Tonantzin, a New Genus of Bess Beetle (Coleoptera, Passalidae) from a Montane Subtropical Forest in Central Mexico, with a Review of the Taxonomic Significance of the Mesofrontal Structure in Proculini

 ,
,
Abstract
:1. Introduction
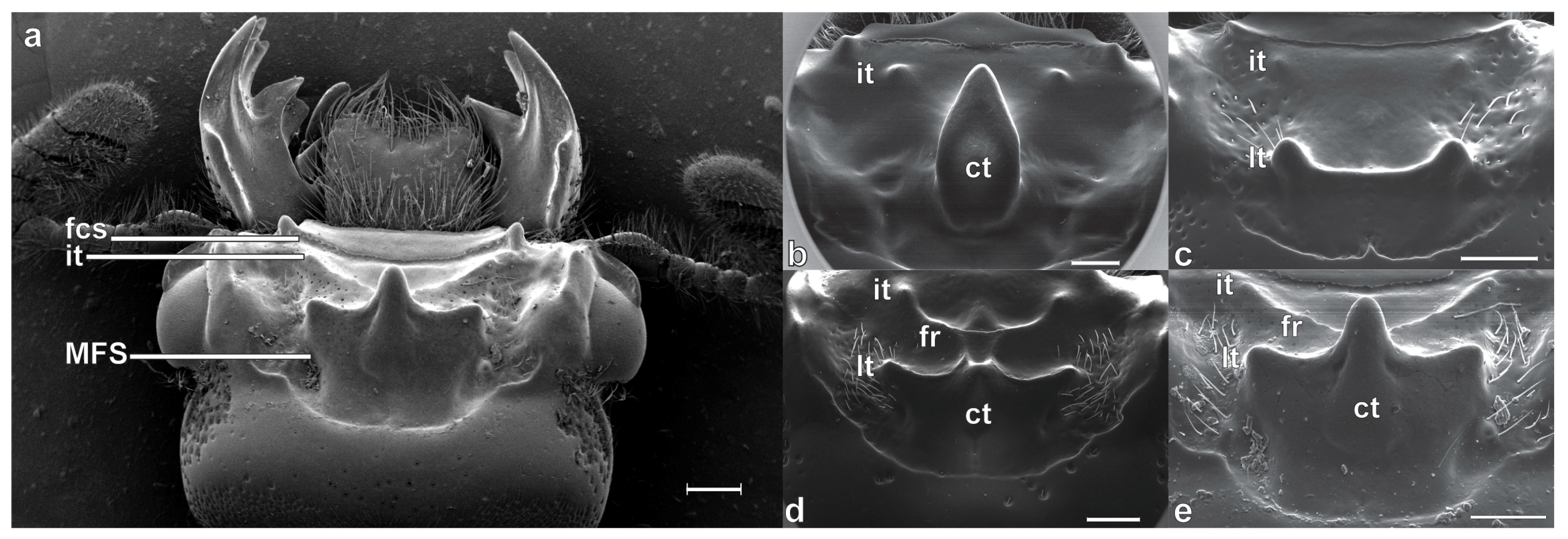
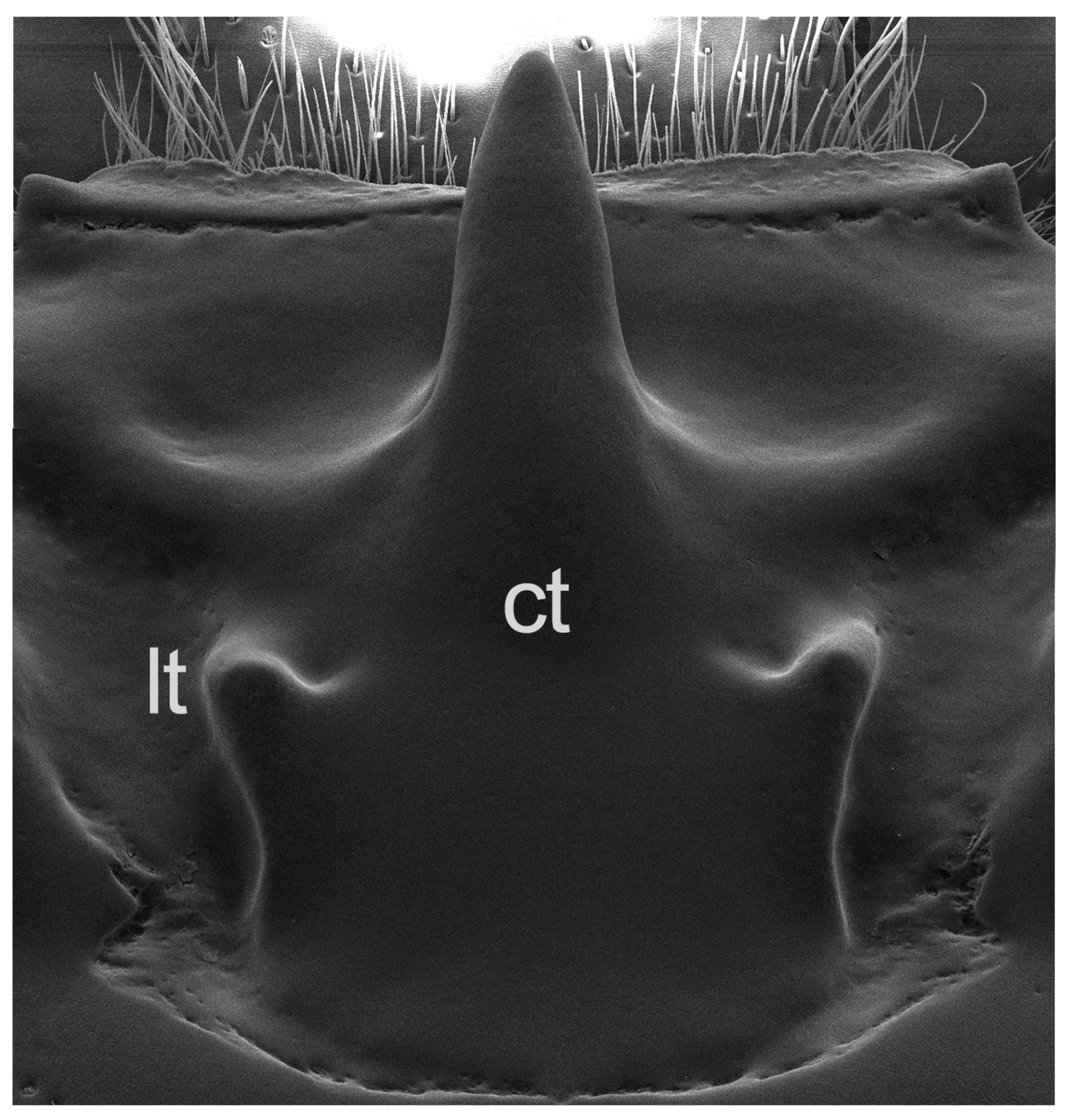
Taxonomic Significance of the Mesofrontal Structure
2. Materials and Methods
2.1. Terminology, Specimen Preparation and Examination
2.2. DNA Extraction and PCR Protocols
2.3. Phylogenetic Analyses
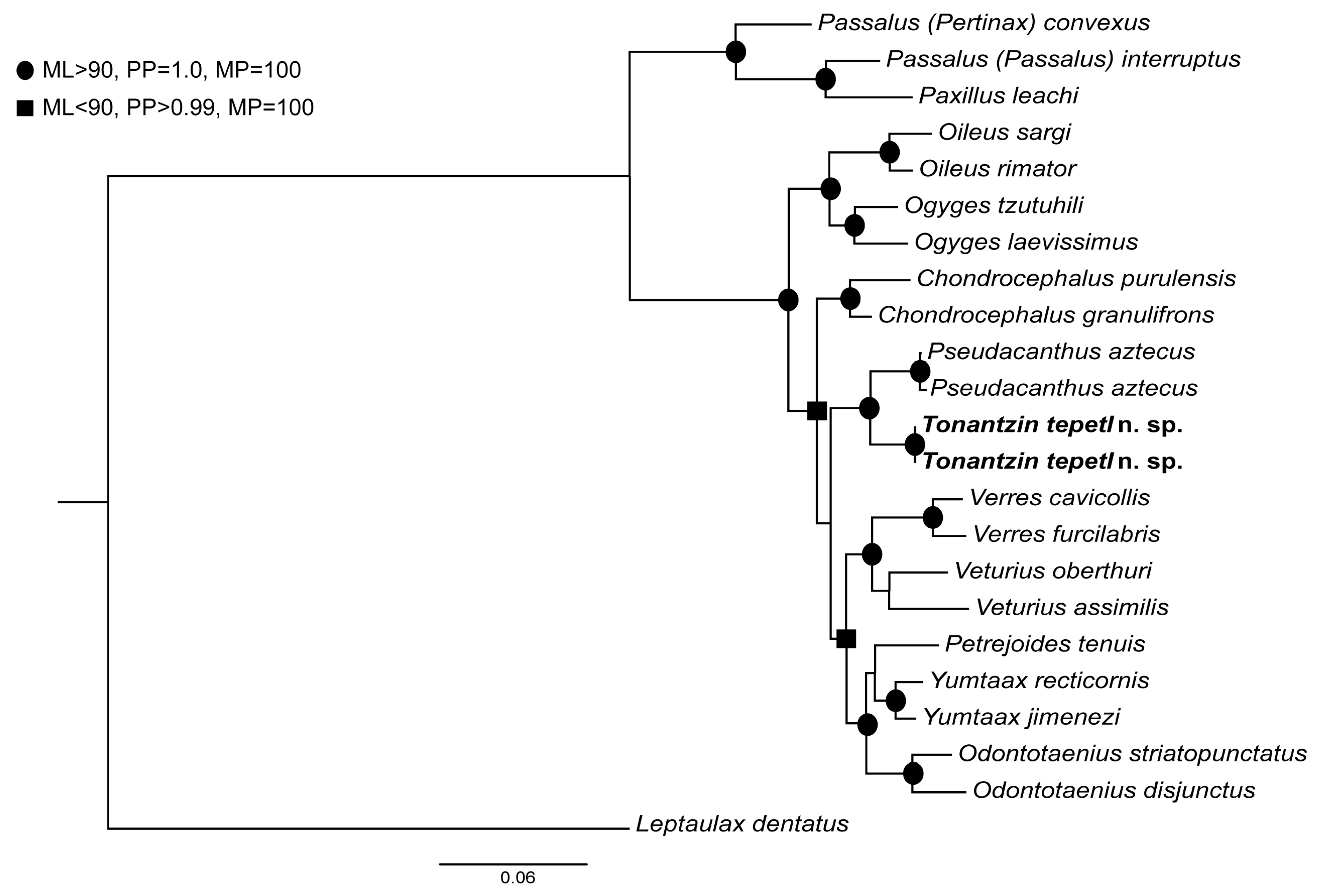
3. Results
3.1. Phylogenetic Analyses
3.2. Taxonomy
3.2.1. Tonantzin Beza-Beza, Jiménez-Ferbans, & Clarke n. gen.
3.2.2. Tonantzin tepetl Beza-Beza, Jiménez-Ferbans, & Clarke n. sp.
 , HOLOTYPE| desig. Beza-Beza et al. 2019.” (IEXA). Paratypes: female. Label data, [hand written]: “? Gen. nov.| det. C. F. Beza-Beza 2013”; [printed] "MEXICO: Veracruz: Municipio Xico,| Ingenio El Rosario, 2920 msnm.| N 19°30′57.3″ W 97°5′32.2″,| 16.vii.2012 P. Reyes-Castillo, C.| Beza-Beza, O. Villerías (cols.)” // “McKenna DNA voucher| DDM3175” // “B197S9” // “Tonantzin tepetl Beza-Beza et al.|
, HOLOTYPE| desig. Beza-Beza et al. 2019.” (IEXA). Paratypes: female. Label data, [hand written]: “? Gen. nov.| det. C. F. Beza-Beza 2013”; [printed] "MEXICO: Veracruz: Municipio Xico,| Ingenio El Rosario, 2920 msnm.| N 19°30′57.3″ W 97°5′32.2″,| 16.vii.2012 P. Reyes-Castillo, C.| Beza-Beza, O. Villerías (cols.)” // “McKenna DNA voucher| DDM3175” // “B197S9” // “Tonantzin tepetl Beza-Beza et al.|  , ALLOTYPE| desig. Beza-Beza et al. 2009.” Designated as Allotype (FMNH). Male. Label data, [hand written]: “? Gen. nove | det. C. F. Beza-Beza 2011”; [printed] "MEXICO: Veracruz: Municipio Xico,| Ingenio El Rosario, 2920 msnm.| N 19°30′57.3″ W 97°5′32.2″,| 16.vii.2012 P. Reyes-Castillo, C.| Beza-Beza, O. Villerías (cols.)” // “McKenna DNA voucher| DDM2883” // “B196S16” // “Tonantzin tepetl Beza-Beza et al.| , PARATYPE| desig. Beza-Beza et al. 2009.” (FMNH).
, ALLOTYPE| desig. Beza-Beza et al. 2009.” Designated as Allotype (FMNH). Male. Label data, [hand written]: “? Gen. nove | det. C. F. Beza-Beza 2011”; [printed] "MEXICO: Veracruz: Municipio Xico,| Ingenio El Rosario, 2920 msnm.| N 19°30′57.3″ W 97°5′32.2″,| 16.vii.2012 P. Reyes-Castillo, C.| Beza-Beza, O. Villerías (cols.)” // “McKenna DNA voucher| DDM2883” // “B196S16” // “Tonantzin tepetl Beza-Beza et al.| , PARATYPE| desig. Beza-Beza et al. 2009.” (FMNH).3.2.3. Taxonomic and Systematic Remarks
4. Discussion
4.1. The Mesofrontal Structure and the Systematics of Proculini
4.2. Biogeography and Micro-Endemism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B

| Voucher | Species | 28s | CAD |
|---|---|---|---|
| DDM3060 | Leptaulax dentatus | MK972968 | MK972161 |
| DDM2869 | Oileus sargi | MK972969 | MK972162 |
| DDM2868 | Odontotaenius striatopunctatus | MK972970 | MK972163 |
| DDM2875 | Verres cavicollis | MK972971 | MK972164 |
| DDM2877 | Pseudacanthus aztecus | MK972972 | MK972165 |
| DDM2878 | Ogyges tzutuhili | MK972973 | MK972166 |
| DDM2879 | Odontotaenius disjunctus | MK972974 | MK972167 |
| DDM2883 | Tonantzin tepetl | MK972975 | MK972168 |
| DDM2886 | Passalus (Pertinax) convexus | MK972976 | MK972169 |
| DDM3051 | Petrejoides tenuis | MK972977 | MK972170 |
| DDM3053 | Veturius oberthuri | MK972978 | MK972171 |
| DDM3057 | Yumtaax recticornis | MK972979 | MK972172 |
| DDM3175 | Tonantzin tepetl | MK972980 | MK972173 |
| DDM3178 | Oileus rimator | MK972981 | MK972174 |
| DDM3186 | Chondrocephalus purulensis | MK972982 | MK972175 |
| DDM3189 | Veturius assimilis | MK972983 | MK972176 |
| DDM3200 | Verres furcilabris | MK972984 | MK972177 |
| DDM3212 | Pseudacanthus aztecus | MK972985 | MK972178 |
| DDM3216 | Yumtaax jimenezi | MK972986 | MK972179 |
| DDM3230 | Chondrocephalus granulifrons | MK972987 | MK972180 |
| DDM3240 | Ogyges laevissimus | MK972988 | MK972181 |
| DDM3262 | Passalus (Passalus) interruptus | MK972989 | MK972182 |
| DDM3282 | Paxillus leachi | MK972990 | MK972183 |
References
- Boucher, S. Évolution et phylogénie des Coléoptères Passalidae (Scarabaeoidea): Les taxons du groupe famille La tribu néotropicale des Proculini et son complexe Veturius. Ann. Société Entomol. Fr. NS 2006, 41, 239–604. [Google Scholar] [CrossRef]
- Jiménez-Ferbans, L.; Amat-García, G.; Reyes-Castillo, P. Estudios de los escarabajos pasálidos (Coleoptera: Passalidae) de Colombia. In Escarabajos del Neotrópico (Insecta: Coleoptera); Deloya, C., Gasca, H., Eds.; S y G Editores: Cuidad de México, México, 2018; pp. 81–94. [Google Scholar]
- Chamé-Vázquez, E.R.C.; Reyes-Castillo, P.; Gómez-Gómez, B.; Ibarra-Núñez, G. Distribución de la familia Passalidae (Coleoptera: Scarabaeoidea) en un gradiente altitudinal en el Soconusco, Chiapas, México. Dugesiana 2018, 25, 115–124. [Google Scholar]
- Reyes-Castillo, P. Coleoptera Passalidae de México. In Hacia un Proyecto CYTED para el Inventario y Estimación de la Diversidad Entomológica en Iberoamérica: PrIBES-2000; Martín-Piera, F., Morrone, J., Melic, A., Eds.; Monografías Tercer Milenio: Zaragoza, España, 2000; Volume 1, pp. 171–182. [Google Scholar]
- Reyes-Castillo, P. Coleoptera, Passalidae: Morfología y división en grandes grupos; géneros Americanos. Folia Entomol. Mex. 1970, 20–22, 1–240. [Google Scholar]
- Schuster, J.; Cano, E. Taxonomy of Passalidae of the New World. Available online: http://unsm-ento.unl.edu/Guide/Scarabaeoidea/Passalidae/Passalidae-Taxonomy.pdf (accessed on 20 May 2019).
- Reyes-Castillo, P.; Chamé-Vázquez, E.R. Nueva especie mexicana de Pseudacanthus Kaup (Coleoptera: Passalidae). Dugesiana 2014, 21, 175–178. [Google Scholar]
- Castillo, C.; Reyes-Castillo, P. Biosistemática del género Petrejoides Kuwert (Coleoptera, Lamelicornia, Passalidae). Acta Zool. Mex. NS 1984, 1, 1–84. [Google Scholar]
- Beza-Beza, C.F.; Beck, J.; Reyes-Castillo, P.; Jameson, M.L. Phylogeny of the genus Yumtaax Boucher (Coleoptera, Passalidae, Proculini): Taxonomic and evolutionary implications with descriptions of three new species. ZooKeys 2017, 667, 95–129. [Google Scholar] [CrossRef]
- Kaup, J.J. Monographie der Passaliden. Berl. Entomol. Z 1871, 15, 1–126. [Google Scholar] [CrossRef]
- Cano, E.; Schuster, J. A new species of Oileus Kaup (Coleoptera, Passalidae) from Guatemala, with a key to the species of the genus. ZooKeys 2012, 194, 81–87. [Google Scholar] [CrossRef]
- Reyes-Castillo, P. Revisión monográfica del género Spurius Kaup (Coleoptera, Passalidae). Stud. Entomol. 1978, 20, 269–290. [Google Scholar]
- Reyes-Castillo, P. Coleoptera: Las especies mexicanas de Heliscus Zang, 1906 (Coleoptera: Passalidae). Dugesiana 2010, 17, 193–196. [Google Scholar]
- Schuster, J.; Cano, E. Key to American Genera of Passalidae. Available online: http://unsm-ento.unl.edu/Guide/Scarabaeoidea/Passalidae/Passalidae-Key/Passalidae-key.pdf (accessed on 20 May 2019).
- Pardo-Locarno, L.C. Escarabajos saproxilófagos Passalidae del chocó biogeográfico. I especies nuevas o poco conocidas y clave genérica para el genero Odontotaenius kuwert. Rev. Agric. Trop. 2012, 35, 36–43. [Google Scholar]
- Pardo-Locarno, L.C. Nueva especie de Verres Kaup (Coleoptera: Passalidae) del Chocó Biogeográfico II: Adiciones y clave para las especies. Boletin Museo Entomológico Francisco Luís Gallego 2014, 6, 7–17. [Google Scholar]
- Reyes-Castillo, P.; Jiménez-Ferbans, L. Aulacocyclus yorkensis a new species of Passalidae (Coleoptera: Scarabaeoidea) from Australia, with a key to the identification of Australian species of the genus. Rev. Bras. Entomol. 2016, 60, 211–213. [Google Scholar] [CrossRef]
- Hanley, R.S.; Ashe, J.S. Techniques for dissecting adult aleocharine beetles (Coleoptera: Staphylinidae). Bull. Entomol. Res. 2003, 93, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Wild, A.L.; Maddison, D.R. Evaluating nuclear protein-coding genes for phylogenetic utility in beetles. Mol. Phylogenet. Evol. 2008, 48, 877–891. [Google Scholar] [CrossRef] [PubMed]
- Whiting, M.F. Mecoptera is paraphyletic: Multiple genes and phylogeny of Mecoptera and Siphonaptera. Zool. Scr. 2002, 31, 93–104. [Google Scholar] [CrossRef]
- Moore, M.R.; Beza-Beza, C.F.; Wickell, D.A.; Beck, J.B.; Jameson, M.L. Molecules, morphology and Mimeoma scarabs: Evolutionary and taxonomic implications for a palm-associated scarab group. Syst. Entomol. 2015, 40, 891–900. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (and other methods) 4.0.b5; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. Partitionfinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Schuster, J.; Reyes-Castillo, P. Coleoptera, Passalidae: Ogyges KAUP, revisión de un género mesoamericano de montaña. Acta Zool. Mex. NS 1990, 40, 1–49. [Google Scholar]
- Schuster, J.C.; Cano, E.B.; Reyes-Castillo, P. Proculus, giant Latin-American passalids: Revision, phylogeny and biogeography. Acta Zool. Mex. NS 2003, 90, 281–306. [Google Scholar]
- Cano, E.B. Ogyges Kaup, a flightless genus of Passalidae (Coleoptera) from Mesoamerica: Nine new species, a key to identify species, and a novel character to support its monophyly. Zootaxa 2014, 3889, 451–484. [Google Scholar] [CrossRef] [PubMed]
- Cano, E.B.; Schuster, J.C.; Morrone, J. Phylogenetics of Ogyges Kaup and the biogeography of Nuclear Central America (Coleoptera, Passalidae). ZooKeys 2018, 737, 81–111. [Google Scholar] [CrossRef] [PubMed]
- Ariza-Marín, E.R.; Reyes-Castillo, P.; Moctezuma, V.; Sánchez-Huerta, J.L. A new species of Vindex Kaup (Coleoptera: Scarabaeoidea: Passalidae: Passalinae: Proculini) from Oaxaca, Mexico. Neotrop. Entomol. 2019, 48, 121–125. [Google Scholar] [CrossRef]
- Jiménez-Ferbans, L.; Reyes-Castillo, P. Phylogeny, biogeography and description of Ameripassalus, a new Mesoamerican genus of Passalidae (Coleoptera). Invertebr. Syst. 2014, 28, 124–144. [Google Scholar] [CrossRef]
- Jiménez-Ferbans, L.; Gónzalez, D.; Reyes-castillo, P. Phylogeny and species delimitation in the group Rhodocanthopus of the genus Passalus (Coleoptera: Passalidae) inferred from morphological and molecular data, with description of two new species. Arthropod syst. Phylo. 2016, 74, 255–266. [Google Scholar]
- Fonseca, C.R.; Reyes-Castillo, P. Novo gênero amazônico de Passalini (Coleoptera, Passalidae, Passalinae). Rev. Bras. Entomol. 1993, 37, 673–681. [Google Scholar]
- Jiménez-Ferbans, L.; Reyes-Castillo, P. Phylogeny and taxonomy of Paxillus Macleay and Passipassalus Reyes-Castillo and Fonseca (Coleoptera: Passalidae: Passalini), with the description of new species. Ann. Entomol. Soc. Am. 2015, 108, 415–434. [Google Scholar] [CrossRef]
- Gillogly, A.R. Review of the Genus Popilius and preliminary phylogeny of Passalidae (Coleoptera). Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2005. [Google Scholar]
- MacVean, C.; Schuster, J.C. Altitudinal distribution of passalid beetles (Coleoptera, Passalidae) and pleistocene dispersal on the volcanic chain of Northern Central America. Biotropica 1981, 13, 29–38. [Google Scholar] [CrossRef]
- Jiménez-Ferbans, L.; Reyes-Castillo, P.; Schuster, J.C. Passalidae (Coleoptera: Scarabaeoidea) of the biogeographical province of Chocó and the west andean region of Colombia, with the description of two new species. Neotrop. Entomol. 2018, 47, 642–667. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, S.R. Geology of Las Cumbres Volcanic Complex, Puebla and Veracruz states, Mexico. Rev. Mex. Cienc. Geológicas 2005, 22, 181–198. [Google Scholar]
- Morales-Mávil, J.E.; Aguilar Rodríguez, S.H. Avifauna del Parque Nacional Cofre de Perote, Veracruz México. For. Veracruzana 2000, 2, 35–40. [Google Scholar]
- Walte-Vega, J.K.; García-Romero, A.; Manzo-Delgado, L.L.; López-García, J. Factores de distribución de las comunidades del bosque húmedo de montaña: Volcán Cofre de Perote, México. Bosque Valdivia 2017, 38, 29–39. [Google Scholar] [CrossRef]
- Cházaro-Basáñez, M. Exploraciones Botánicas en Veracruz y Estados Circunvecinos I. Pisos Altitudinales de Vegetación en el Centro de Veracruz y Zonas Limítrofes con Puebla. Available online: https://cdigital.uv.mx/ (accessed on 17 May 2019).
- De Queiroz, K. The general lineage concept of species, species criteria, and the process of speciation. In Endless forms: Species and Speciation; Howard, D.J., Berlocher, S.H., Eds.; Oxford University Press: Oxford, UK, 1998; pp. 57–75. [Google Scholar]
- Hey, J. On the failure of modern species concepts. Trends Ecol. Evol. 2006, 21, 447–450. [Google Scholar] [CrossRef]
- Wilkinson, M. A comparison of two methods of character construction. Cladistics 1995, 11, 297–308. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beza-Beza, C.F.; Jiménez-Ferbans, L.; Clarke, D.J.; Reyes-Castillo, P.; McKenna, D.D. Tonantzin, a New Genus of Bess Beetle (Coleoptera, Passalidae) from a Montane Subtropical Forest in Central Mexico, with a Review of the Taxonomic Significance of the Mesofrontal Structure in Proculini. Insects 2019, 10, 188. https://doi.org/10.3390/insects10070188
Beza-Beza CF, Jiménez-Ferbans L, Clarke DJ, Reyes-Castillo P, McKenna DD. Tonantzin, a New Genus of Bess Beetle (Coleoptera, Passalidae) from a Montane Subtropical Forest in Central Mexico, with a Review of the Taxonomic Significance of the Mesofrontal Structure in Proculini. Insects. 2019; 10(7):188. https://doi.org/10.3390/insects10070188
Chicago/Turabian StyleBeza-Beza, Cristian Fernando, Larry Jiménez-Ferbans, Dave J. Clarke, Pedro Reyes-Castillo, and Duane D. McKenna. 2019. "Tonantzin, a New Genus of Bess Beetle (Coleoptera, Passalidae) from a Montane Subtropical Forest in Central Mexico, with a Review of the Taxonomic Significance of the Mesofrontal Structure in Proculini" Insects 10, no. 7: 188. https://doi.org/10.3390/insects10070188