Quick-Freeze, Deep-Etch Electron Microscopy Reveals the Characteristic Architecture of the Fission Yeast Spore



1
Department of Biology, Graduate School of Science, Osaka City University, Sumiyoshi-ku, Osaka 558-8585, Japan
2
The OCU Advanced Research Institute for Natural Science and Technology (OCARINA), Osaka City University, Sumiyoshi-ku, Osaka 558-8585, Japan
*
Authors to whom correspondence should be addressed.
J. Fungi 2021, 7(1), 7; https://doi.org/10.3390/jof7010007
Submission received: 30 November 2020
/
Revised: 22 December 2020
/
Accepted: 23 December 2020
/
Published: 26 December 2020
(This article belongs to the Special Issue Formation and Function of Fungal Ascospores)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The spore of the fission yeast Schizosaccharomyces pombe is a dormant cell that is resistant to a variety of environmental stresses. The S. pombe spore is coated by a proteinaceous surface layer, termed the Isp3 layer because it comprises mainly Isp3 protein. Although thin-section electron microscopy and scanning electron microscopy have revealed the fundamental structure of the spore, its architecture remains unclear. Here we visualized S. pombe spores by using a quick-freeze replica electron microscopy (QFDE-EM) at nanometer resolution, which revealed novel characteristic structures. QFDE-EM revealed that the Isp3 layer exists as an interwoven fibrillar layer. On the spore cell membrane, many deep invaginations, which are longer than those on the vegetative cell membrane, are aligned in parallel. We also observed that during spore germination, the cell surface changes from a smooth to a dendritic filamentous structure, the latter being characteristic of vegetative cells. These findings provide significant insight into not only the structural composition of the spore, but also the mechanism underlying the stress response of the cell.
1. Introduction
The spore of the fission yeast Schizosaccharomyces pombe is a quiescent cell that is highly resistant to various stresses, including heat, digestive enzymes, and organic solvents [1,2]. This resistance is associated with various structural features of the spore. Structural characterization of S. pombe spore has been performed mainly by thin-section electron microscopy [3,4,5], which has revealed that the spore wall is more extensive than the vegetative cell wall. The spore wall is also thicker than the vegetative cell wall, and its constituents differ substantially. In the vegetative cell wall, α- and β-glucans and galactomannan are the major components [6,7,8]. By contrast, the spore wall comprises chitosan, in addition to mannan and glucans, and is coated by a proteinaceous layer comprising mainly the Isp3 protein. Since spore mutants with defects in the genes responsible for spore wall formation exhibit sensitivity to stress, the spore wall seems to primarily contribute to the stress resistance of spores [9,10,11,12,13,14,15].
Scanning electron microscopy (SEM) has shown that the S. pombe spore has many protrusions projecting outwards [16]. In terms of probing spore structure, SEM provides three-dimensional images, but the chemical fixation needed for samples may change the original structure, and the resolution of SEM is lower than that of transmission microscopy. On the other hand, thin-section microscopy does not produce three-dimensional images.
Quick-freeze, deep-etch replica EM (QFDE-EM) has emerged as a useful tool that can be applied to the visualization of many biological phenomena [17,18]. It is an advanced technology that is used to capture a snapshot of a biological specimen in an active state, with a spatial resolution on the order of nanometers and a time resolution of sub-milliseconds. In this method, the specimen is frozen in less than a millisecond by pressing it against a metal block chilled with liquid helium or nitrogen, resulting in much faster fixation than can be achieved by chemical methods [18]. The frozen specimen is exposed by fracturing and etching, and is then shadowed by platinum. The resulting metal replica provides high contrast, allowing clear visualization from a single image without the need for image averaging, unlike cryo-electron microscopy [17,18]. Therefore, QFDE-EM has considerable advantages, especially when used to visualize low-density images, relative to other methods of transmission electron microscopy.
In this study, we observed S. pombe spores by QFDE-EM at nanometer resolution, enabling us to identify some novel structural features: namely an interwoven fibrillar layer on the spore surface and deep invaginations of the spore cell membrane. We also used QFDE-EM to visualize the germination process.
2. Materials and Methods
2.1. Yeast Strains, Media, and Culture Conditions
2.2. Isolation of Spores by Density Gradient Centrifugation
The homothallic haploid strains were grown in ME liquid medium for 7 days. The ascal walls were spontaneously dissolved to liberate single spores. The spores were isolated by using linear 25–55% Urografin (Bayer, Leverkusen, Germany) density-gradient centrifugation as described previously [22].
2.3. Quick-Freeze Deep-Etch Electron Microscopy
The isolated spores were washed twice with distilled water, and an aliquot was mixed with a slurry including mica flakes and frozen by a CryoPress (Valiant Instruments, St. Louis, MO, USA) cooled by liquid helium. To prepare a replica, the frozen sample was knife-fractured, and subjected to platinum and carbon rotary-shadowing in a JFDV freeze-etching device (JEOL, Akishima, Japan). The platinum replica was removed by floatation on the surface of hydrofluoric acid, cleaned in commercial bleach, rinsed in distilled water, and transferred to a 400-mesh copper grid as previously described [23,24]. Replicas were viewed under a transmission electron microscope (JEM1010, JEOL) at an acceleration voltage of 80 kV. Electron microscopy images were obtained with a FastScan-F214 (T) charge-coupled device (CCD) camera (TVIPS, Gauting, Germany).
3. Results
3.1. The S. pombe Spore Is Covered by a Fibrillar Structure of Isp3
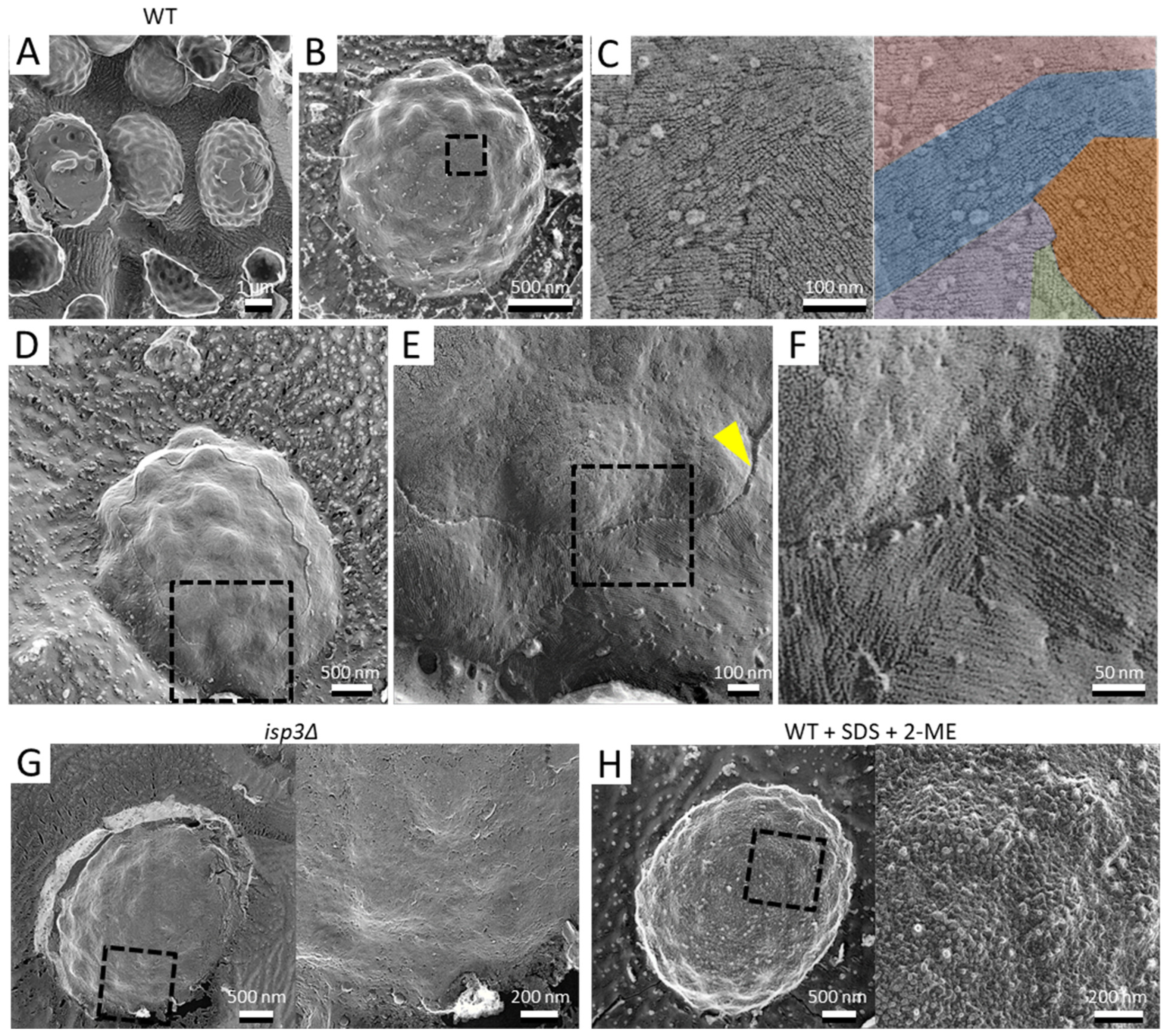
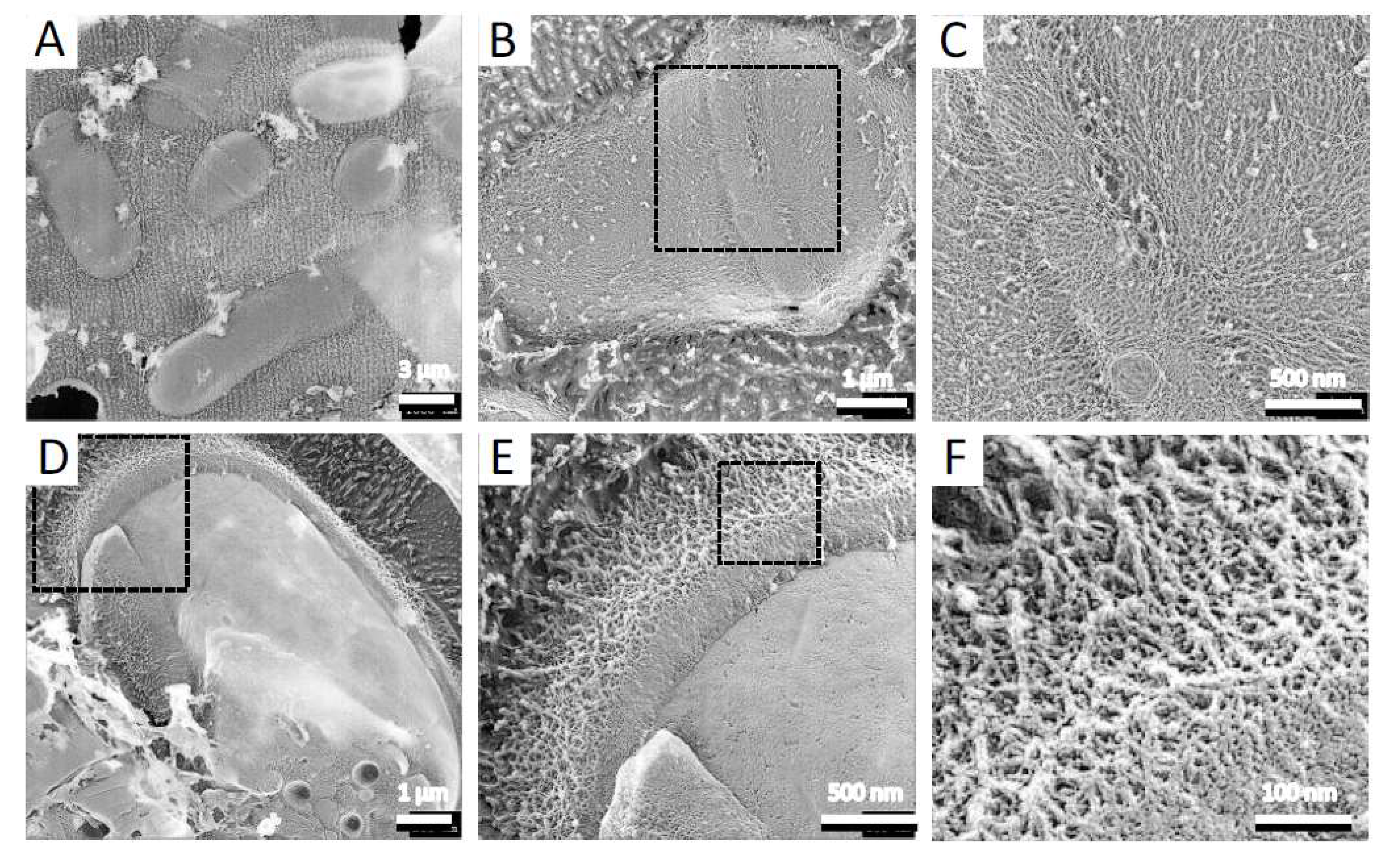
To observe the architecture of S. pombe ascospores in more detail, we conducted QFDE-EM. Isolated spores (see Materials and Methods) were collected by centrifugation and then mixed with mica flakes. The spores were frozen, fractured, deeply etched, and shadowed with platinum, and then the platinum replicas were recovered and observed (Figure 1). The shape and dimensions of the spores were consistent with those observed in images obtained by SEM [16], and each spore had many protrusions that projected outwards. Whereas dendritic structures covered the surface of vegetative cells (Figure 2), the surface of the spore was essentially smooth (Figure 1A,B).
Next, we observed the spore surface at a higher resolution, which revealed fibrillar structures with a width of about 7.0 ± 1.9 nm, a length of hundreds of nanometers, and grouped fascicles (Figure 1B,C). The thickness of the layer was about 12.0 ± 3.0 nm (Figure 1D–F). Previous studies have shown that the S. pombe spore is covered by a proteinaceous layer, termed the “Isp3 layer”, which comprises mainly Isp3 protein [9,15]. To determine whether the fibrillar structure corresponds to the Isp3 layer, we examined the surface of isp3∆ spores. QFDE-EM showed that there were no fibrillar structures on the isp3∆ spore (Figure 1G).
It is known that Isp3 is firmly associated with the spore wall: extraction of Isp3 from the spore wall requires treatment with both 1% SDS and 5% β-mercaptoethanol, and neither treatment works alone [9,15]. As expected, the fibrillar layer was not observed on spores pretreated with both SDS and β-mercaptoethanol (Figure 1H), similar to isp3∆ spores. Collectively, these data indicate that the Isp3 layer is present as a fibrillar structure on the spore surface.
3.2. Deep Invaginations Are Aligned in Parallel on the Spore Cell Membrane
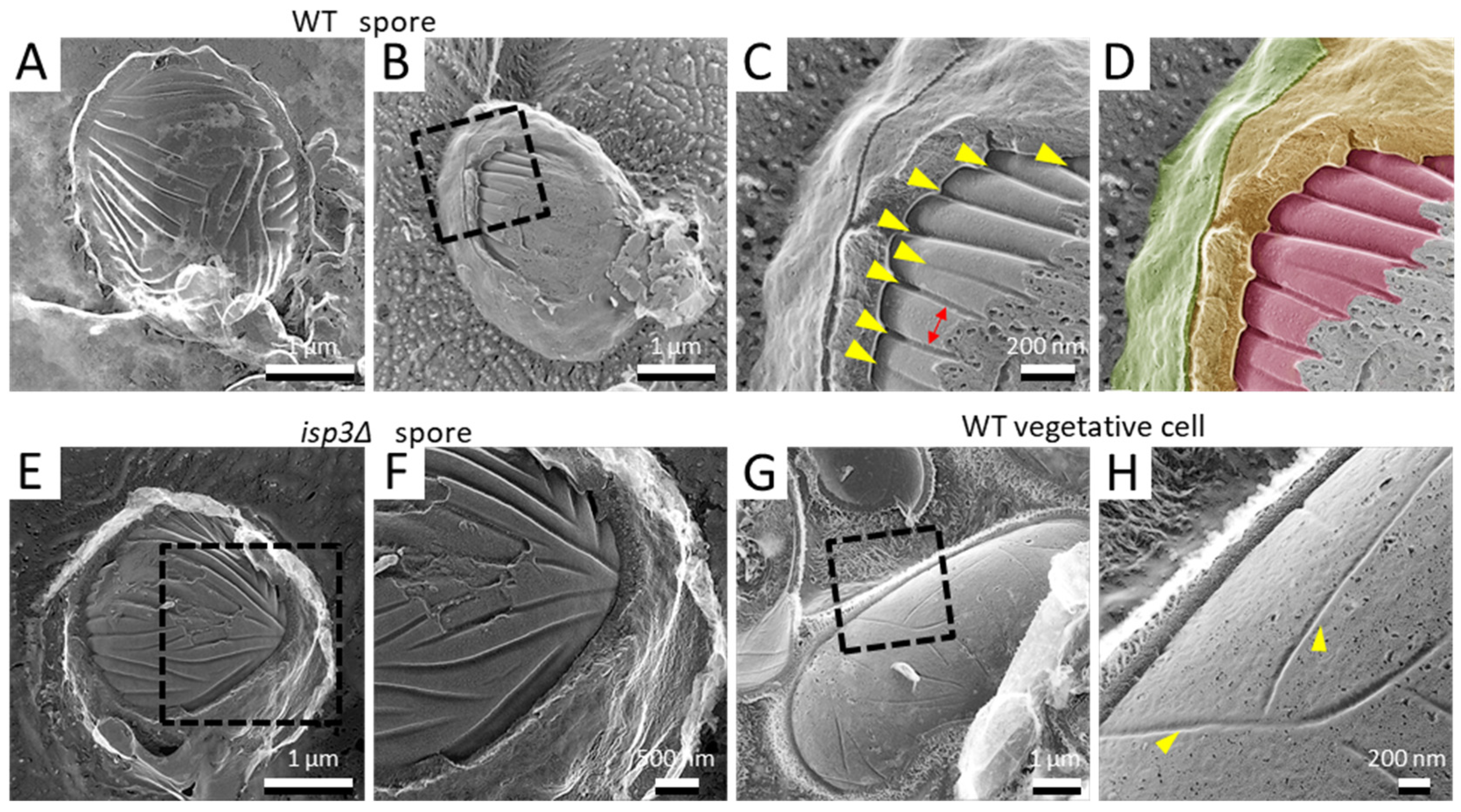
In more deeply fractured samples, we observed spores with the cell wall removed or with the cell membrane exposed (Figure 3A). As described previously, there are many invaginations distributed randomly on the cell membrane of vegetative growing cells ([25]; Figure 3G,H). Using QFDE-EM, many invaginations were also observed on the spore cell membrane. Notably, however, they had a considerably different appearance from those of the vegetative cell membrane: the invaginations of the spore membrane were longer than those of the vegetative cell membrane (Figure 3B–D). In addition, they were aligned in parallel (Figure 3C). The depth and the width were 34.9 ± 4.9 nm and 34.8 ± 6.9 nm, respectively, while the intervals between the invaginations were about 164 ± 34 nm. By contrast, the depth was 32.0 ± 5.3 nm on the vegetative cell membrane. These invaginations were also observed in isp3∆ spores (Figure 3E,F), indicating that the presence of the Isp3 layer does not affect the formation of these invaginations.
3.3. Spore Germination
When appropriate nutrients are supplied, spores germinate to exit dormancy and resume growth. The process of germination involves a marked morphological change from the dormant cell “spore” to the vegetative growing cell. We therefore used QFDE-EM to observe this process.
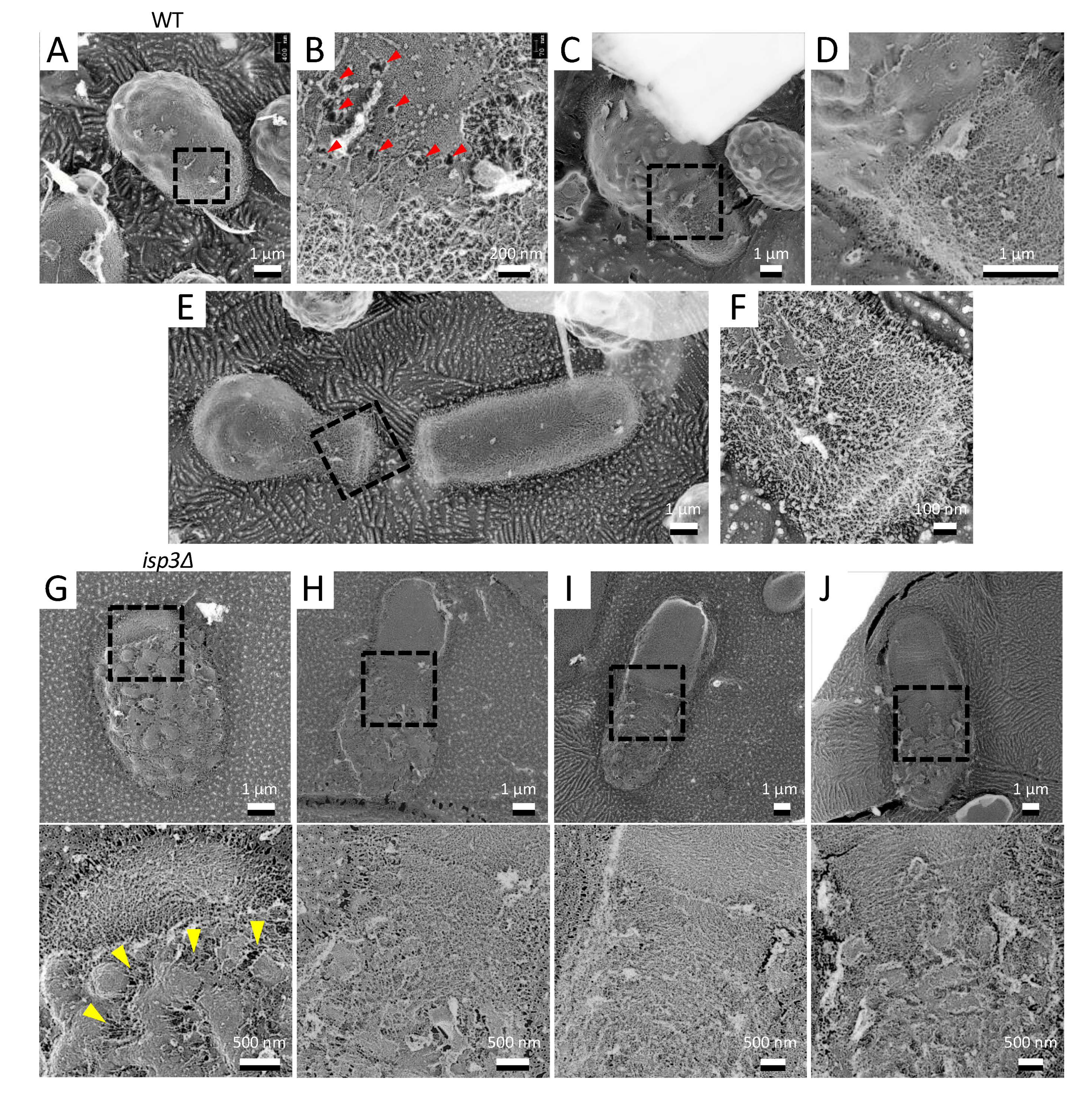
During germination, S. pombe spores develop a polar tube that hatches out of the outer spore wall in a process termed “outgrowth” [25]. QFDE-FM showed that there was no fibrillar layer on the growing tubes, and the inner dendritic structures were exposed (Figure 4A,B). In addition, at the boundary region of the cell surface between the spore and the tube, we frequently observed holes with exposed fibers (Figure 4B). As germination proceeded, the polar tube became coated with the dendritic structure, but the rest of the surface retained the structure normally observed on the spore wall surface (Figure 4C–F).
We also observed the germination process in isp3∆ spores in which the outermost layer of the spore wall is defective. As germination proceeded, the spore wall of isp3∆ collapsed faster than that of wild-type spores. Notably, the collapse was clearly observed outside the tip of the protrusion, suggesting that the Isp3 layer plays an important role in maintaining the cell wall during germination.
4. Discussion
4.1. The Surface of the S. pombe Spore
In the present study, S. pombe spores were visualized at nanometer resolution by using QFDE-EM. First, we found that an interwoven fibrillar layer coats the spore surface, and we showed that this layer corresponds to the Isp3 layer, the outermost proteinaceous layer of the spore wall [9,15]. Interestingly, this fibrillar layer has a very similar appearance to the surface of the conidium, known as the “rodlet layer”, in filamentous fungi [26]. The rodlet layer is a significantly stable protein structure that is resistant to various environmental stresses and comprises a ubiquitously expressed protein named hydrophobin. The primary role of hydrophobins is probably to render the spore surface hydrophobic and water resistant, thereby facilitating spore dispersal in the air [27,28,29]. Notably, however, the Isp3 protein does not show any amino acid sequence similarity to known hydrophobins. Furthermore, the isp3Δ mutation does not affect the hydrophobicity of the spore surface (our unpublished data). Thus, although the morphology of the layers formed by Isp3 and hydrophobin is similar, the roles of these layers may be different.
In the budding yeast Saccharomyces cerevisiae, the ultrastructure of the spore has been well studied by thin-section microscopy and SEM [30,31,32]. Interestingly, the appearance of the spore surface of S. cerevisiae is considerably different from that of S. pombe: The S. cerevisiae spore has a distinctive ridged or scalloped appearance [30], whereas the S. pombe spore has many protrusions. It seems likely that this difference is due to the component of the outermost layer of the spore: S. cerevisiae spores are coated by a dityrosine layer that comprises mainly a cross-linked modified di-amino acid, LL-N,N′-bisformyl dityrosine [33]. In a mutant of the DIT1 gene responsible for formation of the dityrosine layer, the ridged structure on the spore surface is defective, demonstrating that the ridged structure is composed of the dityrosine polymer itself. In S. pombe spores, by contrast, the protrusions still persist on the surface of isp3∆ spores ([15]; Figure 1G), even though the fibrillar layer is defective, suggesting that the protrusion comprises polysaccharides.
QFDE-EM also showed that vegetative cells are covered with a dendritic structure. The structure and composition of the cell wall of S. pombe vegetative cells are well established. The major structural components are α- and β-glucans, both of which form the framework of the cell wall [7,34,35]. High-resolution SEM of intact cells of S. pombe has revealed that the exterior of the vegetative cell wall is primarily composed of α-galactomannoproteins [35], suggesting that the dendritic structure comprises galactomannan.
We also observed the structural change that takes place on the cell surface during spore germination by using QFDE-EM. As germination proceeded, the spore wall collapsed in the region where the germ tube subsequently forms. Furthermore, we found that a number of holes, through which the inner dendritic structures were exposed, emerged on the germinating spore surface. It is unclear how these holes are formed. The protrusions and the fibrillar layer were retained on the mother cell (i.e., spore) after the first cell division (Figure 4E). Notably, the collapse of the structure occurred faster in isp3∆ spores than in wild-type spores. Our previous study has described that isp3∆ spores do not show obvious defects in spore germination (e.g., germination rate and growth speed after germination) [15]. One possibility is that the faster collapse of the spore wall may affect germination under special conditions, including environmental stresses.
4.2. Invaginations on the Spore Cell Membrane
Since the yeast cell is covered by the cell wall, QFDE-EM is a very effective method to observe the cell membrane structure in three dimensions. The present study revealed that many long and deep invaginations are aligned in parallel on the spore cell membrane (Figure 3A). This seems to differ essentially from the vegetative cell membrane, where short invaginations are distributed randomly (Figure 3G; [36]). The invaginations on the vegetative cell membrane resemble those seen on the S. cerevisiae cell membrane, which are called “eisosomes” [37,38] and act as sites of endocytosis. Therefore, it is possible that the invaginations on the S. pombe vegetative cell membrane are similar to the eisosomes of S. cerevisiae. However, it seems unlikely that the invaginations on the spore cell membrane function as eisosomes because the spore is a dormant cell. What might be the physiological function of the invaginations on the spore cell membrane? Lee et al., have reported the presence of invaginations on the cell membrane of various fungi, microalgae, and lichens, using QFDE-EM [39]. In most of the organisms that they examined, the invaginations were observed as randomly distributed patterns. Very interestingly, the red algae Cyanidioshizon merolae has deep and long invaginations that are aligned in parallel similar to the S. pombe spore. C. merolae was originally isolated from hot springs and grows under severe conditions, including high temperature (i.e., 50 °C) and acidic conditions (pH 1–2). Given the high resistance of the spore to various stresses, we presume that the deep and long invaginations are involved in survival under conditions of severe stress.
In summary, by using QFDE-EM, we have identified novel structural features in S. pombe spores. In future studies, elucidation of the molecular mechanisms by which these features are formed will unveil their physiological functions.
Author Contributions
Conceptualization: Y.O.T., M.M., and T.N.; methodology: Y.O.T., M.M., and T.N.; formal analysis: Y.O.T.; resources: Y.O.T., M.M., and T.N.; data curation: Y.O.T.; writing—original draft preparation: Y.O.T., and T.N.; writing—review and editing: M.M., and T.N.; visualization: Y.O.T.; project administration: M.M., and T.N.; funding acquisition: M.M. and T.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Osaka City University (OCU) Strategic Research Grant 2018 for top priority researches to M.M. and a Grant-in Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan: 18K06225 to T.N.
Acknowledgments
We appreciate Kazuki Imada at National Institute of Technology, Suzuka College for fruitful discussions, Kana Fukunishi and Junko Shiomi at Osaka City University for technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Egel, R. Selective spore survival during replica-plating of fission yeast. Arch. Microbiol. 1977, 112, 109–110. [Google Scholar] [CrossRef]
- Kupiec, M.; Byers, B.; Esposito, B. Meiosis and sporulation in Saccharomyces cerevisiae. In The molecular and Cellular Biology of the Yeast Saccharomyces; Jones, E.W., Pringle, J.R., Broach, J.R., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997; pp. 889–1036. [Google Scholar]
- Yoo, B.Y.; Calleja, G.B.; Johnson, B.F. Ultrastructural changes of the fission yeast (Schizosaccharomyces pombe) during ascospore formation. Arch. Microbiol. 1973, 91, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Hirata, A. Ascospore development in the fission yeasts Schizosaccharomyces pombe and S. japonicus. J. Cell Sci. 1982, 56, 263–279. [Google Scholar] [PubMed]
- Hirata, A.; Shimoda, C. Electron microscopic examination of sporulation-deficient mutants of the fission yeast Schizosaccharomyces pombe. Arch. Microbiol. 1992, 158, 249–255. [Google Scholar] [CrossRef]
- Osumi, M. Development of methods for observing of bacteria and fungi by electron microscopy. QJM Int. J. Med. 1989, 38, S150. [Google Scholar]
- Humbel, B.M.; Konomi, M.; Takagi, T.; Kamasawa, N.; Ishijima, S.A.; Osumi, M. In situ localization of β-glucans in the cell wall of Schizosaccharomyces pombe. Yeast 2001, 18, 433–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duran, A.; Perez, P. Cell wall synthesis. In Molecular Biology of Schizosaccharomyces pombe; Springer: Berlin/Heidelberg, Germany, 2004; pp. 269–280. [Google Scholar]
- Shimoda, C. Existence of a major spore wall protein (23K) in the fission yeast Schizosaccharomyces pombe: A possible sporulation-specific protein. Arch. Microbiol. 1983, 136, 26–27. [Google Scholar] [CrossRef]
- Martin, V.; Ribas, J.C.; Carnero, E.; Duran, A.; Sánchez, Y. bgs2+, a sporulation-specific glucan synthase homologue is required for proper ascospore wall maturation in fission yeast. Mol. Microbiol. 2000, 38, 308–321. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Tang, X.; Wang, H.; Balasubramanian, M. Bgs2p, a 1,3-β-glucan synthase subunit, is essential for maturation of ascospore wall in Schizosaccharomyces pombe. FEBS Lett. 2000, 478, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Arellano, M.; Cartagena-Lirola, H.; Hajibagheri, M.A.N.; Duran, A.; Valdivieso, M.-H. Proper ascospore maturation requires the chs1+ chitin synthase gene in Schizosaccharomyces pombe. Mol. Microbiol. 2000, 35, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, Y.; Tanaka, K.; Matsuda, H.; Kawamukai, M. cda1+, encoding chitin deacetylase is required for proper spore formation in Schizosaccharomyces pombe. FEBS Lett. 2005, 579, 2737–2743. [Google Scholar] [CrossRef] [Green Version]
- Garcia, I.; Tajadura, V.; Martín, Y.S.; Toda, T.; Sanchez, Y. Synthesis of α-glucans in fission yeast spores is carried out by three α-glucan synthase paralogues, Mok12p, Mok13p and Mok14p. Mol. Microbiol. 2006, 59, 836–853. [Google Scholar] [CrossRef] [Green Version]
- Fukunishi, K.; Miyakubi, K.; Hatanaka, M.; Otsuru, N.; Hirata, A.; Shimoda, C.; Nakamura, T. The fission yeast spore is coated by a proteinaceous surface layer comprising mainly Isp3. Mol. Biol. Cell 2014, 25, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Abe, H.; Hirata, A.; Shimoda, C. ADAM family protein Mde10 is essential for development of spore envelopes in the fission yeast Schizosaccharomyces pombe. Eukaryot. Cell 2004, 3, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuser, J.E.; Reese, T.S.; Dennis, M.J.; Jan, Y.; Jan, L.; Evans, L. Synaptic vesicle exocytosis captured by quick freezing and correlated with quantal transmitter release. J. Cell Biol. 1979, 81, 275–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuser, J.E. The origins and evolution of freeze-etch electron microscopy. Microbiology 2011, 60, S3–S29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura-Kubo, M.; Nakamura, T.; Hirata, A.; Shimoda, C. The fission yeast spo14+ gene encoding a functional homologue of budding yeast Sec12 is required for the development of forespore membranes. Mol. Biol. Cell 2003, 14, 1109–1124. [Google Scholar] [CrossRef] [Green Version]
- Egel, R.; Egel-Mitani, M. Premeiotic DNA synthesis in fission yeast. Exp. Cell Res. 1974, 88, 127–134. [Google Scholar] [CrossRef]
- Moreno, S.; Klar, A.; Nurse, P. Molecular genetic analysis of fission yeast Schizosaccharomyces pombe. Methods Enzymol. 1991, 194, 795–823. [Google Scholar] [CrossRef]
- Nishi, K.; Shimoda, C.; Hayashibe, M. Germination and outgrowth of Schizosaccharomyces pombe ascospores isolated by Urografin density gradient centrifugation. Can. J. Microbiol. 1978, 24, 893–897. [Google Scholar] [CrossRef]
- Miyata, M.; Petersen, J.D. Spike structure at the interface between gliding Mycoplasma mobile cells and glass surfaces visualized by rapid-freeze-and-fracture electron microscopy. J. Bacteriol. 2004, 186, 4382–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulum, I.; Tahara, Y.O.; Miyata, M. Peptidoglycan layer and disruption processes in Bacillus subtilis cells visualized using quick-freeze, deep-etch electron microscopy. Microbiology 2019, 68, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Hatanaka, M.; Shimoda, C. The cyclic AMP/PKA signal pathway is required for initiation of spore germination in Schizosaccharomyces pombe. Yeast 2001, 18, 207–217. [Google Scholar] [CrossRef]
- Hess, W.M.; Sassen, M.M.; Remsen, C.C. Surface characteristics of Penicillum conidia. Microbiology 1968, 60, 290–303. [Google Scholar]
- Wessels, J.G. Hydrophobins: Proteins that change the nature of the fungal surface. Adv. Microbial. Physiol. 1996, 38, 1–45. [Google Scholar] [CrossRef]
- Wosten, H.A.; de Vocht, M.L. Hydrophobins, the fungal coat unravelled. Biochim. Biiophys. Acta 2000, 1469, 79–86. [Google Scholar] [CrossRef]
- Linder, M.B.; Szilvay, G.R.; Nakari-Setälä, T.; Penttilä, M.E. Hydrophobins: The protein-amphiphiles of filamentous fungi. FEMS Microbiol. Rev. 2005, 29, 877–896. [Google Scholar] [CrossRef] [Green Version]
- Coluccio, A.; Neiman, A.M. Interspore bridges: A new feature of the Saccharomyces cerevisiae spore wall. Microbiology 2004, 150 Pt 10, 3189–3196. [Google Scholar] [CrossRef] [Green Version]
- Coluccio, A.; Bogengruber, E.; Conrad, M.N.; Dresser, M.E.; Briza, P.; Neiman, A.M. Morphogenetic pathway of spore wall assembly in Saccharomyces cerevisiae. Eukaryot. Cell 2004, 3, 1464–1475. [Google Scholar] [CrossRef] [Green Version]
- Neiman, A.M. Sporulation in the budding yeast Saccharomyces cerevisiae. Genetics 2011, 189, 737–765. [Google Scholar] [CrossRef] [Green Version]
- Briza, P.; Kalchhauser, H.; Pittenauer, E.; Allmaier, G.; Breitenbach, M. N,N’ Bisformyl Dityrosine is an in vivo precursor of the yeast ascospore wall. J. Biol. Inorg. Chem. 1996, 239, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Bush, D.A.; Horisberger, M.; Horman, I.; Würsch, P. The wall structure of Schizosaccharomyces pombe. Microbiol. 1974, 81, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osumi, M. Visualization of yeast cells by electron microscopy. Microbiology 2012, 61, 343–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeo, K. Lack of invaginations of the plasma membrane during budding and cell division of Saccharomyces cerevisiae and Schizosaccharomyces pombe. FEMS Microbiol. Lett. 1984, 22, 97–100. [Google Scholar] [CrossRef]
- Walther, T.C.; Brickner, J.H.; Aguilar, P.S.; Bernales, S.; Pantoja, C.; Walter, P. Eisosomes mark static sites of endocytosis. Nat. Cell Biol. 2006, 439, 998–1003. [Google Scholar] [CrossRef] [PubMed]
- Strádalová, V.; Stahlschmidt, W.; Grossmann, G.; Blažíková, M.; Rachel, R.; Tanner, W.; Malinsky, J. Furrow-like invaginations of the yeast plasma membrane correspond to membrane compartment of Can1. J. Cell Sci. 2009, 122, 2887–2894. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Heuser, J.E.; Roth, R.; Goodenough, U. Eisosome ultrastructure and evolution in fungi, microalgae, and lichens. Eukaryot. Cell 2015, 14, 1017–1042. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Surface structure of the S. pombe spore. (A) Field image of wild-type spores. Left spore shows a cytoplasmic cross-section. Center spore shows partial detachment of the surface layer. Right spore shows partial detachment of the spore wall. (B) Single spore. (C) Left, Magnified image of the boxed region in B. Right, the fibrillar structures are shown in various colors. (D) Spore with the outermost layer partly removed by fracture. (E) Magnified image of the boxed region in (D). Arrowhead indicates a cross-section of the fractured surface layer. (F) Magnified image of the boxed region in (E). (G) Left, isp3Δ spore. Right, magnified image of the boxed region. (H) Left, wild-type spore pretreated with 1% SDS and 5% β-mercaptoethanol (2-ME). Right, magnified image of the boxed region.
Figure 1.
Surface structure of the S. pombe spore. (A) Field image of wild-type spores. Left spore shows a cytoplasmic cross-section. Center spore shows partial detachment of the surface layer. Right spore shows partial detachment of the spore wall. (B) Single spore. (C) Left, Magnified image of the boxed region in B. Right, the fibrillar structures are shown in various colors. (D) Spore with the outermost layer partly removed by fracture. (E) Magnified image of the boxed region in (D). Arrowhead indicates a cross-section of the fractured surface layer. (F) Magnified image of the boxed region in (E). (G) Left, isp3Δ spore. Right, magnified image of the boxed region. (H) Left, wild-type spore pretreated with 1% SDS and 5% β-mercaptoethanol (2-ME). Right, magnified image of the boxed region.

Figure 2.
Surface structure of vegetative cell. (A) Field image of S. pombe wild-type vegetative cells. (B) Single vegetative cell. (C) Magnified image of the boxed region in B. (D) Single fractured cell. (E) Magnified image of the boxed region in (D). (F) Magnified image of the boxed region in (E).
Figure 2.
Surface structure of vegetative cell. (A) Field image of S. pombe wild-type vegetative cells. (B) Single vegetative cell. (C) Magnified image of the boxed region in B. (D) Single fractured cell. (E) Magnified image of the boxed region in (D). (F) Magnified image of the boxed region in (E).

Figure 3.
Invagination structure on the membrane surface of spores and vegetative cells. (A) Spore with the cell wall detached by fracture. (B) Spore with the cell wall and cell membrane partially exposed by fracture. (C) Magnified image of the boxed region in B. Yellow arrowheads indicate invaginations of the membrane surface. Arrow indicates the interval of the invaginations. (D) Colored image of (C). The fibrillar layer is colored green, the spore wall yellow, and the spore cell membrane red. (E) isp3Δ spore with a fractured spore wall. (F) Magnified image of the boxed region in (E). (G) Vegetative cell surface with fractured cell walls. (H) Magnified image of the boxed region in (G). Arrowheads indicate invaginations of the membrane surface.
Figure 3.
Invagination structure on the membrane surface of spores and vegetative cells. (A) Spore with the cell wall detached by fracture. (B) Spore with the cell wall and cell membrane partially exposed by fracture. (C) Magnified image of the boxed region in B. Yellow arrowheads indicate invaginations of the membrane surface. Arrow indicates the interval of the invaginations. (D) Colored image of (C). The fibrillar layer is colored green, the spore wall yellow, and the spore cell membrane red. (E) isp3Δ spore with a fractured spore wall. (F) Magnified image of the boxed region in (E). (G) Vegetative cell surface with fractured cell walls. (H) Magnified image of the boxed region in (G). Arrowheads indicate invaginations of the membrane surface.

Figure 4.
Morphological changes on the spore surface during germination. (A) Spore germination of a wild-type cell. (B) Magnified image of the boxed region in A. Arrowheads indicate the holes. (C) Spore in which germination is proceeding. (D) Magnified image in the boxed region in (C). (E) Image of the first cell division after germination. (F) Magnified image of the boxed region in (E). (G–J) Top panels, isp3Δ spore germination. Bottom panels, magnified images of the boxed regions above. Arrowheads indicate the collapsed regions of the spore wall.
Figure 4.
Morphological changes on the spore surface during germination. (A) Spore germination of a wild-type cell. (B) Magnified image of the boxed region in A. Arrowheads indicate the holes. (C) Spore in which germination is proceeding. (D) Magnified image in the boxed region in (C). (E) Image of the first cell division after germination. (F) Magnified image of the boxed region in (E). (G–J) Top panels, isp3Δ spore germination. Bottom panels, magnified images of the boxed regions above. Arrowheads indicate the collapsed regions of the spore wall.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tahara, Y.O.; Miyata, M.; Nakamura, T. Quick-Freeze, Deep-Etch Electron Microscopy Reveals the Characteristic Architecture of the Fission Yeast Spore. J. Fungi 2021, 7, 7. https://doi.org/10.3390/jof7010007
AMA Style
Tahara YO, Miyata M, Nakamura T. Quick-Freeze, Deep-Etch Electron Microscopy Reveals the Characteristic Architecture of the Fission Yeast Spore. Journal of Fungi. 2021; 7(1):7. https://doi.org/10.3390/jof7010007
Chicago/Turabian StyleTahara, Yuhei O., Makoto Miyata, and Taro Nakamura. 2021. "Quick-Freeze, Deep-Etch Electron Microscopy Reveals the Characteristic Architecture of the Fission Yeast Spore" Journal of Fungi 7, no. 1: 7. https://doi.org/10.3390/jof7010007
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.