
Micro-Current Stimulation Has Potential Effects of Hair Growth-Promotion on Human Hair Follicle-Derived Papilla Cells and Animal Model

 , , and
, , and
Abstract
:1. Introduction
2. Results
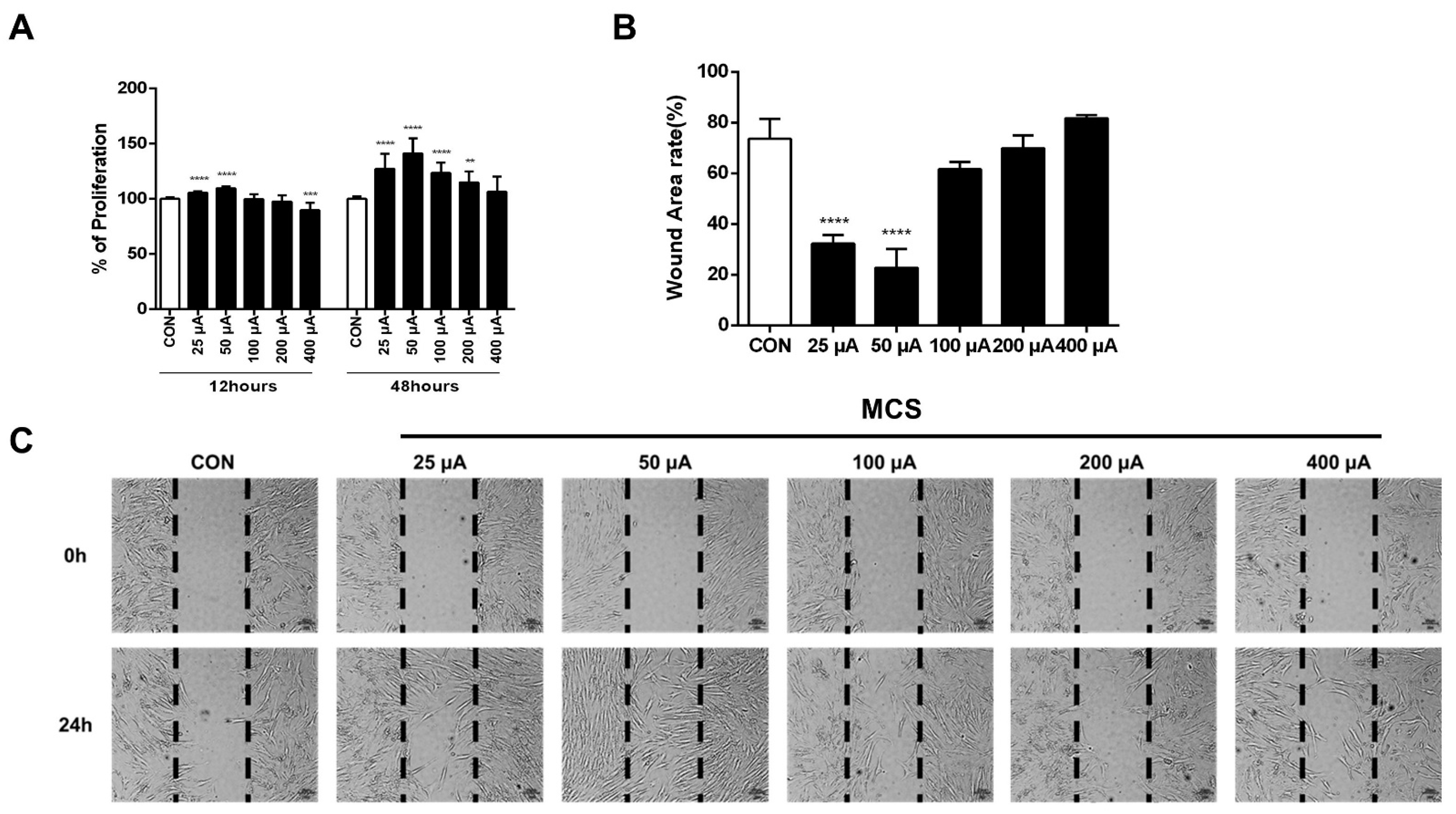
2.1. MCS Stimulated HFDPC Cell Proliferation and Migration
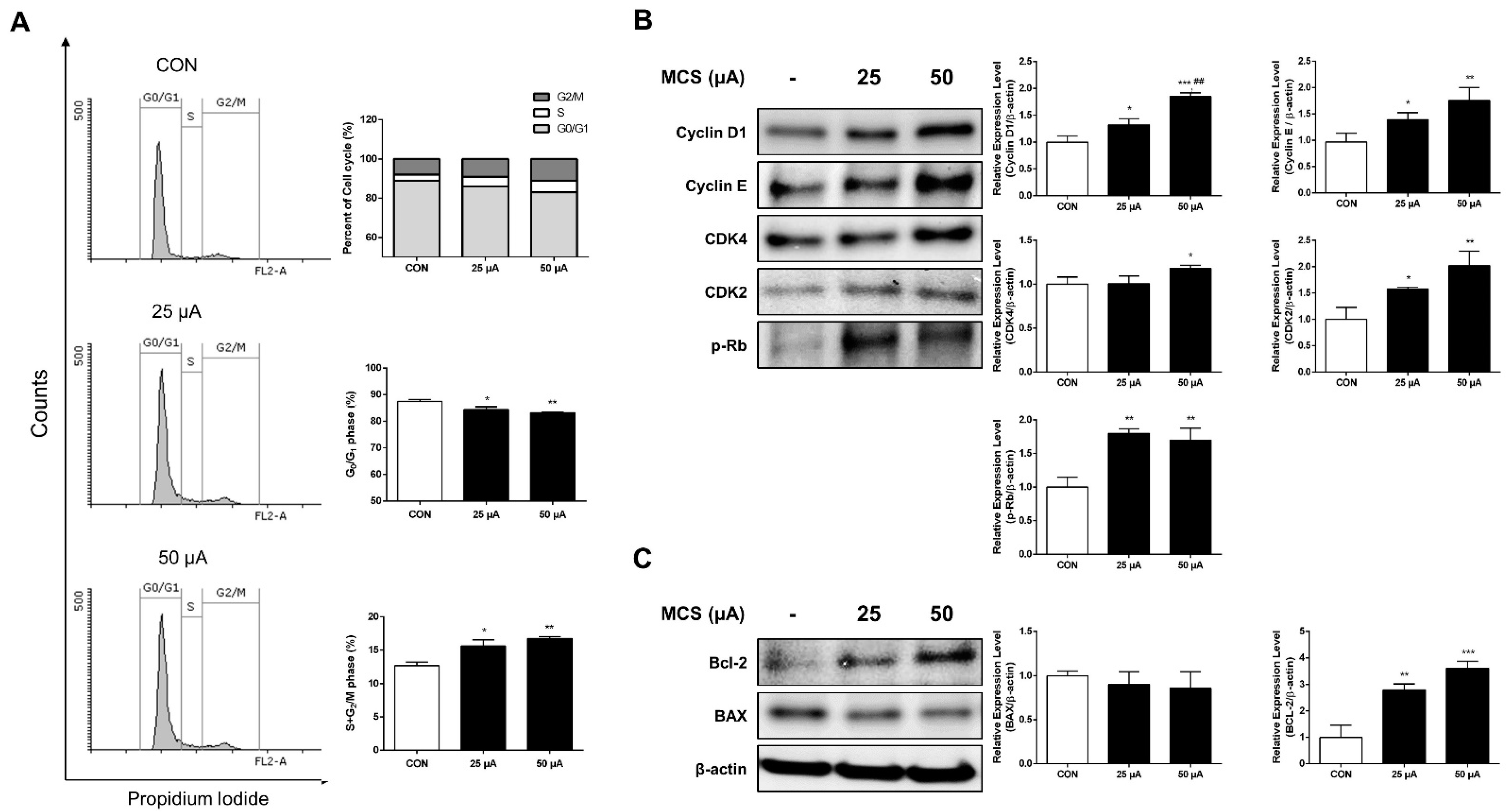
2.2. MCS Enhanced Cell Cycle Progression
2.3. Effect of MCS on G1 Phase Related Protein and Apoptosis Related Protein Expression
2.4. MCS-Induced Activation of the PI3K/AKT/mTOR/Fox01 Signaling Pathway and Wnt Pathway Affects Upregulation of β-Catenin Expression
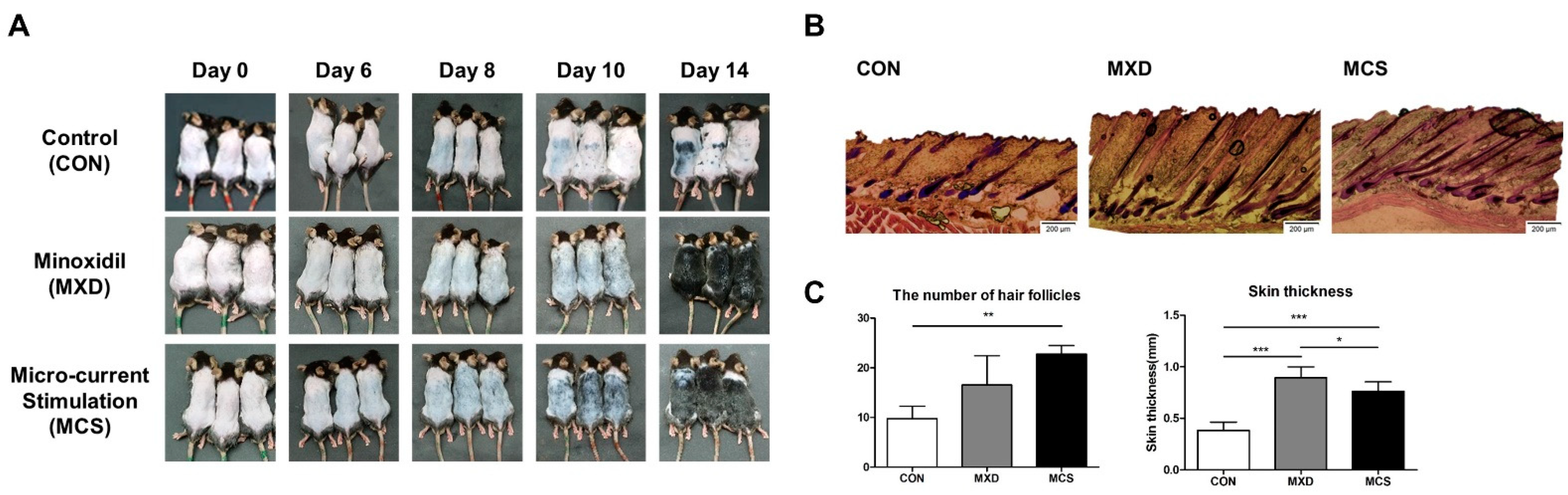
2.5. The Effect of MCS on Hair Growth in the Telogenic C57BL/6 Mice
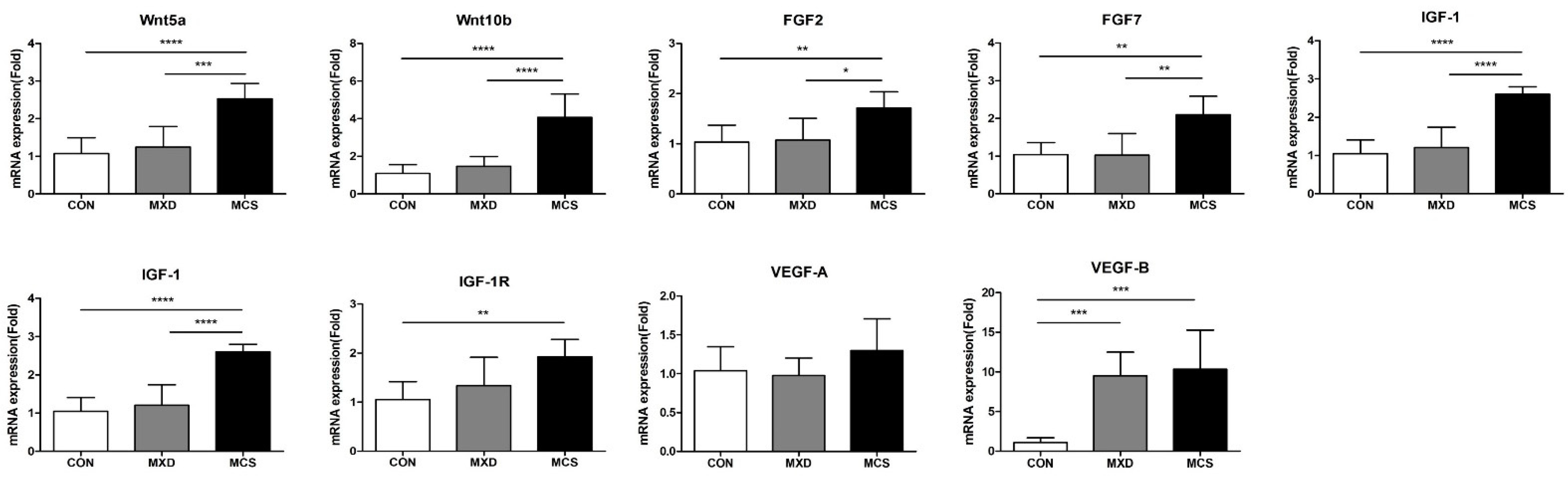
2.6. The mRNA Expression Level of Hair Growth Promotion Associated Genes in Mouse Skin Tissue
3. Discussion
4. Materials and Methods
4.1. Materials
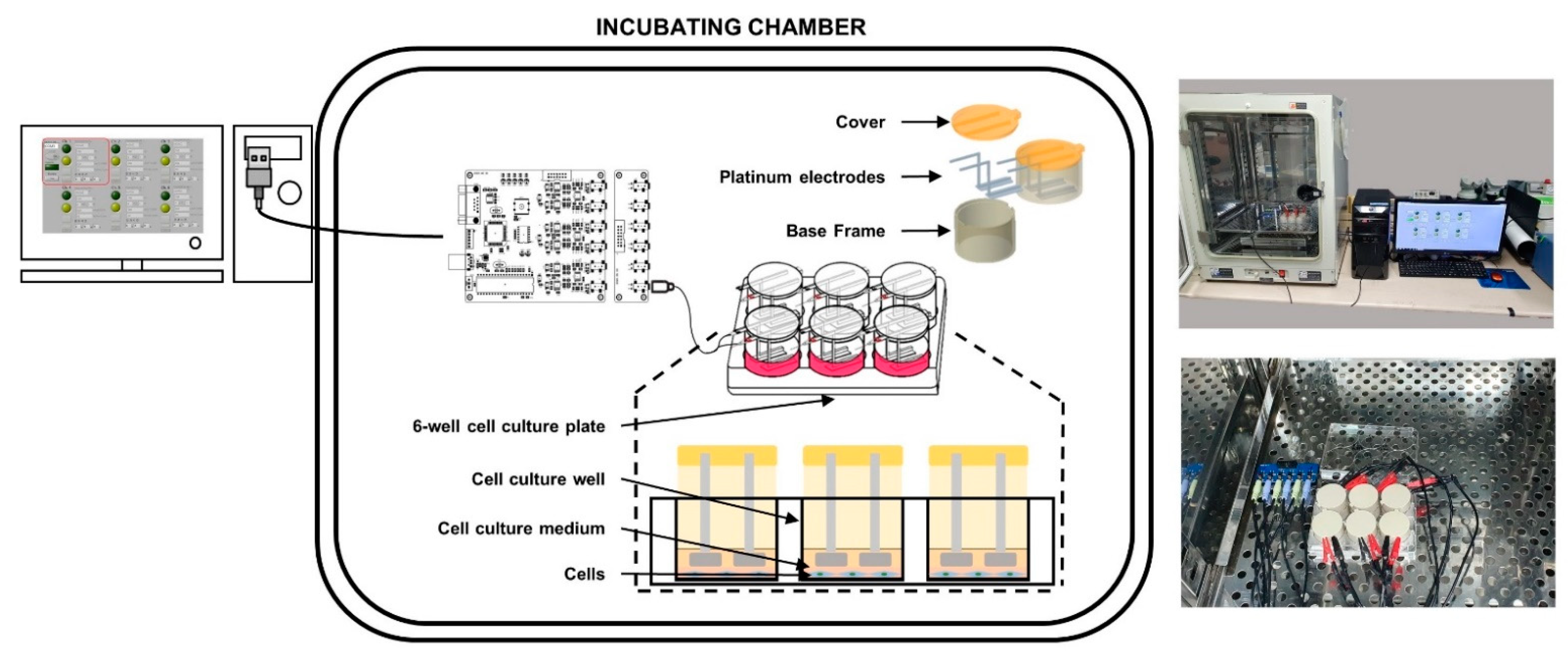
4.2. Design and Implementation of the Custom-Made Microcurrent Stimulating Chanmber System
4.3. Application of MCS for In-Vitro and In-Vivo Experiment
4.4. HFDPC Cell Culture
4.5. Cell Viability
4.6. Cell Migration Assay
4.7. Flow Cytometry Analysis
4.8. Immunoblotting
4.9. Animals
4.10. Histologic Evaluation
4.11. Real-Time Reverse Transcription PCR (qRT-PCR)
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lei, M.; Chuong, C.-M. Aging, alopecia, and stem cells. Science 2016, 351, 559–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, H.; Mohri, Y.; Binh, N.T.; Morinaga, H.; Fukuda, M.; Ito, M.; Nishimura, E.K. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 proteolysis. Science 2016, 351, 6273. [Google Scholar] [CrossRef] [PubMed]
- Price, V.H. Treatment of Hair Loss. N. Engl. J. Med. 1999, 341, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Mounsey, A.L.; Reed, S.W. Diagnosing and treating hair loss. Am. Fam. Physician 2009, 80, 356–362. [Google Scholar] [PubMed]
- Sasson, M.; Shupack, J.L.; Stiller, M.J. status of medical treatment for androgenetic alopecia. Int. J. Dermatol. 1993, 32, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J. Hair Loss in Women. N. Engl. J. Med. 2007, 357, 1620–1630. [Google Scholar] [CrossRef] [Green Version]
- Rogers, N.E.; Avram, M.R. Medical treatments for male and female pattern hair loss. J. Am. Acad. Dermatol. 2008, 59, 547–566. [Google Scholar] [CrossRef]
- Bernstein, E.F. Hair growth induced by diode laser treatment. Dermatol. Surg. 2005, 31, 584–586. [Google Scholar] [CrossRef]
- Lin, T.-Y.D.; Manuskiatti, W.; Dierickx, C.C.; Farinelli, W.A.; Fisher, M.E.; Flotte, T.; Baden, H.P.; Anderson, R.R. Hair Growth Cycle Affects Hair Follicle Destruction by Ruby Laser Pulses. J. Investig. Dermatol. 1998, 111, 107–113. [Google Scholar] [CrossRef]
- Benjamin, B.; Ziginskas, D.; Harman, J.; Meakin, T. Pulsed electrostatic fields (ETG) to reduce hair loss in women undergoing chemotherapy for breast carcinoma: A pilot study. Psycho-Oncology 2002, 11, 244–248. [Google Scholar] [CrossRef]
- Sohn, K.M.; Jeong, K.H.; Park, Y.M.; Kim, J.E.; Kang, H. Hair growth-promotion effects of different alternating current parameter settings are mediated by the activation of Wnt/β-catenin and MAPK pathway. Exp. Dermatol. 2015, 24, 958–963. [Google Scholar] [CrossRef] [PubMed]
- Ngok Cheng, M.D.; van Hoof, H.A.; Bockx, E.; Hoogmartens, M.J.; Mulier, J.C.; de Ducker, F.J.; Sansen, W.M.; de Loecker, W.I. The effects of electric currents on ATP generation, protein synthesis, and membrane transport in rat skin. Clin. Orthop. Relat. Res. 1982, 171, 264–272. [Google Scholar]
- Bureau, J.P.; Ginouves, P.; Guilbaud, J.; Roux, M.E. Essential oils and low-intensity electromagnetic pulses in the treatment of androgen-dependent alopecia. Adv. Ther. 2003, 20, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowski, T. Hair loss as a consequence of cancer chemotherapy—Physical methods of prevention. A review of the literature. Współczesna Onkol. 2011, 2, 95–101. [Google Scholar] [CrossRef]
- Chapman-Jones, D.; Hill, D. Novel microcurrent treatment is more effective than conventional therapy for chronic Achilles tendinopathy: Randomised comparative trial. Physiotherapy 2002, 88, 471–480. [Google Scholar] [CrossRef]
- Cho, M.-S.; Park, R.-J.; Park, S.H.; Cho, Y.-H.; Cheng, G.A. The Effect of Microcurrent-Inducing Shoes on Fatigue and Pain in Middle-Aged People with Plantar Fascitis. J. Phys. Ther. Sci. 2007, 19, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Poltawski, L.; Watson, T. Bioelectricity and microcurrent therapy for tissue healing–a narrative review. Phys. Ther. Rev. 2009, 14, 104–114. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-M.; Li, Y.; Ji, Y.-C.; Keng, H.; Cai, X.-N.; Zhang, J.-K. Microencapsulated human hair dermal papilla cells: A substitute for dermal papilla? Arch. Dermatol. Res. 2008, 300, 531–535. [Google Scholar] [CrossRef]
- Elliott, K.; Messenger, A.G.; Stephenson, T.J. Differences in Hair Follicle Dermal Papilla Volume are Due to Extracellular Matrix Volume and Cell Number: Implications for the Control of Hair Follicle Size and Androgen Responses. J. Investig. Dermatol. 1999, 113, 873–877. [Google Scholar] [CrossRef] [Green Version]
- Stenn, K.S.; Paus, R. Controls of Hair Follicle Cycling. Physiol. Rev. 2001, 81, 449–494. [Google Scholar] [CrossRef]
- Madaan, A.; Verma, R.; Singh, A.T.; Jaggi, M. Review of Hair Follicle Dermal Papilla cells as in vitro screening model for hair growth. Int. J. Cosmet. Sci. 2018, 40, 429–450. [Google Scholar] [CrossRef] [Green Version]
- Kwack, M.H.; Kang, B.M.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Minoxidil activates β-catenin pathway in human dermal papilla cells: A possible explanation for its anagen prolongation effect. J. Dermatol. Sci. 2011, 62, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-I.; Yoo, E.-S.; Hyun, J.-W.; Koh, Y.-S.; Lee, N.H.; Ko, M.-H.; Ko, C.-S.; Kang, H.-K. Promotion Effect of Apo-9′-fucoxanthinone from Sargassum muticum on Hair Growth via the Activation of Wnt/β-Catenin and VEGF-R. Biol. Pharm. Bull. 2016, 39, 1273–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greco, V.; Chen, T.; Rendl, M.; Schober, M.; Pasolli, H.A.; Stokes, N.; Cruz-Racelis, J.D.; Fuchs, E. A Two-Step Mechanism for Stem Cell Activation during Hair Regeneration. Cell Stem Cell 2009, 4, 155–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittaker, S.R.; Mallinger, A.; Workman, P.; Clarke, P.A. Inhibitors of cyclin-dependent kinases as cancer therapeutics. Pharmacol. Ther. 2017, 173, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Sherr, C.J.; Roberts, J.M. CDK inhibitors: Positive and negative regulators of G1-phase progression. Genes Dev. 1999, 13, 1501–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, Y.-C.; Pasolli, H.A.; Fuchs, E. Dynamics between Stem Cells, Niche, and Progeny in the Hair Follicle. Cell 2011, 144, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, G.; Jiang, D.; Li, J.; Kang, L.; Chen, S.; Long, Y.; Wang, Y.; Huang, P.; Lin, Y.; Cai, W.; et al. Self-Activated Electrical Stimulation for Effective Hair Regeneration via a Wearable Omnidirectional Pulse Generator. ACS Nano 2019, 13, 12345–12356. [Google Scholar] [CrossRef]
- Nuccitelli, R.; Lui, K.; Kreis, M.; Athos, B.; Nuccitelli, P. Nanosecond pulsed electric field stimulation of reactive oxygen species in human pancreatic cancer cells is Ca2+-dependent. Biochem. Biophys. Res. Commun. 2013, 435, 580–585. [Google Scholar] [CrossRef] [Green Version]
- Tzoneva, R. Influence of electric field on cell behavior. Electrotreatment of cells for biomedical applications. Asian J. Phys. 2014, 23, 789–814. [Google Scholar]
- Kim, J.; Yang, H.J.; Cho, T.H.; Lee, S.E.; Park, Y.D.; Kim, H.M.; Kim, I.S.; Seo, Y.-K.; Hwang, S.J.; Kim, S.J. Enhanced regeneration of rabbit mandibular defects through a combined treatment of electrical stimulation and rhBMP-2 application. Med. Biol. Eng. Comput. 2013, 51, 1339–1348. [Google Scholar] [CrossRef]
- Paus, R.; Foitzik, K. In search of the “hair cycle clock”: A guided tour. Differentiation 2004, 72, 489–511. [Google Scholar] [CrossRef]
- Botchkareva, N.V.; Ahluwalia, G.; Shander, D. Apoptosis in the Hair Follicle. J. Investig. Dermatol. 2006, 126, 258–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luanpitpong, S.; Nimmannit, U.; Chanvorachote, P.; Leonard, S.S.; Pongrakhananon, V.; Wang, L.; Rojanasakul, Y. Hydroxyl radical mediates cisplatin-induced apoptosis in human hair follicle dermal papilla cells and keratinocytes through Bcl-2-dependent mechanism. Apoptosis 2011, 16, 769–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, K.; Nimnual, A.; Zong, W.-X.; Kennedy, N.J.; Flavell, R.A.; Thompson, C.B.; Bar-Sagi, D.; Davis, R.J. The Bax Subfamily of Bcl2-Related Proteins Is Essential for Apoptotic Signal Transduction by c-Jun NH2-Terminal Kinase. Mol. Cell. Biol. 2002, 22, 4929–4942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manse, Y.; Luo, F.; Kato, K.; Okazaki, A.; Okada-Nishida, E.; Yanagida, M.; Nakamura, S.; Morikawa, T. Ent-kaurane-type diterpenoids from Isodonis Herba activate human hair follicle dermal papilla cells proliferation via the Akt/GSK-3β/β-catenin transduction pathway. J. Nat. Med. 2021, 75, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Jacobson, K.; Schaller, M.D. MAP kinases and cell migration. J. Cell Sci. 2004, 117, 4619–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, H.; Lee, S.; Kim, S.; Park, D.; Jung, E. Effect of sinapic acid on hair growth promoting in human hair follicle dermal papilla cells via Akt activation. Arch. Dermatol. Res. 2017, 309, 381–388. [Google Scholar] [CrossRef]
- Zhou, Q.; Song, Y.; Zheng, Q.; Han, R.; Cheng, H. Expression profile analysis of dermal papilla cells mRNA in response to WNT10B treatment. Exp. Ther. Med. 2019, 19, 1017–1023. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.S.; Jeong, J.; Lee, C.M.; Lee, K.S.; Lee, J.N.; Park, S.M.; Lee, Y.M. Treatment with Malva verticillata seed extracts alleviates alopecia via activation of Wnt/β-catenin signaling. Preprints 2020. [Google Scholar] [CrossRef]
- Choi, B.Y. Targeting Wnt/β-Catenin Pathway for Developing Therapies for Hair Loss. Int. J. Mol. Sci. 2020, 21, 4915. [Google Scholar] [CrossRef] [PubMed]
- Evangelisti, C.; Cenni, V.; Lattanzi, G. Potential therapeutic effects of the MTOR inhibitors for preventing ageing and progeria-related disorders. Br. J. Clin. Pharmacol. 2016, 82, 1229–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisvert, W.A.; Yu, M.; Choi, Y.; Jeong, G.H.; Zhang, Y.-L.; Cho, S.; Choi, C.; Lee, S.; Lee, B.-H. Hair growth-promoting effect of Geranium sibiricum extract in human dermal papilla cells and C57BL/6 mice. BMC Complement. Altern. Med. 2017, 17, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Sima, G.; Wu, C.; Fan, Y.; Tan, Y.; Wang, Z.; Cheng, G.; Li, J. Decreased MiR-17 in glioma cells increased cell viability and migration by increasing the expression of Cyclin D1, p-Akt and Akt. PLoS ONE 2018, 13, e0190515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Diao, S.; Wang, Q.; Zhu, C.; Sun, X.; Yin, B.; Zhang, X.; Meng, X.; Wang, B. IL-17A promotes cell migration and invasion of glioblastoma cells via activation of PI3K/AKT signalling pathway. J. Cell. Mol. Med. 2019, 23, 357–369. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Su, Y.; Wang, J.; Gao, Y.; Yang, F.; Li, G.; Shi, Q. Ginsenoside Rb1 promotes the growth of mink hair follicle via PI3K/AKT/GSK-3β signaling pathway. Life Sci. 2019, 229, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Huelsken, J.; Vogel, R.; Erdmann, B.; Cotsarelis, G.; Birchmeier, W. β-Catenin Controls Hair Follicle Morphogenesis and Stem Cell Differentiation in the Skin. Cell 2001, 105, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Yang, Z.; Andl, T.; Cui, C.; Kim, N.; Millar, S.E.; Cotsarelis, G. Wnt-dependent de novo hair follicle regeneration in adult mouse skin after wounding. Nat. Cell Biol. 2007, 447, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Ouji, Y.; Yoshikawa, M.; Moriya, K.; Nishiofuku, M.; Matsuda, R.; Ishizaka, S. Wnt-10b, uniquely among Wnts, promotes epithelial differentiation and shaft growth. Biochem. Biophys. Res. Commun. 2008, 367, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.L.; Gangadaran, P.; Seo, C.H.; Kwack, M.H.; Oh, J.M.; Lee, H.W.; Gopal, A.; Sung, Y.K.; Jeong, S.Y.; Lee, S.-W.; et al. Macrophage-Derived Extracellular Vesicle Promotes Hair Growth. Cells 2020, 9, 856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andl, T.; Reddy, S.T.; Gaddapara, T.; Millar, S.E. WNT Signals Are Required for the Initiation of Hair Follicle Development. Dev. Cell 2002, 2, 643–653. [Google Scholar] [CrossRef]
- Maretto, S.; Cordenonsi, M.; Dupont, S.; Braghetta, P.; Broccoli, V.; Hassan, A.B.; Piccolo, S. Mapping Wnt/β-catenin signaling during mouse development and in colorectal tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 3299–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enshell-Seijffers, D.; Lindon, C.; Kashiwagi, M.; Morgan, B.A. β-catenin Activity in the Dermal Papilla Regulates Morphogenesis and Regeneration of Hair. Dev. Cell 2010, 18, 633–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clevers, H.; & Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alao, J.P. The regulation of cyclin D1 degradation: Roles in cancer development and the potential for therapeutic invention. Mol. Cancer 2007, 6, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.R.; Bae, S.; Kim, J.Y.; Lee, J.; Cho, D.H.; Kim, H.S.; An, S. Monoterpenoid loliolide regulates hair follicle inductivity of human dermal papilla cells by activating the Akt/β-catenin signaling pathway. J. Microbiol. Biotechnol. 2019, 29, 1830–1840. [Google Scholar] [CrossRef]
- Yamauchi, K.; Kurosaka, A. Inhibition of glycogen synthase kinase-3 enhances the expression of alkaline phosphatase and insulin-like growth factor-1 in human primary dermal papilla cell culture and maintains mouse hair bulbs in organ culture. Arch. Dermatol. Res. 2009, 301, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Nito, C.; Kamada, H.; Nishi, T.; Chan, P.H. Activation of the Akt/GSK3β Signaling Pathway Mediates Survival of Vulnerable Hippocampal Neurons after Transient Global Cerebral Ischemia in Rats. Br. J. Pharmacol. 2006, 26, 1479–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.H.; Xiang, L.J.; Shi, H.X.; Zhang, J.; Jiang, L.P.; Cai, P.T.; Xiao, J. Fibroblast growth factors stimulate hair growth through β-catenin and Shh expression in C57BL/6 mice. BioMed Res. Int. 2015, 2015, 730139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; McElwee, K.J.; Owen, G.R.; Häkkinen, L.; Larjava, H.S. Integrin β6-Deficient Mice Show Enhanced Keratinocyte Proliferation and Retarded Hair Follicle Regression after Depilation. J. Investig. Dermatol. 2012, 132, 547–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, K.; Brown, L.F.; Detmar, M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J. Clin. Investig. 2001, 107, 409–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.H.; Lee, J.S.; Kim, Y.C. Hair Growth-Promoting Effects of Lavender Oil in C57BL/6 Mice. Toxicol. Res. 2016, 32, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, S.; Andl, T.; Bagasra, A.; Lu, M.M.; Epstein, D.J.; Morrisey, E.E.; Millar, S.E. Characterization of Wnt gene expression in developing and postnatal hair follicles and identification of Wnt5a as a target of Sonic hedgehog in hair follicle morphogenesis. Mech. Dev. 2001, 107, 69–82. [Google Scholar] [CrossRef]
- Li, Y.-H.; Zhang, K.; Yang, K.; Ye, J.-X.; Xing, Y.-Z.; Guo, H.-Y.; Deng, F.; Lian, X.-H.; Yang, T. Adenovirus-Mediated Wnt10b Overexpression Induces Hair Follicle Regeneration. J. Investig. Dermatol. 2013, 133, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-H.; Zhang, K.; Ye, J.-X.; Lian, X.-H.; Yang, T. Wnt10b promotes growth of hair follicles via a canonical Wnt signalling pathway. Clin. Exp. Dermatol. 2011, 36, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Bejaoui, M.; Villareal, M.O.; Isoda, H. β-catenin-mediated hair growth induction effect of 3,4,5-tri-O-caffeoylquinic acid. Aging 2019, 11, 4216–4237. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.-Y.; Sennett, R.; Rezza, A.; Clavel, C.; Grisanti, L.; Zemla, R.; Najam, S.; Rendl, M. Wnt/β-catenin signaling in dermal condensates is required for hair follicle formation. Dev. Biol. 2014, 385, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, S.-Y.; Pi, L.-Q.; Hwang, S.T.; Lee, W.-S. Effect of IGF-I on Hair Growth Is Related to the Anti-Apoptotic Effect of IGF-I and Up-Regulation of PDGF-A and PDGF-B. Ann. Dermatol. 2012, 24, 26–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudman, S.M.; Philpott, M.P.; Thomas, G.A.; Kealey, T. The Role of IGF-I in Human Skin and its Appendages: Morphogen as Well as Mitogen? J. Investig. Dermatol. 1997, 109, 770–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.Y.; Hickford, J.G.; Bickerstaffe, R.; Palmer, B.R. Insulin-like growth factor 1 and hair growth. Dermatol. Online J. 1999, 5, 5. [Google Scholar]
- Little, J.C.; Redwood, K.R.; Stones, A.J.; Gibson, W.T.; Granger, S.P. The insulin-like growth-factor-i receptor is important in controlling the hair-growth cycle. J. Investig. Dermatol. 1994, 102, 533. [Google Scholar]
- Ahn, J.H.; Park, Y.E.; Kim, B.; Park, C.W.; Sim, T.H.; Lee, T.-K.; Lee, J.-C.; Park, J.H.; Kim, J.-D.; Lee, H.S.; et al. Hair Growth is Promoted in Mouse Dorsal Skin by a Mixture of Platycladus orientalis (L.) Franco Leaf Extract and Alpha-Terpineol by Increasing Growth Factors and wnt3/β-Catenin. Nat. Prod. Commun. 2020, 15, 1934578X20951433. [Google Scholar] [CrossRef]
- Aase, K.; von Euler, G.; Li, X.; Pontén, A.; Thorén, P.; Cao, R.; Cao, Y.; Olofsson, B.; Gebre-Medhin, S.; Pekny, M.; et al. Vascular Endothelial Growth Factor-B–Deficient Mice Display an Atrial Conduction Defect. Circulation 2001, 104, 358–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellomo, D.; Headrick, J.P.; Silins, G.U.; Paterson, C.A.; Thomas, P.S.; Gartside, M.; Mould, A.; Cahill, M.M.; Tonks, I.D.; Grimmond, S.M.; et al. Mice lacking the vascular endothelial growth factor-B gene (Vegfb) have smaller hearts, dysfunctional coronary vasculature, and impaired recovery from cardiac ischemia. Circ. Res. 2000, 86, 29. [Google Scholar] [CrossRef] [Green Version]
- Reichelt, M.; Shi, S.; Hayes, M.; Kay, G.; Batch, J.; Gole, G.; Browning, J. Vascular endothelial growth factor-B and retinal vascular development in the mouse. Clin. Exp. Ophthalmol. 2003, 31, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Louzier, V.; Raffestin, B.; Leroux, A.; Branellec, D.; Caillaud, J.M.; Levame, M.; Eddahibi, S.; Adnot, S. Role of VEGF-B in the lung during development of chronic hypoxic pulmonary hypertension. Am. J. Physiol. Cell. Mol. Physiol. 2003, 284, L926–L937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.; Kim, S.; Kim, D.; Kim, S.; Lee, H.; Hwang, D.; Kim, H.S. Potential Effects of Micro-Current Stimulation for Reduction of Abdominal Fat and Weight Loss: Clinical Study. J. Biomed. Eng. Res. 2018, 39, 284–296. [Google Scholar]
- Yoshikawa, Y.; Sugimoto, M.; Uemura, M.; Matsuo, M.; Maeshige, N.; Niba, E.T.E.; Shuntoh, H. Monophasic Pulsed Microcurrent of 1–8 Hz Increases the Number of Human Dermal Fibroblasts. Prog. Rehabil. Med. 2016, 1, 20160005. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Primer Name | Forward | Reverse |
|---|---|---|---|
| Mouse | IGF-1 | GTCGTCTTCACACCTCTTCTACCT | GCACAGTACATCTCCAGTCTCCT |
| IGF-1R | CTCAGGCTTCATCCGCAACAG | GTTCTCCAACTCCGAGGCAATG | |
| Wnt5a | CTGGCAGGACTTTCTCAAGG | CTCTAGCGTCCACGAACTCC | |
| Wnt10b | CCTGTCCGGACTGAGTAAGC | TTGCTCACCACTACCCTTCC | |
| FGF2 | CAAGAACGGCGGCTTCTTC | GAAAGAAACAGTATGGCCT | |
| FGF7 | AGACTGTTCTGTCGCACC | CCGCTGTGTGTCCATTTAG | |
| FGF10 | TGTCCGCTGGAGAAGGCTGTTC | CTATGTTTGGATCGTCATGG | |
| VEGF-A | CGAGATAGAGTACATCTTCAAGCC | TCATCGTTACAGCAGCCTGC | |
| VEGF-B | AAAAAAAAAGGAGAGTGCTGTGAAG | TCCCAGCCCGGAACAGA | |
| GAPDH | GCCAAGGTCATCCATGACAACT | GAGGGGCCATCCACAGTCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, D.; Lee, H.; Lee, J.; Lee, M.; Cho, S.; Kim, T.; Kim, H. Micro-Current Stimulation Has Potential Effects of Hair Growth-Promotion on Human Hair Follicle-Derived Papilla Cells and Animal Model. Int. J. Mol. Sci. 2021, 22, 4361. https://doi.org/10.3390/ijms22094361
Hwang D, Lee H, Lee J, Lee M, Cho S, Kim T, Kim H. Micro-Current Stimulation Has Potential Effects of Hair Growth-Promotion on Human Hair Follicle-Derived Papilla Cells and Animal Model. International Journal of Molecular Sciences. 2021; 22(9):4361. https://doi.org/10.3390/ijms22094361
Chicago/Turabian StyleHwang, Donghyun, Hana Lee, Jinho Lee, Minjoo Lee, Seungkwan Cho, Tackjoong Kim, and Hansung Kim. 2021. "Micro-Current Stimulation Has Potential Effects of Hair Growth-Promotion on Human Hair Follicle-Derived Papilla Cells and Animal Model" International Journal of Molecular Sciences 22, no. 9: 4361. https://doi.org/10.3390/ijms22094361