
An Overview of Metabolic Activity, Beneficial and Pathogenic Aspects of Burkholderia Spp.



School of Agricultural, Forestry, Food and Environmental Sciences, University of Basilicata, Via dell’Ateneo Lucano 10, 85100 Potenza, Italy
*
Author to whom correspondence should be addressed.
Metabolites 2021, 11(5), 321; https://doi.org/10.3390/metabo11050321
Submission received: 15 April 2021
/
Revised: 13 May 2021
/
Accepted: 14 May 2021
/
Published: 17 May 2021
(This article belongs to the Special Issue Metabolites from Bacterial Pathogens and Their Role in Disease)
Abstract
:Burkholderia is an important bacterial species which has different beneficial effects, such as promoting the plant growth, including rhizosphere competence for the secretion of allelochemicals, production of antibiotics, and siderophores. In addition, most of Burkholderia species have demonstrated promising biocontrol action against different phytopathogens for diverse crops. In particular, Burkholderia demonstrates significant biotechnological potential as a source of novel antibiotics and bioactive secondary metabolites. The current review is concerned with Burkholderia spp. covering the following aspects: discovering, classification, distribution, plant growth promoting effect, and antimicrobial activity of different species of Burkholderia, shedding light on the most important secondary metabolites, their pathogenic effects, and biochemical characterization of some important species of Burkholderia, such as B. cepacia, B. andropogonis, B. plantarii, B. rhizoxinica, B. glumae, B. caryophylli and B. gladioli.
1. Genus Burkholderia
1.1. Discovering, Classification and Distribution
In 1942, Burkholder described one of the first Burkholderia species, Phytomonas caryophylli [1], later known as Pseudomonas caryophylli. In 1949, Burkholder also described another bacterium that caused rot in onion bulbs, as reported by vegetable growers in New York State in the mid-1940s, and gave it the species name ‘cepacia’, which was later known as Pseudomonas cepacia [2]. Burkholderia spp. was included for many years in the genus Pseudomonas due to broad and vague phenotypic characteristics [3]. However, rRNA–DNA hybridization analyses during the early 1970s indicated considerable genetic diversity among members of this genus which was divided into five rRNA homology groups [4]. Subsequent genotypic analyses have confirmed that these five groups are only distantly related to each other. Consequently, Pseudomonas was restricted to homology group I containing the type species Pseudomonas aeruginosa [5]. In 1992, the seven species belonging to rRNA homology group II (Pseudomonas solanacearum, P. pickettii, P. cepacia, P. gladioli, P. mallei, P. pseudomallei and P. caryophylli) were transferred to the novel genus Burkholderia [6]. The members of genus Burkholderia have a broad distribution occurring commonly in soil, water, in symbiosis with plants and fungi and in association with animals and humans [7,8]. Burkholderia are motile and gram negative (G-ve) rods that may be straight or slightly curved. They are aerobic, catalase positive, urease positive, non-spore formers and non-lactose fermenting [9].
PCR can be used to distinguish between the different Burkholderia species. The ribosomal RNA gene is highly conserved and universally distributed in all living things, and therefore the difference in the DNA sequences between 16S and 23S rRNA genes can be used to differentiate between different species [10].
1.2. Species of Burkholderia
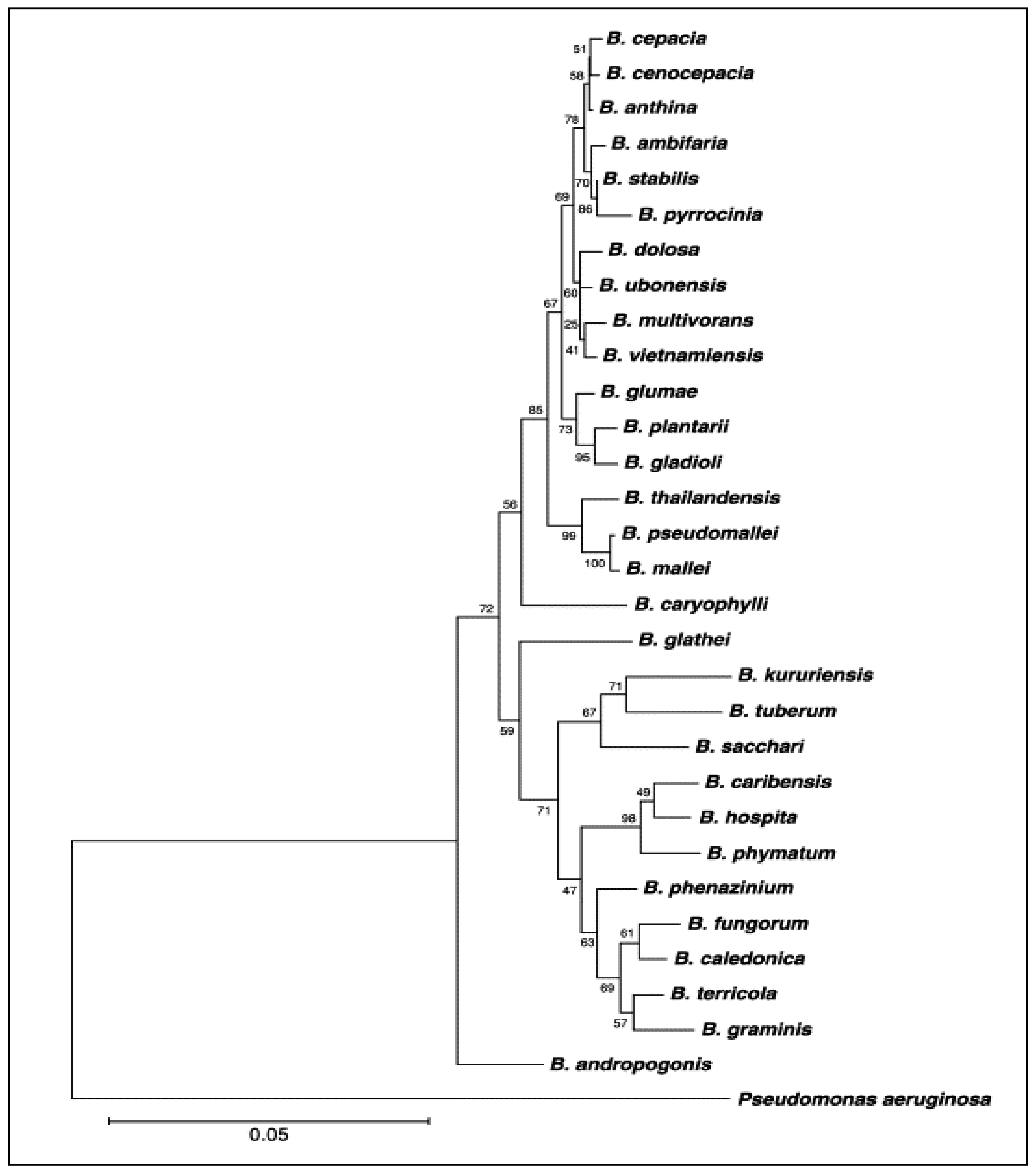
The genus Burkholderia contains about 35 validly named species (Table 1): B. alpina, B. ambifaria, B. andropogonis, B. anthina, B. arboris, B. caryophylli, B. cenocepacia, B. cepacia, B. contaminans, B. diffusa, B. dolosa, B. gladioli, B. glumae, B. humptydooensis, B. lata, B. latens, B. mallei, B. metallica, B. multivorans, B. oklahomensis, B. plantarii, B. pseudomallei, B. pseudomultivorans, B. puraquae, B. pyrrocinia, B. rhizoxinica, B. seminalis, B. singaporensis, B. singularis, B. stabilis, B. stagnalis, B. territorii, B. thailandensis, B. ubonensis and B. vietnamiensis [11]. Figure 1 presents the phylogenetic tree based on the gene sequence of 16S rRNA, showing the position species included in the genus Burkholderia as reported by Coenye and Vandamme [12].
2. Plant Growth Promoting Effect of Burkholderia Spp.
Burkholderia spp. involve diverse mechanisms of action for promoting the plant growth, including rhizosphere competence for secretion of allelochemicals, production of antibiotics and siderophores such as iron chelating compounds [13,14,15]. Ornibactins is considered the predominant siderophores produced by Burkholderia spp. [16].
Pandey et al. [17] reported that the plant growth promoting activity of MSSP strain of genus Burkholderia was determined by some factors such as: 1-aminocyclopropane-1-carboxylic acid deaminase production [18], nitrogen fixation, phosphate solubilization, production of indol acetic acid (IAA), siderophores, and hydrogen cyanide (HCN) [17]. In addition, the same strain showed also an antagonistic activity against different phytopathogens [19].
Karakurt and Aslantas [20] reported that the application of B. gladioli increased the annual shoot diameter of some apple cultivars. Burkholderia spp. strain PsJN is considered an effective plant growth-promoting bacterium since it promotes the growth of potatoes, vegetables and grapevines by producing a high level of 1-aminocyclopropane-1-carboxylic acid deaminase which able to reduce the level of inhibitory hormone ethylene [21]. In addition, Compant et al. [15] studied the growth promoting effect of Burkholderia sp. strain PsJN on Vitis vinifera and concluded that all inoculated plants with this strain have performed better than those non-bacterized and the relative fresh weights of roots and aerial parts were significantly increased compared to the non-bacterized plants.
Karakurt and Aslantas [20] evaluated the effects of some different strains of B. gladioli on the growth and the leaf nutrient content of Starking Delicious, Granny Smith, Starkrimson Delicious, Starkspur Golden Delicious and Golden Delicious apple cultivars grafted on semi-dwarf rootstock and observed an increase of leaf number and area as well as the number of annual shoots and their diameters. Furthermore, the latter authors also reported that the application of B. gladioli has increased the manganese content and did not affect the concentration of sodium and calcium in the leaves [20].
3. Use of Burkholderia Spp. as Biocontrol Agent
Most of Burkholderia species can be used potentially as biocontrol agents against phytopathogenic fungi, bacteria, protozoa and nematodes in many different crops such as: corn, sweet corn, cotton, grapevine, pea, tomato and pepper [22,23]. On the other hand, some Burkholderia species were commercialized and effectively used as biocontrol agents in agriculture [24]. Recently, many researchers have conducted different studies for evaluating the antagonistic effect of Burkholderia spp. for controlling plant diseases since these bacteria are known as producer of many bioactive metabolites such as bacteriocins, alkaloids, lipopeptides and polypeptide [25].
In particular, Holmes et al. [26] studied the capacity of B. cepacia in degradation of chlorinated aromatic substrates in certain synthetic pesticides. Some other strains of Burkholderia produce enzymes able to degrade non-nutritive substrates, such as trichloroethylene (TCE), a major ground water contaminant used in the dry cleaning industry and in degreasing solvents [27]. Other species such as B. bryophila and B. megapolitana showed antifungal activity against some phytopathogens as well as plant growth-promoting properties [28]. Another study has been conducted by Barka et al. [21] reported that strain PsJN of Burkholderia has showed a biocontrol effect against Botrytis cinerea and proved also its growth promoting effect on the grapevine [21].
In addition, several strains of B. gladioli showed an effective in vitro antagonistic activity against a wide range of fungal and bacterial species [29,30,31]. The above-mentioned species could completely inhibit the conidial germination of Penicillium digitatum and Botrytis cinerea, as reported by Walker et al. [32]. In addition, the metabolites produced by B. gladioli caused a significant inhibition of Penicillium expansum, as reported by Elshafie et al. [29]. Apparently, a growth suppression of some pathogenic fungi by B. gladioli strains was more efficient when the bacterial cultures were used than the culture filtrates and this verified the synergic effect of several bioactive substances [29]. However, the antagonizing activity of B. gladioli explained by the combination between competition for nutrients, space and production of antifungal metabolites [30].
Several recent studies showed antimicrobial activity of B. gladioli pv. agaricicola against some serious phytopathogens [29,30,31,32,33,34]. In particular, the pathovar agaricicola showed antagonizing activity against wide range of important phytopathogenic fungi, including Botrytis cinerea, Aspergillus flavus, Aspergillus niger, Penicillium digitatum, Penicillium expansum, Sclerotinia sclerotiorum and Phytophthora cactorum [29]. In the same context, Elshafie et al. [30] reported that four studied strains of B. gladioli pv. agaricicola (ICMP: 11096, 11097, 12220 and 12322) have exerted antifungal activity against above mentioned phytopathogenic fungi by producing diffusible metabolites and extracellular hydrolytic enzymes. The same authors have attributed this bioactivity to the production of two bioactive fatty acids identified as methyl stearate and ethanol 2-butoxy phosphate with mass spectrum m/e 298 and 398, respectively [30].
Another recent study reported that the application of B. gladioli pv. agaricicola strain ICMP 12322 was able to enhance the disease protection and improve the consistency of biological control against tomato-wilt disease caused by Verticillium dahliae [34].
4. Induction of Plant Systemic Resistance (ISR)
The microbial community in soil can play a vital role in stimulation the plant growth and also can suppress the deleterious effect of other soil microorganisms [35]. In particular, Rhizobacteria can reduce the activity of pathogenic microorganisms not only through the microbial antagonism, but also by inducing the plant to defend itself. This phenomenon, named “induced systemic resistance” (ISR), was first described by Van Peer et al. [36]. ISR can be triggered by some specific strains of plant growth promoting bacteria (PGPB) through the production of some plant signaling molecules [37].
Some endophytic bacterial strains belonging to the genus Burkholderia and Bacillus are considered effective biological control agents [38]. The beneficial effects of Burkholderia spp. in agricultural could be explained by induction of plant resistance against abiotic stresses through ISR and others mechanisms [23]. B. phytofirmans strain PsJN-grapevine interaction, a host defense reaction coinciding with phenolic compounds accumulation and strengthening of cell walls in the exodermis and in several cortical cell layers [39]. Sharma and Nowak [40] and Bordiec et al. [41] reported the biocontrol effect of strain PsJN against Verticillium dahliae and Botrytis cinerea, the causal agents of wilt disease and grey mould, respectively.
5. Burkholderia’ Diseases on Human and Animals
Several species of Burkholderia have been reported as rich of virulence factors such as: presence of a flagella, reactive oxygen species resistance and resistance to several antimicrobial drugs [42]. These above mentioned traits of Burkholderia enable them to be adapted perfectly in their different ecological niches [42]. Many species of Burkholderia are known as phytopathogens [43,44] however there are other species belong to Burkholderia have demonstrated some opportunistic infection to animal and human. In particular, B. pseudomallei and B. mallei were considered as pathogens for animals and humans and they are both resistant to a number of antibiotics [45]. B. mallei is responsible for glanders disease, which mostly affected animals, such as horses, mules, donkeys and rarely humans [46]. Whereas, B. pseudomallei is the causal agent of melioidosis, the disease in tropical countries [47]. On the other hand, B. cepacia complex (Bcc) has a natural occurrence in the environment and has both beneficial and detrimental effects on plants, however it is considered an opportunistic human pathogen. Bcc causes severe lung infections in cystic fibrosis patients and it is often resistant to common antibiotics and able to degrade natural and man-made pollutants [48,49,50].
6. Microbial Secondary Metabolites
Most of living organisms, such as invertebrates, plants and microorganisms, are lacking the immune system, hence they have developed the capacity to produce bioactive secondary metabolites including some toxic substances against other harmful microorganisms. These natural products act as specific defense systems against other organisms [51,52]. Secondary metabolites are compounds that are not required for the growth or reproduction but play a vital role in inhibiting the growth of harmful organisms with which they compete and can also inhibit their biologically important processes [53].
Microbial secondary metabolites (MSM) are the most promising source of novel natural products; hence their discovery and characterization are the objective of many researches for controlling important phyto- and human pathogens [53,54,55].
In addition, MSM are low-molecular-mass products of secondary metabolism, usually produced during the late growth phase of microorganisms and their production arises from intracellular intermediates (amino acids, sugars, fatty acids, etc.). MSM are very important for the human health and economics of our society [53,54].
There are thousands of important known MSM, among them penicillin, which was discovered in 1940, obtained from Penicillium moulds, such as P. chrysogenum and P. rubens, which began the era of antibiotics. The penicillin has been recognized as one of the greatest advances in therapeutic medicine [56].
Aspergillus terreus has been reported to produce biological drugs known as statins. The statins are class of drugs that inhibit HMG-CoA reductase and lead to lower cholesterol level [57]. On the other hand, a new substance named lovastatin, with a similar structure of statin, has been extracted from Monascus purpureus and Monascus ruber [58].
6.1. Secondary Metabolites Produced by Genus Burkholderia
Bacterial secondary metabolites (BSM) are considered one of the most promising sources among the novel bioactive pharmaceutical compounds. In particular, Actinobacteria are considered the major source of bioactive BSM, such as different antibiotics, which usually used for human being and animals [59]. Generally, the majority of discovered antimicrobials substances have been isolated from Actinomycetes especially from genus Streptomyces Waksman and Henrici. Among the most important common antibiotics: tetracycline and aminoglycoside or glycopeptide [60].
Several Burkholderia species, considered as beneficial bacteria in the natural environment, have the ability to produce compounds with antimicrobial activity [61] and can be used as biocontrol agents for phytopathogenic fungi and able to inhibit the growth of other bacteria, protozoa and nematodes in many different crops, such as corn, sweet corn, cotton, grapevine, pea, tomato, and pepper [22]. In general, Burkholderia demonstrate significant biotechnological potential as a source of novel antibiotics and bioactive secondary metabolites [62,63].
In particular, genus Burkholderia showed high ability to produce several extracellular hydrolytic enzymes such as chitinase, protease, cellulase, amylase and glucanase [64], which may have important applications in both pharmaceutical industry [29,65]. On the other hand, genus Burkholderia produced also a wide range of secondary metabolites such as pyrrolnitrin, phenazine, cepabactin, and other bioactive diffusible and volatile compounds [34,66,67,68,69].
In the current review, the most important species of genus Burkholderia were reported as following: B. cepacia Palleroni and Holmes (Yabuuchi et al.), B. andropogonis Smith (Gillis et al.), B. plantarii Azegami et al. (Urakami et al.), B. rhizoxinica Partida-Martinez et al., B. glumae Kurita and Tabei (Urakami et al.), B. caryophylli Burkholder (Yabuuchi et al.), and B. gladioli Severini (Yabuuchi et al.).
6.1.1. Burkholderia cepacia
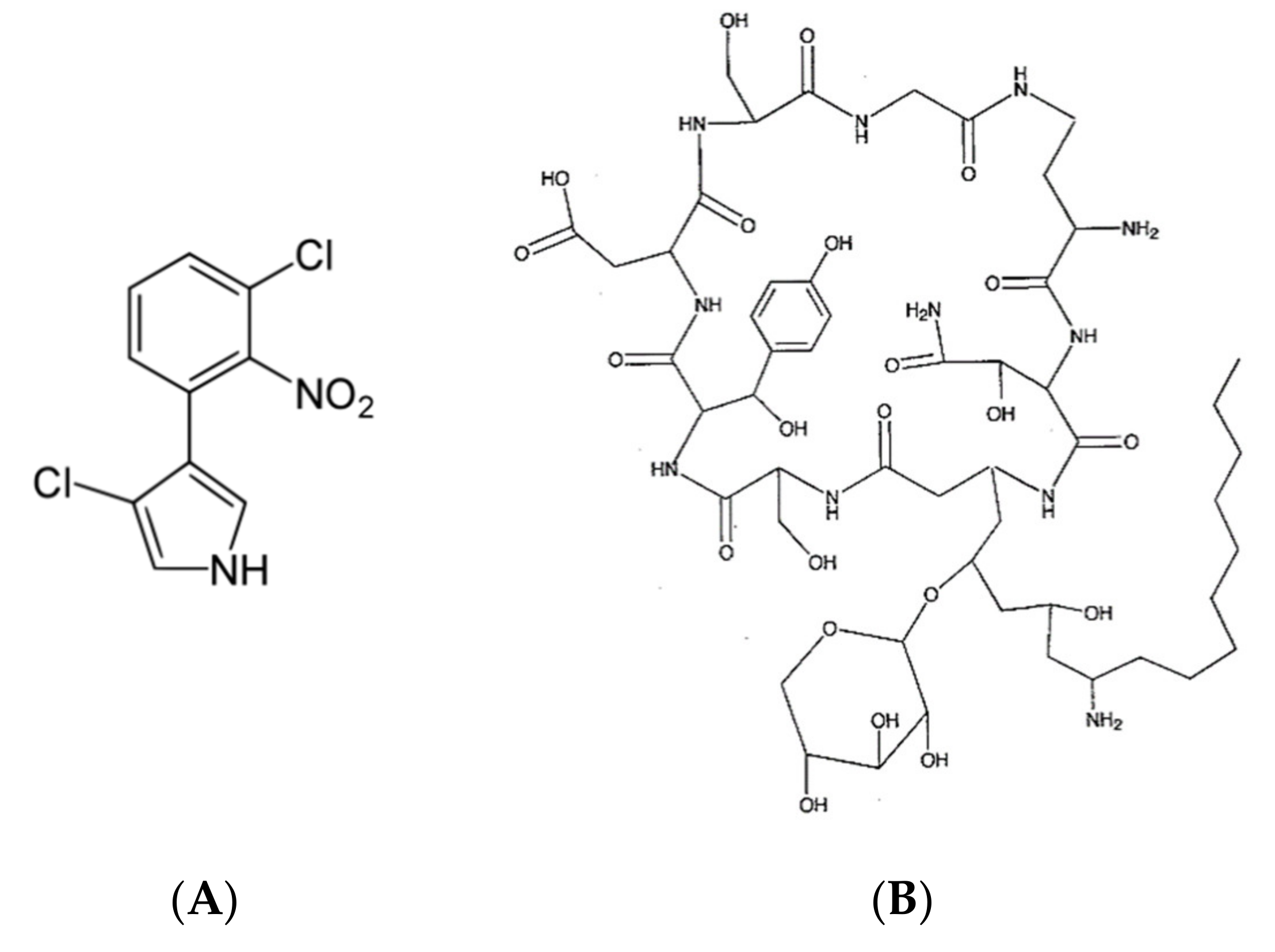
B. cepacia produces a bioactive compound called 3-chloro-4- (2′nitro-3′cloro-phenyl) pyrrole pyrrolnitrin [25,70] (Figure 2A) which showed antimicrobial activity against some pathogenic fungi, yeast and Gram-positive (G+ve) bacteria as reported by Arima et al. [71,72]. Arima et al. [71] reported the molecular formula of pyrrolnitrin C10H6O2N2Cl2 and observed that this compound is a pale-yellow crystal and can loss its bioactivity if exposed to sun light or acidic conditions. Pyrrolnitrin is well solubilized in different organic solvents such as methanol, ethanol, butanol, acetone, ethyl acetate, etc., whereas it is slightly solubilized in water [71]. Rahman et al. [70] reported that the antifungal activity of B. cepacia is due to the chemical toxicity nature of pyrrolnitrin, which can penetrate the cell membrane and leads to the protoplasmic dissolution and disintegration and finally inhibit the cell growth. On the other hand, the same authors explained that the vacuolar appearance of the mycelium may be due to the antibiotic metabolites [70].
Another important antifungal compound produced by B. cepacia is called cepacidine A (Figure 2B) [73] which demonstrated a strong activity against Trichophyton spp. and Epidermophyton spp. Cepacidine A consists of two closely related compounds, cepacidin A1 and A2, with molecular weights of 270.29 Da and 286.29 Da, respectively, in a ratio 9:1. Furthermore, Parker et al. [74] discovered cepacina A and B, two important bioactive compounds, produced by B. cepacia which were able to inhibit the growth of Staphylococcus spp. and some G-ve bacteria. In particular, cepacina B is significantly more active than cepacina A against both G+ve and G-ve [74], whereas both compounds are slightly active against streptococci bacteria. On the other hand, Santos-Villalobos et al. [75] found that siderophores, volatile metabolites produced by B. cepacia, were able to control the growth of Colletotrichum gloeosporioides.
6.1.2. Burkholderia andropogonis
B. andropogonis was first described by Smith [76] as the causal agent of stripe disease of sorghum [77]. It was named before as Pseudomonas andropogonis and then transferred to genus Burkholderia by Gillis et al. [78]. B. andropogonis has been reported also to cause bacterial stripe disease sudangrass, teosinte, johnsongrass, field corn, broomcorn, and sweet corn [79].
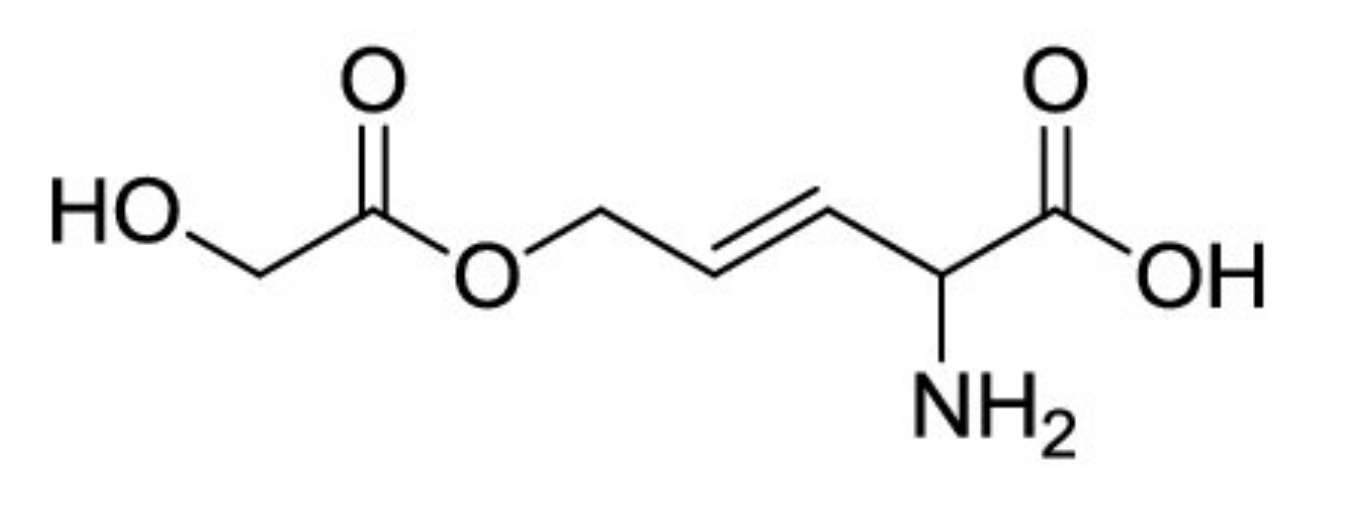
This bacterium produces amino enol ether rhizobitoxine which is responsible for the chlorosis of soybean [80]. The structure of rhizobitoxine, illustrated in Figure 3, was identified by Owens et al. [81]. Rhizobitoxine, with a molecular weight of 190 Da, was able to inhibit the ethylene biosynthesis in apple tissues [82] and reduce the defense reaction by the host plants [83]. The capacity of rhizobitoxine to inhibit the ethylene production may enhance the nodulation and competitiveness in Macroptilium atropurpureum and Vigna radiata [84]. Yasuta et al. [85] reported that rhizobitoxine has significantly inhibited 1-aminocyclopropane-1-carboxylate synthase bLE-ACS2 from tomato, which considered the key enzyme in the pathway of ethylene biosynthesis.
Furthermore, Sugawara et al. [83] explained that rhizobitoxine has strongly inhibited the enzyme [1-aminocyclopropane-1-carboxylate (ACC) synthase] in the ethylene biosynthesis pathway which would explain the early observation of rhizobitoxine inhibition of ethylene evolution in apple tissues. On the other hand, Sugawara et al. [83] reported also the positive role of rhizobitoxine in the symbiosis between Bradyrhizobium elkanii strains and their host legumes. The latter coauthors also reported that rhizobitoxine, as an analog of cystathionine, can irreversibly inhibit β-cystathionase in bacteria and plants.
6.1.3. Burkholderia plantarii
The name of this species derived from the Latin word plantarium (seedbed). B. plantarii is responsible for root rot, seedling blight, chlorosis and reduction of root growth of rice [86]. B. plantarii was found to be distributed on the weeds in fields and in seed stored at room temperature and it was often isolated in association with B. glumae indicating that these two species may have similar transmission path and life cycle [86].
B. plantarii produces a compound called tropolone with molecular weight of 122 Da, which has phenolic and acidic characteristics with antimicrobial activity and phytotoxic effect on rice. Tropolone, identified in 1945, is a non-benzenoid aromatic compound and has similar characteristics of phenols and acids [86]. Trust [87] reported that tropolone showed bacteriostatic and bactericidal effect against wide range of bacterial species such as Bacillus subtilis, Escherichia coli, Pseudomonas aeruginosa, Salmonella typhi and Serratia marcescens. The mechanism of the biological activity of this compound is based mainly on its ability to penetrate the plasma membrane and cell wall of microbes and increasing the cell permeability and leads to cell lysis and subsequent loss of cell contents after rupture of the bleb [87]. Azegami et al. [86] observed that the mere addition of iron to the MA broth culture media has greatly enhanced the growth of P. plantarii. However, the mere addition of ferric chloride has markedly reduced the amount of dissolved tropolone [86].
Furthermore, this bacterium produces another two bioactive compounds identified as: 2-methylene-3-imino-5-L (carboxy-L-valine)-pyrrolidine and 2-methylene-3-imino-5-L (carboxy -L-treoninil)-pyrrolidine with molecular weights of 242.11 Da and 240.13 Da, respectively [88]. Mitchell and Katrina [88] reported that the last two compounds are amino acid conjugates to a new iminopyrrolidine carboxylic acid structure and this is in keeping with the amino acid conjugation characteristic related to many natural compounds that exhibited biological activity [88]. The last two bioactive compounds are able to inhibit Erwinia aylovora, which is responsible for fire blight disease of pome fruit, especially for apple and pear trees [88]. Moreover, B. plantarii strain DSM 9509 produces extracellular rhamnolipids when grown in glucose supplemented rich medium [89]. Rhamnolipids have been used in different applications as detergents and in the pharmaceutical industry [89].
6.1.4. Burkholderia rhizoxinica
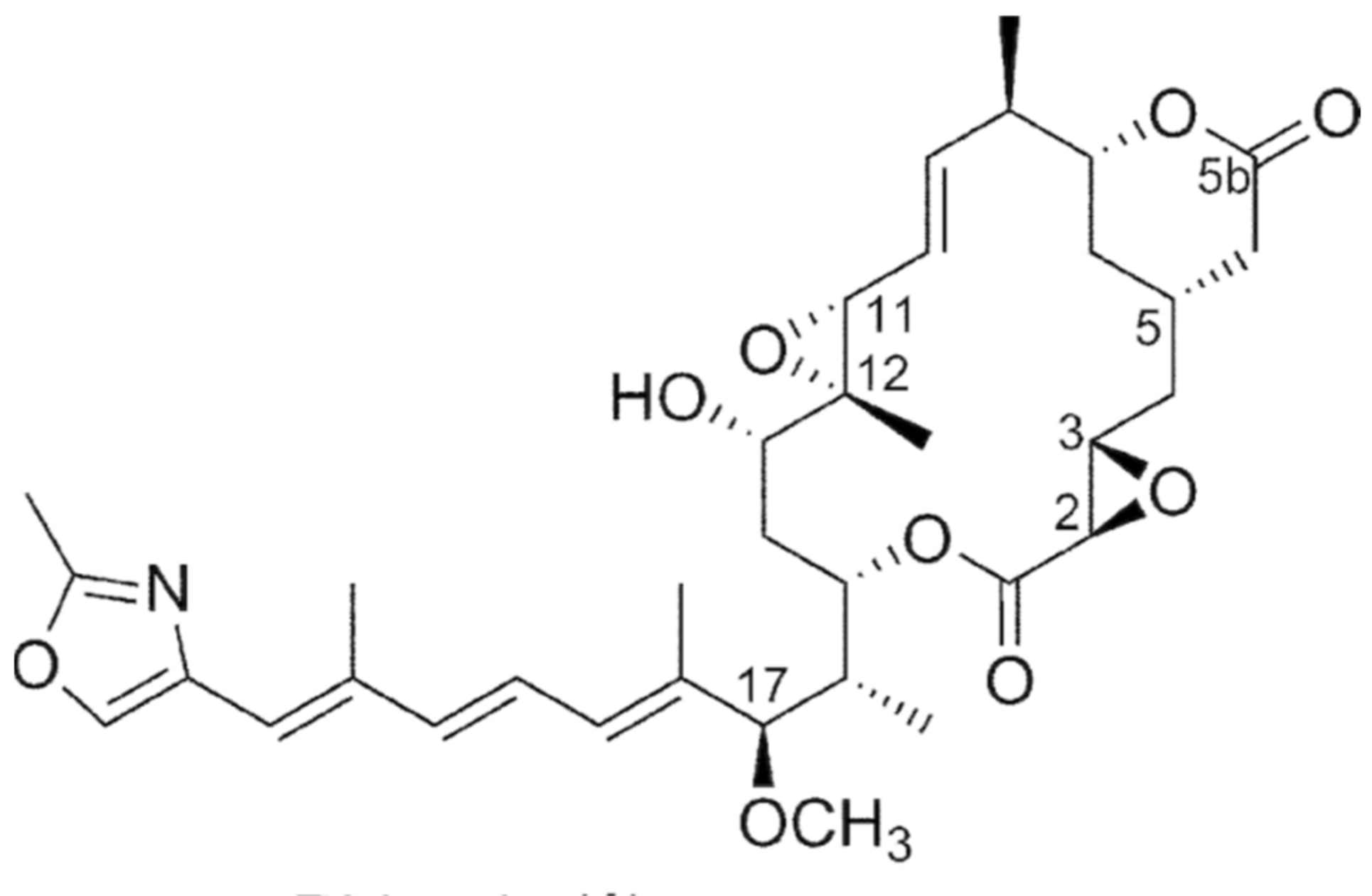
The specific name of this species refers to its ability to produce the rhizoxin antibiotic (Figure 4) [90,91]. B. rhizoxinica is able to grow under aerobic and microaerophilic conditions, but not in an anaerobic atmosphere containing CO2 [91]. Recently, it has been reclassified as Paraburkholderia rhizoxinica Partida-Martinez (Sawana) [92]. B. rhizoxinica is an intracellular symbiont endophytic and was isolated from the phytopathogenic fungus Rhizopus microsporus, a common pathogen for food and feed stuff which causes rice seedling blight [93,94,95]. B. rhizoxinica is now associated with the ability of Rhizopus to cause rice seedling blight [96]. Rhizoxin is an important virulence factor for infection of plants and has phytotoxic, antifungal and anticancer activities [91,96,97].
6.1.5. Burkholderia glumae
B. glumae, the causal agent of bacterial grain rot and seedling rot of rice, was isolated from hot and high relative humid areas [99]. B. glumae was first reported in Japan, but later it was distributed in different countries producing rice such as: Japan, Thailand, Vietnam, South Korea, Malaysia, Philippines, Sri Lanka, United States, Panama, Nicaragua, Costa Rica, and Colombia [100]. The incidence of B. glumae has been increased recently due to climate changes, as well as the deficiency of appropriate management and biocontrol strategies [100].
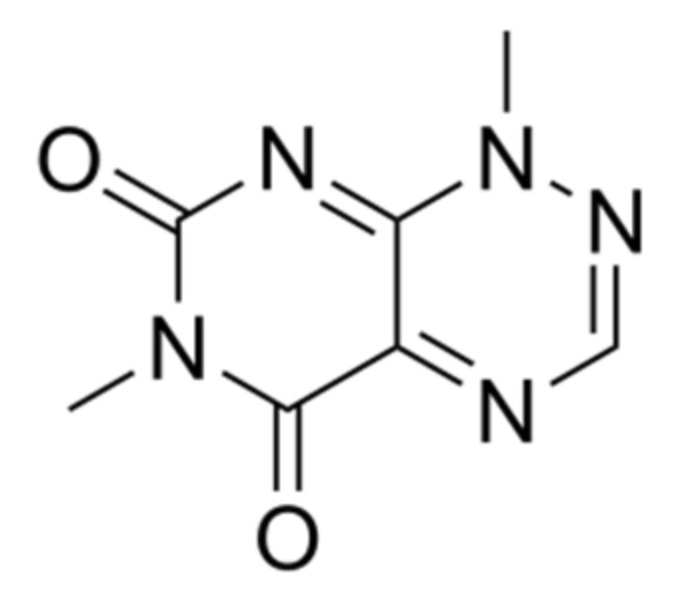
B. glumae produces a range of secondary metabolites and lipase on agar media. In particular, antibiotic production is stimulated by some substrates presented in agar such as K+, Ca2+, Mg2+ and NH4+. Among the active metabolites produced by this bacterium is toxoflavin (Figure 5), which plays a role in pathogenicity of this bacterium and is involved in the rice grain rot [101].
Toxoflavin is a bright yellow color and is highly toxic to plants, fungi, animals and microorganisms [102]. In addition, the toxicity of toxoflavin to plants has led to severe losses in rice crops around the world [102]. The production of this molecule is influenced by temperature whereas the maximum suitable temperature is at 37 °C [103]. Lee et al. [102] reported also that the toxoflavin biosynthesis process is regulated by QS mechanism depending on the homoserine lactone synthesized by cognate receptors TofI and TofR, through the activation of ToxJ and ToxR as transcriptional regulators of toxoflavin biosynthesis.
6.1.6. Burkholderia caryophylli
B. caryophylli, a parasitic endophyte infecting vascular plants, was previously classified as Pseudomonas caryophylli (Burkholder) Starr & Burkholder, is the causal agent of wilt stem cracking and a progressive rot of stems and roots of carnation [104]. It used to be a major problem in carnation production in the USA [105]. B. caryophylli is a soil borne bacterium that overwinters in the rhizosphere of soil forming close interactions with the host plant and soil itself. This species can survive in infected host debris and can infect many different species of the dianthus plant [106].
B. caryophylli produces caryoynencine toxin with molecular weight of 280.31 Da [104]. Caryoynencines are unstable C18 carboxylic acids with conjugated dienetetrayne and polymerize structures [107]. On the other hand, caryoynencine showed potent antimicrobial activity against G+ve and G-ve bacteria especially against the growth of methicillin-resistant Staphylococcus aureus (MESA) [107]. In addition, some analogs of caryoynencine exhibited a broad spectra of activity against the following pathogenic fungi: Tricophyton mentagrophytes, T. interdigitale and T. rubrum which are the causal agents of onychomycosis and tinea pedis in humans [107].
6.1.7. Burkholderia gladioli
B. gladioli is an aerobic G-ve rod-shaped bacterium that may cause disease in human, plants and mushrooms [108]. This species is included in phylum Proteobacteria; class Betaproteobacteria; order Burkholderiales; family Burkholderiaceae and genus Burkholderia. B. gladioli can be distinguished from the other Burkholderia species because it is oxidase negative [108].
B. gladioli was initially identified in gladiolus and successively, associated with other plant diseases such as onions, iris, freesia, dendrobium, cymbidium, tulip, green gram and rice [109]. Disease symptoms varied from the spotting of foliar parts to scabbing and rotting of storage tissues [109]. In the last decade, different strains of B. gladioli have demonstrated the ability to infect human causing severe pulmonary infections in cystic fibrosis and other immune-compromised human patients [110,111]. B. gladioli is closely related to a member of B. cepacia complex that includes ten closely related species which are all plant pathogens [112].
B. gladioli is negative for indole production, nitrate utilization and lysine decorboxylation [9]. On the molecular level, two primers (GLA-f 5′-CGAGCTAATACCGCGAAA-3′ and GLA-r 5′-AGACTCGAGTCAACTGA-3′) were used for the amplification from 16S to 23S region in the B. gladioli genome [10]. The obtained amplicon by using these two above mentioned primers in PCR assay was approximately 300bp [10].
B. gladioli contains four pathovars. Three pathovars, gladioli, alliicola and agaricicola causing soft rots on gladiolus, onion bulbs and mushroom, respectively [7,29,113]. Whereas, the fourth pathovar, cocovenenans causes food spoilage which can be toxic to animal and human being consumers [114]. Differentiation of these four pathovars was made based on hosts, molecular basis and biochemical properties [7,113,114,115,116,117].
B. gladioli pv. gladioli Severini (Yabuuchi et al.) is the causal agent of soft rot of stem bases and corms [118]. On the fern Asplenium nidus (bird’s nest fern), leaf spot and blight have been observed, causing extensive losses in many nurseries in Florida, USA [119].
B. gladioli pv. alliicola Burkholder (Starr and Burkholder) has been isolated recently from onion in the Northeastern Slovenia infecting about 30% of onion bulbs. The internal layers were found to have water-soaked and brown-colored lesions [120]. This pathovar exhibited two different white-yellowish color colonies; one has a slightly wrinkled surface where the other has a smooth surface.
B. gladioli pv. cocovenenas van Damme et al. (Gillis et al.) was isolated from a petroleum- contaminated soil [121] and described as producer for lethal toxins (Bongkrekic acid and toxoflavin) which are toxic to animals [114,122] and was also reported sometimes to cause pneumonia for humans [114].
B. gladioli pv. agaricicola (Bga) Yabuuchi is considered an important pathogen for mushroom [123] because it may cause a significant crop loss [109]. B. gladioli pv. agaricicola causing soft rot and cavity disease on mushroom [114,124]. In particular, some strains of this pathovar causes soft rot on a number of commercially important mushrooms, such as Lentinula edodes, Pleurotus ostreatus, Flammulina velutipes, Pholiota nameko, Hypsizygus marmoreus and Grifola frondosa in Japan and different cultivated Agaricus species in New Zealand and Europe [109,124].
Based on the proposal of Yabuuchi [6] who proposed a new genus, Burkholderia, to include members of the “pseudomallei group”, the pathovar “agaricicola”, previously classified as Pseudomonas gladioli pv. agaricicola [125], was subsequently transferred to the new genus B. gladioli pv. agaricicola [114,126,127].
Secondary metabolites produced from Bga are implicated within the quorum sensing (QS) phenomenon [124,128]. This mechanism enable the bacterial cells to communicate to each other by responding to different signal molecules such as N-Acyl homoserine lactones (N.AHLs) in case of G-ve bacteria [49,129,130]. In particular, recent investigations reported that this pathovar produces N.AHLs which regulates the virulence and other biological activities [29,124].
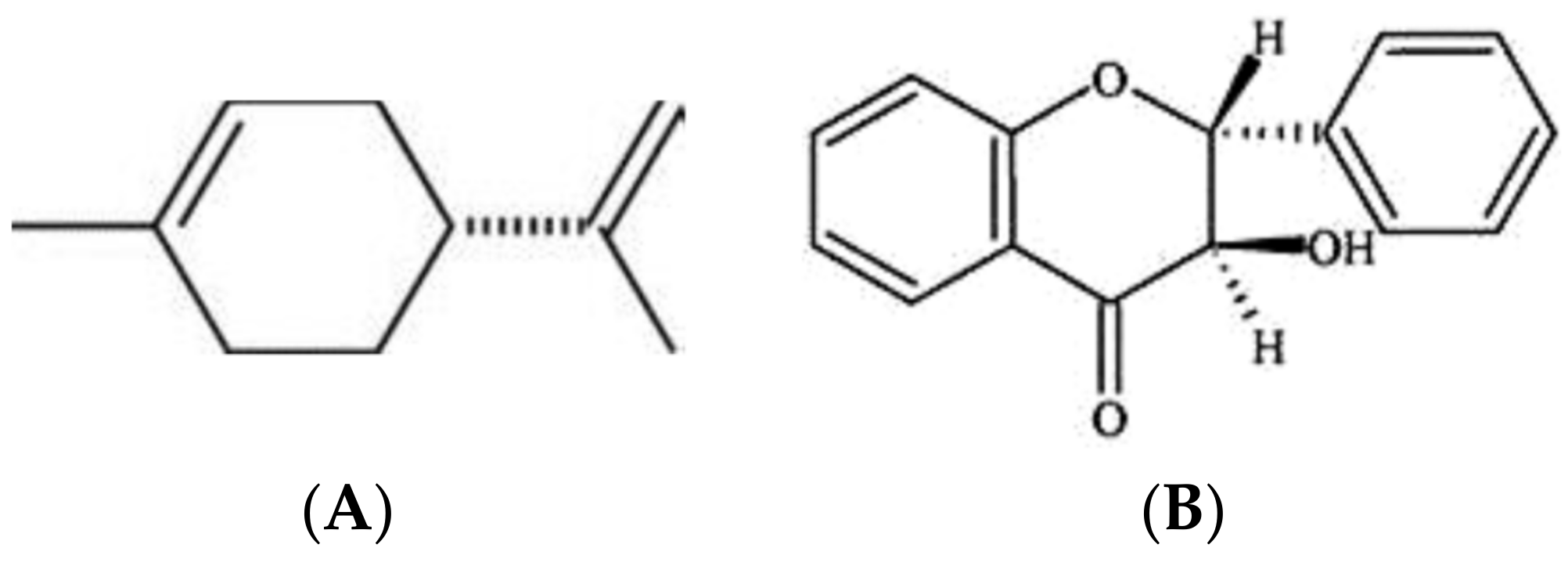
Regarding the volatile organic compounds (VOCs) produced by Bga, they induced a reduction of fungal growth of Fusarium oxysporum and Rhizoctonia solani [131]. The biochemical characterization of VOCs produced by strain ICMP 11096 from this pathovar has identified two bioactive compounds. The first one was a liquid hydrocarbon cyclic terpene identified as cyclohexene 1-methyl-4-(1-methylethenyl) (Figure 6A), as the more frequent d-isomers of limonene [131]. The second one was identified as 4-flavanone (4H-1-Benzopyran-4-one, 2, 3-dihydro-2-phenyl) (Figure 6B). The two produced VOCs could be mainly responsible for the antifungal activity of this pathovar against phytopathogenic and plant-associated fungi [131,132].
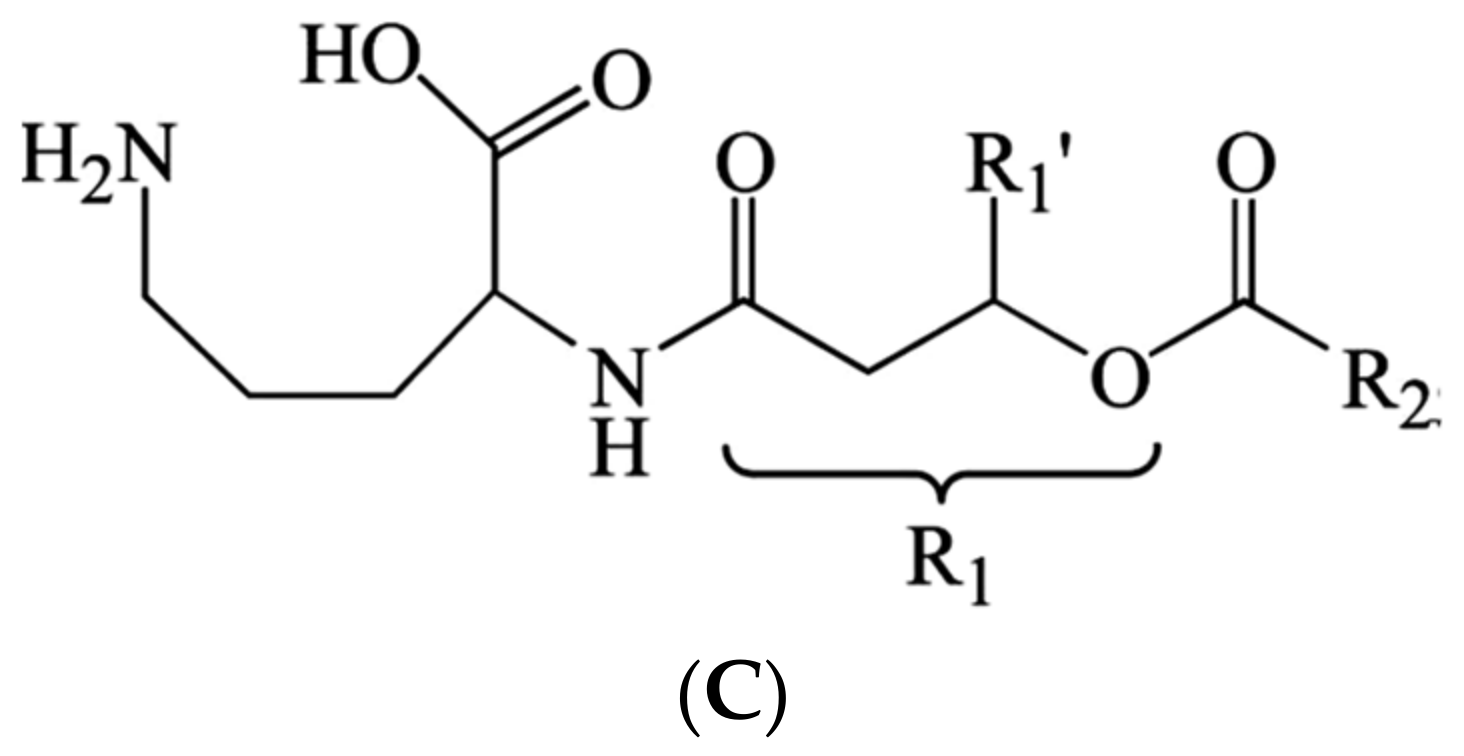
The chemical analysis of the main diffusible secondary metabolites of Bga by using Liquid Chromatography-Mass Spectroscopy (LC-MS) and Nuclear Magnetic Resonance (NMR) investigations demonstrated that the main isolated bioactive diffusible substance is an amino lipid compound identified as ornithine lipid (Figure 6C) [31]. On the other hand, the same authors reported that the ornithine lipid represented a major polar lipid constituent of the whole bacterial cell.
In Table 2, we reported the most important secondary metabolites produced by the above seven species of Burkholderia spp. with their related references.
7. Conclusions
Genus Burkholderia is one of the most important group of plant, animal, and human associated bacteria. It is well-known for its virulence, bioactivity and microbicide properties. This genus includes different species which occupy wide range of ecological niches, such as B. cepacia, B. andropogonis, B. plantarii, B. caryophylli, B. glumae and B. gladioli, which are the causal agents for different plant, animal and human diseases. The current review deals with some important species of Burkholderia which have been manipulated in different studies. It is worth noting to underline that the study of metabolic profile of this genus could aid in revealing different aspects of this group related to its pathogenicity, virulence, plant-microbe interaction and role of produced metabolites in controlling phytopathogens. It is beyond doubt that the knowledge of synthesized secondary metabolites of this group will also support differentiation between different species and eventually strains and pathovars. Detailed information has been reported here regarding some important identified secondary metabolites from different species and pathovars of Burkholderia, their chemical structures, biological activities and modes of action against several phytopathogens. It is concluded that genus Burkholderia has important biological and metabolic properties and can be exploited in promising ways as antagonising biocontrol agents, for soil bioremediation and plant growth promoting purposes. Finally, different synthesized metabolites by Burkholderia can be used effectively in human and agro-pharmaceutical industry.
Author Contributions
Conceptualization, H.S.E. and I.C.; data curation, H.S.E. and I.C.; writing—original draft preparation, H.S.E. and I.C.; writing—review and editing, H.S.E. and I.C.; supervision, I.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Burkholder, W.H. Three bacterial plant pathogens. Phytomonas caryophylli sp.n., Phytomonas alliicola. sp.n., and Phytomonas manihotis (Arthaud-Berthet et Sondar) Viégas. Phytopathology 1942, 32, 141–149. [Google Scholar]
- Burkholder, W.H. Sour skin, a bacterial rot of onion bulbs. Phytopathology 1950, 40, 115–117. [Google Scholar]
- Ferreira, A.S.; Silva, I.N.; Moreira, L.M. Mechanisms controlling the expression of the exopolysaccharide of Burkholderia and role in niche adaptation. Biotechnol. Biopolym. 2011, 8, 147–164. [Google Scholar]
- Wiedmann, M.; Weilmeier, D.; Dineen, S.S.; Ralyea, R.; Boor, K.J. Molecular and Phenotypic Characterization of Pseudomonas spp. Isolated from Milk. Appl. Environ. Microbiol. 2000, 66, 2085–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compant, S.; Kaplan, H.; Sessitsch, A.; Nowak, J.; Ait Barka, E.; Clement, C. Endophytic colonization of Vitis vinifera L. by Burkholderia phytofirmans strain PsJN: From the rhizosphere to inflorescence tissues. FEMS Microbiol. Ecol. 2008, 63, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Yabuuchi, E.; Kosako, Y.; Oyaizu, H.; Yano, I.; Hotta, H.; Hashimoto, Y.; Ezaki, T.; Arakawa, M. Proposal of Burkholderia gen. nov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes 1981) comb. nov. Microbiol. Immunol. 1992, 36, 1251–1275. [Google Scholar] [CrossRef] [PubMed]
- Stoyanova, M.; Pavlina, I.; Moncheva, P.; Bogatzevska, N. Biodiversity and Incidence of Burkholderia Species. Biotechnol. Biotechnol. 2007, 47, 306–310. [Google Scholar] [CrossRef]
- Compant, S.M.; Nowak, J.; Coenye, T.; Clement, C.; Ait Barka, E. Diversity and occurrence of Burkholderia spp. In the natural environment. FEMS Microbiol. Rev. 2008, 32, 607–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graves, M.; Robin, T.; Chipman, A.M.; Wong, J.; Khashe, S.; Janda, J.M. Four additional cases of Burkholderia gladioli infection with microbiological correlates and review. Clin. Infect. 1997, 25, 838–842. [Google Scholar] [CrossRef] [Green Version]
- Furuya, N.; Ura, H.; Iiyama, K.; Matsumoto, M.; Takeshita, M.; Takanami, Y. Specific oligonucleotide primers based on sequences of the 16S-23S rDNA spacer region for the detection of Burkholderia gladioli by PCR. J. Gen. Plant Pathol. 2002, 68, 220–224. [Google Scholar] [CrossRef]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of prokaryotic names with standing in nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Coenye, T.; Vandamme, P. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ. Microbiol. 2003, 5, 719–729. [Google Scholar] [CrossRef] [Green Version]
- Bevivino, A.; Tabacchioni, S.; Chiarini, L.; Carusi, M.V.; Del Gallo, M.; Visca, P. Phenotypic comparison between rhizosphere and clinical isolated of Burkholderia cepacia. Microbiology 1994, 140, 1069–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldani, V.L.; Baldani, J.I.; Dobereiner, J. Inoculation of rice plants with the endophytic diazotrophs Herbaspirillum seropedicae and Burkholderia spp. Biol. Fert. Soils 2000, 30, 485–491. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clement, C.; Barka, E. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.M.; Tran van, V.; Strintzi, A.; Berge, O.; Winkelmann, G. Ornibactin production and transport proprieties in strains of Burkholderia vietnamensis and Burkholderia cepacia. Biometals 1995, 8, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Kang, S.C.; Maheshwari, D.K. Isolation of endophytic plant growth promoting Burkholderia sp. MSSP from root nodules of Mimosa pudica. Curr. Sci. 2005, 98, 177–180. [Google Scholar]
- Honma, M.; Shimomura, T. Metabolism of 1-aminocyclopropane-1- carboxylic acid. Agr. Biol. Chem. 1978, 42, 1825–1831. [Google Scholar]
- Estrade-de los, S.; Bustillos-Cristales, R.; Caballero-Mellado, J. Burkholderia, a genus rich in plant- associated nitrogen fixers with wide environmental and geographic distribution. Appl. Environ. Microbiol. 2001, 67, 2790–2798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karakurt, H.; Aslantas, R. Effects of some plant growth promoting Rhizobacteria (PGPR) strains on plant Growth and leaf nutrient content of apple. J. Fruit Ornamen. Plant Res. 2010, 18, 101–110. [Google Scholar]
- Barka, E.; Gognies, S.; Nowak, J.; Audran, J.C.; Belarbi, A. Inhibitory effect of endophyte bacteria on Botrytis cinerea and its influence to promote the grapevine growth. Biol. Control 2002, 24, 135–142. [Google Scholar] [CrossRef]
- Cain, C.C.; Henry, A.T.; Waldo, R.H.; Casida, L.J.; Falkinham, J.O. Identification and characteristics of a novel Burkholderia strain with broad-spectrum antimicrobial activity. Appl. Environ. Microbiol. 2000, 66, 4139–4141. [Google Scholar] [CrossRef] [Green Version]
- Perin, L.; Martinez-aguilar, L.; Paredes-Valdez, G.; Baldani, J.I.; Estrada-de los, S.; Reis, V.M.; Caballero-Mellado, J. Burkholderia silvatlantica sp. nov. a diazotrophic bacterium associated with sugar cane and maize. Int. J. Syst. Evol. Micriobiol. 2006, 56, 1931–1937. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Ahmedna, M.; Goktepe, I. Effects of processing methods and extraction solvents on concentration and antioxidant activity of peanut skin phenolics. Food Chem. 2005, 90, 199–206. [Google Scholar] [CrossRef]
- El-Banna, N.; Winkelmann, G. Pyrrolnitrin from Burkholderia cepacia: Antibiotic activity against fungi and novel activities against streptomycetes. J. Appl. Microbiol. 1998, 85, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.; Govan, J.; Goldstein, R. Agricultural use of Burkholderia (Pseudomonas) cepacia: A threat to human health. Emerg. Infect. Dis. 1998, 4, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.W.; Doty, S.L. Co-metabolic degradation of trichloroethylene by Burkholderia cepacia G4 with poplar leaf homogenate. Can. J. Microbiol. 2014, 60, 487–490. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, P.; Opelt, K.; Knochel, N.; Berg, C.; Schonmann, S.; De Brandt, E.; Eberl, L.; Falsen, E.; Berg, G. Burkholderia bryophyla sp. nov. and Burkholderia megapolitana sp. nov., moss-associated species with antifungal and plant growthpromoting properties. Int. J. Syst. Evol. Microbiol. 2007, 57, 2228–2235. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Camele, I.; Racioppi, R.; Scrano, L.; Iacobellis, N.S.; Bufo, S.A. In vitro antifungal activity of Burkholderia gladioli pv. agaricicola against some phytopathogenic fungi. Int. J. Mol. Sci. 2012, 13, 6291. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, H.S.; Racioppi, R.; Bufo, S.A.; Camele, I. In vitro study of biological activity of four strains of Burkholderia gladioli pv. agaricicola and identification of their bioactive metabolites using GC–MS. Saudia J. Biol. Sci. 2017, 24, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, H.S.; Viggiani, L.; Mostafa, M.S.; El-Hashash, M.A.; Bufo, S.A.; Camele, I. Biological activity and chemical identification of ornithine lipid produced by Burkholderia gladioli pv. agaricicola ICMP 11096 using LC-MS and NMR analyses. J. Biol. Res. 2017, 9, 96–103. [Google Scholar] [CrossRef]
- Walker, R.; Innes, C.M.; Allan, E.J. The potential biocontrol agent Pseudomonas antimicrobica inhibits germination of conidia and outgrowth of Botrytis cinerea. Lett. Appl. Microbiol. 2001, 32, 346–348. [Google Scholar] [CrossRef] [Green Version]
- Altindag, M.; Sahin, M.; Esitken, A.; Ercisli, S.; Guleryuz, M.; Donmez, M.F. Biological control of brown rot (Moniliana laxa Ehr.) on apricot (Prunus armeniaca L. cv. Hacihaliloglu) by Bacillus, Burkholderia and Pseudomonas application under in vitro and in vivo conditions. Biol. Control 2006, 38, 369–372. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Sakr, S.; Bufo, S.A.; Camele, I. An attempt of biocontrol the tomato-wilt disease caused by Verticillium dahliae using Burkholderia gladioli pv. agaricicola and its bioactive secondary metabolites. Int. J. Plant Biol. 2017, 8, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.C. Plant responses to plant growth-promoting rhizobacteria. Eur. J. Plant Pathol. 2007, 119, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Van Peer, R.; Niemann, G.J.; Schippers, B. Induced resistance and phytoalexin accumulation in biological control of fusarium wilt of carnation by Pseudomonas sp. strain WCS417r. Phytopathology 1991, 81, 728–734. [Google Scholar] [CrossRef]
- Zhang, S.; Moyne, A.L.; Reddy, M.S.; Kloepper, J.W. The role of salicylic acid in induced systemic resistance elicited by plant growth promoting rhizobacteria against blue mould of tobacco. Biol. Control 2002, 25, 288–296. [Google Scholar] [CrossRef]
- Wang, Y.; Zeng, Q.; Zhang, Z.; Yan, R.; Zhu, D. Antagonistic bioactivity of an endophytic bacterium H- 6. Afr. J. Biotechnol. 2010, 9, 6140–6145. [Google Scholar]
- Compant, S.; Reiter, B.; Sessitsch, A.; Nowak, J.; Clement, C.; Barka, E. Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685–1693. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.K.; Nowak, J. Enhancement of verticillium wilt resistance in tomato transplants by in vitro co-culture of seedlings with a plant growth-promoting rhizobacterium (Pseudomonas sp. strain PsJN). Can. J. Microbiol. 1998, 44, 528–536. [Google Scholar] [CrossRef]
- Bordiec, S.; Paquis, S.; Lacroix, H.; Dhond, S.; Barka, E.A.; Kauffmann, S.; Jeandet, P.; Gourbeyre, F.M.; Clement, C.; Baillieul, F.; et al. Comparative analysis of defence responses induced by the endophytic plant growth-promoting rhizobacterium Burkholderia phytofirmans strain PsJN and the non-host bacterium Pseudomonas syringae pv. pisi in grapevine cell suspensions. J. Exp. Bot. 2010, 62, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Chain, P.S.G.; Denef, V.J.; Konstantinidis, K.T.; Vergez, L.M.; Agulló, L.; Reyes, V.L.; Hauser, L.; Córdova, M.; Gómez, L.; González, M.; et al. Burkholderia xenovorans LB400 harbors a multi-replicon, 9.73-Mbp genome shaped for versatility. Proc. Natl. Acad. Sci. USA 2006, 103, 15280–15287. [Google Scholar] [CrossRef] [Green Version]
- European and Mediterranean Plant Protection Organization. Burkholderia caryophylli. OEPP/EPPO Bull. 2006, 36, 95–98.
- OEPP/EPPO, EPPO Standard PP 2/1(1) Guideline on good plant protection practice: Principles of good plant protection practice. OEPP/EPPO Bull. 1994, 24, 233–240. [CrossRef]
- Dance, D.A.B. Melioidosis And Glanders as Possible Biological Weapons. In Bioterrorism and Infectious Agents: A New Dilemma for the 21st Century; Fong, I.W., Alibek, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 99–145. [Google Scholar]
- Whitlock, G.C.; Estes, D.M.; Torres, A.G. Glanders: Off to the races with Burkholderia mallei. FEMS Microbiol. Lett. 2007, 277, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Ngamdee, W.; Tandhavanant, S.; Wikraiphat, C.; Reamtong, O.; Wuthiekanun, V.; Salje, J.; Low, D.A.; Peacock, S.J.; Chantratita, N. Competition between Burkholderia pseudomallei and B. thailandensis. BMC Microbiol. 2015, 15, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahenthiralingam, E.; Urban, T.A.; Goldberg, J.B. The multifarious, multireplicon Burkholderia cepacia complex. Nat. Rev. Microbiol. 2005, 3, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Lipuma, J.J. Update on the Burkholderia cepacia complex. Curr. Opin. Pulm. Med. 2005, 11, 528–533. [Google Scholar] [CrossRef]
- Peeters, C.; Zlosnik, J.E.; Spilker, T.; Hird, T.J.; LiPuma, J.J.; Vandamme, P. Burkholderia pseudomultivorans sp. nov., a novel Burkholderia cepacian complex species from human respiratory samples and the rhizosphere. Syst. Appl. Microbiol. 2013, 36, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Wright, J.L. Attack and Defend: The Function and Evolution of Bioactive or Toxic Metabolites. In Proceedings of the Xth International Conference on Harmful Algae, St. Petersburg, FL, USA, 21–25 October 2002. [Google Scholar]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. “Chapter 24: Natural Products (Secondary Metabolites)”. In Natural Products in Chemical Biology; Civjan, N., Ed.; Wiley: Hoboken, NJ, USA, 2012; pp. 1250–1319. ISBN 978-1-118-10117-9. [Google Scholar]
- Sanchez, S.; Demain, A.L. Comprehensive Biotechnology, 3rd ed.; Moo-Young, M., Ed.; Pergamon: Oxford, UK, 2011; pp. 131–143. [Google Scholar]
- Camele, I.; Elshafie, H.S.; Caputo, L.; Sakr, S.H.; De Feo, V. Bacillus mojavensis: Biofilm formation and biochemical investigation of its bioactive metabolites. J. Biol. Res. 2019, 92, 39–45. [Google Scholar] [CrossRef]
- Samson, R.A.; Pitt, J.I. Integration of Modern Taxonomic Methods for Penicillium and Aspergillus Classification; CRC Press: Boca Raton, FL, USA, 2000; p. 66. [Google Scholar]
- Subhan, M.; Faryal, R.; Macreadie, I. Exploitation of Aspergillus terreus for the Production of Natural Statins. J. Fungi 2016, 2, 13. [Google Scholar] [CrossRef]
- Panda, B.; Javed, S.; Ali, M. Optimization of fermentation parameters for higher lovastatin production in red mold rice through co-culture of Monascus purpureus and Monascus ruber. Food Bioprocess Technol. 2010, 3, 373–378. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Mast, Y.; Stegmann, E. Actinomycetes: The Antibiotics Producers. Antibiotics 2019, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.; Young, J.M. Biocidal activity in plant pathogenic Acidovorax, Burkholderia, Herbaspirillum, Ralstonia, and Xanthomonas sp. J. Appl. Microbiol. 1998, 84, 263–271. [Google Scholar] [CrossRef]
- Depoorter, E.; Bull, M.J.; Peeters, C.; Coenye, T.; Vandamme, P.; Mahenthiralingam, E. Burkholderia: An update on taxonomy and biotechnological potential as antibiotic producers. Appl. Microbiol. Biotechnol. 2016, 100, 5215–5229. [Google Scholar] [CrossRef] [PubMed]
- Kunakom, S.; Eustaquio, A.S. Burkholderia as a source of natural products. J. Nat. Prod. 2019, 82, 2018–2037. [Google Scholar] [CrossRef] [PubMed]
- Al-ahmadi, J.K.; Yazdi, T.M.; Najafi, F.M.; Shahverdi, A.R.; Faramarzi, M.A.; Zarrini, G.; Behravan, J. Optimization of medium and cultivation conditions for chitinase production by the newly isolated: Aeromonas sp. Biotechnology 2008, 7, 266–272. [Google Scholar]
- Wang, S.; Hwang, J. Microbial reclamation of shellfish wastes for the production of chitinases. Enzyme. Microb. Technol. 2001, 28, 376–382. [Google Scholar] [CrossRef]
- Roitman, J.N.; Mahoney, N.E.; Janisiewicz, W.J. Production and composition of phenylpyrrole metabolites produced by Pseudomonas cepacia. Appl. Microbiol. Biotechnol. 1990, 34, 381–386. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Iacobellis, N.S.; Simmaco, M.; Graniti, A. Biological properties and spectrum of activity of Pseudomonas syringae pv. syringae toxins. Physiol. Mol. Plant Pathol. 1997, 50, 129–140. [Google Scholar] [CrossRef]
- Cartwright, D.K.; Chilton, W.S.; Benson, D.M. Pyrrolnitrin and phenazine production by Pseudomonas cepacia, strain 5.5B, a biocontrol agent of Rhizoctonia solani. Appl. Microbiol. Biotechnol. 1995, 43, 211–216. [Google Scholar] [CrossRef]
- Liu, X.; Bimerew, M.; Ma, Y.; Muller, H.; Ovadis, M.; Eberl, L.; Berg, G.; Chernin, L. Quorum-sensing signaling is required for production of the antibiotic pyrrolnitrin in a rhizospheric biocontrol strain of Serratia plymuthica. FEMS Microbiol. Lett. 2007, 270, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Mahmud, T.M.; Kadir, J.; Abdul Rahman, R.; Begum, M.M. Screening of antagonistic bacteria for biocontrol activities on Colletotrichum gloeosporioides in papaya. Asian J. Plant Sci. 2007, 6, 12–20. [Google Scholar] [CrossRef]
- Arima, K.; Imanaki, H.; Kousaka, M.; Fukuda, A.; Tamura, G. Studies on pyrrolnitrin, a new antibiotic. I. Isolation and properties of pyrrolnitrin. J. Antibiot. 1965, 8, 201–204. [Google Scholar]
- Arima, K.; Imanaki, H.; Kousaka, M.; Fukuta, A.; Tamura, G. Pyrrolnitrin, a new antibiotic substance, produced by Psneudomonas. Agr. Biol. Chem. 1964, 28, 575–576. [Google Scholar] [CrossRef]
- Lee, C.; Kim, S.; Hyun, B.; Suh, J.; Yon, C.; Kim, C.; Lim, Y.; Kim, C. Cepacidine A, a novel antifungal antibiotic produced by Pseudomonas cepacia. J. Antibiot. 1994, 47, 1402–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, W.L.; Rathnum, M.L.; Seiner, V.; Trejo, W.H.; Principe, P.A.; Sykes, R.B. Cepacin A and Cepacin B, two new antibiotics produced by Pseudomonas cepacia. J. Antibiot. 1984, 37, 431–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Villalobos, S.; Barrera-Galicia, G.C.; Miranda-Salcedo, M.A.; Peña-Cabriales, J.J. Burkholderia cepacia XXVI siderophore with biocontrol capacity against Colletotrichum gloeosporioides. World. J. Microbiol. Biotechnol. 2012, 28, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.F. Bacteria in relation to plant diseases. Carnegie Inst Washington Publ. 1911, 2, 1–368. [Google Scholar]
- Takahashi, Y.; Takahashi, K.; Watanabe, K.; Kawano, T. Bacterial black spot caused by Burkholderia andropogonis on Ondontoglossum and intergeneric hybrid orchids. J. Gen. Plant Pathol. 2004, 70, 284–287. [Google Scholar] [CrossRef]
- Gillis, M.; Van, T.; Bardin, R.; Goor, M.; Hebbar, P.; Willems, A.; Segers, P.; Kersters, K.; Heulin, T.; Fernandez, M.P. Polyphasic taxonomy in the genus Burkholderia leading to an emended description of the genus and proposition of Burkholderia vietnamiensis sp. nov. for N2 Fixing isolates from rice in Vietnam. Int. J. Syst. Bacteriol. 1995, 45, 274–289. [Google Scholar] [CrossRef]
- Vidaver, A.K.; Carlson, R.R. Leaf spot of field corn caused by Pseudomonas andropogonis. Plant Dis. Rep. 1978, 62, 213–216. [Google Scholar]
- Coenye, T.; Laevens, S.; Gillis, M.; Vandamme, P. Genotypic and chemotaxonomic evidence for the reclassification of Pseudomonas woodsii (Smith 1911) Stevens 1925 as Burkholderia andropogonis (Smith 1911) Gillis et al. 1995. Int. J. Syst. Evol. Microbiol. 2001, 51, 183–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, L.D.; Thompson, J.F.; Pitcher, R.G.; Williams, T. Structure of rhizobitoxine, an antimetabolic enol-ether amino acid from Rhizobium japonicum. J. Chem. Soc. Chem. Commun. 1972, 12, 714. [Google Scholar] [CrossRef]
- Owens, L.D.; Lieberman, M.; Kunishi, A. Inhibition of ethylene production by rhizobitoxine. Plant Physiol. 1971, 48, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, M.; Okazaki, S.; Nukui, N.; Ezura, H.; Mitsui, H.; Minamisawa, K. Rhizobitoxine modulates plant-microbe interactions by ethylene inhibition. Biotech. Adv. 2006, 24, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Duodu, S.; Bhuvaneswari, T.V.; Stokkermans, T.J.W.; Peters, N.K. A Positive role for rhizobitoxine in Rhizobium-legume symbiosis. Mol. Plant-Microbe Interact. 1999, 12, 1082–1089. [Google Scholar] [CrossRef]
- Yasuta, T.; Satoh, S.; Minamisawa, K. New assay for rhizobitoxine based on inhibition of 1-Aminocyclopropane-1-Carboxylate Synthase. Appl. Environ. Microbiol. 1999, 65, 849–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azegami, K.; Nishiyama, K.; Kato, H. Effect of iron limitation on Pseudomonas plantarii growth and tropolone and protein production. Appl. Environ. Microbiol. 1988, 54, 844–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trust, T.J. Antibacterial Activity of Tropolone. Antimicrob. Agen. Chemoth. 1975, 7, 500–506. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.E.; Katrina, L.T. Antibacterial iminopyrrolidines from Burkholderia plantarii, a bacterial pathogen of rice. Org. Biomol. Chem. 2005, 3, 3540–3543. [Google Scholar] [CrossRef]
- Hörmann, B.; Müller, M.M.; Syldatk, C.; Hausmann, R. Rhamnolipid production by Burkholderia plantarii DSM 9509T. Eur. J. Lipid Sci. Technol. 2010, 112, 674–680. [Google Scholar] [CrossRef]
- Laila, P.; Hertweck, C. Pathogenic fungus harbours endosymbiotic bacteria for toxin production. Nature 2005, 437, 884–888. [Google Scholar]
- Partida-Martinez, L.P.; Groth, I.; Schmitt, I.; Richter, W.; Roth, M.; Hertweck, C. Burkholderia rhizoxinica sp. nov. and Burkholderia endofungorum sp. nov., bacterial endosymbionts of the plant-pathogenic fungus Rhizopus microsporus. Int. J. Sys. Evolut. Microbiol. 2007, 57, 2583–2590. [Google Scholar] [CrossRef]
- Sawana, A.; Adeolu, M.; Gupta, R.S. Molecular signatures and phylogenomic analysis of the genus Burkholderia: Proposal for division of this genus into the emended genus Burkholderia containing pathogenic organisms and a new genus Paraburkholderia gen. nov. harboring environmental species. Front. Genet. 2014, 5, 429. [Google Scholar] [CrossRef] [Green Version]
- Jennessen, J.; Nielsen, K.F.; Houbraken, J.; Lyhne, E.K.; Schnrer, J.; Frisvad, J.C.; Samson, R.A. Secondary metabolite and mycotoxin production by the Rhizopus microsporus group. J. Agric. Food Chem. 2005, 53, 1833–1840. [Google Scholar] [CrossRef]
- Gee, J.E.; Glass, M.B.; Lackner, G.; Helsel, L.O.; Daneshvar, M.; Hollis, D.G.; Jordan, J.; Morey, R.; Steigerwalt, A.; Hertweck, C. Characterization of Burkholderia rhizoxinica and Burkholderia endofungorum isolated from clinical specimens. PLoS ONE 2011, 6, e15731. [Google Scholar] [CrossRef] [PubMed]
- Lackner, G.; Moebius, N.; Partida-Martinez, L.; Hertweck, C. Complete genome sequence of Burkholderia rhizoxinica, an endosymbiont of Rhizopus microspores. J. Bacteriol. 2011, 193, 783–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikubo, S.; Takigawa, N.; Ueoka, H.; Kiura, K.; Tabata, M.; Shibayama, T.; Chikamori, M.; Aoe, K.; Matsushita, A.; Harada, M. In vitro evaluation of antimicrotubule agents in human small-cell lung cancer cell lines. Anticancer Res. 1999, 19, 3985–3988. [Google Scholar]
- Loper, J.E.; Henkels, M.D.; Shaffer, B.; Valeriote, F.A.; Gross, H. Isolation and identification of rhizoxin analogs from Pseudomonas fluorescens Pf-5 by using a genomic mining strategy. App. Environ. Microbiol. 2008, 74, 3085–3093. [Google Scholar] [CrossRef] [Green Version]
- Partida-Martinez, L.P.; Hertweck, C. A gene cluster encoding rhizoxin biosynthesis in “Burkholderia rhizoxina”,the bacterial endosymbiont of the fungus Rhizopus Microsporus. Chem. Biochem. 2007, 8, 41–45. [Google Scholar]
- Agbagala, M.L. Occurrence and Survival of Burkholderia glumae (Kurita and Tabei, Comb. nov.) Yabuuchi et.al. in Rice (Oryza sativa L.) Seeds. Master’s Thesis, University of Philippines, Los Baños, Laguna, Philippines, 1997. [Google Scholar]
- Pedraza, L.A.; Bautista, J.; Uribe-Vélez, D. Seed-born Burkholderia glumae infects rice seedling and maintains bacterial population during vegetative and reproductive growth stage. Plant Pathol. J. 2018, 34, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Yeonhwa, J.; Jinwoo, K.; Suhyun, K.; Yongsung, K.; Tomohisa, N.; Tsushima, O. Toxoflavin Produced by Burkholderia glumae causing rice grain rot is responsible for inducing bacterial wilt in many field crops. Plant Dis. 2007, 87, 890–895. [Google Scholar]
- Lee, J.; Park, J.; Kim, S.; Park, I.; Seo, Y. Differential regulation of toxoflavin production and its role in the enhanced virulence of Burkholderia gladioli. Mol. Plant Pathol. 2016, 17, 65–76. [Google Scholar] [CrossRef]
- Kim, J.; Kim, J.G.; Kang, Y.; Nagamatsu, T.; Hwang, I. Quorum sensing and the Lys-R type transcriptional activator ToxR regulate toxoflavin biosynthesis and transport in Burkholderia glumae. Mol. Microbiol. 2004, 54, 21–34. [Google Scholar] [CrossRef]
- Kusumi, T.; Ohtani, I.; Nishiyamaa, K.; KLakisawa, H. Caryoynencins, potent antibiotics from a plant pathogen Pseudomonas caryophylli. Tetrahedron Lett. 1987, 34, 3981–3984. [Google Scholar] [CrossRef]
- Gregory, C.L. Carnation bacterial wilt in Indiana. USDA Plant Dis. Rep. 1942, 26, 413. [Google Scholar]
- Scientific Opinion on the risk to plant health posed by Burkholderia caryophylli for the EU territory with the identification and evaluation of risk reduction options. EFSA J. 2013, 11, 3071. [CrossRef]
- Yamaguchi, M.; Park, H.; Ishizuka, S.; Omata, K.; Hirama, M. Chemistry and antimicrobial activity of caryoynencins analogs. Afric. J. Biotechnol. 1995, 7, 2227–2231. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Vandamme, P. Burkholderia: Molecular Microbiology and Genomics; Horizon Bioscience: Norfolk, UK, 2007; pp. 203–228. [Google Scholar]
- Chowdhury, P.R.; Heinemann, J.A. The General secretory pathway of Burkholderia gladioli pv. agaricicola BG164R is necessary for cavity disease in white button mushrooms . Appl. Environ. Microbiol. 2006, 72, 3558–3565. [Google Scholar]
- Wilsher, M.L.; Kolbe, J.; Morris, A.J.; Welch, D.F. Nosocomial acquisition of Burkholderia gladioli in patients with cystic fibrosis. Am. J. Resp. Crit. Care Med. 1997, 156, 1436–1440. [Google Scholar] [CrossRef]
- UK Cystic Fibrosis Trust Infection Control Group. The Burkholderia Cepacia Complex. In Suggestions for Prevention and Infection Control, 2nd ed.; 2004; p. 32.
- Henry, D.A.; Mahenthiralingam, E.; Vandamme, P.; Coenye, T.; Speert, D.P. Phenotypic methods for determining genomovar status of the Burkholderia cepacia Complex. J. Clin. Microbiol. 2001, 39, 1073–1078. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, J. Guide to Plant Pathogenic Bacteria; CAB International: Wallingford, UK, 1986. [Google Scholar]
- Jiao, Z.; Kawamura, Y.; Mishima, N.; Yang, R.; Li, N.; Liu, X.; Ezaki, T. Need to differentiate lethal toxin-producing strains of Burkholderia gladioli, which cause severe food poisoning: Description of Burkholderia gladioli pv. cocovenenans and an emended description of Burkholderia gladioli. Microbiol. Immunol. 2003, 47, 915–925. [Google Scholar] [CrossRef]
- Uchida, J. Bacterial Diseases of Dendrobium; HITAHR, College of Tropical Agriculture and Human Resources, University of Hawaii: Honolulu, HI, USA, 1995; p. 158. [Google Scholar]
- Keith, L.M.; Kewake, S.; Zee, F.T. Isolation and characterization of Burkholderia gladioli from orchids in Hawaii. Plant Dis. 2005, 89, 1273–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ura, H.; Furuya, N.; Iiyama, K.; Hidaka, M.; Tsuchiya, K.; Matsuyama, N. Burkholderia gladioli associated with symptoms of bacterial grain rot and leaf- sheath browning of rice plants. J. Gen. Pl. Pathol. 2006, 72, 98–103. [Google Scholar] [CrossRef]
- Scortichini, M. Pseudomonas gladioli pv. gladioli causal agent of bacterial scab of Gladiolus in Lazio. Informatore Fitopatologico 1993, 43, 49–51. [Google Scholar]
- Chase, A.R.; Miller, J.W.; Jones, J.B. Leaf spot and blight of Asplenium nidus caused by Pseudomonas gladioli. Plant Dis. 1984, 68, 344–347. [Google Scholar] [CrossRef]
- Lamovšek, J.; Gerič Stare, B.; Žerjav, M.; Urek, G. Soft rot of onion bulbs caused by Burkholderia gladioli pv. alliicola in slovenia. J. Plant Pathol. 2016, 98, 369–377. [Google Scholar]
- Wong, J.W.; Lai, K.M.; Wan, C.K.; Ma, K.K.; Fang, M. Isolation and optimisation of PAH-degradative bacteria from contaminated soil for PAH bioremediation. Water Air Soil Pollut. 2002, 139, 1–13. [Google Scholar] [CrossRef]
- Levenberg, B.; Linton, S.N. On biosynthesis of toxoflavin, an azapteridine antibiotic produced by Pseudomonas cocovenenans. J. Biolog. Chem. 1966, 241, 846–852. [Google Scholar] [CrossRef]
- Gill, W.M. Bacterial diseases of Agaricus Mushrooms. Rep. Tottori. Mycol. Inst. 1995, 33, 34–55. [Google Scholar]
- Elshafie, H.S.; Devescovi, G.; Venturi, V.; Camele, I.; Bufo, S.A. Study of the regulatory role of N-Acyl homoserine lactones mediated quorum sensing in the biological activity of Burkholderia gladioli pv. agaricicola causing soft rot of Agaricus spp. Front. Microbiol. 2019, 10, 2695. [Google Scholar] [CrossRef] [PubMed]
- Gill, W.M.; Tsuneda, A. The interaction of the soft rot bacterium Pseudomonas gladioli pv. agaricicola with Japanese cultivated mushrooms. Can. J. Microbiol. 1997, 43, 639–648. [Google Scholar] [CrossRef]
- Chowdhury, P.R. Biology and Genetic Characterization of Burkholderia gladioli pv. agaricicola, the Causal Organism of ‘Cavity Disease’ of White Button Mushrooms. Ph.D. Thesis, School of Biological Sciences at the University of Canterbury, Christchurch, New Zealand, 2004; p. 208. [Google Scholar]
- Lunch, K.H.; Dennis, J.J. Burkholderia. In Molecular Detection of Foodborn Pathogens; Taylor and Francis Group, Liu, D., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 331–340. [Google Scholar]
- Steindler, L.; Venturi, V. Detection of quorum-sensing N-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol. Lett. 2007, 266, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Camele, I.; Elshafie, H.S.; De Feo, V.; Caputo, L. Anti-quorum sensing and antimicrobial effect of mediterranean plant essential oils against phytopathogenic bacteria. Front. Microbiol. 2019, 10, 2019. [Google Scholar]
- Withers, H.; Swift, S.; Williams, P. Quorum sensing as an integral component of gene regulatory networks in Gram-negative bacteria. Curr. Opin. Microbiol. 2001, 4, 186–193. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Bufo, S.A.; Racioppi, R.; Camele, I. Biochemical characterization of volatile secondary metabolites produced by Burkholderia gladioli pv. agaricicola. Int. J. Drug Discov. 2013, 5, 181–184. [Google Scholar]
- Ayoola, G.A.; Johnson, O.O.; Adelowotan, T.; Aibinu, I.E.; Adenipekun, E.; Adepoju-Bello, A.A.; Coker, H.A.; Odugbemi, T.O. Evaluation of the chemical constituents and the antimicrobial activity of the volatile oil of Citrus reticulata fruit (Tangerine fruit peel) from South West Nigeria. Afr. J. Biotechnol. 2008, 7, 2227–2231. [Google Scholar]
Figure 1.
Phylogenetic tree of genus Burkholderia based on 16S rRNA gene sequence. This phylogenetic tree is in agree with Coenye and Vandamme [12].
Figure 1.
Phylogenetic tree of genus Burkholderia based on 16S rRNA gene sequence. This phylogenetic tree is in agree with Coenye and Vandamme [12].

Figure 2.
Secondary metabolites produced by Burkholderia cepacia. Where (A) 3-chloro-4-(2′nitro-3′cloro-phenyl) pyrrole pyrolnitrin and (B) Cepacidine A.
Figure 2.
Secondary metabolites produced by Burkholderia cepacia. Where (A) 3-chloro-4-(2′nitro-3′cloro-phenyl) pyrrole pyrolnitrin and (B) Cepacidine A.

Figure 3.
Chemical structure of rhizobitoxine.

Figure 4.
Chemical structure of rhizoxin.

Figure 5.
Chemical structure of toxoflavin.

Figure 6.
Secondary metabolites produced by B. gladioli pv. agaricicola. Where (A) D-Limonene, (B) 4-Flavanone, (C) Ornithine lipid.
Figure 6.
Secondary metabolites produced by B. gladioli pv. agaricicola. Where (A) D-Limonene, (B) 4-Flavanone, (C) Ornithine lipid.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of species of Burkholderia mentioned in the current review.
| Species of Burkholderia | Author | Year of Description | Disease | Host | |
|---|---|---|---|---|---|
| 1 | B. alpina | Weber and King | 2017 | - | - |
| 2 | B. ambifaria | Coenye et al. | 2001 | belongs to B. cepacia complex | |
| 3 | B. andropogonis | Smith (Gillis et al.) | 1911, 1995 | bacterial leaf stripe | sorghum and corn |
| 4 | B. anthina | Vandamme et al. | 2002 | belongs to B. cepacia complex | |
| 5 | B. arboris | Vanlaere et al. | 2008 | belongs to B. cepacia complex | |
| 6 | B. caryophylli | Burkholder (Yabuuchi et al.) | 1942, 1993 | wilt, stem cracking and rot of stems and roots | carnation |
| 7 | B. cenocepacia | Vandamme et al. | 2003 | cystic fibrosis | humans |
| 8 | B. cepacia | Palleroni and Holmes (Yabuuchi et al.) | 1981, 1993 |
cystic fibrosis soft-rotting |
humans onion |
| 9 | B. contaminans | Vanlaere et al. | 2009 | belongs to B. cepacia complex | |
| 10 | B. diffusa | Vanlaere et al. | 2008 | belongs to B. cepacia complex | |
| 11 | B. dolosa | Vermis et al. | 2004 | belongs to B. cepacia complex | |
| 12 | B. gladioli | Severini (Yabuuchi et al.) | 1931, 1993 | a. Scabdisease b. severe pulmonary infections c. soft rot | - gladiolus corms - humans - mushroom |
| 13 | B. glumae | Kurita and Tabei (Urakami et al.) | 1967, 1994 | panicle blight | rice |
| 14 | B. humptydooensis | Vanlaere et al. | 2009 | melioidosis disease | humans and animals |
| 15 | B. lata | Vanlaere et al. | 2009 | belongs to B. cepacia complex | |
| 16 | B. latens | Vanlaere et al. | 2008 | belongs to B. cepacia complex | |
| 17 | B. mallei | Zopf (Yabuuchi et al.) | 1885, 1993 | glanders disease | animals |
| 18 | B. metallica | Vanlaere et al. | 2008 | belongs to B. cepacia complex | |
| 19 | B. multivorans | Vandamme et al. | 1997 | belongs to B. cepacia complex | |
| 20 | B. oklahomensis | Glass et al. | 2006 | melioidosis | humans |
| 21 | B. plantarii | Azegami et al. (Urakami et al.) | 1987, 1994 | seedling blight | rice |
| 22 | B. pseudomallei | Whitmore (Yabuuchi et al.) | 1913, 1993 | melioidosis disease | humans and animals |
| 23 | B. pseudomultivorans | Peeters et al. | 2014 | belongs to B. cepacia complex | |
| 24 | B. puraquae | Martina et al. | 2018 | belongs to B. cepacia complex | |
| 25 | B. pyrrocinia | Imanaka et al. (Vandamme et al.), (Storms et al.) | 1965, 1997, 2004 | cystic fibrosis | humans |
| 26 | B. rhizoxinica | Partida-Martinez et al. | 2007 | rice seedling blight, associated with Rhizopus microsporus | rice |
| 27 | B. seminalis | Vanlaere et al. | 2008 | belongs to B. cepacia complex | |
| 28 | B. singaporensis | Wang et al. | 2003 | - | - |
| 29 | B. singularis | Vandamme et al. | 2017 | respiratory system disease | humans |
| 30 | B. stabilis | Vandamme et al. | 2000 | belongs to B. cepacia complex | |
| 31 | B. stagnalis | De Smet et al. | 2015 | B. stagnalis | |
| 32 | B. territorii | De Smet et al. | 2015 | belongs to B. cepacia complex | |
| 33 | B. thailandensis | Brett et al. | 1998 | melioidosis disease | humans and animals |
| 34 | B. ubonensis | Yabuuchi et al. | 2000 | - | - |
| 35 | B. vietnamiensis | Gillis et al. | 1995 | cystic fibrosis | humans |
Table 2.
List of secondary metabolites synthesized by some species of Burkholderia spp.
| No. | Species | Synthesized Metabolites | References |
|---|---|---|---|
| 1 | B. cepacia | Pyrrolnitrin | [26,72] |
| Cepacidine A | [75,76] | ||
| 2 | B. andropogonis | Rhizobitoxine | [81,82] |
| 3 | B. plantarii | Tropolone | [87] |
| 4 | B. plantarii strain DSM 9509 | Rhamnolipids | [88] |
| 5 | B. rhizoxinica | Rhizoxin | [89,90] |
| 6 | B. glumae | Toxoflavin | [97,98] |
| 7 | B. caryophylli | Caryoynencine | [99,101] |
| 8 | B. gladioli pv. cocovenenas | Bongkrekic acid and toxoflavin | [108,116] |
| 9 | B. gladioli pv. agaricicola | d-Limonene | [127] |
| 4-Flavanone | [128] | ||
| Ornithine lipid | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Elshafie, H.S.; Camele, I. An Overview of Metabolic Activity, Beneficial and Pathogenic Aspects of Burkholderia Spp. Metabolites 2021, 11, 321. https://doi.org/10.3390/metabo11050321
AMA Style
Elshafie HS, Camele I. An Overview of Metabolic Activity, Beneficial and Pathogenic Aspects of Burkholderia Spp. Metabolites. 2021; 11(5):321. https://doi.org/10.3390/metabo11050321
Chicago/Turabian StyleElshafie, Hazem S., and Ippolito Camele. 2021. "An Overview of Metabolic Activity, Beneficial and Pathogenic Aspects of Burkholderia Spp." Metabolites 11, no. 5: 321. https://doi.org/10.3390/metabo11050321
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.