

Four New Species of Torula (Torulaceae, Pleosporales) from Sichuan, China


School of Life Science and Technology, Center for Informational Biology, University of Electronic Science and Technology of China, Chengdu 611731, China
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(2), 150; https://doi.org/10.3390/jof9020150
Submission received: 30 December 2022
/
Revised: 18 January 2023
/
Accepted: 19 January 2023
/
Published: 22 January 2023
(This article belongs to the Special Issue Polyphasic Identification of Fungi 2.0)
Abstract
:Torula is an asexual and hyphomycetous genus in the family Torulaceae. Torula species are generally saprophytic. They have a worldwide distribution and abound in humid or freshwater habitats. In order to better understand this genus, we carried out several field collections from Sichuan, China. As a result, we obtained nine Torula isolates from dead woody substrates in terrestrial and freshwater habitats. Based on a biphasic approach of morphological examination and multi-locus phylogenetic analyses (ITS, SSU, LSU, TEF, RPB2), these collections were identified as belonging to seven Torula species. Four of them were new species (Torula chinensis, T. longiconidiophora, T. sichuanensis and T. submersa), and the other three belonged to existing species, though one was found for the first time in China (T. masonii). Morphological and updated phylogenetic delamination of the new discoveries is also discussed. This study provides further insights into our understanding of wood-based Torula species in China.
1. Introduction
The family Torulaceae was introduced by Corda with Torula as the type [1]. Torulaceae is known only by asexual characteristics of its effuse and powdery colony; mostly immersed mycelia; micro- or macronematous, erect and subcylindrical conidiophores; doliiform to ellipsoid or clavate, cupulate, smooth to verruculose and mono- to polyblastic conidiogenous cells; subcylindrical, phragmosporous, acrogenous, brown, and smooth to verrucose conidia [1,2,3,4,5]. Based on combined ITS and LSU sequence data analyses, Crous et al. [1] revised the family Torulaceae and included the genus Dendryphion. Su et al. [2] introduced Neotorula from freshwater habitats, and Li et al. [6] reported a Sporidesmium-like genus, Sporidesmioides, within Torulaceae. Su et al. [5] provided an updated phylogeny for Torulaceae based on the combined ITS, LSU, RPB2, and TEF sequence data analyses and established the monotypic genus Rostriconidium from freshwater habitats. Based on ITS and LSU sequence data and morphology, Crous et al. [7] established the genus Rutola within the Torulaceae. Later, Boonmee et al. [8] introduced Cylindrotorula within the Torulaceae with evidence from morphology and multi-gene phylogenetic analyses (ITS, LSU, RPB2, TEF). At present, the family Torulaceae includes genera Cylindrotorula, Dendryphion, Neotorula, Rostriconidium, Rutola, Sporidesmioides, and Torula [1,2,5,6,7,8].
The genus Torula was initially described by Persoon [9]. Conidiogenesis in Torula was observed by Crane and Schoknecht [10] using light and transmission electron microscopy. The phylogenetic relationships of the Torulaceae (Pleosporales) were studied by Crous et al. [1], and Torula was accepted within the family. Additionally, they identified three new species based on the number of septa per conidium and designated a neotype (CBS H-22275) for the generic type, Torula herbarum [1]. Torula has an ancient origin and has 541 species epithets in Index Fungorum (http://www.indexfungorum.org/; accessed on 20 January 2023). However, most species of Torula were introduced before the arrival of Sanger sequencing; therefore, there are a few molecular data available. Torula species are commonly collected from dead branches and submerged wood in terrestrial or freshwater habitats [4,5]. The genus is characterized by terminal or lateral, monoblastic or polyblastic conidiogenous cells with a basally thickened and heavily melanized wall and a thin-walled apex, frequently collapsing and becoming coronate [1,4,11]. This genus is a mold that lacks a known sexual state. The different species of Torula can be distinguished by conidial size and the number of septa and chains [1,4]. Recently, some species were introduced based on both molecular data and morphology [5,12,13,14].
We are studying the diversity of fungi in southwestern China, Sichuan, along the Yangtze River, and surveying the taxonomy of hyphomycetes [15,16,17,18,19]. During the survey of torula-like species in Chengdu Province (July to September 2021), nine isolates were obtained from wood-based substrates. Based on the in-depth phylogenetic analysis of combined (ITS, LSU, SSU, RPB2 and TEF) sequence data and morphological examination, these isolates were identified as three known Torula species, including a new geographic record (T. masonii) and four new species, viz., T. chinensis sp. nov., T. submersa sp. nov., T. sichuanensis sp. nov. and T. longiconidiophora sp. nov., which are introduced herein.
2. Materials and Methods
2.1. Sample Collection, Isolation and Morphological Examination
A survey on the diversity of hyphomycetes in Sichuan, China, was conducted between July and September of 2021. Dead branches of unknown hosts were collected from 5 locations (Yunqiao Wetland, Chengdu City; Baiyungou, Chongzhou City; Huilonggou, Pengzhou City; Hongfengling, Deyang City; Longchang, Neijiang City) in Sichuan. The specimens were taken into the laboratory in paper envelopes for examination. Microscopic characteristics were observed and recorded using a Nikon SMZ800N stereo microscope equipped with a Nikon DS-Fi3 microscope camera and a Nikon ECLIPSE Ni-U microscope fitted with a Nikon DS-Ri2 microscope camera, respectively. Measurements were conducted using the Nikon NIS-Elements Documentation Imaging Software Version 5.21.00. All photos were processed using Adobe Photoshop software version 22.0. Single conidium isolation was performed following the method described by Senanayake et al. [20]. Germinated conidia were individually transferred to potato dextrose agar (PDA) media plates and incubated in the dark at 25 °C. Culture characteristics were examined and recorded after one week and at regular intervals.
Herbarium specimens were deposited in the Herbarium of Cryptogams Kunming Institute of Botany Academia Sinica (HKAS), Kunming, China; and the Herbarium of the University of Electronic Science and Technology (HUEST), Chengdu, China. The living cultures were deposited in the China General Microbiological Culture Collection Center (CGMCC) in Beijing, China, and the University of Electronic Science and Technology Culture Collection (UESTCC) in Chengdu, China.
2.2. DNA Extraction, PCR Amplification and Sequencing
Fungal genomic DNA was extracted from mycelia using TreliefTM Plant Genomic DNA Kit (TSINGKE Biotech, Shanghai, P.R. China) according to the manufacturer’s protocol. Five partial loci, including the nuclear ribosomal internal transcribed spacer (ITS: ITS1-5.8S-ITS2), the nuclear ribosomal small subunit rRNA gene (SSU), the nuclear ribosomal large subunit rRNA gene (LSU), the translation elongation factor 1-alpha gene (TEF) and the second largest subunit of RNA polymerase II gene (RPB2) were amplified by polymerase chain reaction (PCR). The corresponding primer pairs and PCR conditions are listed in Table 1. The final reaction volume of the PCR reagent was 25 µL containing 2 µL of DNA template, 1 µL each of the forward and reverse primer, 8.5 µL of double-distilled water (ddH2O) and 12.5 µL of 2 × lash PCR MasterMix (mixture of DNA Polymerase, dNTPs, Mg2+ and optimized buffer; CoWin Biosciences, Jiangsu, China). The PCR products were visualized by 1% agarose gel electrophoresis. Sanger sequencing was conducted by Tsingke Biological Technology (Beijing, China).
2.3. Phylogenetic Analyses
According to the corresponding Sanger sequencing chromatograms, misleading data from the ends of raw sequencing fragments were manually trimmed and assembled into consensus sequences using SeqMan Pro version 7.1.0 (DNASTAR, Inc. Madison, WI, USA). Barcode sequences of all Torula species for which the sequence data are available and the outgroup taxon Sporidesmioides thailandica (MFLUCC 13-0840) were downloaded from the NCBI nucleotide database (Table 2) using the function read. GenBank data were integrated within the R package Analysis of Phylogenetics and Evolution (APE) [29].
The multiple sequence alignments were conducted using MAFFT [30] version 7.310 with options “--adjustdirectionaccurately --auto”, and the alignment files were further trimmed using trimAl version 1.4 [31] with the option “-gapthreshold 0.5”, which only allows 50% of taxa with a gap in each site. The best-fit nucleotide substitution models for each locus were selected using PartitionFinder version 2.1.1 [32] under the corrected Akaike information criterion (AICC). All sequence alignments were combined using an in-house Python script.
Maximum likelihood (ML) and Bayesian analysis were conducted based on individual and combined datasets. Five alignment datasets of SSU, ITS, LSU, TEF, and RPB2 were concatenated for multi-locus phylogenetic analysis. ML phylogenetic tree was obtained using the IQ-TREE version 2.0.3 [33], and the topology was evaluated using 1,000 ultrafast bootstrap replicates. The Bayesian analysis was conducted using parallel MrBayes version 3.2.7a [34]. Two different runs with 20 million generations and four chains were executed, and the initial 25% of sample trees were treated as burn-in. Tracer version 1.7.1 [35] was used to confirm that the MCMC runs reached convergence, and all ESS values were above 200. Then, the ML tree was annotated by TreeAnnotator version 2.6.4 implemented in BEAST [36] based on Mrbayes MCMC trees with no discard of burn-in and no posterior probability limit. The ML tree was visualized using ggtree [37] and further edited in Adobe Illustrator software version 16.0.0.
3. Results
3.1. Molecular Phylogeny
The combined dataset includes five loci (LSU: 1–846, ITS: 847–1397, SSU: 1398–2411, TEF: 2412–3278, RPB2: 3279–4161) from 32 strains of Torula and the outgroup taxon Sporidesmioides thailandica (MFLUCC 13-0840). The combined dataset is composed of 970 distinct patterns, 511 parsimony-informative sites, 411 singleton sites and 3239 constant sites. The best-fit evolution models were determined—GTR+I+G for the ITS, LSU and TEF partitions; K80+G for the SSU partition; and SYM+G for the RPB2 partition.
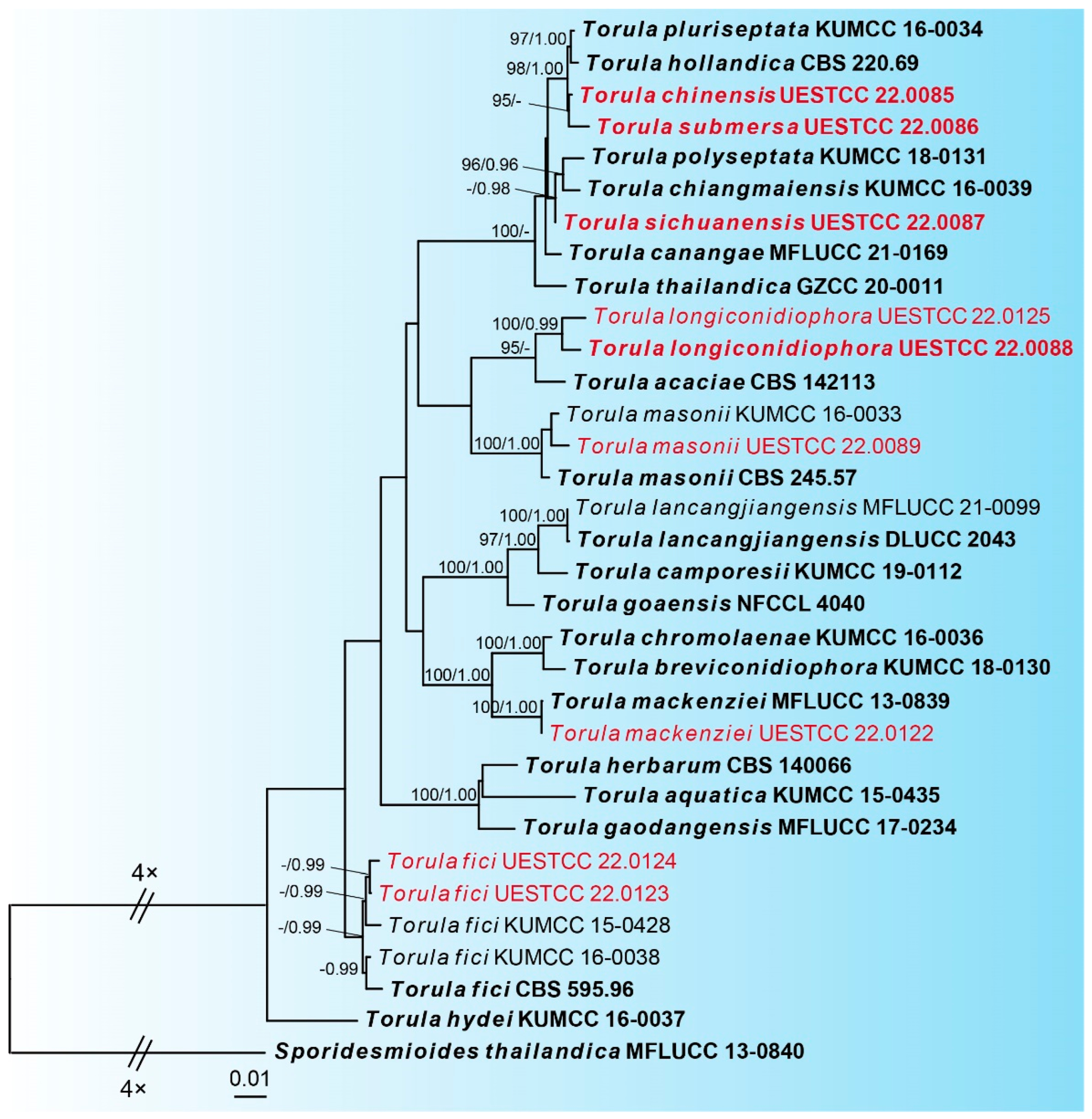
The best-scoring ML tree (lnL = −14437.4141) with ultrafast bootstrap values from ML analyses and posterior probabilities from MrBayes analysis at the node is shown in Figure 1. Nine newly obtained Torula isolates represent seven different species. Torula isolate UESTCC 22.0089 is clustered with the ex-type stain of T. masonii (CBS 245.57). Our Torula isolate UESTCC 22.0122 and the ex-type stain T. mackenziei (MFLUCC 13-0839) are grouped into a statistically well-supported clade (100% ML, 1.00 BYPP). Two Torula isolates (UESTCC 22.0123 and UESTCC 22.0124) are clustered with the three T. fici strains, including the ex-type (CBS 595.96). The new species Torula longiconidiophora (UESTCC 22.0088 and UESTCC 22.0125) is separate from T. acacia (CBS 142113) with 95% ML support. The new species Torula chinensis (UESTCC 22.0085) is a sister to another new species, Torula submersa (UESTCC 22.0086), with 95% ML support. Torula sichuanensis (UESTCC 22.0087) formed a subclade with significant statistical support (91% ML/0.98 BYPP) and is sister to T. polyseptata (KUMCC 18-0131) and T. chiangmaiensis (KUMCC 16-0039).
3.2. Taxonomy
Torula chinensis W.H. Tian, Y.P. Chen and Maharachch., sp. nov. (Figure 2).
MycoBank: MB 847014
Etymology: Named after the country, China, where it was collected.
Saprobic on decaying wood in a damp environment. Asexual morph: Colonies on natural substrate—effuse, black and powdery. Mycelium immersed or superficial, composed of septate, branched, dark brown to black hyphae. Conidiophores 2–4 μm wide, macronematous to semi-macronematous, septate, smooth, straight or slightly flexuous, dark brown, with 1–2 doliiform to globose cells. Conidiogenous cells 5–8 × 5–7 μm (x = 6.1 × 6.2, n = 25), polyblastic, terminal, dark brown to black, doliiform to subglobose, smooth to minutely verruculose and thick-walled. Conidia 6–37 × 4–8 μm (x = 21.5 × 6.0, n = 35), catenated, acrogenous, simple, phragmosporous, dark brown to black, minutely verruculose, 1–7-septate, rounded at both ends, composed of subglobose cells, often smaller at the apex, slightly constricted at some septa and chiefly subcylindrical. Conidial secession schizolytic.
Material examined: CHINA: Sichuan, Chongzhou City, Baiyungou, on a submerged decaying branch of an unknown host, N 30°47’56, E 103°24’15, elevation 990 m, 27 September 2021, W.H. Tian and Y.P. Chen BY45-2 (HKAS 126509, holotype), ex-type culture; CGMCC 3.24282 = UESTCC 22.0085.
Culture characteristics: Conidia germinated on PDA within 24 h at 25 °C. Colonies reached 32 mm after 10 days in an incubator under dark conditions at 25 °C. Colonies were raised in the middle; they were irregular circles; surface velvety; they had a white center and were yellowish-brown at the edges, with clear margins; reverse, yellowish brown with pale edges.
Notes: The phylogenetic tree shows that the isolate UESTCC 22.0085 is clustered with the ex-type strain of Torula submersa (UESTCC 22.0086) (Figure 1). Torula chinensis differs from T. submersa by having longer conidia (6–37 vs. 6–20 μm) and more septa in conidia (1–7 vs. 1–4). Torula chinensis resembles T. pluriseptata by having phragmosporous, minutely verruculose, subglobose cells and catenate conidia, but differs by having bigger conidiogenous cells (5–8 × 5–7 μm vs. 3.2–3.5 × 3.8–4.6) and fewer septa and shorter conidia (1–7-septa conidia 6–37 × 4–8 μm vs. 3–10-septa conidia 23.5–36 × 3.6–4.4 μm) [38]. Thus, considering the significant differences in morphology and molecular data, we describe the isolate UESTCC 22.0085 as T. chinensis sp. nov.
Torula longiconidiophora W.H. Tian, Y.P. Chen and Maharachch., sp. nov. (Figure 3).
MycoBank: MB 847015
Etymology: Referring to the longer conidiophores.
Saprobic on decaying wood in a damp environment. Asexual morph: Colonies effuse on the host, velvety, dark brown to black and powdery. Mycelium partly immersed, composed of septate, branched, smooth, dark brown to black hyphae. Conidiophores 9–166 × 2–6 μm (x = 74.1 × 5.3, n = 20), septate, solitary, straight or curly winded, light brown to dark brown, consisting of many subcylindrical to subglobose cells that are smooth to minutely verruculose, without apical branches and thick-walled, and some doliiform to globose cells, forming scars at the junction with the conidiogenous cells. Conidiogenous cells 6–12 × 6–11 μm (x = 7.2 × 8.3, n = 25), polyblastic, terminal or intercalary, produced branched chain; base cell slightly truncated, black, ellipsoid to coronal, smooth to verruculose and thick-walled. Conidia 12–46 × 6–11 μm (x = 36.6 × 9.4, n = 35), solitary to catenate, acrogenous, simple, phragmosporous, dark brown to black, minutely verruculose, 1–7-septate, rounded at both ends, composed of subglobose cells, wider cells in the middle, often smaller at apex, slightly constricted at some septa and chiefly subcylindrical. Conidial secession schizolytic.
Material examined: CHINA: Sichuan, Pengzhou City, Huilonggou, on decaying branch of unknown host, N 31°11′6, E 103°54′56, elevation 1400 m, 28 July 2021, Y.P. Chen HLG 0728821 (HKAS 126512, holotype), ex-type culture, CGMCC 3.24283 = UESTCC 22.0088.
Culture characteristics: Conidia germinating on PDA within 24 h at 25 °C. Colonies reaching 34 mm after 10 days in an incubator in the dark at 25 °C. Colonies raised in the middle, irregularly circular, surface velvety, with white centers and becoming light brown, with a white and clear margin; reverse, yellow to light brown in the center and white at the margin.
Notes: In an NCBI BLASTn search based on ITS sequences, the closest match of T. longiconidiophora was T. acacia (CBS 142113) with 97% sequence identity; they differ in 538/555 bp (3%) with two gaps [39]. In addition, multi-gene phylogenetic analysis of a combined dataset of the LSU, ITS, SSU, TEF and RPB2 showed that they are close relatives with 95% ML support (Figure 1). Torula longiconidiophora also differs from T. acacia in having significantly wider conidia (6–11 μm vs. 5–6 μm) and fewer septa (1–7 vs. 2–15). Furthermore, the conidial cells of T. longiconidiophora are wider in the middle and narrow at the ends, whereas the cells of T. acacia are more uniform [39]. Torula longiconidiophora differs from other species in the genus by having two types of conidiophores. Thus, considering the significant differences in morphology and molecular data, we describe the isolate UESTCC 22.0088 as Torula longiconidiophora sp. nov.
Torula masonii P.W. Crous, IMA Fungus 6 (1): 192 (2015) (Figure 4).
MycoBank: MB 812806
Saprobic on decaying wood in a damp environment. Asexual morph: Colonies sparse, hairy, velvety, black on the substrate. Mycelium immersed or superficial, composed of septate, branched, dark brown to black hyphae. Conidiophores 2–4 μm wide, macronematous to semi-macronematous, septate, erect, straight or slightly flexuous, dark brown, subcylindrical to subglobose and thick-walled, with 1–2 doliiform to globose cells. Conidiogenous cells 4–7 × 5–8 μm (x = 5.5 × 6.5, n = 25), polyblastic, terminal or intercalary, dark brown to black, doliiform to subglobose, smooth to verruculose and thick-walled. Conidia 10–67 × 5–9 μm (?x = 35.2 × 6.5, n = 35), catenated, acrogenous, simple, phragmosporous, dark brown to black, minutely verruculose, 1–12-septate, rounded at both ends, composed of subglobose cells, often smaller at the apex (1–2 cells black at the apex), slightly constricted at some septa and chiefly subcylindrical. Conidial secession schizolytic.
Material examined: CHINA: Sichuan, Pengzhou City, Huilonggou, on decaying branch of unknown host, N 31°11′6, E 103°54′56, elevation 1400 m, 28 July 2021, Y.P. Chen HLG 072856 (HUEST 22.0090), living culture UESTCC 22.0089.
Culture characteristics: Conidia germinated on PDA within 24 h at 25 °C. The colony reached 21 mm after 10 days in an incubator in the dark at 25 °C. Colonies were white and irregularly circular with a velvety surface, with denser mycelium in the center and becoming sparser towards the edge, with clear margins; in reverse, pale green in the center and becoming white towards the edge.
Notes: Torula masonii was introduced by Crous et al. [1] based on the fungus sporulating in culture, which was collected from Brassica sp. in the UK. In this study, the phylogenetic tree shows that our isolate (UESTCC 22.0089) from decaying wood in a damp environment is clustered with the strain T. masonii (KUMCC 16-0033). In addition, isolate HUEST 22.0089 displays similar morphological characteristics with the type species of T. masonii (CBS 245.57) in colony shape, conidiophores, conidiogenous cells and conidia. We identified the isolate UESTCC 22.0089 as T. masonii, a new geographic record from a humid habitat in China.
Torula sichuanensis W.H. Tian, Y.P. Chen and Maharachch., sp. nov. (Figure 5).
MycoBank: MB 847016
Etymology: Named after the province Sichuan, China, where it was collected.
Saprobic on decaying wood in a damp environment. Asexual morph: Colonies effuse on host, black and powdery. Mycelium partly immersed and composed of septate, branched, dark brown to black hyphae. Conidiophores 2–3 μm wide, macronematous to semi-macronematous, erect, solitary, thick-walled, brown, verruculose, consisting of 1–2 cells or reduced to conidiogenous cells and subcylindrical to subglobose. Conidiogenous cells 5–9 × 4–7 μm (x = 6.5 × 5.3, n = 25), polyblastic, terminal, dark brown to black, doliiform to subglobose, smooth to minutely verruculose and thick-walled. Conidia 7–80 × 5–7 μm (x = 31.3 × 6.3, n = 35), solitary to catenated, acrogenous, simple, phragmosporous, dark brown to black, minutely verruculose, 1–12-septate, rounded at both ends, composed of subglobose cells, often smaller at apex, slightly constricted at some septa and chiefly subcylindrical. Conidial secession schizolytic.
Material examined: CHINA: Sichuan, Deyang City, Yinghua town, Hongfengling, on submerged wood, N 31°20’56, E 103°59’47, elevation 1200 m, 16 October 2021, W.H. Tian and Y.P. Chen HFL16 (HKAS 126511, holotype), ex-type culture CGMCC 3.24284 = UESTCC 22.0087.
Culture characteristics: Conidia germinating on PDA within 24 h at 25 °C. Colonies reached 34 mm after 10 days in an incubator under dark conditions at 25 °C. Colony margins, regular; colonies have white aerial mycelia and a woolly center, with a sparse, thin layer and snowflake margin; reverse, brown in center and white in the margin.
Notes: Morphologically, T. sichuanensis is similar to T. chiangmaiensis and T. pluriseptata. However, they differ in the number of septa (1–12- epta vs. 4–12 septa and 3–10 septa) and the size of conidia (7–80 × 5–7 μm vs. 25.5–70 × 5.6–7.8 μm and 23.5–36 × 3.6–4.4 μm) [4]. The color of the conidia of T. chiangmaiensis is light brown to greyish-brown, T. pluriseptata is dark brown and T. sichuanensis is dark brown to black [4]. Phylogenetic analyses also confirmed they are distinct species (Figure 1). Therefore, we introduce the isolate UESTCC 22.0087 as a new species.
Torula submersa W.H. Tian, Y.P. Chen and Maharachch., sp. nov. (Figure 6).
MycoBank: MB 847013
Etymology: Named referring to the submerged habitat of this fungus.
Saprobic on submerged decaying wood. Asexual morph: Colonies effuse, black, velvety and powdery on the host. Mycelium partly immersed and composed of septate, branched, black hyphae. Conidiophores 1–4 μm wide, macronematous to semi-macronematous, solitary, smooth, erect, thick-walled and dark brown, with one doliiform to globose cell. Conidiogenous cells 5–6 × 5–6 μm (x = 5.6 × 5.5, n = 25), monoblastic, terminal, dark brown to black, paler at apex, doliiform to subglobose, smooth to minutely verruculose and thick-walled. Conidia 6–20 × 4–8 μm (x = 14.5 × 6.0, n = 35), catenated, acrogenous, simple, phragmosporous, black, minutely verruculose, 1–4-septate, rounded at both ends, composed of subglobose cells, often paler at the apex, slightly constricted at some septa and chiefly subcylindrical. Conidial secession schizolytic. Sexual morph: Unknown.
Material examined: CHINA: Sichuan, Chongzhou City, Baiyungou, on a submerged decaying branch of unknown host, N 30°47’56, E 103°24’15, elevation 990 m, 27 September 2021, W.H. Tian and Y.P. Chen BY60-2 (HKAS 126510, holotype), ex-type culture CGMCC 3.24281 = UESTCC 22.0086.
Culture characteristics: Conidia germinated on PDA within 24 h at 25 °C. Colonies reached 26 mm after 10 days in an incubator in the dark at 25 °C. Colonies were raised in the middle, irregularly circular, velvety on the surface and had a white center fading to yellowish brown and white edges; they hadclear margins; reverse, yellowish brown in center with white margin.
Notes: The phylogenetic tree shows that the isolate UESTCC 22.0086 is clustered with the ex-type strain of Torula chinensis (UESTCC 22.0085) (Figure 1). Multigene phylogenetic analysis of a combined dataset of the LSU, ITS, SSU, TEF, and RPB2 showed that T. chinensis and T. submersa are close relatives with 95% ML support (Figure 1). Morphologically, T. submersa differs from other Torula species by having black conidia composed of subglobose cells and 1–4 septa (see notes under T. chinensis). Therefore, we introduce the isolate UESTCC 22.0086 as a new species.
4. Discussion
Even though over 541 epithets of Torula have been reported, previously, many Torula species were identified based on morphology alone, and there were very little data relating to the phylogenetic relationships until the study of Crous et al. [1]. Therefore, it is possible that a number of the species in Torula are conspecific or belong to different genera. Re-examining type specimens of Torula-like species described prior to the advent of molecular technology is necessary to address this issue. Additionally, fresh specimens should be collected, sequenced and combined with multi-locus phylogenetic analysis and morphological examination, and designation of epi-types is essential. For instance, Crane et al. [11] re-examined several hyphomycetous species previously placed in the Torula and found that T. rhombica and T. terrestris do not seem to be congenerous with T. herbarum, and transferred to them Bahusandhika. Recently, new species have been identified based on molecular data and morphology, and to date, only 23 species have sequence data [1,3,4,5,8,11,12,13,14,20,38,39,40].
The genus Torula contains many diverse species, frequently isolated from submerged decaying wood, living leaves, dead wood and twigs of various terrestrial plants, soil and earthworm casts [41]. They are commonly isolated as saprobes in temperate and tropical climate regions [42]. A few species are plant pathogens, such as T. herbarum, causing stem blight in Indian jujube (Ziziphus mauritiana) [43]. They are also isolated from the air of natural environments, parks and industrial zones and are reported to cause seasonal fungal allergies in humans [44,45,46,47,48]. The genus has been shown to produce a wide range of chemically novel diverse secondary metabolites. For example, T. herbarum displays antibacterial, antifungal, antiamoebic and potentially anti-cancer properties [49,50,51]. Therefore, more taxonomic, phylogenic and biochemical studies of this bioprospecting genus should be performed.
Author Contributions
Conceptualization, W.T., Y.C. and S.S.N.M.; methodology, W.T. and Y.C.; writing—original draft preparation, W.T.; writing—review and editing, S.S.N.M.; visualization, W.T., Y.C. and P.S.; supervision, S.S.N.M.; funding acquisition, S.S.N.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Talent Introduction and Cultivation Project, University of Electronic Science and Technology of China, grant number A1098531023601245.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequence data are available in NCBI GenBank with the accession numbers in the manuscript.
Acknowledgments
We thank the University of Electronic Science and Technology of China for funding this research. We thank D. Wanasinghe for careful reading of the manuscript and for valuable feedback.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Crous, P.W.; Carris, L.M.; Giraldo, A.; Groenewald, J.Z.; Hawksworth, D.L.; Hemández-Restrepo, M.; Jaklitsch, W.M.; Lebrun, M.H.; Schumacher, R.K.; Stielow, J.B.; et al. The Genera of Fungi—Fixing the application of the type species of generic names—G2: Allantophomopsis, Latorua, Macrodiplodiopsis, Macrohilum, Milospium, Protostegia, Pyricularia, Robillarda, Rotula, Septoriella, Torula, and Wojnowicia. IMA Fungus 2015, 6, 163–198. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Ariyawansa, H.A.; Luo, Z.; Promputtha, I.; Tian, Q.; Lin, C.; Shang, Q.; Zhao, Y.; et al. The families Distoseptisporaceae fam. nov., Kirschsteiniotheliaceae, Sporormiaceae and Torulaceae, with new species from freshwater in Yunnan Province, China. Fungal Divers. 2016, 80, 375–409. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Li, J.F.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Mapook, A.; Camporesi, E.; Shang, Q.J.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. Molecular taxonomy and morphological characterization reveal new species and new host records of Torula species (Torulaceae, Pleosporales). Mycol. Prog. 2017, 16, 447–461. [Google Scholar] [CrossRef]
- Su, X.J.; Luo, Z.L.; Jeewon, R.; Bhat, D.J.; Bao, D.F.; Li, W.L.; Hao, Y.E.; Su, H.Y.; Hyde, K.D. Morphology and multigene phylogeny reveal new genus and species of Torulaceae from freshwater habitats in northwestern Yunnan, China. Mycol. Prog. 2018, 17, 531–545. [Google Scholar] [CrossRef]
- Li, J.; Bhat, D.J.; Phookamsak, R.; Mapook, A.; Lumyong, S.; Hyde, K. Sporidesmioides thailandica gen. et sp. nov. (Dothideomycetes) from northern Thailand. Mycol. Prog. 2016, 15, 1169–1178. [Google Scholar] [CrossRef]
- Crous, P.W.; Schumacher, R.K.; Wood, A.R.; Groenewald, J.Z. The Genera of Fungi—G5: Arthrinium, Ceratosphaeria, Dimerosporiopsis, Hormodochis, Lecanostictopsis, Lembosina, Neomelanconium, Phragmotrichum, Pseudomelanconium, Rutola, and Trullula. Fungal Syst. Evol. 2020, 5, 77–98. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, S.K.U.; Jones, G.E.B.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387–1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–335. [Google Scholar] [CrossRef] [PubMed]
- Persoon, C.H. Neuer versuch einer systematischen eintheilung der schwaümme. N. Mag. Bot. Ihrem Ganzen Umfange 1794, 1, 63–128. [Google Scholar]
- Crane, J.L.; Schoknecht, J.D. Revision of Torula species. Rutola, a new genus for Torula graminis. Botany 1977, 55, 3013–3019. [Google Scholar] [CrossRef]
- Crane, J.L.; Miller, A.N. Studies in genera similar to Torula: Bahusaganda, Bahusandhika, Pseudotorula, and Simmonsiella gen. nov. IMA Fungus 2016, 7, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Thilini Chethana, K.W.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.Q.; Wang, B.; Kang, J.C. Several rare entopathogenic fungi from the Western Sichuan mountains. Fungal Divers. 2003, 12, 129–134. [Google Scholar]
- Zhou, Y.; He, S.; Gong, G.; Zhang, S.; Chang, X.; Liu, N.; Sun, X.; Qi, X.; Ye, K.; Wang, Y. Soil fungal diversity in three nature reserves of Jiuzhaigou County, Sichuan Province, China. Ann. Microbiol. 2014, 64, 1275–1290. [Google Scholar] [CrossRef]
- Zhou, W.; Wei, Q.; Feng, R.; Liu, Y.; Liang, H.; Li, J.; Yan, K. Diversity and spatial distribution of endophytic fungi in Cinnamomum longepaniculatum of Yibin, China. Arch. Microbiol. 2021, 203, 3361–3372. [Google Scholar] [CrossRef]
- Yan, K.; Abbas, M.; Meng, L.; Cai, H.; Peng, Z.; Li, Q.; El-Sappah, A.H.; Yan, L.; Zhao, X. Analysis of the fungal diversity and community structure in Sichuan dark tea during pile-fermentation. Front. Microbiol. 2021, 12, 706714. [Google Scholar] [CrossRef]
- Chen, Y.P.; Tian, W.H.; Guo, Y.B.; Madrid, H.; Maharachchikumbura, S.S.N. Synhelminthosporium gen. et sp. nov. and two new species of Helminthosporium (Massarinaceae, Pleosporales) from Sichuan Province, China. J. Fungi 2022, 8, 712. [Google Scholar] [CrossRef]
- Senanayake, I.; Rathnayaka, A.; Sandamali, D.; Calabon, M.; Gentekaki, E.; Lee, H.; Pem, D.; Dissanayake, L.; Wijesinghe, S.; Bundhun, D.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-coverage ITS primers for the DNA-based identification of Ascomycetes and Basidiomycetes in environmental samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38—Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Cubeta, M.A.; Echandi, E.; Abernethy, T.; Vilgalys, R. Characterization of anastomosis groups of binucleate Rhizoctonia species using restriction analysis of an amplified ribosomal RNA gene. Phytopathology 1991, 81, 1395–1400. [Google Scholar] [CrossRef]
- Hibbett, D.S. Phylogenetic evidence for horizontal transmission of group I introns in the nuclear ribosomal DNA of mushroom-forming fungi. Mol. Biol. Evol. 1996, 13, 903–917. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-alpha sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Komon, M.; Kubicek, C.P.; Druzhinina, I.S. Hypocrea voglmayrii sp. nov. from the Austrian Alps represents a new phylogenetic clade in Hypocrea/Trichoderma. Mycologia 2005, 97, 1365–1378. [Google Scholar] [CrossRef]
- Voglmayr, H.; Akulov, O.Y.; Jaklitsch, W.M. Reassessment of Allantonectria, phylogenetic position of Thyronectroidea, and Thyronectria caraganae sp. nov. Mycol. Prog. 2016, 15, 921–937. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for selecting partitioned models of evolution for molecular and morphological phylogenetic Analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Yu, G. Using ggtree to visualize data on Tree-like structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef]
- Pratibha, J.; Prabhugaonkar, A. Torula goaensis, a new asexual ascomycetous fungus in Torulaceae. Webbia 2017, 72, 171–175. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; Le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal Planet description sheets: 469–557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef]
- De Silva, N.I.; Hyde, K.D.; Lumyong, S.; Phillips, A.J.L.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Thambugala, K.M.; Tennakoon, D.S.; Suwannarach, N.; Karunarathna, S.C. Morphology, phylogeny, host association and geography of fungi associated with plants of Annonaceae, Apocynaceae and Magnoliaceae. Mycosphere 2022, 13, 955–1076. [Google Scholar] [CrossRef]
- Tiwari, S.C.; Mishra, R.R. Fungal abundance and diversity in earthworm casts and in uningested soil. Biol. Fertil. Soils 1993, 16, 131–134. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 105, 317–318. [Google Scholar] [CrossRef]
- Nallathambi, P.; Umamaheswari, C. A new disease of ber (Ziziphus mauritiana Lim) caused by Torula herbarum (Pers) link. J. Mycol. Plant Pathol. 2001, 31, 92. [Google Scholar]
- Kasprzyk, I.; Grinn-Gofroń, A.; Ćwik, A.; Kluska, K.; Cariñanos, P.; Wójcik, T. Allergenic fungal spores in the air of urban parks. Aerobiologia 2021, 37, 39–51. [Google Scholar] [CrossRef]
- Knutsen, A.P.; Bush, R.K.; Demain, J.G.; Denning, D.W.; Dixit, A.; Fairs, A.; Greenberger, P.A.; Kariuki, B.; Kita, H.; Kurup, V.P.; et al. Fungi and allergic lower respiratory tract diseases. J. Allergy. Clin. Immunol. 2012, 129, 280–291. [Google Scholar] [CrossRef]
- Korneykova, M.V.; Soshina, A.S.; Gavrichkova, O.V. Culturable airborne fungi of urban, forest and coastal areas of the Kola Peninsula. In Proceedings of the Advanced Technologies for Sustainable Development of Urban Green Infrastructure; Springer: Cham, Switzerland, 2021; pp. 150–160. [Google Scholar]
- Korneykova, M.V.; Evdokimova, G.A. Microbiota of the ground air layers in natural and industrial zones of the Kola Arctic. J. Environ. Sci. Health. A Tox. Hazard. Subst. Environ. Eng. 2018, 53, 271–277. [Google Scholar] [CrossRef]
- Anjali, A.R.; Bhajbhuje, M.N. Aeromycoflora of the lake of Futala area, Nagpur (M. S.) India. Int. J. Life Sci. 2018, 6, 635–642. [Google Scholar]
- Kadkol, M.V.; Gopalkrishnan, K.S.; Narasimhachari, N. Isolation and characterization of naphthaquinone pigments from Torula herbarum (Pers.). herbarin and dehydroherbarin. J. Antibiot. 1971, 24, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Cole, R.J.; Jarvis, B.B.; Schweikert, M.A. 7—Paspaline and Related Metabolites. In Handbook of Secondary Fungal Metabolites; Cole, R.J., Jarvis, B.B., Schweikert, M.A., Eds.; Academic Press: San Diego, CA, USA, 2003; pp. 441–482. [Google Scholar]
- Narasimhachari, N.; Gopalkrishnan, L.S. Naphthaquinone pigments from Torula herbarum: Structure of o-methylherbarin. J. Antibiot. 1974, 27, 283–287. [Google Scholar] [CrossRef]
Figure 1.
Phylogram of the best ML tree of species of Torula based on a combined dataset of LSU, ITS, SSU, TEF, and RPB2. Novel isolates are indicated in red. Type isolates are in bold. The ML ultrafast bootstrap values/Bayesian PP greater than 95%/0.95 are indicated at the respective nodes. The tree is rooted with Sporidesmioides thailandica (MFLUCC 13-0840).
Figure 1.
Phylogram of the best ML tree of species of Torula based on a combined dataset of LSU, ITS, SSU, TEF, and RPB2. Novel isolates are indicated in red. Type isolates are in bold. The ML ultrafast bootstrap values/Bayesian PP greater than 95%/0.95 are indicated at the respective nodes. The tree is rooted with Sporidesmioides thailandica (MFLUCC 13-0840).

Figure 2.
Torula chinensis (HKAS 126509, holotype). (a–c) Colonies on the natural substrate, arrows indicate the locations of colonies. (d–h) Conidiophores, conidiogenous cells and conidia. (i–p) Conidia. (q,r) Conidia in catenated chain. (s) Germinated conidium. (t,u) Culture on PDA from the surface and reverse. Scale bars: (d–s) = 10 μm. Scale bar of (d) applies to (d–h). Scale bar of (i) applies to (i–n). Scale bar of (o) applies to (o–r).
Figure 2.
Torula chinensis (HKAS 126509, holotype). (a–c) Colonies on the natural substrate, arrows indicate the locations of colonies. (d–h) Conidiophores, conidiogenous cells and conidia. (i–p) Conidia. (q,r) Conidia in catenated chain. (s) Germinated conidium. (t,u) Culture on PDA from the surface and reverse. Scale bars: (d–s) = 10 μm. Scale bar of (d) applies to (d–h). Scale bar of (i) applies to (i–n). Scale bar of (o) applies to (o–r).

Figure 3.
Torula longiconidiophora (HKAS 126512, holotype). (a–c) Colonies on the natural substrate; arrows indicate the locations of colonies. (d–f) Conidiophores with conidiogenous cells. (g) Mass of conidiophores. (h–j) Conidiophores. (k) Conidiogenous cells. (l–p) Conidia. (q,r) Culture on PDA from the surface and reverse. Scale bars: (d–p) = 10 μm. Scale bar of (d) applies to (d–f). Scale bar of (h) applies to (h–j). Scale bar of (l) applies to (l–p).
Figure 3.
Torula longiconidiophora (HKAS 126512, holotype). (a–c) Colonies on the natural substrate; arrows indicate the locations of colonies. (d–f) Conidiophores with conidiogenous cells. (g) Mass of conidiophores. (h–j) Conidiophores. (k) Conidiogenous cells. (l–p) Conidia. (q,r) Culture on PDA from the surface and reverse. Scale bars: (d–p) = 10 μm. Scale bar of (d) applies to (d–f). Scale bar of (h) applies to (h–j). Scale bar of (l) applies to (l–p).

Figure 4.
Torula masonii (HUEST 22.0090). (a–c) Colonies on the natural substrate. (d–g) Conidiophores, conidiogenous cells and conidia. (h–q) Conidia; (r) Germinated conidium. (s,t) Culture on PDA from surface and reverse. Scale bars: (d–r) = 10 μm. Scale bar of (d) applies to (d–g). Scale bar of (h) applies to (h–m). Scale bar of (n) applies to (n–q).
Figure 4.
Torula masonii (HUEST 22.0090). (a–c) Colonies on the natural substrate. (d–g) Conidiophores, conidiogenous cells and conidia. (h–q) Conidia; (r) Germinated conidium. (s,t) Culture on PDA from surface and reverse. Scale bars: (d–r) = 10 μm. Scale bar of (d) applies to (d–g). Scale bar of (h) applies to (h–m). Scale bar of (n) applies to (n–q).

Figure 5.
Torula sichuanensis (HKAS 126511, holotype). (a–c) Colonies on the natural substrate; arrows indicate the locations of colonies. (d,e) Conidiophores, conidiogenous cells and conidia. (f) Squash mount of partial colony. (g–q) Conidia; (r) Germinated conidium; (s,t) Culture on PDA from surface and reverse. Scale bars: (d–r) = 10 μm. Scale bar of (g) applies to (g–q).
Figure 5.
Torula sichuanensis (HKAS 126511, holotype). (a–c) Colonies on the natural substrate; arrows indicate the locations of colonies. (d,e) Conidiophores, conidiogenous cells and conidia. (f) Squash mount of partial colony. (g–q) Conidia; (r) Germinated conidium; (s,t) Culture on PDA from surface and reverse. Scale bars: (d–r) = 10 μm. Scale bar of (g) applies to (g–q).

Figure 6.
Torula submersa (HKAS 126510, holotype). (a–c) Colonies on the natural substrate; (d–g) Conidiophores with conidiogenous cells. (h,i) Branched chains of conidia. (j–q) Conidia. (r) Germinated conidium. (s,t) Culture on PDA from the surface and reverse. Scale bars: (d–r) = 10 μm. Scale bar of (d) applies to (d–g). Scale bar of (h) applies to (h,i). Scale bar of (j) applies to (j–q).
Figure 6.
Torula submersa (HKAS 126510, holotype). (a–c) Colonies on the natural substrate; (d–g) Conidiophores with conidiogenous cells. (h,i) Branched chains of conidia. (j–q) Conidia. (r) Germinated conidium. (s,t) Culture on PDA from the surface and reverse. Scale bars: (d–r) = 10 μm. Scale bar of (d) applies to (d–g). Scale bar of (h) applies to (h,i). Scale bar of (j) applies to (j–q).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Loci used in this study with the corresponding PCR primers and conditions.
| Locus | PCR Primers | PCR: Thermal Cycles | References |
|---|---|---|---|
| ITS | ITS9mun or ITS5/ITS4_KYO1 or ITS4 | (94 °C: 30 s, 56 °C: 30 s, 72 °C: 30 s) × 35 cycles | [21,22] |
| LSU | LR0R/LR5 | (94 °C: 30 s, 56 °C: 30 s, 72 °C: 1 min) × 35 cycles | [23,24] |
| SSU | PNS1/NS41 | (94 °C: 30 s, 56 °C: 30 s, 72 °C: 1 min) × 35 cycles | [25] |
| TEF | EF1-983/EF1-2218R | (94 °C: 30 s, 52 °C: 30 s, 72 °C: 1 min) × 35 cycles | [26,27] |
| RPB2 | dRPB2-5f/dRPB2-7r | (94 °C: 30 s, 52 °C: 30 s, 72 °C: 1 min) × 35 cycles | [28] |
Table 2.
Species details and their GenBank accession numbers used in phylogenetic analyses. Type/epitype strains are in bold, and newly generated sequences are in red.
Table 2.
Species details and their GenBank accession numbers used in phylogenetic analyses. Type/epitype strains are in bold, and newly generated sequences are in red.
| Species | Strain/Voucher No. | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| ITS | LSU | SSU | TEF | RPB2 | ||
| Sporidesmioides thailandica | MFLUCC 13-0840 | MN061347 | NG_059703 | NG_061242 | KX437766 | KX437761 |
| Torula acaciae | CBS 142113 | NR_155944 | NG_059764 | - | - | KY173594 |
| T. aquatica | KUMCC 15-0435 | MG208167 | MG208146 | - | - | MG207977 |
| T. breviconidiophora | KUMCC 18-0130 | MK071670 | MK071672 | MK071697 | MK077673 | - |
| T. camporesii | KUMCC 19-0112 | MN507400 | MN507402 | MN507401 | MN507403 | MN507404 |
| T. canangae | MFLUCC 21-0169 | OL966950 | OL830816 | - | ON032379 | - |
| T. chiangmaiensis | KUMCC 16-0039 | MN061342 | KY197856 | KY197863 | KY197876 | - |
| T. chinensis | UESTCC 22.0085 | OQ127986 | OQ128004 | OQ127995 | - | - |
| T. chromolaenae | KUMCC 16-0036 | MN061345 | KY197860 | KY197867 | KY197880 | KY197873 |
| T. fici | CBS 595.96 | KF443408 | KF443385 | KF443387 | KF443402 | KF443395 |
| T. fici | KUMCC 15-0428 | MG208172 | MG208151 | - | MG207999 | MG207981 |
| T. fici | KUMCC 16-0038 | MN061341 | KY197859 | KY197866 | KY197879 | KY197872 |
| T. fici | UESTCC 22.0123 | OQ127978 | OQ127996 | OQ127987 | OQ158972 | OQ158969 |
| T. fici | UESTCC 22.0124 | OQ127979 | OQ127997 | OQ127988 | OQ158972 | OQ158970 |
| T. gaodangensis | MFLUCC 17-0234 | MF034135 | NG_059827 | NG_063641 | - | - |
| T. goaensis | NFCCL 4040 | NR_159045 | NG_060016 | - | - | - |
| T. herbarum | CBS 140066 | KR873260 | KR873288 | - | - | - |
| T. hollandica | CBS 220.69 | NR_132893 | NG_064274 | KF443389 | KF443401 | KF443393 |
| T. hydei | KUMCC 16-0037 | MN061346 | MH253926 | MH253928 | MH253930 | - |
| T. lancangjiangensis | DLUCC 2043 | MW723059 | MW879526 | MW774582 | MW729785 | MW729780 |
| T. lancangjiangensis | MFLUCC 21-0099 | MZ538529 | MZ538563 | - | MZ567104 | - |
| T. longiconidiophora | UESTCC 22.0088 | OQ127983 | OQ128001 | OQ127992 | OQ158972 | OQ158967 |
| T.longiconidiophora | UESTCC 22.0125 | OQ127984 | OQ128002 | OQ127993 | OQ158972 | OQ158972 |
| T. mackenziei | MFLUCC 13-0839 | MN061344 | KY197861 | KY197868 | KY197881 | KY197874 |
| T. mackenziei | UESTCC 22.0122 | OQ127980 | OQ127998 | OQ127989 | OQ158972 | OQ158971 |
| T. masonii | CBS 245.57 | NR_145193 | NG_058185 | - | - | - |
| T. masonii | KUMCC 16-0033 | MN061339 | KY197857 | KY197864 | KY197877 | KY197870 |
| T. masonii | UESTCC 22.0089 | OQ127982 | OQ128000 | OQ127991 | - | - |
| T. pluriseptata | KUMCC 16-0034 | MN061338 | KY197855 | KY197862 | KY197875 | KY197869 |
| T. polyseptata | KUMCC 18-0131 | MK071671 | MK071673 | MK071698 | MK077674 | - |
| T. sichuanensis | UESTCC 22.0087 | OQ127981 | OQ127999 | OQ127990 | - | - |
| T. submersa | UESTCC 22.0086 | OQ127985 | OQ128003 | OQ127994 | OQ158972 | OQ158968 |
| T. thailandica | GZCC 20-0011 | MN907426 | MN907428 | MN907427 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tian, W.; Su, P.; Chen, Y.; Maharachchikumbura, S.S.N. Four New Species of Torula (Torulaceae, Pleosporales) from Sichuan, China. J. Fungi 2023, 9, 150. https://doi.org/10.3390/jof9020150
AMA Style
Tian W, Su P, Chen Y, Maharachchikumbura SSN. Four New Species of Torula (Torulaceae, Pleosporales) from Sichuan, China. Journal of Fungi. 2023; 9(2):150. https://doi.org/10.3390/jof9020150
Chicago/Turabian StyleTian, Wenhui, Pengwei Su, Yanpeng Chen, and Sajeewa S. N. Maharachchikumbura. 2023. "Four New Species of Torula (Torulaceae, Pleosporales) from Sichuan, China" Journal of Fungi 9, no. 2: 150. https://doi.org/10.3390/jof9020150
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.