
The Molecular Context of Oxidant Stress Response in Cancer Establishes ALDH1A1 as a Critical Target: What This Means for Acute Myeloid Leukemia

{kind=link}
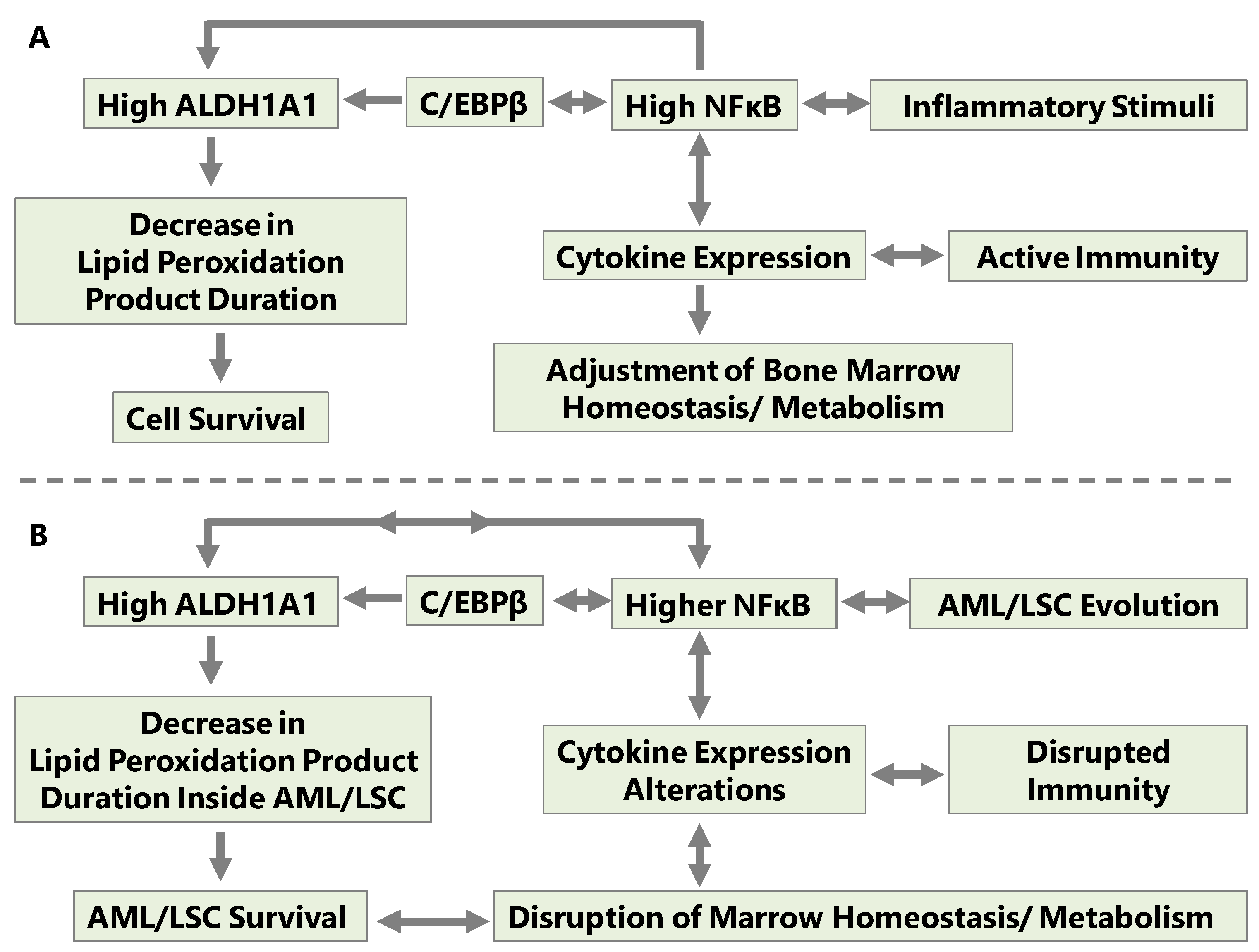{kind=link}
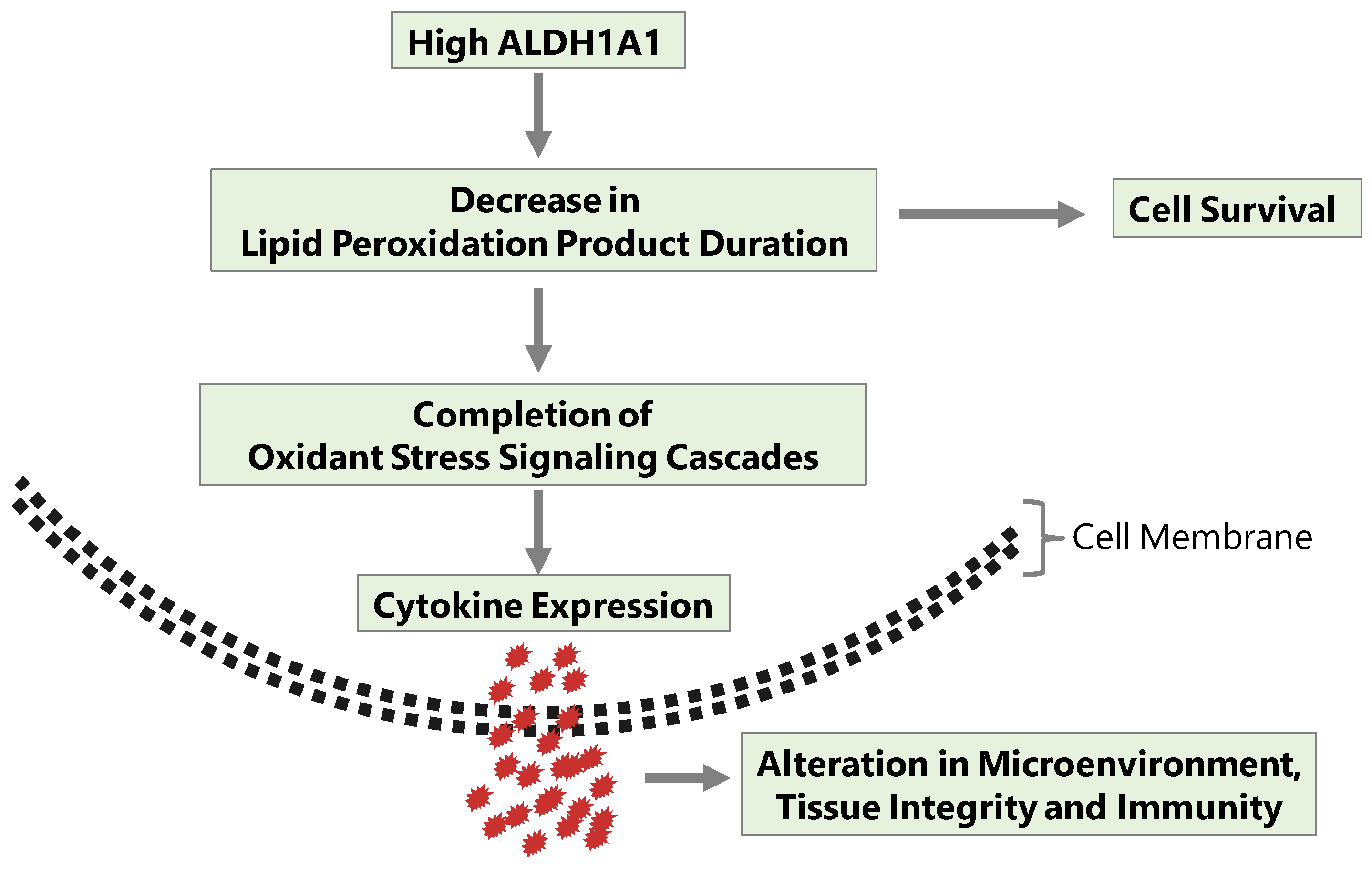{kind=link}
Abstract
:1. Introduction
2. Basic Functions of ALDH1 in Acute Myeloid Leukemia
3. Emerging Role of ALDH1A1 in Cancer Stem Cells: Basic Signaling Pathways
3.1. Regulation of ALDH1A1 in Solid Cancers and Linked Molecular Effects
3.1.1. Non-Small Cell Lung Cancer and Pancreatic Cancer
3.1.2. Intrahepatic Cholangiocarcinoma
3.1.3. Prostate Cancer
3.1.4. Ovarian Cancer
3.1.5. Breast Cancer, Colorectal Cancer, and Melanoma
3.2. ALDH1A1 Is Expressed in Hematologic Cancers: Potential for Repression by ARID1A
4. AML Stem Cells (LSC) and Their Supporting Network
5. AML Poor Prognosis and Coordination of Cellular Turnover and Oxidant Stress
6. An Agent Development Strategy Can Be Based on the Alteration of NFκB Signaling Cascades in AML
6.1. Interfering with Cell Stress Response Kills AML Cells
6.2. Repurposing Existing Molecules as a Prospective Addition to Treatments of AML
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kusuma, G.D.; Abumaree, M.H.; Pertile, M.D.; Perkins, A.V.; Brennecke, S.P.; Kalionis, B. Mesenchymal Stem/Stromal Cells Derived from a Reproductive Tissue Niche under Oxidative Stress Have High Aldehyde Dehydrogenase Activity. Stem Cell Rev. Rep. 2016, 12, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Marchitti, S.A.; Brocker, C.; Stagos, D.; Vasiliou, V. Non-P450 Aldehyde Oxidizing Enzymes: The Aldehyde Dehydrogenase Superfamily. Expert Opin. Drug Metab. Toxicol. 2008, 4, 697–720. [Google Scholar] [CrossRef]
- MacDonagh, L.; Santiago, R.M.; Gray, S.G.; Breen, E.; Cuffe, S.; Finn, S.P.; O’Byrne, K.J.; Barr, M.P. Exploitation of the Vitamin A/Retinoic Acid Axis Depletes ALDH1-Positive Cancer Stem Cells and Re-Sensitises Resistant Non-Small Cell Lung Cancer Cells to Cisplatin. Transl. Oncol. 2021, 14, 101025. [Google Scholar] [CrossRef] [PubMed]
- di Martino, O.; Welch, J.S. Retinoic Acid Receptors in Acute Myeloid Leukemia Therapy. Cancers 2019, 11, 1915. [Google Scholar] [CrossRef]
- Yoval-Sánchez, B.; Rodríguez-Zavala, J.S. Differences in Susceptibility to Inactivation of Human Aldehyde Dehydrogenases by Lipid Peroxidation Byproducts. Chem. Res. Toxicol. 2012, 25, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Suman, S.; Kumar, S.; N’Gouemo, P.; Datta, K. Increased DNA Double-Strand Break Was Associated with Downregulation of Repair and Upregulation of Apoptotic Factors in Rat Hippocampus after Alcohol Exposure. Alcohol 2016, 54, 45–50. [Google Scholar] [CrossRef]
- Feng, Z.; Hu, W.; Tang, M.-S. Trans-4-Hydroxy-2-Nonenal Inhibits Nucleotide Excision Repair in Human Cells: A Possible Mechanism for Lipid Peroxidation-Induced Carcinogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 8598–8602. [Google Scholar] [CrossRef]
- Winczura, A.; Czubaty, A.; Winczura, K.; Masłowska, K.; Nałęcz, M.; Dudzińska, D.A.; Saparbaev, M.; Staroń, K.; Tudek, B. Lipid Peroxidation Product 4-Hydroxy-2-Nonenal Modulates Base Excision Repair in Human Cells. DNA Repair 2014, 22, 1–11. [Google Scholar] [CrossRef]
- Gasparetto, M.; Sekulovic, S.; Brocker, C.; Tang, P.; Zakaryan, A.; Xiang, P.; Kuchenbauer, F.; Wen, M.; Kasaian, K.; Witty, M.F.; et al. Aldehyde Dehydrogenases Are Regulators of Hematopoietic Stem Cell Numbers and B-Cell Development. Exp. Hematol. 2012, 40, 318–329.e2. [Google Scholar] [CrossRef]
- Levi, B.P.; Yilmaz, O.H.; Duester, G.; Morrison, S.J. Aldehyde Dehydrogenase 1a1 Is Dispensable for Stem Cell Function in the Mouse Hematopoietic and Nervous Systems. Blood 2009, 113, 1670–1680. [Google Scholar] [CrossRef]
- Chute, J.P.; Muramoto, G.G.; Whitesides, J.; Colvin, M.; Safi, R.; Chao, N.J.; McDonnell, D.P. Inhibition of Aldehyde Dehydrogenase and Retinoid Signaling Induces the Expansion of Human Hematopoietic Stem Cells. Proc. Natl. Acad. Sci. USA 2006, 103, 11707–11712. [Google Scholar] [CrossRef] [PubMed]
- Gasparetto, M.; Pei, S.; Minhajuddin, M.; Khan, N.; Pollyea, D.A.; Myers, J.R.; Ashton, J.M.; Becker, M.W.; Vasiliou, V.; Humphries, K.R.; et al. Targeted Therapy for a Subset of Acute Myeloid Leukemias That Lack Expression of Aldehyde Dehydrogenase 1A1. Haematologica 2017, 102, 1054–1065. [Google Scholar] [CrossRef] [PubMed]
- Venton, G.; Pérez-Alea, M.; Baier, C.; Fournet, G.; Quash, G.; Labiad, Y.; Martin, G.; Sanderson, F.; Poullin, P.; Suchon, P.; et al. Aldehyde Dehydrogenases Inhibition Eradicates Leukemia Stem Cells While Sparing Normal Progenitors. Blood Cancer J. 2016, 6, e469. [Google Scholar] [CrossRef]
- Mochizuki-Kashio, M.; Shiozaki, H.; Suda, T.; Nakamura-Ishizu, A. Mitochondria Turnover and Lysosomal Function in Hematopoietic Stem Cell Metabolism. Int. J. Mol. Sci. 2021, 22, 4627. [Google Scholar] [CrossRef] [PubMed]
- Anastasiou, V.; Ninou, E.; Alexopoulou, D.; Stertmann, J.; Müller, A.; Dahl, A.; Solimena, M.; Speier, S.; Serafimidis, I.; Gavalas, A. Aldehyde Dehydrogenase Activity Is Necessary for Beta Cell Development and Functionality in Mice. Diabetologia 2016, 59, 139–150. [Google Scholar] [CrossRef]
- Honeth, G.; Lombardi, S.; Ginestier, C.; Hur, M.; Marlow, R.; Buchupalli, B.; Shinomiya, I.; Gazinska, P.; Bombelli, S.; Ramalingam, V.; et al. Aldehyde Dehydrogenase and Estrogen Receptor Define a Hierarchy of Cellular Differentiation in the Normal Human Mammary Epithelium. Breast Cancer Res. BCR 2014, 16, R52. [Google Scholar] [CrossRef]
- Serio, R.N.; Laursen, K.B.; Urvalek, A.M.; Gross, S.S.; Gudas, L.J. Ethanol Promotes Differentiation of Embryonic Stem Cells through Retinoic Acid Receptor-γ. J. Biol. Chem. 2019, 294, 5536–5548. [Google Scholar] [CrossRef]
- Gudas, L.J. Emerging Roles for Retinoids in Regeneration and Differentiation in Normal and Disease States. Biochim. Biophys. Acta 2012, 1821, 213–221. [Google Scholar] [CrossRef]
- Yang, X.; Yao, R.; Wang, H. Update of ALDH as a Potential Biomarker and Therapeutic Target for AML. BioMed Res. Int. 2018, 2018, 9192104. [Google Scholar] [CrossRef]
- Schuurhuis, G.J.; Meel, M.H.; Wouters, F.; Min, L.A.; Terwijn, M.; de Jonge, N.A.; Kelder, A.; Snel, A.N.; Zweegman, S.; Ossenkoppele, G.J.; et al. Normal Hematopoietic Stem Cells within the AML Bone Marrow Have a Distinct and Higher ALDH Activity Level than Co-Existing Leukemic Stem Cells. PLoS ONE 2013, 8, e78897. [Google Scholar] [CrossRef]
- Dancik, G.M.; Voutsas, I.F.; Vlahopoulos, S. Lower RNA Expression of ALDH1A1 Distinguishes the Favorable Risk Group in Acute Myeloid Leukemia. Mol. Biol. Rep. 2022, 49, 3321–3331. [Google Scholar] [CrossRef] [PubMed]
- Dancik, G.M.; Voutsas, I.F.; Vlahopoulos, S. Aldehyde Dehydrogenase Enzyme Functions in Acute Leukemia Stem Cells. Front. Biosci. Sch. Ed. 2022, 14, 8. [Google Scholar] [CrossRef]
- Rahmati, A.; Goudarzi, S.; Sheikhi, M.; Siyadat, P.; Ferns, G.A.; Ayatollahi, H. The Emerging Roles of Aldehyde Dehydrogenase in Acute Myeloid Leukemia and Its Therapeutic Potential. Anticancer Agents Med. Chem. 2022, 23, 246–255. [Google Scholar] [CrossRef]
- Johansson, B. A Review of the Pharmacokinetics and Pharmacodynamics of Disulfiram and Its Metabolites. Acta Psychiatr. Scand. Suppl. 1992, 369, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xiang, Y.; Xiang, L.; Xiao, Y.; Li, F.; Hao, P. ALDH Maintains the Stemness of Lung Adenoma Stem Cells by Suppressing the Notch/CDK2/CCNE Pathway. PLoS ONE 2014, 9, e92669. [Google Scholar] [CrossRef]
- Mohan, A.; Raj, R.R.; Mohan, G.; Padmaja, K.P.; Maliekal, T.T. Reporters of Cancer Stem Cells as a Tool for Drug Discovery. Front. Oncol. 2021, 11, 669250. [Google Scholar] [CrossRef]
- Wang, D.; Wang, H. Oxazaphosphorine Bioactivation and Detoxification The Role of Xenobiotic Receptors. Acta Pharm. Sin. B 2012, 2, 107–117. [Google Scholar] [CrossRef]
- Yue, H.; Hu, Z.; Hu, R.; Guo, Z.; Zheng, Y.; Wang, Y.; Zhou, Y. ALDH1A1 in Cancers: Bidirectional Function, Drug Resistance, and Regulatory Mechanism. Front. Oncol. 2022, 12, 918778. [Google Scholar] [CrossRef] [PubMed]
- Nacarino-Palma, A.; Rejano-Gordillo, C.M.; González-Rico, F.J.; Ordiales-Talavero, A.; Román, Á.C.; Cuadrado, M.; Bustelo, X.R.; Merino, J.M.; Fernández-Salguero, P.M. Loss of Aryl Hydrocarbon Receptor Favors K-RasG12D-Driven Non-Small Cell Lung Cancer. Cancers 2021, 13, 4071. [Google Scholar] [CrossRef]
- Liu, S.; Cao, W.; Niu, Y.; Luo, J.; Zhao, Y.; Hu, Z.; Zong, C. Single-PanIN-Seq Unveils That ARID1A Deficiency Promotes Pancreatic Tumorigenesis by Attenuating KRAS-Induced Senescence. eLife 2021, 10, e64204. [Google Scholar] [CrossRef]
- Liu, L.; Salnikov, A.V.; Bauer, N.; Aleksandrowicz, E.; Labsch, S.; Nwaeburu, C.; Mattern, J.; Gladkich, J.; Schemmer, P.; Werner, J.; et al. Triptolide Reverses Hypoxia-Induced Epithelial-Mesenchymal Transition and Stem-like Features in Pancreatic Cancer by NF-ΚB Downregulation. Int. J. Cancer 2014, 134, 2489–2503. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, J.; Akiyama, Y.; Shimada, S.; Ogura, T.; Ogawa, K.; Ono, H.; Mitsunori, Y.; Ban, D.; Kudo, A.; Yamaoka, S.; et al. Loss of ARID1A Induces a Stemness Gene ALDH1A1 Expression with Histone Acetylation in the Malignant Subtype of Cholangiocarcinoma. Carcinogenesis 2020, 41, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, Q.; Han, Y.; Pei, S.; Cheng, B.; Xu, J.; Miao, X.; Pan, Q.; Wang, H.; Guo, J.; et al. ARID1A Loss Induces Polymorphonuclear Myeloid-Derived Suppressor Cell Chemotaxis and Promotes Prostate Cancer Progression. Nat. Commun. 2022, 13, 7281. [Google Scholar] [CrossRef]
- Matsika, A.; Srinivasan, B.; Day, C.; Mader, S.A.; Kiernan, D.M.; Broomfield, A.; Fu, J.; Hooper, J.D.; Kench, J.G.; Samaratunga, H. Cancer Stem Cell Markers in Prostate Cancer: An Immunohistochemical Study of ALDH1, SOX2 and EZH2. Pathology 2015, 47, 622–628. [Google Scholar] [CrossRef]
- Federer-Gsponer, J.R.; Müller, D.C.; Zellweger, T.; Eggimann, M.; Marston, K.; Ruiz, C.; Seifert, H.-H.; Rentsch, C.A.; Bubendorf, L.; Le Magnen, C. Patterns of Stemness-Associated Markers in the Development of Castration-Resistant Prostate Cancer. Prostate 2020, 80, 1108–1117. [Google Scholar] [CrossRef] [PubMed]
- Webber, J.P.; Spary, L.K.; Mason, M.D.; Tabi, Z.; Brewis, I.A.; Clayton, A. Prostate Stromal Cell Proteomics Analysis Discriminates Normal from Tumour Reactive Stromal Phenotypes. Oncotarget 2016, 7, 20124–20139. [Google Scholar] [CrossRef]
- Meng, E.; Mitra, A.; Tripathi, K.; Finan, M.A.; Scalici, J.; McClellan, S.; Madeira da Silva, L.; Reed, E.; Shevde, L.A.; Palle, K.; et al. ALDH1A1 Maintains Ovarian Cancer Stem Cell-like Properties by Altered Regulation of Cell Cycle Checkpoint and DNA Repair Network Signaling. PLoS ONE 2014, 9, e107142. [Google Scholar] [CrossRef] [PubMed]
- Nwani, N.G.; Condello, S.; Wang, Y.; Swetzig, W.M.; Barber, E.; Hurley, T.; Matei, D. A Novel ALDH1A1 Inhibitor Targets Cells with Stem Cell Characteristics in Ovarian Cancer. Cancers 2019, 11, 502. [Google Scholar] [CrossRef]
- Allison, S.E.; Chen, Y.; Petrovic, N.; Zhang, J.; Bourget, K.; Mackenzie, P.I.; Murray, M. Activation of ALDH1A1 in MDA-MB-468 Breast Cancer Cells That over-Express CYP2J2 Protects against Paclitaxel-Dependent Cell Death Mediated by Reactive Oxygen Species. Biochem. Pharmacol. 2017, 143, 79–89. [Google Scholar] [CrossRef]
- Canter, R.J.; Ames, E.; Mac, S.; Grossenbacher, S.K.; Chen, M.; Li, C.-S.; Borys, D.; Smith, R.C.; Tellez, J.; Sayers, T.J.; et al. Anti-Proliferative but Not Anti-Angiogenic Tyrosine Kinase Inhibitors Enrich for Cancer Stem Cells in Soft Tissue Sarcoma. BMC Cancer 2014, 14, 756. [Google Scholar] [CrossRef]
- Antar, A.; Otrock, Z.K.; El-Cheikh, J.; Kharfan-Dabaja, M.A.; Battipaglia, G.; Mahfouz, R.; Mohty, M.; Bazarbachi, A. Inhibition of FLT3 in AML: A Focus on Sorafenib. Bone Marrow Transplant. 2017, 52, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Xuan, L.; Wang, Y.; Huang, F.; Fan, Z.; Xu, Y.; Sun, J.; Xu, N.; Deng, L.; Li, X.; Liang, X.; et al. Sorafenib Maintenance in Patients with FLT3-ITD Acute Myeloid Leukaemia Undergoing Allogeneic Haematopoietic Stem-Cell Transplantation: An Open-Label, Multicentre, Randomised Phase 3 Trial. Lancet Oncol. 2020, 21, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Solomon, H.; Dinowitz, N.; Pateras, I.S.; Cooks, T.; Shetzer, Y.; Molchadsky, A.; Charni, M.; Rabani, S.; Koifman, G.; Tarcic, O.; et al. Mutant P53 Gain of Function Underlies High Expression Levels of Colorectal Cancer Stem Cells Markers. Oncogene 2018, 37, 1669–1684. [Google Scholar] [CrossRef]
- Allen, J.E.; Hart, L.S.; Dicker, D.T.; Wang, W.; El-Deiry, W.S. Visualization and Enrichment of Live Putative Cancer Stem Cell Populations Following P53 Inactivation or Bax Deletion Using Non-Toxic Fluorescent Dyes. Cancer Biol. Ther. 2009, 8, 2194–2205. [Google Scholar] [CrossRef]
- Ciccone, V.; Terzuoli, E.; Ristori, E.; Filippelli, A.; Ziche, M.; Morbidelli, L.; Donnini, S. ALDH1A1 Overexpression in Melanoma Cells Promotes Tumor Angiogenesis by Activating the IL-8/Notch Signaling Cascade. Int. J. Mol. Med. 2022, 50, 99. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Lu, K.; Ye, T.; Zhang, Z. MicroRNA-223 Attenuates LPS-induced Inflammation in an Acute Lung Injury Model via the NLRP3 Inflammasome and TLR4/NF-κB Signaling Pathway via RHOB. Int. J. Mol. Med. 2019, 43, 1467–1477. [Google Scholar] [CrossRef]
- Zhou, W.; Pal, A.S.; Hsu, A.Y.-H.; Gurol, T.; Zhu, X.; Wirbisky-Hershberger, S.E.; Freeman, J.L.; Kasinski, A.L.; Deng, Q. MicroRNA-223 Suppresses the Canonical NF-ΚB Pathway in Basal Keratinocytes to Dampen Neutrophilic Inflammation. Cell Rep. 2018, 22, 1810–1823. [Google Scholar] [CrossRef]
- Gentner, B.; Pochert, N.; Rouhi, A.; Boccalatte, F.; Plati, T.; Berg, T.; Sun, S.M.; Mah, S.M.; Mirkovic-Hösle, M.; Ruschmann, J.; et al. MicroRNA-223 Dose Levels Fine Tune Proliferation and Differentiation in Human Cord Blood Progenitors and Acute Myeloid Leukemia. Exp. Hematol. 2015, 43, 858–868.e7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhou, Y.-F.; Cao, J.; Burley, S.K.; Wang, H.-Y.; Zheng, X.F.S. MTORC1 Promotes ARID1A Degradation and Oncogenic Chromatin Remodeling in Hepatocellular Carcinoma. Cancer Res. 2021, 81, 5652–5665. [Google Scholar] [CrossRef]
- Řezníčková, E.; Krajčovičová, S.; Peřina, M.; Kovalová, M.; Soural, M.; Kryštof, V. Modulation of FLT3-ITD and CDK9 in Acute Myeloid Leukaemia Cells by Novel Proteolysis Targeting Chimera (PROTAC). Eur. J. Med. Chem. 2022, 243, 114792. [Google Scholar] [CrossRef]
- Hynes-Smith, R.W.; Swenson, S.A.; Vahle, H.; Wittorf, K.J.; Caplan, M.; Amador, C.; Hyde, R.K.; Buckley, S.M. Loss of FBXO9 Enhances Proteasome Activity and Promotes Aggressiveness in Acute Myeloid Leukemia. Cancers 2019, 11, 1717. [Google Scholar] [CrossRef] [PubMed]
- Göllner, S.; Oellerich, T.; Agrawal-Singh, S.; Schenk, T.; Klein, H.-U.; Rohde, C.; Pabst, C.; Sauer, T.; Lerdrup, M.; Tavor, S.; et al. Loss of the Histone Methyltransferase EZH2 Induces Resistance to Multiple Drugs in Acute Myeloid Leukemia. Nat. Med. 2017, 23, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, B.; Verzella, D.; Capece, D.; Vecchiotti, D.; Di Vito Nolfi, M.; Flati, I.; Cornice, J.; Di Padova, M.; Angelucci, A.; Alesse, E.; et al. NF-ΚB: A Druggable Target in Acute Myeloid Leukemia. Cancers 2022, 14, 3557. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Wang, J.; Tang, W.; Chen, D.; Wang, S.; Zhang, X.; Yang, D. ARID1A Has Prognostic Value in Acute Myeloid Leukemia and Promotes Cell Proliferation via TGF-Β1/SMAD3 Signaling. Clin. Exp. Med. 2022. [Google Scholar] [CrossRef] [PubMed]
- Fabiani, E.; Cicconi, L.; Nardozza, A.M.; Cristiano, A.; Rossi, M.; Ottone, T.; Falconi, G.; Divona, M.; Testi, A.M.; Annibali, O.; et al. Mutational Profile of ZBTB16-RARA-Positive Acute Myeloid Leukemia. Cancer Med. 2021, 10, 3839–3847. [Google Scholar] [CrossRef]
- Loberg, M.A.; Bell, R.K.; Goodwin, L.O.; Eudy, E.; Miles, L.A.; SanMiguel, J.M.; Young, K.; Bergstrom, D.E.; Levine, R.L.; Schneider, R.K.; et al. Sequentially Inducible Mouse Models Reveal That Npm1 Mutation Causes Malignant Transformation of Dnmt3a-Mutant Clonal Hematopoiesis. Leukemia 2019, 33, 1635–1649. [Google Scholar] [CrossRef]
- Loveday, T.; Duns, G.; Rimsza, L.M.; Rech, K.L.; Cook, J.R.; Robetorye, R.S.; Rosenthal, A.C.; Ramsower, C.A.; Yip, T.K.; McKinney, C.L.; et al. Transformation of FL into DLBCL with a PMBL Gene Expression Signature. Blood Adv. 2023, 7, 893–899. [Google Scholar] [CrossRef]
- Song, Y.-H.; Zhong, M.-Z.; Gan, P.-P.; Yi, P.-Y.; Tang, Y.-H.; Liu, Y.-P.; Jiang, J.-Q.; Li, L. ALDH1A1 Mediates Resistance of Diffuse Large B Cell Lymphoma to the CHOP Regimen. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2014, 35, 11809–11817. [Google Scholar] [CrossRef]
- Song, S.; Li, Y.; Zhang, K.; Zhang, X.; Huang, Y.; Xu, M.; Li, S.; Guan, X.; Yang, T.; Liu, Z.; et al. Cancer Stem Cells of Diffuse Large B Cell Lymphoma Are Not Enriched in the CD45+CD19- Cells but in the ALDHhigh Cells. J. Cancer 2020, 11, 142–152. [Google Scholar] [CrossRef]
- Maynard, R.S.; Hellmich, C.; Bowles, K.M.; Rushworth, S.A. Acute Myeloid Leukaemia Drives Metabolic Changes in the Bone Marrow Niche. Front. Oncol. 2022, 12, 924567. [Google Scholar] [CrossRef]
- Yun, H.; Narayan, N.; Vohra, S.; Giotopoulos, G.; Mupo, A.; Madrigal, P.; Sasca, D.; Lara-Astiaso, D.; Horton, S.J.; Agrawal-Singh, S.; et al. Mutational Synergy during Leukemia Induction Remodels Chromatin Accessibility, Histone Modifications and Three-Dimensional DNA Topology to Alter Gene Expression. Nat. Genet. 2021, 53, 1443–1455. [Google Scholar] [CrossRef]
- Vlahopoulos, S.A.; Cen, O.; Hengen, N.; Agan, J.; Moschovi, M.; Critselis, E.; Adamaki, M.; Bacopoulou, F.; Copland, J.A.; Boldogh, I.; et al. Dynamic Aberrant NF-ΚB Spurs Tumorigenesis: A New Model Encompassing the Microenvironment. Cytokine Growth Factor Rev. 2015, 26, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Haladyna, J.N.; Pastuer, T.; Riedel, S.S.; Perraud, A.-L.; Bernt, K.M. Transient Potential Receptor Melastatin-2 (Trpm2) Does Not Influence Murine MLL-AF9-Driven AML Leukemogenesis or in Vitro Response to Chemotherapy. Exp. Hematol. 2016, 44, 596–602.e3. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, A.D.; Hoff, F.W.; Qiu, Y.; Gerbing, R.B.; Gamis, A.S.; Aplenc, R.; Kolb, E.A.; Alonzo, T.A.; Meshinchi, S.; Jenkins, G.N.; et al. Bortezomib Is Significantly Beneficial for de Novo Pediatric AML Patients with Low Phosphorylation of the NF-ΚB Subunit RelA. Proteom. Clin. Appl. 2022, 16, e2100072. [Google Scholar] [CrossRef] [PubMed]
- Bosman, M.C.J.; Schepers, H.; Jaques, J.; Brouwers-Vos, A.Z.; Quax, W.J.; Schuringa, J.J.; Vellenga, E. The TAK1-NF-ΚB Axis as Therapeutic Target for AML. Blood 2014, 124, 3130–3140. [Google Scholar] [CrossRef]
- Bueso-Ramos, C.E.; Rocha, F.C.; Shishodia, S.; Medeiros, L.J.; Kantarjian, H.M.; Vadhan-Raj, S.; Estrov, Z.; Smith, T.L.; Nguyen, M.H.; Aggarwal, B.B. Expression of Constitutively Active Nuclear-Kappa B RelA Transcription Factor in Blasts of Acute Myeloid Leukemia. Hum. Pathol. 2004, 35, 246–253. [Google Scholar] [CrossRef]
- Klimentova, M.; Shelikhova, L.; Ilushina, M.; Kozlovskaya, S.; Blagov, S.; Popov, A.; Kashpor, S.; Fadeeva, M.; Olshanskaya, J.; Glushkova, S.; et al. Targeted Therapy with Venetoclax and Daratumumab as Part of HSCT Preparative Regimen in Children with Chemorefractory Acute Myeloid Leukemia.: Venetoclax and Daratumumab as Part of HSCT Preparative Regimen for AD AML. Transplant. Cell. Ther. 2023, 29, 127.e1–127.e9. [Google Scholar] [CrossRef]
- Weng, G.; Zhang, Y.; Yu, G.; Luo, T.; Yu, S.; Xu, N.; Sun, Z.; Lin, D.; Deng, L.; Liang, X.; et al. Genetic Characteristics Predict Response to Venetoclax plus Hypomethylating Agents in Relapsed or Refractory Acute Myeloid Leukemia. J. Intern. Med. 2023, 293, 329–339. [Google Scholar] [CrossRef]
- Vlahopoulos, S.A. Aberrant Control of NF-ΚB in Cancer Permits Transcriptional and Phenotypic Plasticity, to Curtail Dependence on Host Tissue: Molecular Mode. Cancer Biol. Med. 2017, 14, 254–270. [Google Scholar] [CrossRef]
- Stroopinsky, D.; Rajabi, H.; Nahas, M.; Rosenblatt, J.; Rahimian, M.; Pyzer, A.; Tagde, A.; Kharbanda, A.; Jain, S.; Avigan, D.; et al. MUC1-C Drives Myeloid Leukaemogenesis and Resistance to Treatment by a Survivin-Mediated Mechanism. J. Cell. Mol. Med. 2018, 22, 3887–3898. [Google Scholar] [CrossRef]
- Alam, M.; Ahmad, R.; Rajabi, H.; Kharbanda, A.; Kufe, D. MUC1-C Oncoprotein Activates ERK→C/EBPβ Signaling and Induction of Aldehyde Dehydrogenase 1A1 in Breast Cancer Cells. J. Biol. Chem. 2013, 288, 30892–30903. [Google Scholar] [CrossRef]
- Cirovic, B.; Schönheit, J.; Kowenz-Leutz, E.; Ivanovska, J.; Klement, C.; Pronina, N.; Bégay, V.; Leutz, A. C/EBP-Induced Transdifferentiation Reveals Granulocyte-Macrophage Precursor-like Plasticity of B Cells. Stem Cell Rep. 2017, 8, 346–359. [Google Scholar] [CrossRef]
- Paz-Priel, I.; Houng, S.; Dooher, J.; Friedman, A.D. C/EBPα and C/EBPα Oncoproteins Regulate NFκB1 and Displace Histone Deacetylases from NF-ΚB P50 Homodimers to Induce NF-ΚB Target Genes. Blood 2011, 117, 4085–4094. [Google Scholar] [CrossRef]
- Horton, T.M.; Perentesis, J.P.; Gamis, A.S.; Alonzo, T.A.; Gerbing, R.B.; Ballard, J.; Adlard, K.; Howard, D.S.; Smith, F.O.; Jenkins, G.; et al. A Phase 2 Study of Bortezomib Combined with Either Idarubicin/Cytarabine or Cytarabine/Etoposide in Children with Relapsed, Refractory or Secondary Acute Myeloid Leukemia: A Report from the Children’s Oncology Group. Pediatr. Blood Cancer 2014, 61, 1754–1760. [Google Scholar] [CrossRef]
- Hansen, Q.; Bachas, C.; Smit, L.; Cloos, J. Characteristics of Leukemic Stem Cells in Acute Leukemia and Potential Targeted Therapies for Their Specific Eradication. Cancer Drug Resist. 2022, 5, 344–367. [Google Scholar] [CrossRef]
- Swords, R.T.; Coutre, S.; Maris, M.B.; Zeidner, J.F.; Foran, J.M.; Cruz, J.; Erba, H.P.; Berdeja, J.G.; Tam, W.; Vardhanabhuti, S.; et al. Pevonedistat, a First-in-Class NEDD8-Activating Enzyme Inhibitor, Combined with Azacitidine in Patients with AML. Blood 2018, 131, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Fathi, A.T. Pevonedistat, a New Partner for 5-Azacitidine. Blood 2018, 131, 1391–1392. [Google Scholar] [CrossRef] [PubMed]
- Verzella, D.; Cornice, J.; Arboretto, P.; Vecchiotti, D.; Di Vito Nolfi, M.; Capece, D.; Zazzeroni, F.; Franzoso, G. The NF-ΚB Pharmacopeia: Novel Strategies to Subdue an Intractable Target. Biomedicines 2022, 10, 2233. [Google Scholar] [CrossRef]
- Guzman, M.L.; Neering, S.J.; Upchurch, D.; Grimes, B.; Howard, D.S.; Rizzieri, D.A.; Luger, S.M.; Jordan, C.T. Nuclear Factor-KappaB Is Constitutively Activated in Primitive Human Acute Myelogenous Leukemia Cells. Blood 2001, 98, 2301–2307. [Google Scholar] [CrossRef]
- Khan, I.; Eklund, E.E.; Gartel, A.L. Therapeutic Vulnerabilities of Transcription Factors in AML. Mol. Cancer Ther. 2021, 20, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Moreb, J.; Zucali, J.R.; Zhang, Y.; Colvin, M.O.; Gross, M.A. Role of Aldehyde Dehydrogenase in the Protection of Hematopoietic Progenitor Cells from 4-Hydroperoxycyclophosphamide by Interleukin 1 Beta and Tumor Necrosis Factor. Cancer Res. 1992, 52, 1770–1774. [Google Scholar] [PubMed]
- Moreb, J.S.; Turner, C.; Sreerama, L.; Zucali, J.R.; Sladek, N.E.; Schweder, M. Interleukin-1 and Tumor Necrosis Factor Alpha Induce Class 1 Aldehyde Dehydrogenase MRNA and Protein in Bone Marrow Cells. Leuk. Lymphoma 1995, 20, 77–84. [Google Scholar] [CrossRef]
- Zhou, H.-S.; Carter, B.Z.; Andreeff, M. Bone Marrow Niche-Mediated Survival of Leukemia Stem Cells in Acute Myeloid Leukemia: Yin and Yang. Cancer Biol. Med. 2016, 13, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi, P.; Borsani, O. Targeting Leukemia Stem Cell-Niche Dynamics: A New Challenge in AML Treatment. J. Oncol. 2019, 2019, 8323592. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Aziz, A.M.; Sun, Y.; Hellmich, C.; Marlein, C.R.; Mistry, J.; Forde, E.; Piddock, R.E.; Shafat, M.S.; Morfakis, A.; Mehta, T.; et al. Acute Myeloid Leukemia Induces Protumoral P16INK4a-Driven Senescence in the Bone Marrow Microenvironment. Blood 2019, 133, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Terreros, M.J.; Vernot, J.-P. In Vitro and In Vivo Modeling of Normal and Leukemic Bone Marrow Niches: Cellular Senescence Contribution to Leukemia Induction and Progression. Int. J. Mol. Sci. 2022, 23, 7350. [Google Scholar] [CrossRef] [PubMed]
- Man, C.H.; Fung, T.K.; Ho, C.; Han, H.H.C.; Chow, H.C.H.; Ma, A.C.H.; Choi, W.W.L.; Lok, S.; Cheung, A.M.S.; Eaves, C.; et al. Sorafenib Treatment of FLT3-ITD(+) Acute Myeloid Leukemia: Favorable Initial Outcome and Mechanisms of Subsequent Nonresponsiveness Associated with the Emergence of a D835 Mutation. Blood 2012, 119, 5133–5143. [Google Scholar] [CrossRef]
- Annageldiyev, C.; Gowda, K.; Patel, T.; Bhattacharya, P.; Tan, S.-F.; Iyer, S.; Desai, D.; Dovat, S.; Feith, D.J.; Loughran, T.P.; et al. The Novel Isatin Analog KS99 Targets Stemness Markers in Acute Myeloid Leukemia. Haematologica 2020, 105, 687–696. [Google Scholar] [CrossRef]
- Hoang, V.T.; Buss, E.C.; Wang, W.; Hoffmann, I.; Raffel, S.; Zepeda-Moreno, A.; Baran, N.; Wuchter, P.; Eckstein, V.; Trumpp, A.; et al. The Rarity of ALDH(+) Cells Is the Key to Separation of Normal versus Leukemia Stem Cells by ALDH Activity in AML Patients. Int. J. Cancer 2015, 137, 525–536. [Google Scholar] [CrossRef]
- Blume, R.; Rempel, E.; Manta, L.; Saeed, B.R.; Wang, W.; Raffel, S.; Ermakova, O.; Eckstein, V.; Benes, V.; Trumpp, A.; et al. The Molecular Signature of AML with Increased ALDH Activity Suggests a Stem Cell Origin. Leuk. Lymphoma 2018, 59, 2201–2210. [Google Scholar] [CrossRef]
- Döhner, H.; Estey, E.; Grimwade, D.; Amadori, S.; Appelbaum, F.R.; Büchner, T.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Larson, R.A.; et al. Diagnosis and Management of AML in Adults: 2017 ELN Recommendations from an International Expert Panel. Blood 2017, 129, 424–447. [Google Scholar] [CrossRef]
- Estey, E.H. Acute Myeloid Leukemia: 2019 Update on Risk-Stratification and Management. Am. J. Hematol. 2018, 93, 1267–1291. [Google Scholar] [CrossRef] [PubMed]
- Masciarelli, S.; Capuano, E.; Ottone, T.; Divona, M.; Lavorgna, S.; Liccardo, F.; Śniegocka, M.; Travaglini, S.; Noguera, N.I.; Picardi, A.; et al. Retinoic Acid Synergizes with the Unfolded Protein Response and Oxidative Stress to Induce Cell Death in FLT3-ITD+ AML. Blood Adv. 2019, 3, 4155–4160. [Google Scholar] [CrossRef] [PubMed]
- Poturnajova, M.; Kozovska, Z.; Matuskova, M. Aldehyde Dehydrogenase 1A1 and 1A3 Isoforms—Mechanism of Activation and Regulation in Cancer. Cell. Signal. 2021, 87, 110120. [Google Scholar] [CrossRef] [PubMed]
- Mendez, L.M.; Posey, R.R.; Pandolfi, P.P. The Interplay Between the Genetic and Immune Landscapes of AML: Mechanisms and Implications for Risk Stratification and Therapy. Front. Oncol. 2019, 9, 1162. [Google Scholar] [CrossRef]
- Pallis, M.; Grundy, M.; Turzanski, J.; Kofler, R.; Russell, N. Mitochondrial Membrane Sensitivity to Depolarization in Acute Myeloblastic Leukemia Is Associated with Spontaneous in Vitro Apoptosis, Wild-Type TP53, and Vicinal Thiol/Disulfide Status. Blood 2001, 98, 405–413. [Google Scholar] [CrossRef]
- Saha, S.; Hossain, D.M.S.; Mukherjee, S.; Mohanty, S.; Mazumdar, M.; Mukherjee, S.; Ghosh, U.K.; Nayek, C.; Raveendar, C.; Khurana, A.; et al. Calcarea Carbonica Induces Apoptosis in Cancer Cells in P53-Dependent Manner via an Immuno-Modulatory Circuit. BMC Complement. Altern. Med. 2013, 13, 230. [Google Scholar] [CrossRef]
- Prokocimer, M.; Molchadsky, A.; Rotter, V. Dysfunctional Diversity of P53 Proteins in Adult Acute Myeloid Leukemia: Projections on Diagnostic Workup and Therapy. Blood 2017, 130, 699–712. [Google Scholar] [CrossRef]
- Algariri, E.S.; Mydin, R.B.S.M.N.; Moses, E.J.; Okekpa, S.I.; Rahim, N.A.A.; Yusoff, N.M. Knockdown of Stromal Interaction Molecule 1 (STIM1) Suppresses Acute Myeloblastic Leukemia-M5 Cell Line Survival through Inhibition of Reactive Oxygen Species Activities. Turk. J. Haematol. Off. J. Turk. Soc. Haematol. 2022, 40, 11–17. [Google Scholar] [CrossRef]
- Wang, W.; Liang, Q.; Zhao, J.; Pan, H.; Gao, Z.; Fang, L.; Zhou, Y.; Shi, J. Low Expression of the Metabolism-Related Gene SLC25A21 Predicts Unfavourable Prognosis in Patients with Acute Myeloid Leukaemia. Front. Genet. 2022, 13, 970316. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A. DNA Damage Accumulation and Repair Defects in FLT3-ITD Acute Myeloid Leukemia: Implications for Clonal Evolution and Disease Progression. Hematol. Oncol. 2023, 41, 26–38. [Google Scholar] [CrossRef]
- Kannan, S.; Irwin, M.E.; Herbrich, S.M.; Cheng, T.; Patterson, L.L.; Aitken, M.J.L.; Bhalla, K.; You, M.J.; Konopleva, M.; Zweidler-McKay, P.A.; et al. Targeting the NRF2/HO-1 Antioxidant Pathway in FLT3-ITD-Positive AML Enhances Therapy Efficacy. Antioxidants 2022, 11, 717. [Google Scholar] [CrossRef] [PubMed]
- Panina, S.B.; Pei, J.; Baran, N.; Tjahjono, E.; Patel, S.; Alatrash, G.; Konoplev, S.; Stolbov, L.A.; Poroikov, V.V.; Konopleva, M.; et al. Novel Mitochondria-Targeting Compounds Selectively Kill Human Leukemia Cells. Leukemia 2022, 36, 2009–2021. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Khan, D.H.; Hurren, R.; Xu, M.; Na, Y.; Kang, H.; Mirali, S.; Wang, X.; Gronda, M.; Jitkova, Y.; et al. Venetoclax Enhances T Cell-Mediated Antileukemic Activity by Increasing ROS Production. Blood 2021, 138, 234–245. [Google Scholar] [CrossRef]
- Cai, H.; Kondo, M.; Sandhow, L.; Xiao, P.; Johansson, A.-S.; Sasaki, T.; Zawacka-Pankau, J.; Tryggvason, K.; Ungerstedt, J.; Walfridsson, J.; et al. Critical Role of Lama4 for Hematopoiesis Regeneration and Acute Myeloid Leukemia Progression. Blood 2022, 139, 3040–3057. [Google Scholar] [CrossRef]
- Marlein, C.R.; Zaitseva, L.; Piddock, R.E.; Robinson, S.D.; Edwards, D.R.; Shafat, M.S.; Zhou, Z.; Lawes, M.; Bowles, K.M.; Rushworth, S.A. NADPH Oxidase-2 Derived Superoxide Drives Mitochondrial Transfer from Bone Marrow Stromal Cells to Leukemic Blasts. Blood 2017, 130, 1649–1660. [Google Scholar] [CrossRef]
- Dong, C.; Zhang, N.; Zhang, L. The Multi-Omic Prognostic Model of Oxidative Stress-Related Genes in Acute Myeloid Leukemia. Front. Genet. 2021, 12, 722064. [Google Scholar] [CrossRef]
- Trombetti, S.; Cesaro, E.; Catapano, R.; Sessa, R.; Lo Bianco, A.; Izzo, P.; Grosso, M. Oxidative Stress and ROS-Mediated Signaling in Leukemia: Novel Promising Perspectives to Eradicate Chemoresistant Cells in Myeloid Leukemia. Int. J. Mol. Sci. 2021, 22, 2470. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.-L.; Zhang, W.-G.; Wei, Y.-C.; Meng, S.; Bai, G.-G.; Wang, B.-Y.; Yang, H.-Y.; Tian, W.; Meng, X.; Zhang, H.; et al. Involvement of Oxidative Stress in the Relapse of Acute Myeloid Leukemia. J. Biol. Chem. 2010, 285, 15010–15015. [Google Scholar] [CrossRef]
- Pan, L.; Hao, W.; Zheng, X.; Zeng, X.; Ahmed Abbasi, A.; Boldogh, I.; Ba, X. OGG1-DNA Interactions Facilitate NF-ΚB Binding to DNA Targets. Sci. Rep. 2017, 7, 43297. [Google Scholar] [CrossRef]
- Chen, M.-C.; Baskaran, R.; Lee, N.-H.; Hsu, H.-H.; Ho, T.-J.; Tu, C.-C.; Lin, Y.-M.; Viswanadha, V.P.; Kuo, W.-W.; Huang, C.-Y. CXCL2/CXCR2 Axis Induces Cancer Stem Cell Characteristics in CPT-11-Resistant LoVo Colon Cancer Cells via Gαi-2 and Gαq/11. J. Cell. Physiol. 2019, 234, 11822–11834. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhao, L.; Man, J.; Liu, B. CXCL2 Benefits Acute Myeloid Leukemia Cells in Hypoxia. Int. J. Lab. Hematol. 2021, 43, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wu, S.; Wu, G.; Yang, J. Inhibition of 8-Oxoguanine DNA Glycosylase (OGG1) Expression Suppresses Polycystic Ovarian Syndrome via the NF-ΚB Signaling Pathway. Reprod. Biol. 2022, 22, 100679. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Qi, T.; Pan, L.; Wang, R.; Zhu, B.; Aguilera-Aguirre, L.; Radak, Z.; Hazra, T.K.; Vlahopoulos, S.A.; Bacsi, A.; et al. Effects of the Stimuli-Dependent Enrichment of 8-Oxoguanine DNA Glycosylase1 on Chromatinized DNA. Redox Biol. 2018, 18, 43–53. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, K.; Pan, L.; Hao, W.; Xue, Y.; Bacsi, A.; Vlahopoulos, S.A.; Radak, Z.; Hazra, T.K.; Brasier, A.R.; et al. Innate Immune Responses to RSV Infection Facilitated by OGG1, an Enzyme Repairing Oxidatively Modified DNA Base Lesions. J. Innate Immun. 2022, 14, 594–615. [Google Scholar] [CrossRef]
- Hao, W.; Wang, J.; Zhang, Y.; Wang, C.; Xia, L.; Zhang, W.; Zafar, M.; Kang, J.-Y.; Wang, R.; Ali Bohio, A.; et al. Enzymatically Inactive OGG1 Binds to DNA and Steers Base Excision Repair toward Gene Transcription. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 7427–7441. [Google Scholar] [CrossRef]
- Vlahopoulos, S.; Adamaki, M.; Khoury, N.; Zoumpourlis, V.; Boldogh, I. Roles of DNA Repair Enzyme OGG1 in Innate Immunity and Its Significance for Lung Cancer. Pharmacol. Ther. 2019, 194, 59–72. [Google Scholar] [CrossRef]
- Lei, H.-M.; Zhang, K.-R.; Wang, C.H.; Wang, Y.; Zhuang, G.-L.; Lu, L.-M.; Zhang, J.; Shen, Y.; Chen, H.-Z.; Zhu, L. Aldehyde Dehydrogenase 1A1 Confers Erlotinib Resistance via Facilitating the Reactive Oxygen Species-Reactive Carbonyl Species Metabolic Pathway in Lung Adenocarcinomas. Theranostics 2019, 9, 7122–7139. [Google Scholar] [CrossRef]
- Mihatsch, J.; Toulany, M.; Bareiss, P.M.; Grimm, S.; Lengerke, C.; Kehlbach, R.; Rodemann, H.P. Selection of Radioresistant Tumor Cells and Presence of ALDH1 Activity in Vitro. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2011, 99, 300–306. [Google Scholar] [CrossRef]
- Allison Stewart, C.; Tong, P.; Cardnell, R.J.; Sen, T.; Li, L.; Gay, C.M.; Masrorpour, F.; Fan, Y.; Bara, R.O.; Feng, Y.; et al. Dynamic Variations in Epithelial-to-Mesenchymal Transition (EMT), ATM, and SLFN11 Govern Response to PARP Inhibitors and Cisplatin in Small Cell Lung Cancer. Oncotarget 2017, 8, 28575–28587. [Google Scholar] [CrossRef]
- Liu, C.; Qiang, J.; Deng, Q.; Xia, J.; Deng, L.; Zhou, L.; Wang, D.; He, X.; Liu, Y.; Zhao, B.; et al. ALDH1A1 Activity in Tumor-Initiating Cells Remodels Myeloid-Derived Suppressor Cells to Promote Breast Cancer Progression. Cancer Res. 2021, 81, 5919–5934. [Google Scholar] [CrossRef]
- Owen, N.; Minko, I.G.; Moellmer, S.A.; Cammann, S.K.; Lloyd, R.S.; McCullough, A.K. Enhanced Cytarabine-Induced Killing in OGG1-Deficient Acute Myeloid Leukemia Cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2016833118. [Google Scholar] [CrossRef] [PubMed]
- Liddiard, K.; Hills, R.; Burnett, A.K.; Darley, R.L.; Tonks, A. OGG1 Is a Novel Prognostic Indicator in Acute Myeloid Leukaemia. Oncogene 2010, 29, 2005–2012. [Google Scholar] [CrossRef]
- Gotoh, N.; Saitoh, T.; Takahashi, N.; Kasamatsu, T.; Minato, Y.; Lobna, A.; Oda, T.; Hoshino, T.; Sakura, T.; Shimizu, H.; et al. Association between OGG1 S326C CC Genotype and Elevated Relapse Risk in Acute Myeloid Leukemia. Int. J. Hematol. 2018, 108, 246–253. [Google Scholar] [CrossRef]
- Liu, L.; Cai, S.; Han, C.; Banerjee, A.; Wu, D.; Cui, T.; Xie, G.; Zhang, J.; Zhang, X.; McLaughlin, E.; et al. ALDH1A1 Contributes to PARP Inhibitor Resistance via Enhancing DNA Repair in BRCA2-/- Ovarian Cancer Cells. Mol. Cancer Ther. 2020, 19, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.; Deshpande, M.; Pang, H.; Stemmer, P.M.; Carruthers, N.J.; Shearn, C.T.; Backos, D.S.; Palaniyandi, S.S. 4-Hydroxy-2-Nonenal Attenuates 8-Oxoguanine DNA Glycosylase 1 Activity. J. Cell. Biochem. 2020, 121, 4887–4897. [Google Scholar] [CrossRef]
- Smoak, K.A.; Cidlowski, J.A. Mechanisms of Glucocorticoid Receptor Signaling during Inflammation. Mech. Ageing Dev. 2004, 125, 697–706. [Google Scholar] [CrossRef]
- Bewersdorf, J.P.; Zeidan, A.M. Hyperleukocytosis and Leukostasis in Acute Myeloid Leukemia: Can a Better Understanding of the Underlying Molecular Pathophysiology Lead to Novel Treatments? Cells 2020, 9, 2310. [Google Scholar] [CrossRef] [PubMed]
- Cerrano, M.; Chevret, S.; Raffoux, E.; Rabian, F.; Sébert, M.; Valade, S.; Itzykson, R.; Lemiale, V.; Adès, L.; Boissel, N.; et al. Benefits of Dexamethasone on Early Outcomes in Patients with Acute Myeloid Leukemia with Hyperleukocytosis: A Propensity Score Matched Analysis. Ann. Hematol. 2023, 102, 761–768. [Google Scholar] [CrossRef]
- Gutjahr, J.C.; Bayer, E.; Yu, X.; Laufer, J.M.; Höpner, J.P.; Tesanovic, S.; Härzschel, A.; Auer, G.; Rieß, T.; Salmhofer, A.; et al. CD44 Engagement Enhances Acute Myeloid Leukemia Cell Adhesion to the Bone Marrow Microenvironment by Increasing VLA-4 Avidity. Haematologica 2021, 106, 2102–2113. [Google Scholar] [CrossRef]
- Walker, A.R.; Wang, H.; Walsh, K.; Bhatnagar, B.; Vasu, S.; Garzon, R.; Canning, R.; Geyer, S.; Wu, Y.-Z.; Devine, S.M.; et al. Midostaurin, Bortezomib and MEC in Relapsed/Refractory Acute Myeloid Leukemia. Leuk. Lymphoma 2016, 57, 2100–2108. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Liu, Y.; Pang, Y.; Qian, T.; Quan, L.; Cheng, Z.; Dai, Y.; Ye, X.; Pang, Y.; Shi, J.; et al. Emerging Agents and Regimens for Treatment of Relapsed and Refractory Acute Myeloid Leukemia. Cancer Gene Ther. 2020, 27, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Döhner, H.; Wei, A.H.; Appelbaum, F.R.; Craddock, C.; DiNardo, C.D.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Godley, L.A.; Hasserjian, R.P.; et al. Diagnosis and Management of AML in Adults: 2022 Recommendations from an International Expert Panel on Behalf of the ELN. Blood 2022, 140, 1345–1377. [Google Scholar] [CrossRef]
- Wei, A.H.; Montesinos, P.; Ivanov, V.; DiNardo, C.D.; Novak, J.; Laribi, K.; Kim, I.; Stevens, D.A.; Fiedler, W.; Pagoni, M.; et al. Venetoclax plus LDAC for Newly Diagnosed AML Ineligible for Intensive Chemotherapy: A Phase 3 Randomized Placebo-Controlled Trial. Blood 2020, 135, 2137–2145. [Google Scholar] [CrossRef]
- Csizmar, C.M.; Kim, D.-H.; Sachs, Z. The Role of the Proteasome in AML. Blood Cancer J. 2016, 6, e503. [Google Scholar] [CrossRef] [PubMed]
- DiNardo, C.D.; Tiong, I.S.; Quaglieri, A.; MacRaild, S.; Loghavi, S.; Brown, F.C.; Thijssen, R.; Pomilio, G.; Ivey, A.; Salmon, J.M.; et al. Molecular Patterns of Response and Treatment Failure after Frontline Venetoclax Combinations in Older Patients with AML. Blood 2020, 135, 791–803. [Google Scholar] [CrossRef]
- Lachowiez, C.; DiNardo, C.D.; Konopleva, M. Venetoclax in Acute Myeloid Leukemia—Current and Future Directions. Leuk. Lymphoma 2020, 61, 1313–1322. [Google Scholar] [CrossRef] [PubMed]
- Schimmer, A.D.; Raza, A.; Carter, T.H.; Claxton, D.; Erba, H.; DeAngelo, D.J.; Tallman, M.S.; Goard, C.; Borthakur, G. A Multicenter Phase I/II Study of Obatoclax Mesylate Administered as a 3- or 24-Hour Infusion in Older Patients with Previously Untreated Acute Myeloid Leukemia. PLoS ONE 2014, 9, e108694. [Google Scholar] [CrossRef]
- Rahmani, M.; Aust, M.M.; Attkisson, E.; Williams, D.C., Jr.; Ferreira-Gonzalez, A.; Grant, S. Inhibition of Bcl-2 Antiapoptotic Members by Obatoclax Potently Enhances Sorafenib-Induced Apoptosis in Human Myeloid Leukemia Cells through a Bim-Dependent Process. Blood 2012, 119, 6089–6098. [Google Scholar] [CrossRef]
- Koschade, S.E.; Klann, K.; Shaid, S.; Vick, B.; Stratmann, J.A.; Thölken, M.; Meyer, L.M.; Nguyen, T.D.; Campe, J.; Moser, L.M.; et al. Translatome Proteomics Identifies Autophagy as a Resistance Mechanism to On-Target FLT3 Inhibitors in Acute Myeloid Leukemia. Leukemia 2022, 36, 2396–2407. [Google Scholar] [CrossRef]
- Zalpoor, H.; Bakhtiyari, M.; Akbari, A.; Aziziyan, F.; Shapourian, H.; Liaghat, M.; Zare-Badie, Z.; Yahyazadeh, S.; Tarhriz, V.; Ganjalikhani-Hakemi, M. Potential Role of Autophagy Induced by FLT3-ITD and Acid Ceramidase in Acute Myeloid Leukemia Chemo-Resistance: New Insights. Cell Commun. Signal. CCS 2022, 20, 172. [Google Scholar] [CrossRef] [PubMed]
- Joffre, C.; Ducau, C.; Poillet-Perez, L.; Courdy, C.; Mansat-De Mas, V. Autophagy a Close Relative of AML Biology. Biology 2021, 10, 552. [Google Scholar] [CrossRef]
- Folkerts, H.; Wierenga, A.T.; van den Heuvel, F.A.; Woldhuis, R.R.; Kluit, D.S.; Jaques, J.; Schuringa, J.J.; Vellenga, E. Elevated VMP1 Expression in Acute Myeloid Leukemia Amplifies Autophagy and Is Protective against Venetoclax-Induced Apoptosis. Cell Death Dis. 2019, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Varisli, L.; Cen, O.; Vlahopoulos, S. Dissecting Pharmacological Effects of Chloroquine in Cancer Treatment: Interference with Inflammatory Signaling Pathways. Immunology 2020, 159, 257–278. [Google Scholar] [CrossRef]
- Grønningsæter, I.S.; Reikvam, H.; Aasebø, E.; Bartaula-Brevik, S.; Hernandez-Valladares, M.; Selheim, F.; Berven, F.S.; Tvedt, T.H.; Bruserud, Ø.; Hatfield, K.J. Effects of the Autophagy-Inhibiting Agent Chloroquine on Acute Myeloid Leukemia Cells; Characterization of Patient Heterogeneity. J. Pers. Med. 2021, 11, 779. [Google Scholar] [CrossRef]
- Cuesta-Casanovas, L.; Delgado-Martínez, J.; Cornet-Masana, J.M.; Carbó, J.M.; Clément-Demange, L.; Risueño, R.M. Lysosome-Mediated Chemoresistance in Acute Myeloid Leukemia. Cancer Drug Resist. 2022, 5, 233–244. [Google Scholar] [CrossRef]
- Piao, S.; Ojha, R.; Rebecca, V.W.; Samanta, A.; Ma, X.-H.; Mcafee, Q.; Nicastri, M.C.; Buckley, M.; Brown, E.; Winkler, J.D.; et al. ALDH1A1 and HLTF Modulate the Activity of Lysosomal Autophagy Inhibitors in Cancer Cells. Autophagy 2017, 13, 2056–2071. [Google Scholar] [CrossRef]
- Cheng, C.K.; Chan, N.P.H.; Wan, T.S.K.; Lam, L.Y.; Cheung, C.H.Y.; Wong, T.H.Y.; Ip, R.K.L.; Wong, R.S.M.; Ng, M.H.L. Helicase-like Transcription Factor Is a RUNX1 Target Whose Downregulation Promotes Genomic Instability and Correlates with Complex Cytogenetic Features in Acute Myeloid Leukemia. Haematologica 2016, 101, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Krohne, T.U.; Kaemmerer, E.; Holz, F.G.; Kopitz, J. Lipid Peroxidation Products Reduce Lysosomal Protease Activities in Human Retinal Pigment Epithelial Cells via Two Different Mechanisms of Action. Exp. Eye Res. 2010, 90, 261–266. [Google Scholar] [CrossRef]
- Zhong, F.-M.; Yao, F.-Y.; Liu, J.; Zhang, H.-B.; Zhang, J.; Zhang, N.; Lin, J.; Li, S.-Q.; Li, M.-Y.; Jiang, J.-Y.; et al. Ferroptosis-Related Molecular Patterns Reveal Immune Escape, Inflammatory Development and Lipid Metabolism Characteristics of the Tumor Microenvironment in Acute Myeloid Leukemia. Front. Oncol. 2022, 12, 888570. [Google Scholar] [CrossRef]
- Wu, Y.; Kram, H.; Gempt, J.; Liesche-Starnecker, F.; Wu, W.; Schlegel, J. ALDH1-Mediated Autophagy Sensitizes Glioblastoma Cells to Ferroptosis. Cells 2022, 11, 4015. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Pan, C.; Zhang, X.; Yang, T.; Hu, T.; Zheng, L.; Cao, S.; Feng, C.; Hu, X.; Chai, X.; et al. Nuclear Factor Nrf2 Promotes Glycosidase OGG1 Expression by Activating the AKT Pathway to Enhance Leukemia Cell Resistance to Cytarabine. J. Biol. Chem. 2022, 299, 102798. [Google Scholar] [CrossRef] [PubMed]
- Bruserud, Ø.; Tsykunova, G.; Hernandez-Valladares, M.; Reikvam, H.; Tvedt, T.H.A. Therapeutic Use of Valproic Acid and All-Trans Retinoic Acid in Acute Myeloid Leukemia-Literature Review and Discussion of Possible Use in Relapse after Allogeneic Stem Cell Transplantation. Pharmaceuticals 2021, 14, 423. [Google Scholar] [CrossRef] [PubMed]
- Elizondo, G.; Corchero, J.; Sterneck, E.; Gonzalez, F.J. Feedback Inhibition of the Retinaldehyde Dehydrogenase Gene ALDH1 by Retinoic Acid through Retinoic Acid Receptor Alpha and CCAAT/Enhancer-Binding Protein Beta. J. Biol. Chem. 2000, 275, 39747–39753. [Google Scholar] [CrossRef] [PubMed]
- Caminear, M.W.; Harrington, B.S.; Kamdar, R.D.; Kruhlak, M.J.; Annunziata, C.M. Disulfiram Transcends ALDH Inhibitory Activity When Targeting Ovarian Cancer Tumor-Initiating Cells. Front. Oncol. 2022, 12, 762820. [Google Scholar] [CrossRef]
- Yasgar, A.; Titus, S.A.; Wang, Y.; Danchik, C.; Yang, S.-M.; Vasiliou, V.; Jadhav, A.; Maloney, D.J.; Simeonov, A.; Martinez, N.J. A High-Content Assay Enables the Automated Screening and Identification of Small Molecules with Specific ALDH1A1-Inhibitory Activity. PLoS ONE 2017, 12, e0170937. [Google Scholar] [CrossRef]
- Bista, R.; Lee, D.W.; Pepper, O.B.; Azorsa, D.O.; Arceci, R.J.; Aleem, E. Disulfiram Overcomes Bortezomib and Cytarabine Resistance in Down-Syndrome-Associated Acute Myeloid Leukemia Cells. J. Exp. Clin. Cancer Res. CR 2017, 36, 22. [Google Scholar] [CrossRef]
- Conticello, C.; Martinetti, D.; Adamo, L.; Buccheri, S.; Giuffrida, R.; Parrinello, N.; Lombardo, L.; Anastasi, G.; Amato, G.; Cavalli, M.; et al. Disulfiram, an Old Drug with New Potential Therapeutic Uses for Human Hematological Malignancies. Int. J. Cancer 2012, 131, 2197–2203. [Google Scholar] [CrossRef]
- Hassani, S.; Ghaffari, P.; Chahardouli, B.; Alimoghaddam, K.; Ghavamzadeh, A.; Alizadeh, S.; Ghaffari, S.H. Disulfiram/Copper Causes ROS Levels Alteration, Cell Cycle Inhibition, and Apoptosis in Acute Myeloid Leukaemia Cell Lines with Modulation in the Expression of Related Genes. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 99, 561–569. [Google Scholar] [CrossRef]
- Yang, W.; Xie, J.; Hou, R.; Chen, X.; Xu, Z.; Tan, Y.; Ren, F.; Zhang, Y.; Xu, J.; Chang, J.; et al. Disulfiram/Cytarabine Eradicates a Subset of Acute Myeloid Leukemia Stem Cells with High Aldehyde Dehydrogenase Expression. Leuk. Res. 2020, 92, 106351. [Google Scholar] [CrossRef]
- Nechushtan, H.; Hamamreh, Y.; Nidal, S.; Gotfried, M.; Baron, A.; Shalev, Y.I.; Nisman, B.; Peretz, T.; Peylan-Ramu, N. A Phase IIb Trial Assessing the Addition of Disulfiram to Chemotherapy for the Treatment of Metastatic Non-Small Cell Lung Cancer. Oncologist 2015, 20, 366–367. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Bhagwati, S.; Siddiqi, M.I.; Chattopadhyay, N. A Critical Assessment of the Potential of Pharmacological Modulation of Aldehyde Dehydrogenases to Treat the Diseases of Bone Loss. Eur. J. Pharmacol. 2020, 886, 173541. [Google Scholar] [CrossRef] [PubMed]
- Saracino, M.A.; Marcheselli, C.; Somaini, L.; Gerra, G.; De Stefano, F.; Pieri, M.C.; Raggi, M.A. Simultaneous Determination of Disulfiram and Bupropion in Human Plasma of Alcohol and Nicotine Abusers. Anal. Bioanal. Chem. 2010, 398, 2155–2161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jiang, Y.; Jing, G.; Tang, Y.; Chen, X.; Yang, D.; Zhang, Y.; Tang, X. A Novel UPLC-ESI-MS/MS Method for the Quantitation of Disulfiram, Its Role in Stabilized Plasma and Its Application. J. Chromatogr. B Analyt. Technol. Biomed. Life. Sci. 2013, 937, 54–59. [Google Scholar] [CrossRef]
- Lee, S.A.; Elliott, J.H.; McMahon, J.; Hartogenesis, W.; Bumpus, N.N.; Lifson, J.D.; Gorelick, R.J.; Bacchetti, P.; Deeks, S.G.; Lewin, S.R.; et al. Population Pharmacokinetics and Pharmacodynamics of Disulfiram on Inducing Latent HIV-1 Transcription in a Phase IIb Trial. Clin. Pharmacol. Ther. 2019, 105, 692–702. [Google Scholar] [CrossRef]
- Elliott, J.H.; McMahon, J.H.; Chang, C.C.; Lee, S.A.; Hartogensis, W.; Bumpus, N.; Savic, R.; Roney, J.; Hoh, R.; Solomon, A.; et al. Short-Term Administration of Disulfiram for Reversal of Latent HIV Infection: A Phase 2 Dose-Escalation Study. Lancet HIV 2015, 2, e520–e529. [Google Scholar] [CrossRef]
- Spivak, A.M.; Andrade, A.; Eisele, E.; Hoh, R.; Bacchetti, P.; Bumpus, N.N.; Emad, F.; Buckheit, R.; McCance-Katz, E.F.; Lai, J.; et al. A Pilot Study Assessing the Safety and Latency-Reversing Activity of Disulfiram in HIV-1-Infected Adults on Antiretroviral Therapy. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2014, 58, 883–890. [Google Scholar] [CrossRef]
- Li, H.; Liu, B.; Ao, H.; Fu, J.; Wang, Y.; Feng, Y.; Guo, Y.; Wang, X. Soybean Lecithin Stabilizes Disulfiram Nanosuspensions with a High Drug-Loading Content: Remarkably Improved Antitumor Efficacy. J. Nanobiotechnol. 2020, 18, 4. [Google Scholar] [CrossRef]
- Ullah, A.; Ullah, N.; Nawaz, T.; Aziz, T. Molecular Mechanisms of Sanguinarine in Cancer Prevention and Treatment. Anticancer Agents Med. Chem. 2023, 23, 765–778. [Google Scholar] [CrossRef]
- Ullah, A.; Leong, S.W.; Wang, J.; Wu, Q.; Ghauri, M.A.; Sarwar, A.; Su, Q.; Zhang, Y. Cephalomannine Inhibits Hypoxia-Induced Cellular Function via the Suppression of APEX1/HIF-1α Interaction in Lung Cancer. Cell Death Dis. 2021, 12, 490. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dancik, G.M.; Varisli, L.; Vlahopoulos, S.A. The Molecular Context of Oxidant Stress Response in Cancer Establishes ALDH1A1 as a Critical Target: What This Means for Acute Myeloid Leukemia. Int. J. Mol. Sci. 2023, 24, 9372. https://doi.org/10.3390/ijms24119372
Dancik GM, Varisli L, Vlahopoulos SA. The Molecular Context of Oxidant Stress Response in Cancer Establishes ALDH1A1 as a Critical Target: What This Means for Acute Myeloid Leukemia. International Journal of Molecular Sciences. 2023; 24(11):9372. https://doi.org/10.3390/ijms24119372
Chicago/Turabian StyleDancik, Garrett M., Lokman Varisli, and Spiros A. Vlahopoulos. 2023. "The Molecular Context of Oxidant Stress Response in Cancer Establishes ALDH1A1 as a Critical Target: What This Means for Acute Myeloid Leukemia" International Journal of Molecular Sciences 24, no. 11: 9372. https://doi.org/10.3390/ijms24119372