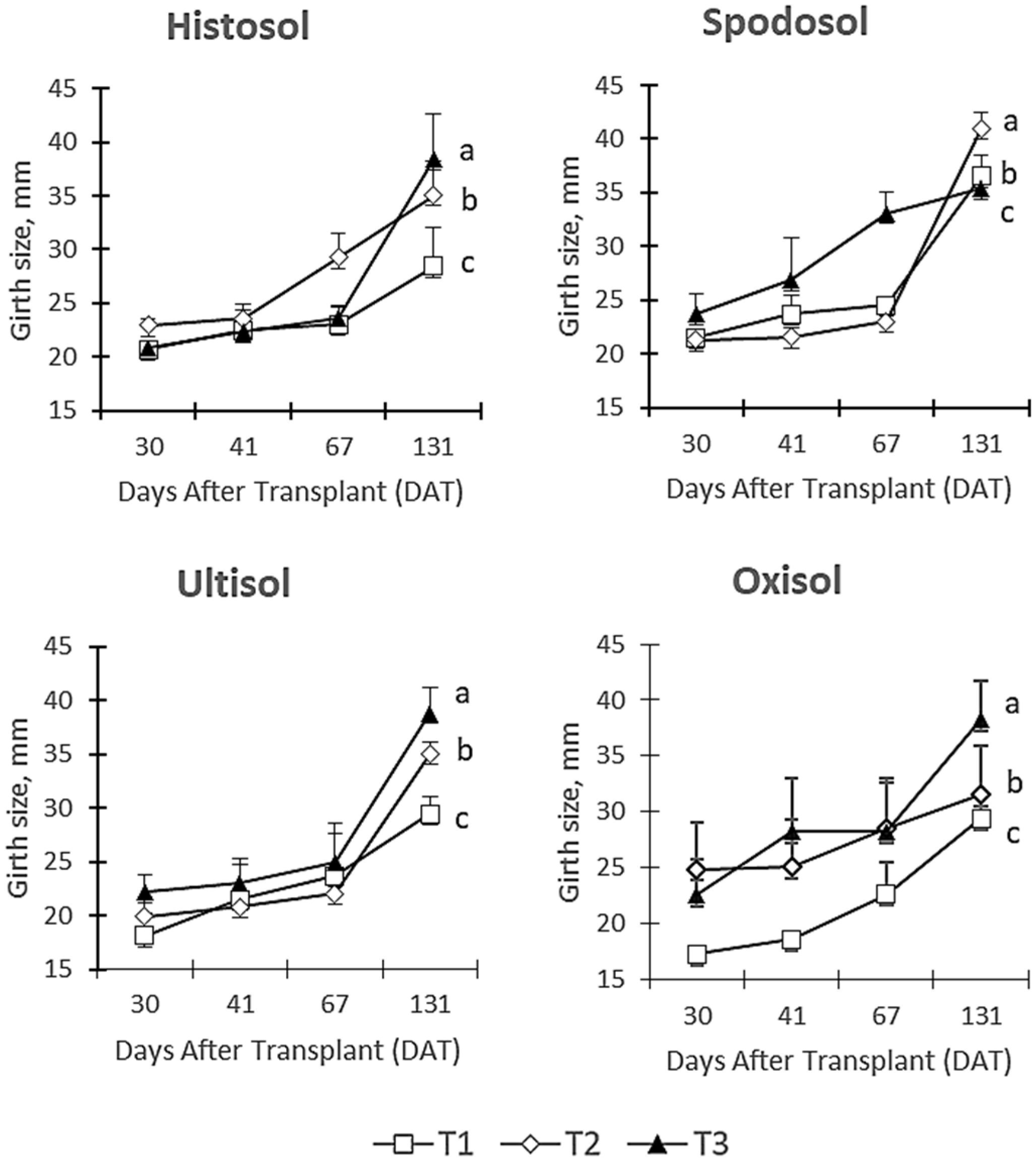
3.2. Stem Girth Size
3.3. Aboveground Biomass (ABG) and Root Dry Mass Ratio
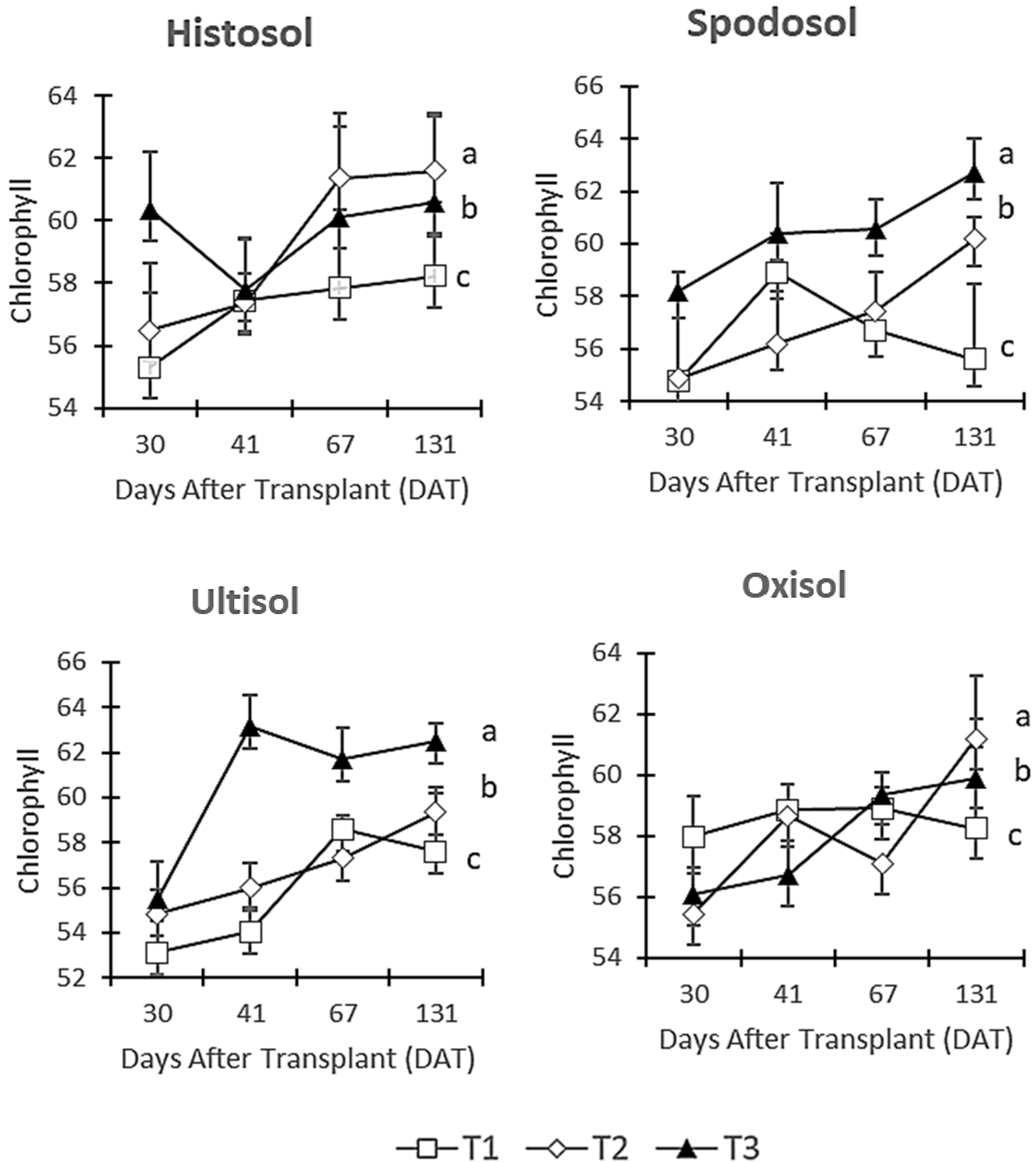
3.4. Chlorophyll Content
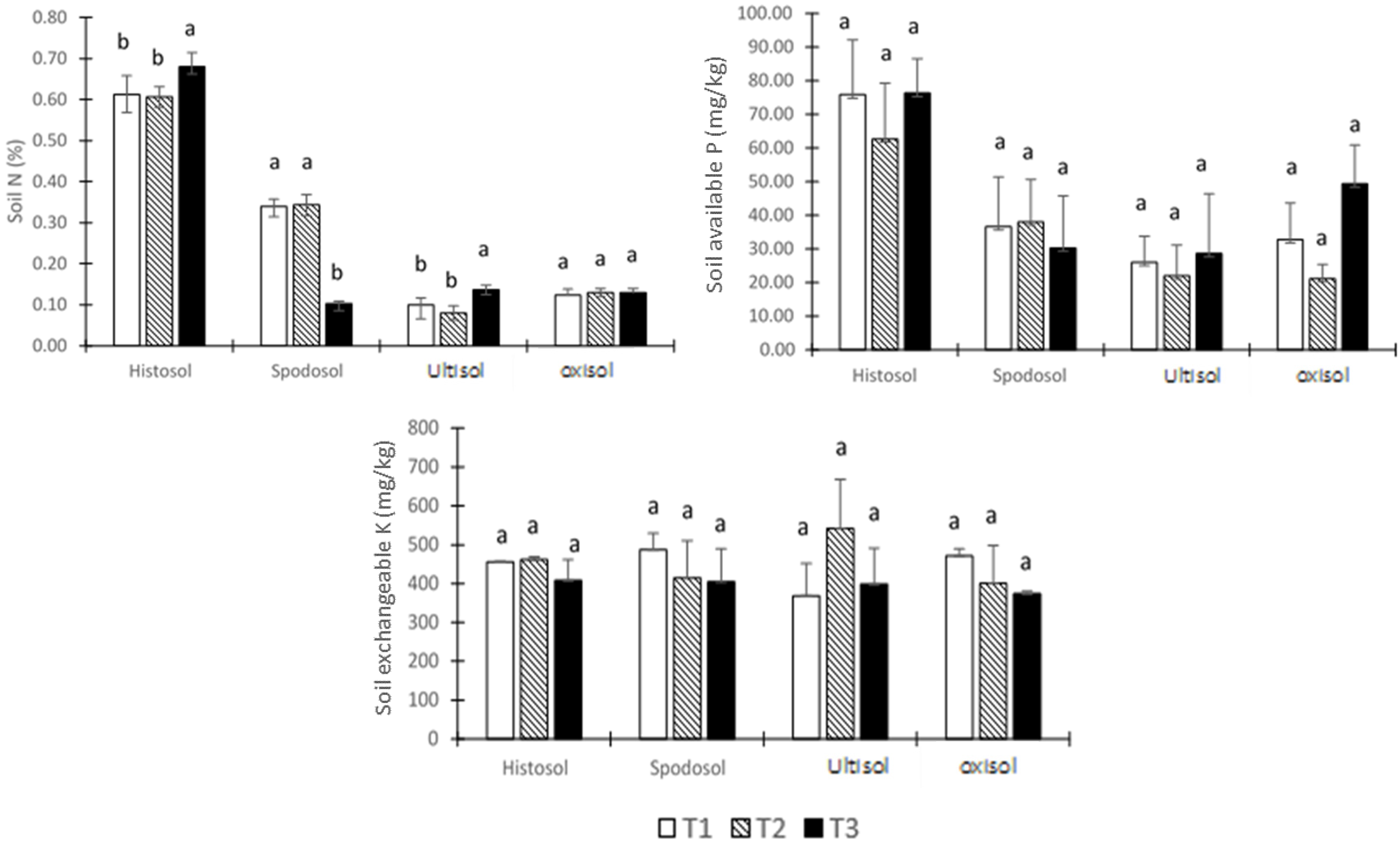
3.5. Soil Macronutrient Status
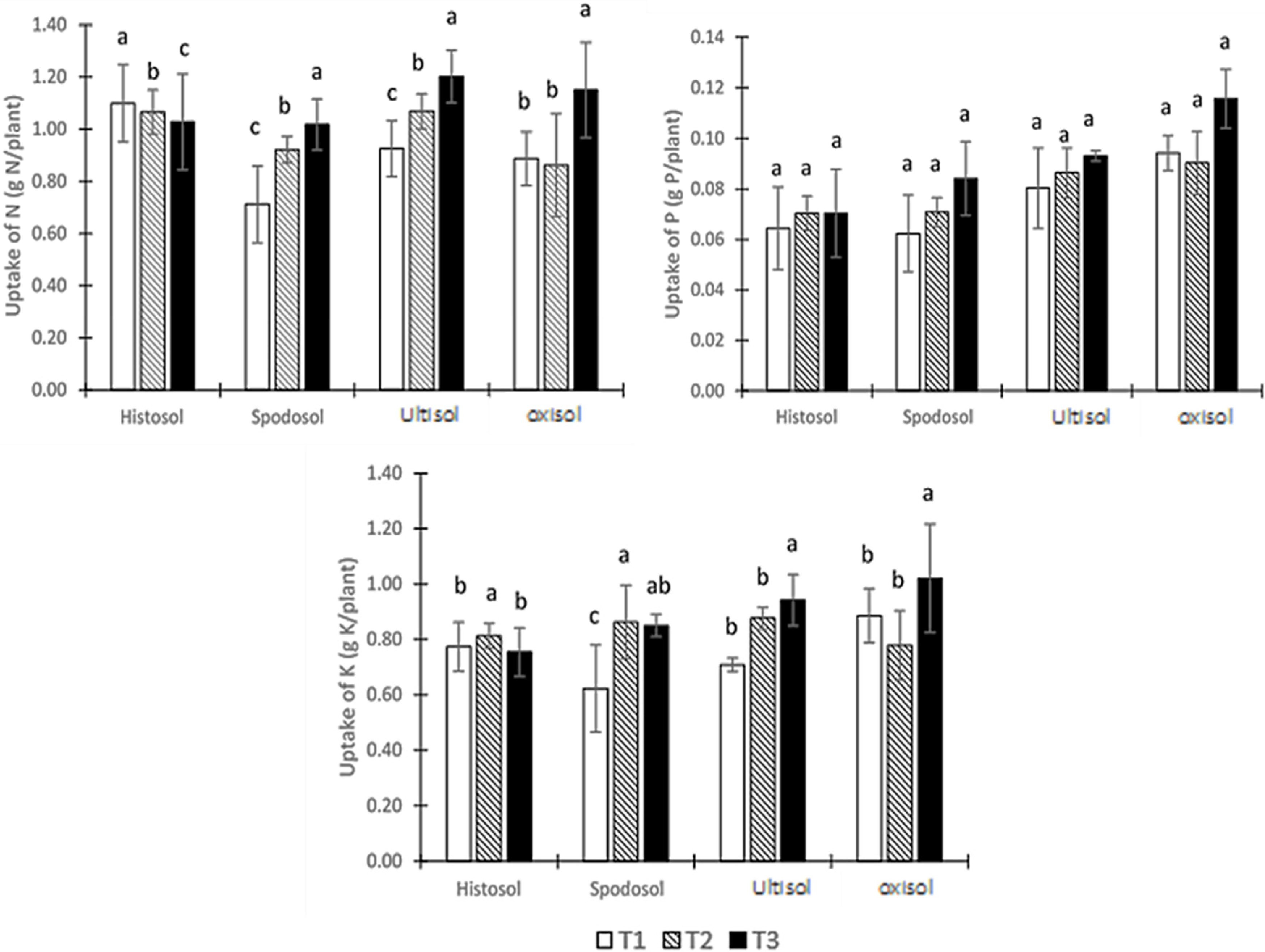
3.6. NPK Uptake by the Oil Palm Seedlings
4. Discussion
4.1. Growth Performance of Fronds
4.2. Stem Girth Size
4.3. Aboveground Biomass (ABG) and Root Dry Mass Ratio
4.4. Chlorophyll Content
4.5. Soil Macronutrient Status
4.6. NPK Uptake by the Oil Palm Seedlings
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hoffmann, U. Assuring food security in developing countries under the challenges of climate change: Key trade and development issues of a fundamental transformation of agriculture. In Proceedings of the United Nations Conference on Trade and Development, Geneva, Switzerland, 22 February 2011; pp. 35–40. [Google Scholar]
- Ward, M.H.; Brender, J.D. Drinking water nitrate and human health. In Encyclopedia of Environmental Health; Elsevier BV: Amsterdam, The Netherlands, 2019; pp. 173–186. [Google Scholar]
- Blankson, G.K.; Osei-Fosu, P.; Adeendze, E.; Ashie, D. Contamination levels of organophosphorus and synthetic pyrethroid pesticides in vegetables marketed in Accra, Ghana. Food Control 2016, 68, 174–180. [Google Scholar] [CrossRef]
- Domínguez, I.; Romero-González, R.; Liébanas, F.J.A.; Vidal, J.L.M.; Frenich, A.G. Automated and semi-automated extraction methods for GC–MS determination of pesticides in environmental samples. Trends Environ. Anal. Chem. 2016, 12, 1–12. [Google Scholar] [CrossRef]
- Wang, B.; Lai, T.; Huang, Q.-W.; Yang, X.-M.; Shen, Q.-R. Effect of N Fertilizers on Root Growth and Endogenous Hormones in Strawberry. Pedosphere 2009, 19, 86–95. [Google Scholar] [CrossRef]
- VoonKheong, L.; Rahman, Z.A.; Musa, M.H.; Hussein, A. Effects of severing oil palm roots on leaf nutrient levels and P uptake. J. Oil Palm Res. 2012, 24, 1343–1348. [Google Scholar]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factories 2014, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Itelima, J.U.; Bang, W.J.; Onyimba, I.A.; Sila, M.D.; Egbere, O.J. Bio-fertilizer as key player in enhancing soil fertility and crop productivity: A Review. Direct Res. J. Agric. Food Sci. 2018, 6, 73–83. [Google Scholar] [CrossRef]
- Zainuddin, N. Effect of biofertiliser containing different percentage rates of chemical fertiliser on oil palm seedlings. J. Oil Palm Res. 2019. [Google Scholar] [CrossRef]
- Naher, U.A.; Panhwar, Q.A.; Othman, R.; Shamshuddin, J.; Ismail, M.R.; Zhou, E. Proteomic study on growth promotion of PGPR inoculated aerobic rice (Oryza sativa L.) cultivar MR219-9. Pak. J. Bot. 2018, 50, 1843–1852. [Google Scholar]
- Vejan, P.; Abdullah, R.; Tumirah, K.; Ismail, S.; Boyce, A.N. Role of plant growth promoting rhizobacteria in agricultural sustainability—A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Amir, H.G.; Shamsuddin, Z.H.; Halimi, M.S.; Marziah, M.; Ramlan, M.F. Enhancement in nutrient accumulation and growth of oil palm seedlings caused by PGPR under field nursery conditions. Commun. Soil Sci. Plant Anal. 2005, 36, 2059–2066. [Google Scholar] [CrossRef]
- Jiménez-Gómez, A.; Flores-Félix, J.D.; García-Fraile, P.; Mateos, P.F.; Menéndez, E.; Velazquez, E.; Rivas, R. Probiotic activities of Rhizobium laguerreae on growth and quality of spinach. Sci. Rep. 2018, 8, 295. [Google Scholar] [CrossRef] [PubMed]
- Vyas, P.; Gulati, A. Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbiol. 2009, 9, 174. [Google Scholar] [CrossRef] [Green Version]
- Fasim, F.; Ahmed, N.; Parsons, R.; Gadd, G.M. Solubilization of zinc salts by a bacterium isolated from the air environment of a tannery. FEMS Microbiol. Lett. 2002, 213, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gulati, A.; Sharma, N.; Vyas, P.; Sood, S.; Rahi, P.; Pathania, V.; Prasad, R. Organic acid production and plant growth promotion as a function of phosphate solubilization by Acinetobacter rhizosphaerae strain BIHB 723 isolated from the cold deserts of the trans-Himalayas. Arch. Microbiol. 2010, 192, 975–983. [Google Scholar] [CrossRef]
- Saeid, A.; Prochownik, E.; Dobrowolska-Iwanek, J. Phosphorus solubilization by Bacillus species. Molecules 2018, 23, 2897. [Google Scholar] [CrossRef] [Green Version]
- Rajput, M.S.; Kumar, G.N.; Rajkumar, S. Repression of oxalic acid-mediated mineral phosphate solubilization in rhizospheric isolates of Klebsiella pneumoniae by succinate. Arch. Microbiol. 2012, 195, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Leite, A.D.A.; Cardoso, A.A.D.S.; Leite, R.D.A.; De Oliveira-Longatti, S.M.; Filho, J.F.L.; Moreira, F.M.D.S.; Melo, L.C.A. Selected bacterial strains enhance phosphorus availability from biochar-based rock phosphate fertilizer. Ann. Microbiol. 2020, 70, 1–13. [Google Scholar] [CrossRef]
- Rosenani, A.B.; Rovica, R.; Cheah, P.M.; Lim, C.T. Growth performance and nutrient uptake of oil palm seedling in prenursery stage as influenced by oil palm waste compost in growing media. Int. J. Agron. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Halim, N.S.A.; Abdullah, R.; Karsani, S.A.; Osman, N.; Panhwar, Q.A.; Ishak, C.F. Influence of soil amendments on the growth and yield of rice in acidic soil. Agronomy 2018, 8, 165. [Google Scholar] [CrossRef] [Green Version]
- Sung, C.T.; Ishak, C.F.; Abdullah, R.; Othman, R.; Qurban; Panhwar, A.O. Soil properties (physical, chemical, biological, mechanical). In Soils of Malaysia; Ashraf, M.A., Othman, R., Ishak, C.F., Eds.; CRC Press, Taylor & Francis Group: New York, NY, USA, 2017. [Google Scholar]
- Nelson, D.W.; Sommers, L.E.; Sparks, D.L.; Page, A.; Helmke, P.; Loeppert, R. Total carbon, organic carbon, and organic matter. In Soil Science Society of America; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 2018; pp. 961–1010. [Google Scholar]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen-Total. In Methods of Soil Analysis. Part 2 Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Ed.; Soil Science Society of America, Inc.: Madison, WI, USA, 1982. [Google Scholar]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Sim, C.C.; Zaharah, A.R.; Tan, M.S.; Goh, K.J. Rapid determination of leaf chlorophyll concentration, photosynthetic activity and NK concentration of Elaies guineensis via correlated SPAD-502 chlorophyll index. Asian J. Agric. Res. 2015, 9, 132–138. [Google Scholar] [CrossRef]
- Havlin, J.L.; Soltanpour, P.N. A nitric acid plant tissue digest method for use with inductively coupled plasma spectrometry 1. Commun. Soil Sci. Plant Anal. 1980, 11, 969–980. [Google Scholar] [CrossRef]
- Hendershot, W.; Lalande, H.; Duquette, M. Ion Exchange and Exchangeable Cations. In Soil Sampling and Methods of Analysis, 2nd ed.; Carter, M.R., Gregorich, E.G., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2006. [Google Scholar]
- FAO; IFAD; IMF; OECD; UNCTAD; WFP; World Bank; WTO; IFPRI; UNHLTF. Price Volatility in Food and Agricultural Markets: Policy Responses; Policy Report; FAO: Roma, Italy, 2 June 2011. [Google Scholar]
- Almaghrabi, O.A.; Massoud, S.I.; Abdelmoneim, T.S. Influence of inoculation with plant growth promoting rhizobacteria (PGPR) on tomato plant growth and nematode reproduction under greenhouse conditions. Saudi J. Boil. Sci. 2013, 20, 57–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.-S.; Park, K.; Kloepper, J.W.; Ryu, C.-M. Plant Growth-Promoting Rhizobacteria stimulate vegetative growth and asexual reproduction of Kalanchoe daigremontiana. Plant Pathol. J. 2015, 31, 310–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adiprasetyo, T.; Purnomo, B.; Handajaningsih, M.; Hidayat, H. The usage of BIOM3G-Biofertilizer to improve and support sustainability of land system of independent oil palm smallholders. Int. J. Adv. Sci. Eng. Inf. Technol. 2014, 4, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-H.; Vater, J.; Piel, J.; Franke, P.; Scholz, R.; Schneider, K.; Koumoutsi, A.; Hitzeroth, G.; Grammel, N.; Strittmatter, A.W.; et al. Structural and functional characterization of three polyketide synthase gene clusters in Bacillus amyloliquefaciens FZB 42. J. Bacteriol. 2006, 188, 4024–4036. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.J.; Ran, C.; Liu, K.; Ryu, C.-M.; Rasmussen-Ivey, C.R.; Williams, M.A.; Hassan, M.K.; Choi, S.-K.; Jeong, H.; Newman, M.; et al. Deciphering the conserved genetic loci implicated in plant disease control through comparative genomics of Bacillus amyloliquefaciens subsp. plantarum. Front. Plant Sci. 2015, 6, 631. [Google Scholar] [CrossRef] [Green Version]
- Illmer, P.; Schinner, F. Solubilization of inorganic phosphates by microorganisms isolated from forest soils. Soil Boil. Biochem. 1992, 24, 389–395. [Google Scholar] [CrossRef]
- Yanti, Y.; Warnita, W.; Reflin, R.; Nasution, C.R. Characterizations of endophytic Bacillus strains from tomato roots as growth promoter and biocontrol of Ralstonia solanacearum. Biodiversitas J. Boil. Divers. 2018, 19, 906–911. [Google Scholar] [CrossRef]
- Abidemi, A.A.; Akinrinde, E.A.; Obigbesan, G.O. Oil Palm (Elaeis guineensis) seedling performance in response to phosphorus fertilization in two benchmark soils of Nigeria. Asian J. Plant Sci. 2006, 5, 767–775. [Google Scholar] [CrossRef]
- Zakry, F.A.A.; Shamsuddin, Z.H.; Khairuddin, A.R.; Zin, Z.Z.; Anuar, A.R. Inoculation of Bacillus sphaericus UPMB-10 to young oil palm and measurement of its uptake of fixed nitrogen using the 15N isotope dilution technique. Microbes Environ. 2012, 27, 257–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batool, S.; Iqbal, A. Phosphate solubilizing rhizobacteria as alternative of chemical fertilizer for growth and yield of Triticum aestivum (Var. Galaxy 2013). Saudi J. Boil. Sci. 2019, 26, 1400–1410. [Google Scholar] [CrossRef] [PubMed]
- Amir, G.H.; Shamsuddin, Z.H.; Saud, M.M.H.; Ramlan, M.F.; Marziah, M. Effects of Azosprillum inoculation on N2 fixation and growth of oil palm plantlets at nursery stage. J. Oil Palm Res. 2001, 13, 42–49. [Google Scholar]
- Om, A.C.; Ghazali, A.H.A.; Keng, C.L.; Ishak, Z. Microbial inoculation improves growth of oil palm plants (Elaeis guineensis Jacq.). Trop. Life Sci. Res. 2009, 20, 71–77. [Google Scholar] [PubMed]
- Dastager, S.G.; Deepa, C.K.; Pandey, A. Growth enhancement of black pepper (Piper nigrum) by a newly isolated Bacillus tequilensis NII-0943. Biologia 2011, 66. [Google Scholar] [CrossRef] [Green Version]
- Safitri, L.; Hermantoro, H.; Purboseno, S.; Kautsar, V.; Saptomo, S.K.; Kurniawan, A. water footprint and crop water usage of oil palm (Eleasis guineensis) in Central Kalimantan: Environmental sustainability indicators for different crop age and soil conditions. Water 2018, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Yahya, Z.; Husin, A.; Talib, J.; Othman, J.; Ahmed, O.H.; Jalloh, M.B.; Yahya, O.H. Oil Palm (Elaeis guineensis) roots response to mechanization in Bernam Series soil. Am. J. Appl. Sci. 2010, 7, 343–348. [Google Scholar] [CrossRef] [Green Version]
- De Souza, R.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Boil. 2015, 38, 401–419. [Google Scholar] [CrossRef]
- Auxtero, E.A.; Shamshuddin, J. Growth of oil palm (Elaeis guineensis) seedlings on acid sulfate soils as affected by water regime and aluminium. Plant Soil 1991, 137, 243–257. [Google Scholar] [CrossRef]
- Woittiez, L.S.; Van Wijk, M.T.; Slingerland, M.; Van Noordwijk, M.; Giller, K.E. Yield gaps in oil palm: A quantitative review of contributing factors. Eur. J. Agron. 2017, 83, 57–77. [Google Scholar] [CrossRef]
- Dudeja, S.S.; Chaudhary, P. Fast chlorophyll fluorescence transient and nitrogen fixing ability of chickpea nodulation variants. Photosynthetica 2005, 43, 253–259. [Google Scholar] [CrossRef]
- Nursu’aidah, H.; Motior, M.R.; Nazia, A.; Islam, M.A. Growth and photosynthetic responses of long bean (Vigna unguiculata) and mung bean (Vigna radiata) response to fertilization. J. Anim. Plant Sci. 2014, 24, 573–578. [Google Scholar]
- Padilla, F.; De Souza, R.; Peña-Fleitas, M.T.; Grasso, R.; Gallardo, M.; Thompson, R. Influence of time of day on measurement with chlorophyll meters and canopy reflectance sensors of different crop N status. Precis. Agric. 2019, 20, 1087–1106. [Google Scholar] [CrossRef] [Green Version]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Florio, A.; Pommier, T.; Gervaix, J.; Bérard, A.; Le Roux, X. Soil C and N statuses determine the effect of maize inoculation by plant growth-promoting rhizobacteria on nitrifying and denitrifying communities. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mia, M.A.B.; Shamsuddin, Z.H.; Wahab, Z.; Marziah, M. High-yielding and quality banana production through plant growth-promoting rhizobacterial (PGPR) inoculation. Fruits 2005, 60, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Liang, B.; Yang, X.; Zhou, J. Fate of residual 15 N-labeled fertilizer in dryland farming systems on soils of contrasting fertility. Soil Sci. Plant Nutr. 2015, 61, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.-H.; Zhang, S.; Mo, X.-D.; Li, Y.; Fu, Y.-Q.; Liu, Z. Effects of Plant Growth-Promoting Rhizobacteria and N source on Plant Growth and N and P uptake by Tomato Grown on Calcareous Soils. Pedosphere 2017, 27, 1027–1036. [Google Scholar] [CrossRef]
- Mohidin, H.; Rafii, M.Y.; Man, S.; Idris, J.; Hanafi, M.M.; Abdullah, S.N.A.; Idris, A.S.; Sahebi, M. Determination of optimum levels of nitrogen, phosphorus and potassium of oil palm seedlings in solution culture. Bragantia 2015, 74, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Radin, R.; Abu Bakar, R.; Ishak, C.F.; Ahmad, S.H.; Tsong, L.C. Biochar-compost mixture as amendment for improvement of polybag-growing media and oil palm seedlings at main nursery stage. Int. J. Recycl. Org. Waste Agric. 2017, 7, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Gholizadeh, A.; Saberioon, M.; Boruvka, L.; Wayayok, A.; Soom, M.A.M. Leaf chlorophyll and nitrogen dynamics and their relationship to lowland rice yield for site-specific paddy management. Inf. Process. Agric. 2017, 4, 259–268. [Google Scholar] [CrossRef]
- Lim, S.-L.; Subramaniam, S.; Zamzuri, I.; Amir, H.G. Growth and biochemical profiling of artificially associated micropropagated oil palm plantlets with Herbaspirillum seropedicae. J. Plant Interact. 2018, 13, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Mia, M.A.B.; Shamsuddin, Z.H.; Wahab, Z.; Marziah, M. Effect of plant growth promoting rhizobacterial (PGPR) inoculation on growth and nitrogen incorporation of tissue-cultured Musa plantlets under nitrogen-free hydroponics condition. Aust. J. Crop Sci. 2010, 4, 85–90. [Google Scholar]
- A Van Veen, J.; Van Overbeek, L.S.; Van Elsas, J.D. Fate and activity of microorganisms introduced into soil. Microbiol. Mol. Boil. Rev. 1997, 61. [Google Scholar] [CrossRef]
- Veeramachaneni, S.; Ramachandrudu, K. Changes in growth, microbial and enzyme activities in oil palm nursery in response to bioinoculants and chemical fertilizers. Arch. Agron. Soil Sci. 2019, 66, 545–558. [Google Scholar] [CrossRef]
- Egamberdieva, D. The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl. Soil Ecol. 2007, 36, 184–189. [Google Scholar] [CrossRef]
- Teo, C.B.; Chew, P.S.; Goh, K.J.; Kee, K.K. Optimising return from fertilizer for oil palms: An integrated agronomic approach. In Proceedings of the IFA Regional Conference for Asia and the Pacific, Hong Kong, China, 7–10 December 1998. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Histosol | Spodosol | Ultisol | Oxisol |
|---|---|---|---|---|
| pH | 3.23 | 5.49 | 3.83 | 4.33 |
| Total N (%) | 0.61 | 0.34 | 0.10 | 0.12 |
| Availlable P (mg/kg) | 75.81 | 36.66 | 25.99 | 32.78 |
| Exchangeable K (mg/kg) | 455.2 | 487.93 | 358.33 | 471.1 |
| Micro and Macro Nutrients | Biofertilizer A | Biofertilizer B |
|---|---|---|
| N | 7% | 5–6% |
| P | 6% | 8–9% |
| K | 9% | 10–11% |
| Ca | 2% | - |
| Mg | 1% | 0.5–1.0% |
| Su | 1% | - |
| Bo | 0.5% | 0.9–1.1% |
| Fe | 50 ppm | 282 ppm |
| Cu | 15 ppm | 18.4 ppm |
| Mn | 10 ppm | 35.8 ppm |
| Zn | 15 ppm | 51.4 ppm |
| Mo | 12 ppm | - |
| Organic matter | Aloe vera Seaweed extract Fulvic acid Amino acid Protein | Aloe vera Seaweed extract Humic acid Amino acid Fish emulsify |
| Month | Control Plot | Treatment Plot | |
|---|---|---|---|
| Dosage per Palm (g seedlings−1) (NPK 12-12-17-2 + TE) | Biofertilizer (mL) | ||
| 100% Chemical Fertilizer | 75% Chemical Fertilizer | ||
| 1 | 15 | 10 | 2 |
| 2 | 20 | 15 | 2 |
| 3 | 25 | 20 | 3 |
| 4 | 30 | 25 | 3 |
| Soil | Treatment | ABG | Root | Root:ABG |
|---|---|---|---|---|
| Histosol | T1 | 57.97 ± 9.92b | 16.22 ± 4.46a | 0.28 ± 0.03a |
| T2 | 62.07 ± 3.47a | 16.10 ± 3.31a | 0.26 ± 0.04ab | |
| T3 | 59.50 ± 17.47b | 15.09 ± 3.61a | 0.26 ± 0.04b | |
| Spodosol | T1 | 49.62 ± 14.32b | 14.25 ± 4.21a | 0.29 ± 0.02a |
| T2 | 63.48 ± 7.08ab | 16.52 ± 0.92a | 0.26 ± 0.02ab | |
| T3 | 64.53 ± 4.99a | 15.84 ± 1.17a | 0.25 ± 0.02b | |
| Ultisol | T1 | 53.61 ± 3.80b | 11.70 ± 0.68a | 0.22 ± 0.01ab |
| T2 | 66.34 ± 2.50ab | 15.20 ± 1.26a | 0.23 ± 0.02a | |
| T3 | 70.39 ± 7.98a | 13.92 ± 1.60a | 0.20 ± 0.00b | |
| Oxisol | T1 | 65.97 ± 4.61b | 15.55 ± 2.95a | 0.24 ± 0.06ab |
| T2 | 58.70 ± 11.13ab | 14.30 ± 1.03a | 0.25 ± 0.02a | |
| T3 | 78.21 ± 14.91a | 16.44 ± 0.95a | 0.22 ± 0.00b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ajeng, A.A.; Abdullah, R.; Malek, M.A.; Chew, K.W.; Ho, Y.-C.; Ling, T.C.; Lau, B.F.; Show, P.L. The Effects of Biofertilizers on Growth, Soil Fertility, and Nutrients Uptake of Oil Palm (Elaeis Guineensis) under Greenhouse Conditions. Processes 2020, 8, 1681. https://doi.org/10.3390/pr8121681
Ajeng AA, Abdullah R, Malek MA, Chew KW, Ho Y-C, Ling TC, Lau BF, Show PL. The Effects of Biofertilizers on Growth, Soil Fertility, and Nutrients Uptake of Oil Palm (Elaeis Guineensis) under Greenhouse Conditions. Processes. 2020; 8(12):1681. https://doi.org/10.3390/pr8121681
Chicago/Turabian StyleAjeng, Aaronn Avit, Rosazlin Abdullah, Marlinda Abdul Malek, Kit Wayne Chew, Yeek-Chia Ho, Tau Chuan Ling, Beng Fye Lau, and Pau Loke Show. 2020. "The Effects of Biofertilizers on Growth, Soil Fertility, and Nutrients Uptake of Oil Palm (Elaeis Guineensis) under Greenhouse Conditions" Processes 8, no. 12: 1681. https://doi.org/10.3390/pr8121681