
Impact of Bee Venom Enzymes on Diseases and Immune Responses



Abstract
:1. Introduction
2. Methodology
3. Bee Venom Enzymes
3.1. Phospholipase A2
3.2. Phospholipase B
3.3. Hyaluronidase
3.4. Acid Phosphatase
3.5. α-Glucosidase
4. Investigated Mechanistic Roles of BV PLA2 and Others on Several Diseases
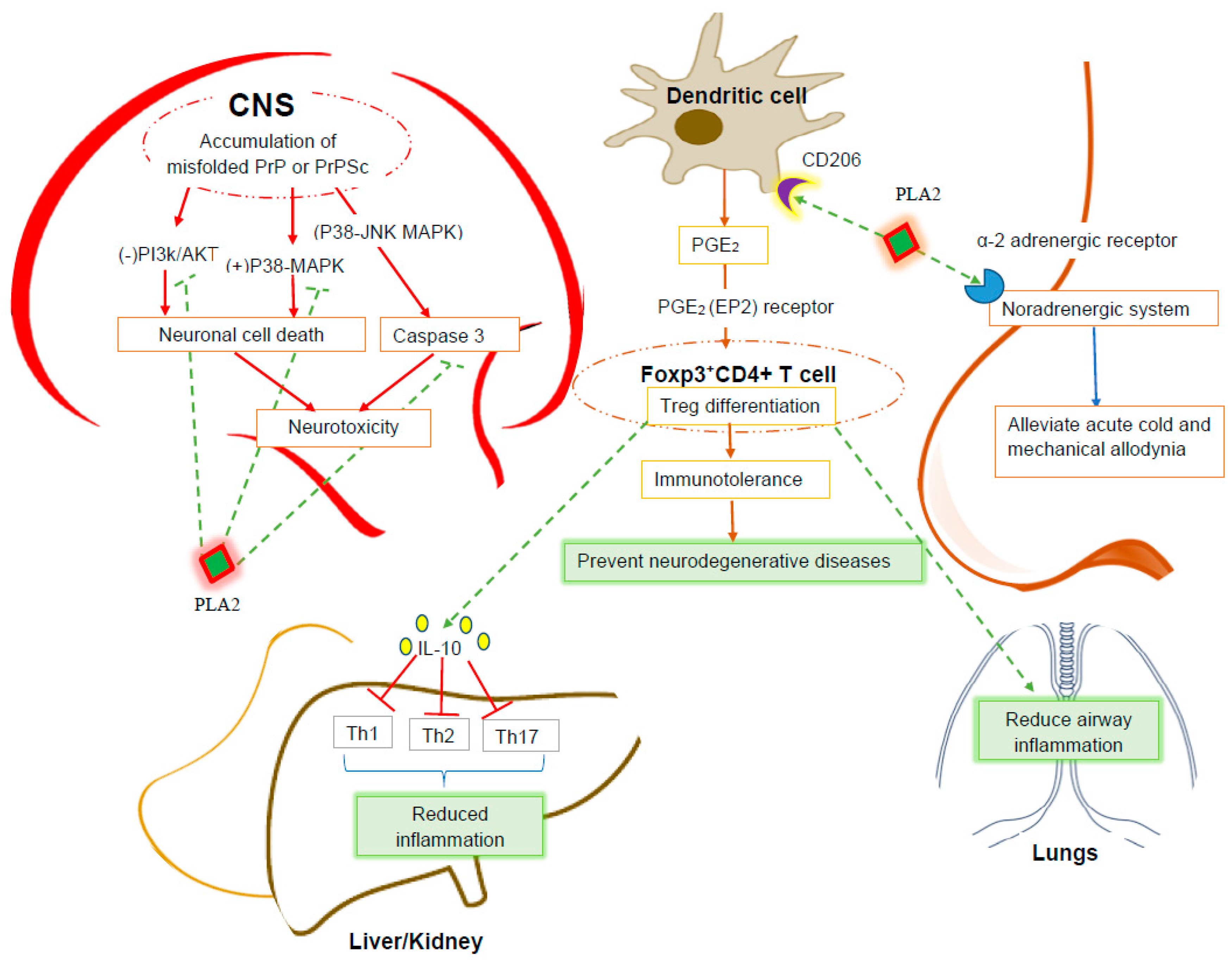
4.1. Anti-Neurodegenerative Effect of PLA2
4.2. Anti-Inflammatory Effect of BV PLA2
4.3. Anti-Nociceptive Effect of BV PLA2
4.4. Anti-Cancer, Anti-Bacterial, Anti-Parasitic and Immunotherapeutic Effects of BV PLA2
4.5. Biochemical and Physiological Roles of Hyaluronidase
4.6. Physiological Role of BV Acid Phosphatase
5. Future Approaches
6. Conclusions
Acknowledgments
Author Contributions
Conflict of Interest
References
- Lee, J.Y.; Kang, S.S.; Kim, J.-H.; Bae, C.S.; Choi, S.H. Inhibitory effect of whole bee venom in adjuvant-induced arthritis. In Vivo 2005, 19, 801–805. [Google Scholar] [PubMed]
- Gajski, G.; Garaj-Vrhovac, V. Radioprotective effects of honeybee venom (Apis mellifera) against 915-mhz microwave radiation–induced DNA damage in wistar rat lymphocytes: In vitro study. Int. J. Toxicol. 2009, 28, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.C.; Koh, P.S.; Seo, B.K.; Lee, J.W.; Cho, N.S.; Park, H.S.; Park, D.S.; Baek, Y.H. Long-term effectiveness of bee venom acupuncture and physiotherapy in the treatment of adhesive capsulitis: A one-year follow-up analysis of a previous randomized controlled trial. J. Altern. Complement. Med. 2014, 20, 919–924. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, G.; Park, S.; Chung, H.-S.; Lee, H.; Kim, J.-Y.; Nam, S.; Kim, S.K.; Bae, H. Bee venom mitigates cisplatin-induced nephrotoxicity by regulating CD4. Evid. Based Complement. Altern. Med. 2013, 2013, 879845. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.S.; Park, S.; Choi, T.; Lee, G.; Haam, K.-K.; Hong, M.-C.; Min, B.-I.; Bae, H. Bee venom ameliorates ovalbumin induced allergic asthma via modulating cd4+ cd25+ regulatory t cells in mice. Cytokine 2013, 61, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-G.; Cho, H.-J.; Bae, Y.-S.; Park, K.-K.; Choe, J.-Y.; Chung, I.-K.; Kim, M.; Yeo, J.-H.; Park, K.-H.; Lee, Y.-S. Bee venom suppresses LPS-mediated no/inos induction through inhibition of PKC-α expression. J. Ethnopharmacol. 2009, 123, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.-S.; Moon, H.J.; Li, D.X.; Gil, M.; Min, J.K.; Lee, G.; Bae, H.; Kim, S.K.; Min, B.-I. Effect of bee venom acupuncture on oxaliplatin-induced cold allodynia in rats. Evid. Based Complement. Altern. Med. 2013, 2013, 369324. [Google Scholar] [CrossRef] [PubMed]
- Van Vaerenbergh, M.; Debyser, G.; Devreese, B.; de Graaf, D.C. Exploring the hidden honeybee (Apis mellifera) venom proteome by integrating a combinatorial peptide ligand library approach with ftms. J. Proteom. 2014, 99, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, S. Bee venom: Composition, health, medicine: A review. Peptides 2015, 1, 1–20. [Google Scholar]
- Shimpi, R.; Chaudhari, P.; Deshmukh, R.; Devare, S.; Bagad, Y.; Bhurat, M. A Review: Pharmacotherapeutics of Bee Venom. WJPPS 2016, 5, 656–667. [Google Scholar]
- Chung, E.S.; Lee, G.; Lee, C.; Ye, M.; Chung, H.-S.; Kim, H.; Sung-joo, S.B.; Hwang, D.-S.; Bae, H. Bee venom phospholipase a2, a novel foxp3+ regulatory t cell inducer, protects dopaminergic neurons by modulating neuroinflammatory responses in a mouse model of parkinson’s disease. J. Immunol. 2015, 195, 4853–4860. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; Xu, J.; Jensen, M.D.; Simonyi, A. Phospholipase a2 in the central nervous system implications for neurodegenerative diseases. J. Lipid Res. 2004, 45, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Landucci, E.C.; Toyama, M.; Marangoni, S.; Oliveira, B.; Cirino, G.; Antunes, E.; de Nucci, G. Effect of crotapotin and heparin on the rat paw oedema induced by different secretory phospholipases A 2. Toxicon 2000, 38, 199–208. [Google Scholar] [CrossRef]
- Park, S.; Baek, H.; Jung, K.H.; Lee, G.; Lee, H.; Kang, G.H.; Lee, G.; Bae, H. Bee venom phospholipase a2 suppresses allergic airway inflammation in an ovalbumin-induced asthma model through the induction of regulatory t cells. Immun. Inflamm. Dis. 2015, 3, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Marty, J.P.; Clauss, C.; Michel, F.B. Enzymes of bee venom, sac and whole body. Ann. Allergy 1979, 43, 110–114. [Google Scholar] [PubMed]
- Doery, H.M.; Pearson, J.E. Phospholipase b in snake venoms and bee venom. Biochem. J. 1964, 92, 599–602. [Google Scholar] [CrossRef] [PubMed]
- Szókán, G.; Horvath, J.; Almas, M.; Saftics, G.; Palocz, A. Liquid chromatographic analysis and separation of polypeptide components from honey bee venoms. J. Liquid Chromatogr. Relat. Technol. 1994, 17, 3333–3349. [Google Scholar] [CrossRef]
- Jeong, J.-K.; Moon, M.-H.; Bae, B.-C.; Lee, Y.-J.; Seo, J.; Park, S.-Y. Bee venom phospholipase a2 prevents prion peptide induced-cell death in neuronal cells. Int. J. Mol. Med. 2011, 28, 867–873. [Google Scholar] [PubMed]
- Son, D.J.; Lee, J.W.; Lee, Y.H.; Song, H.S.; Lee, C.K.; Hong, J.T. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 2007, 115, 246–270. [Google Scholar] [CrossRef] [PubMed]
- Urtubey, N. Apitoxin: From Bee Venom to Apitoxin for Medical Use; Termas de Rio Grande Santiago del Estero: Rio Hondo, Argentina, 2005. [Google Scholar]
- Kim, H.; Keum, D.J.; won Kwak, J.; Chung, H.-S.; Bae, H. Bee venom phospholipase a2 protects against acetaminophen-induced acute liver injury by modulating regulatory t cells and IL-10 in mice. PLoS ONE 2014, 9, e114726. [Google Scholar] [CrossRef] [PubMed]
- Banks, B.E.; Shipolini, R.A. Chemistry and pharmacology of honey-bee venom. In Venoms Hymenoptera: Biochemtry, Pharmacological Behavioral Aspects; Academic press, Inc.: Orlando, Florida, USA, 1986. [Google Scholar]
- Grunwald, T.; Bockisch, B.; Spillner, E.; Ring, J.; Bredehorst, R.; Ollert, M.W. Molecular cloning and expression in insect cells of honeybee venom allergen acid phosphatase (api m 3). J. Allergy Clin. Immunol. 2006, 117, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Kubota, M.; Tsuji, M.; Nishimoto, M.; Wongchawalit, J.; Okuyama, M.; Mori, H.; Matsui, H.; Surarit, R.; Svasti, J.; Kimura, A. Localization of α-glucosidases i, ii, and iii in organs of european honeybees, Apis mellifera L., and the origin of α-glucosidase in honey. Biosci. Biotechnol. Biochem. 2004, 68, 2346–2352. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, M.; Kubota, M.; Tsuji, M.; Mori, H.; Kimura, A.; Matsui, H.; Chiba, S. Purification and substrate specificity of honeybee, Apis mellifera L., α-glucosidase iii. Biosci. Biotechnol. Biochem. 2001, 65, 1610–1616. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Sato, H.; Miki, Y.; Yamamoto, K.; Taketomi, Y. A new era of secreted phospholipase a2. J. Lipid Res. 2015, 56, 1248–1261. [Google Scholar] [CrossRef] [PubMed]
- Welker, S.; Markert, Y.; Koditz, J.; Mansfeld, J.; Ulbrich-Hofmann, R. Disulfide bonds of phospholipase a2 from bee venom yield discrete contributions to its conformational stability. Biochimie 2011, 93, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Gomez, F.; Vandermeers, A.; Vandermeers-Piret, M.C.; Herzog, R.; Rathe, J.; Stievenart, M.; Winand, J.; Christophe, J. Purification and characterization of five variants of phospholipase A2 and complete primary structure of the main phospholipase a2 variant in heloderma suspectum (gila monster) venom. FEBS J. 1989, 186, 23–33. [Google Scholar] [CrossRef]
- Ferreira Junior, R.S.; Sciani, J.M.; Marques-Porto, R.; Junior, A.L.; Orsi Rde, O.; Barraviera, B.; Pimenta, D.C. Africanized honey bee (Apis mellifera) venom profiling: Seasonal variation of melittin and phospholipase A 2 levels. Toxicon 2010, 56, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Habermann, E.; Hardt, K. A sensitive and specific plate test for the quantitation of phospholipases. Anal. Biochem. 1972, 50, 163–173. [Google Scholar] [CrossRef]
- Gmachl, M.; Kreil, G. Bee venom hyaluronidase is homologous to a membrane protein of mammalian sperm. Proc. Natl. Acad. Sci. USA 1993, 90, 3569–3573. [Google Scholar] [CrossRef] [PubMed]
- März, L.; Kühne, C.; Michl, H. The glycoprotein nature of phospholipase A2, hyaluronidase and acid phosphatase from honey-bee venom. Toxicon 1983, 21, 893–896. [Google Scholar] [CrossRef]
- Chiba, S. Molecular mechanism in α-glucosidase and glucoamylase. Biosci. Biotechnol. Biochem. 1997, 61, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Takewaki, S.-I.; Matsui, H.; Kubota, M.; Chiba, S. Allosteric properties, substrate specificity, and subsite affinities of honeybee α-glucosidase I. J. Biochem. 1990, 107, 762–768. [Google Scholar] [PubMed]
- Masuda, S.; Yamamoto, K.; Hirabayashi, T.; Ishikawa, Y.; Ishii, T.; Kudo, I.; Murakami, M. Human group iii secreted phospholipase a2 promotes neuronal outgrowth and survival. Biochem. J. 2008, 409, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Yawo, H.; Kuno, M. How a nerve fiber repairs its cut end: Involvement of phospholipase A2. Science 1983, 222, 1351–1353. [Google Scholar] [CrossRef] [PubMed]
- Vila, M.; Jackson-Lewis, V.; Guégan, C.; Teismann, P.; Choi, D.-K.; Tieu, K.; Przedborski, S. The role of glial cells in Parkinson’s disease. Curr. Opin. Neurol. 2001, 14, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Appel, S.H. CD4+ t cells mediate cytotoxicity in neurodegenerative diseases. J. Clin. Investig. 2009, 119, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Chung, H.-S.; Lee, C.; Yoon, M.S.; Yu, A.R.; Kim, J.S.; Hwang, D.-S.; Shim, I.; Bae, H. Neuroprotective effects of bee venom phospholipase A2 in the 3xTg ad mouse model of alzheimer’s disease. J. Neuroinflamm. 2016, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, H.A. Molecular pathogenesis of prion diseases. Eur. Arch. Psychiatry Clin. Neurosci. 1999, 249, S56–S63. [Google Scholar] [CrossRef]
- Kim, H.; Lee, H.; Lee, G.; Jang, H.; Kim, S.-S.; Yoon, H.; Kang, G.-H.; Hwang, D.-S.; Kim, S.K.; Chung, H.-S. Phospholipase a2 inhibits cisplatin-induced acute kidney injury by modulating regulatory t cells by the CD206 mannose receptor. Kidney Int. 2015, 88, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Tsoutsou, P.G.; Koukourakis, M.I. Radiation pneumonitis and fibrosis: Mechanisms underlying its pathogenesis and implications for future research. Int. J. Radiat. Oncol. Biol. Phys. 2006, 66, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Lee, G.; Sohn, S.-H.; Park, S.; Jung, K.-H.; Lee, J.M.; Yang, J.; Cho, J.; Bae, H. Regulatory t cells contribute to the inhibition of radiation-induced acute lung inflammation via bee venom phospholipase A2 in mice. Toxins 2016, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Lee, Y.; Kim, W.; Lee, K.; Bae, H.; Kim, S.K. Analgesic effects of bee venom derived phospholipase a2 in a mouse model of oxaliplatin-induced neuropathic pain. Toxins 2015, 7, 2422–2434. [Google Scholar] [CrossRef] [PubMed]
- Putz, T.; Ramoner, R.; Gander, H.; Rahm, A.; Bartsch, G.; Thurnher, M. Antitumor action and immune activation through cooperation of bee venom secretory phospholipase a2 and phosphatidylinositol-(3, 4)-bisphosphate. Cancer Immunol. Immunother. 2006, 55, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Putz, T.; Ramoner, R.; Gander, H.; Rahm, A.; Bartsch, G.; Bernardo, K.; Ramsay, S.; Thurnher, M. Bee venom secretory phospholipase A2 and phosphatidylinositol-homologues cooperatively disrupt membrane integrity, abrogate signal transduction and inhibit proliferation of renal cancer cells. Cancer Immunol. Immunother. 2007, 56, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-C.; Yen, M.-H.; Liu, H.-W.; Lau, Y.-T. Lysophosphatidylcholine induces apoptotic and non-apoptotic death in vascular smooth muscle cells: In comparison with oxidized ldl. Atherosclerosis 2000, 151, 481–491. [Google Scholar] [CrossRef]
- Ohara, Y.; Peterson, T.E.; Zheng, B.; Kuo, J.; Harrison, D.G. Lysophosphatidylcholine increases vascular superoxide anion production via protein kinase c activation. Arterioscler. Thromb. Vasc. Biol. 1994, 14, 1007–1013. [Google Scholar] [CrossRef]
- Boutrin, M.-C.; Foster, H.; Pentreath, V. The effects of bee (Apis mellifera) venom phospholipase a2 on trypanosoma brucei brucei and enterobacteria. Exp. Parasitol. 2008, 119, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.A.; Ito, J.; Ghosh, A.; Devenport, M.; Zieler, H.; Abraham, E.G.; Crisanti, A.; Nolan, T.; Catteruccia, F.; Jacobs-Lorena, M. Bee venom phospholipase inhibits malaria parasite development in transgenic mosquitoes. J. Biol. Chem. 2002, 277, 40839–40843. [Google Scholar] [CrossRef] [PubMed]
- Kugler, A.; Stuhler, G.; Walden, P.; Zöller, G.; Zobywalski, A.; Brossart, P.; Trefzer, U.; Ullrich, S.; Müller, C.A.; Becker, V. Regression of human metastatic renal cell carcinoma after vaccination with tumor cell–dendritic cell hybrids. Nat. Med. 2000, 6, 332–336. [Google Scholar] [PubMed]
- Herr, W.; Ranieri, E.; Olson, W.; Zarour, H.; Gesualdo, L.; Storkus, W.J. Mature dendritic cells pulsed with freeze–thaw cell lysates define an effective in vitro vaccine designed to elicit ebv-specific CD4+ and CD8+ t lymphocyte responses. Blood 2000, 96, 1857–1864. [Google Scholar] [PubMed]
- Arrode, G.; Boccaccio, C.; Lulé, J.; Allart, S.; Moinard, N.; Abastado, J.-P.; Alam, A.; Davrinche, C. Incoming human cytomegalovirus pp65 (ul83) contained in apoptotic infected fibroblasts is cross-presented to CD8+ t cells by dendritic cells. J. Virol. 2000, 74, 10018–10024. [Google Scholar] [CrossRef] [PubMed]
- Tamura, Y.; Peng, P.; Liu, K.; Daou, M.; Srivastava, P.K. Immunotherapy of tumors with autologous tumor-derived heat shock protein preparations. Science 1997, 278, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Boczkowski, D.; Nair, S.K.; Nam, J.-H.; Lyerly, H.K.; Gilboa, E. Induction of tumor immunity and cytotoxic t lymphocyte responses using dendritic cells transfected with messenger rna amplified from tumor cells. Cancer Res. 2000, 60, 1028–1034. [Google Scholar] [PubMed]
- Hacein-Bey-Abina, S.; von Kalle, C.; Schmidt, M.; le Deist, F.; Wulffraat, N.; McIntyre, E.; Radford, I.; Villeval, J.-L.; Fraser, C.C.; Cavazzana-Calvo, M. A serious adverse event after successful gene therapy for x-linked severe combined immunodeficiency. N. Engl. J. Med. 2003, 348, 255–256. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.H.; Engelhardt, J.F.; Yang, Y.; Zepeda, M.; Weber-Pendleton, S.; Grossman, M.; Wilson, J.M. Adenovirus-mediated transfer of the cftr gene to lung of nonhuman primates: Toxicity study. Hum. Gene Ther. 1993, 4, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Almunia, C.; Bretaudeau, M.; Held, G.; Babon, A.; Marchetti, C.; Castelli, F.A.; Ménez, A.; Maillere, B.; Gillet, D. Bee venom phospholipase a2, a good “chauffeur” for delivering tumor antigen to the mhc i and mhc ii peptide-loading compartments of the dendritic cells: The case of ny-eso-1. PLoS ONE 2013, 8, e67645. [Google Scholar] [CrossRef] [PubMed]
- Müller, U.; Crameri, R.; Soldatova, L. Diagnostik mit rekombinenten/synthetischen bienengiftallergenen. Allergologie 1999, 22, 51–52. [Google Scholar]
- Bordon, K.C.; Wiezel, G.A.; Amorim, F.G.; Arantes, E.C. Arthropod venom hyaluronidases: Biochemical properties and potential applications in medicine and biotechnology. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, D.; Greunke, K.; Genov, N.; Betzel, C. 3-D model of the bee venom acid phosphatase: Insights into allergenicity. Biochem. Biophys. Res. Commun. 2009, 378, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Barboni, E.; Kemeny, D.; Campos, S.; Vernon, C. The purification of acid phosphatase from honey bee venom (Apis mellifica). Toxicon 1987, 25, 1097–1103. [Google Scholar] [CrossRef]
- Nicolas, J.-P.; Lin, Y.; Lambeau, G.; Ghomashchi, F.; Lazdunski, M.; Gelb, M.H. Localization of structural elements of bee venom phospholipase A2 involved in n-type receptor binding and neurotoxicity. J. Biol. Chem. 1997, 272, 7173–7181. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.K.; Zhang, Y.P.; Titsworth, W.L.; Jiang, X.; Han, S.; Lu, P.H.; Shields, C.B.; Xu, X.M. A novel role of phospholipase A2 in mediating spinal cord secondary injury. Ann. Neurol. 2006, 59, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Titsworth, W.L.; Onifer, S.M.; Liu, N.-K.; Xu, X.-M. Focal phospholipases a 2 group iii injections induce cervical white matter injury and functional deficits with delayed recovery concomitant with schwann cell remyelination. Exp. Neurol. 2007, 207, 150–162. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| BV Enzyme | Identification Method | Biological Function | Reference |
|---|---|---|---|
| PLA2 | Reversed phase HPLC on different columns with an acetonitrile-water-trifluoro acetic acid eluent system, nano-LC/MALDI-TOF/TOF-MS system, shotgun LC-FT-ICR MS analysis | Causes destruction of phospholipids and dissolves the cell membrane of blood cells. Lowers blood coagulation and blood pressure, prevents neuronal cell death caused by prion peptides. Prevents acetaminophen-induced hepatotoxicity through CD4+, CD25+, Foxp3+ and T cells (Treg) in mice. | [17,18,19,20,21] |
| Phospholipase B | Paper chromatography and also paper electrophoresis | Causes cleavage of lysolectin and thus detoxification. Has the combined activity of PLA1 and PLA2. Enhances the activity of PLA2. Cleaves lysophospholipid into glycerophosphocholine and anionic fatty acids. | [16,18,19,20,22] |
| Hyaluronidase | Reverse-phase HPLC on different columns with an acetonitrile-water-trifluoroacetic acid eluent system, nano-LC/MALDI-TOF/TOF-MS system, shotgun LC-FT-ICR MS analysis | Degrades hyaluronic acid, enabling the penetration of venom into tissue. Causes dilation and increased permeability of blood vessels and thus increased blood circulation. | [17,18,19,20] |
| Acid phosphatase | 1-D SDS PAGE in gel digest followed by LC-ESI-LTQ-FT-ICR-MS, nano-LC/MALDI-TOF/TOF-MS system, shotgun LC-FT-ICR MS analysis | Potent histamine releaser from human sensitized basophils. May be exploited in immunotherapy against BV. | [23] |
| α-glucosidase | Salting-out chromatography, DEAE-cellulose, DEAE-sepharose CL-6B, Bio-Gel P-150, and CM-toyopearl 650M column chromatographies, shotgun LC-FT-ICR MS analysis | Involved in honey production. | [24,25] |
| Phospholipase A2 | Hyaluronidase | Acid Phosphatase | α-Glucosidase | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nomenclature and Source | ||||||||||||||||
| Recommended name | Phospholipase A2 | Hyaluronoglucosaminidase | Venom acid phosphatase | Alpha glucosidase | ||||||||||||
| Alternative names | Allergen Api m I, Allergen: Api m 1, lecithinase A, phosphatidase, phosphatidolipase, phospholipase A | Allergen Api m II, Allergen: Api m 2, hyaluronoglucosidase, chondroitinase, chondroitinase I, BVH, HAse, HyaI, Mu toxin, sperm surface protein PH-20 | Allergen: Api m 3, acid phosphatase | Acid maltase, AGL, α-1,4-glucosidase, α-d-glucosidase, α-glucopyranosidase, α-glucoside hydrolase, α-glucosidase III, glucoinvertase, glucosidoinvertase, glucosidosucrase, maltase, maltase glucoamylase, HBG III, HBGase I, HBGase II, HBGase III | ||||||||||||
| Systematic name | Phosphatidylcholine 2-acylhydrolase | Hyaluronate 4-glycanohydrolase | Phosphate-monoester phosphohydrolase (acid optimum) | α-d-glucosideglucohydrolase | ||||||||||||
| Short name | bvPLA2 | Hya | Acph-1 | AGL | ||||||||||||
| Popular sources | Mouse, human, bovine, rat, fruit fly | Human, zebrafish, rat, bovine | Rice, Arabidopsis thaliana, fruit fly, human, Caenorhabditis elegan, bees, viper snakes | Fruit fly, human, mouse, Saccharomyces cerevisiae, Arabidopsis thaliana | ||||||||||||
| Organism-based information | ||||||||||||||||
| Source tissue | Venom gland | Expressed in the venom glands of worker bees; it is also detected in the testes of drones but not in the queen’s BV gland or in pupae | Venom gland | Hypopharyngeal gland of worker bees | ||||||||||||
| Subcellular location | Extracellular | Extracellular | Extracellular | No data | ||||||||||||
| Maximal levels | Approximately 40 micrograms PLA2/venom sac | No data | No data | No data | ||||||||||||
| Percentage of dry weight (%) | 10–12 | 1–2 | 1 | 0.6 | ||||||||||||
| Purification |
|
|
|
| ||||||||||||
| Enzymatics | ||||||||||||||||
| EC number | 3.1.1.4 | 3.2.1.35 | 3.1.3.2 | 3.2.1.20 | ||||||||||||
| Family | Phospholipase A2 family; group III subfamily | Glycosyl hydrolase 56 family | Venom acid phosphatase | Glycoside hydrolase family 13 | ||||||||||||
| Reaction type | Hydrolysis of carboxylic ester | Hydrolysis of O-glycosyl bond | Acid phosphatase activity | Hydrolysis | ||||||||||||
| Reaction | Phosphatidylcholine + H2O = 1-acylglycerophosphocholine + a carboxylate | Random hydrolysis of (1→4)-linkages between N-acetyl-beta-d-glucosamine and-glucuronate residues in hyaluronate | Phosphatidylcholine + H2O = 1-acylglycerophosphocholine + a carboxylate | Hydrolysis of terminal, non-reducing (1→4)-linked α-d-glucose residues with release of α-d-glucose | ||||||||||||
| Metabolic pathways |
|
|
|
| ||||||||||||
| Metals and ions | Ca2+, Others: Ba2+, Mg2+, Sr2+ | No data | No data | Ca2+ | ||||||||||||
| Inhibitors |
|
| Not found |
| ||||||||||||
| Activators |
| Not investigated | Not investigated | Not investigated | ||||||||||||
| Active sites |
| Not investigated |
|
| ||||||||||||
| Optimum pH |
|
|
|
| ||||||||||||
| Optimum temperature (°C) |
|
|
| No data | ||||||||||||
| Temperature stability ((°C) |
|
| No data |
| ||||||||||||
| Genomics | ||||||||||||||||
| Gene symbol |
|
|
|
| ||||||||||||
| Other names |
|
|
|
| ||||||||||||
| Gene type |
|
|
|
| ||||||||||||
| RefSeq status |
|
|
|
| ||||||||||||
| Location |
|
|
|
| ||||||||||||
| Exon count |
|
|
|
| ||||||||||||
| Proteomics | ||||||||||||||||
| Formula |
|
|
|
| ||||||||||||
| Total number of atoms |
|
|
|
| ||||||||||||
| Molecular weight (Dalton) |
|
|
|
| ||||||||||||
| Length |
|
|
|
| ||||||||||||
| Theoretical pI |
|
|
|
| ||||||||||||
| Total number of negatively charged residues (Asp + Glu) |
|
|
|
| ||||||||||||
| Total number of positively charged residues (Arg + Lys) |
|
|
|
| ||||||||||||
| Estimated half-life |
|
|
|
| ||||||||||||
| Instability index |
|
|
|
| ||||||||||||
| Grand average hydropathicity (GRAVY) |
|
|
|
| ||||||||||||
| Graphical overview |  |  |  |  | ||||||||||||
| Source | Domain | Start | End | Source | Domain | Start | End | Source | Domain | Start | End | Source | Domain | Start | End | |
| 1 = Sig P | n/a | 1 | 18 | 1 = Transmembrane | n/a | 12 | 30 | 1 = Sig P | n/a | 1 | 15 | 1 = Sig P | n/a | 1 | 17 | |
| 14 = pfam | Phospholipid A2-2 | 35 | 132 | 5 = pfam | Glyco hydro 56 | 38 | 367 | 6 = pfam | His_Phos_2 | 16 | 187 | 2 = Low complexity | n/a | 44 | 56 | |
| Low complexity | n/a | 4 | 15 | Low complexity | His_Phos_2 | 15 | 26 | 7 = pfam | His_Phos_2 | 162 | 315 | 3 = pfam | α-Amylase | 49 | 394 | |
| Disorder | n/a | 231 | 236 | Low complexity | n/a | 233 | 244 | 6 = disorder | n/a | 400 | 410 | |||||
| 7 = disorder | n/a | 412 | 414 | |||||||||||||
| 8 = Low complexity | n/a | 424 | 433 | |||||||||||||
| Disulfide | Disulfide | |||||||||||||||
| 9 = Disulfide, coordinates 42–64 | 2 = Disulfide, coordinates 54-345 | |||||||||||||||
| 10 = Disulfide, coordinates 63–103 | 3 = Disulfide, coordinates 221-233 | |||||||||||||||
| 11 = Disulfide, coordinates 70–96 | ||||||||||||||||
| 12 = Disulfide, coordinates 94–128 | ||||||||||||||||
| 13 = Disulfide, coordinates 138–146 | ||||||||||||||||
| Annotation | Annotation | Annotation | Annotation | |||||||||||||
| 7 = H-active site, position 67 | 4 = E-active site: proton donor, position 145 | 2 = H-active site nucleophile, position 26 | 4 = D-active site nucleophile, position 223 | |||||||||||||
| 8 = D-active site, position 97 | 4 = E-active site: proton donor, position 273 | 5 = E-active site: proton donor, position 286 | ||||||||||||||
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossen, M.S.; Shapla, U.M.; Gan, S.H.; Khalil, M.I. Impact of Bee Venom Enzymes on Diseases and Immune Responses. Molecules 2017, 22, 25. https://doi.org/10.3390/molecules22010025
Hossen MS, Shapla UM, Gan SH, Khalil MI. Impact of Bee Venom Enzymes on Diseases and Immune Responses. Molecules. 2017; 22(1):25. https://doi.org/10.3390/molecules22010025
Chicago/Turabian StyleHossen, Md. Sakib, Ummay Mahfuza Shapla, Siew Hua Gan, and Md. Ibrahim Khalil. 2017. "Impact of Bee Venom Enzymes on Diseases and Immune Responses" Molecules 22, no. 1: 25. https://doi.org/10.3390/molecules22010025