
Identification and Analysis of Axolotl Homologs for Proteins Implicated in Human Neurodegenerative Proteinopathies
 , ,
, ,
Abstract
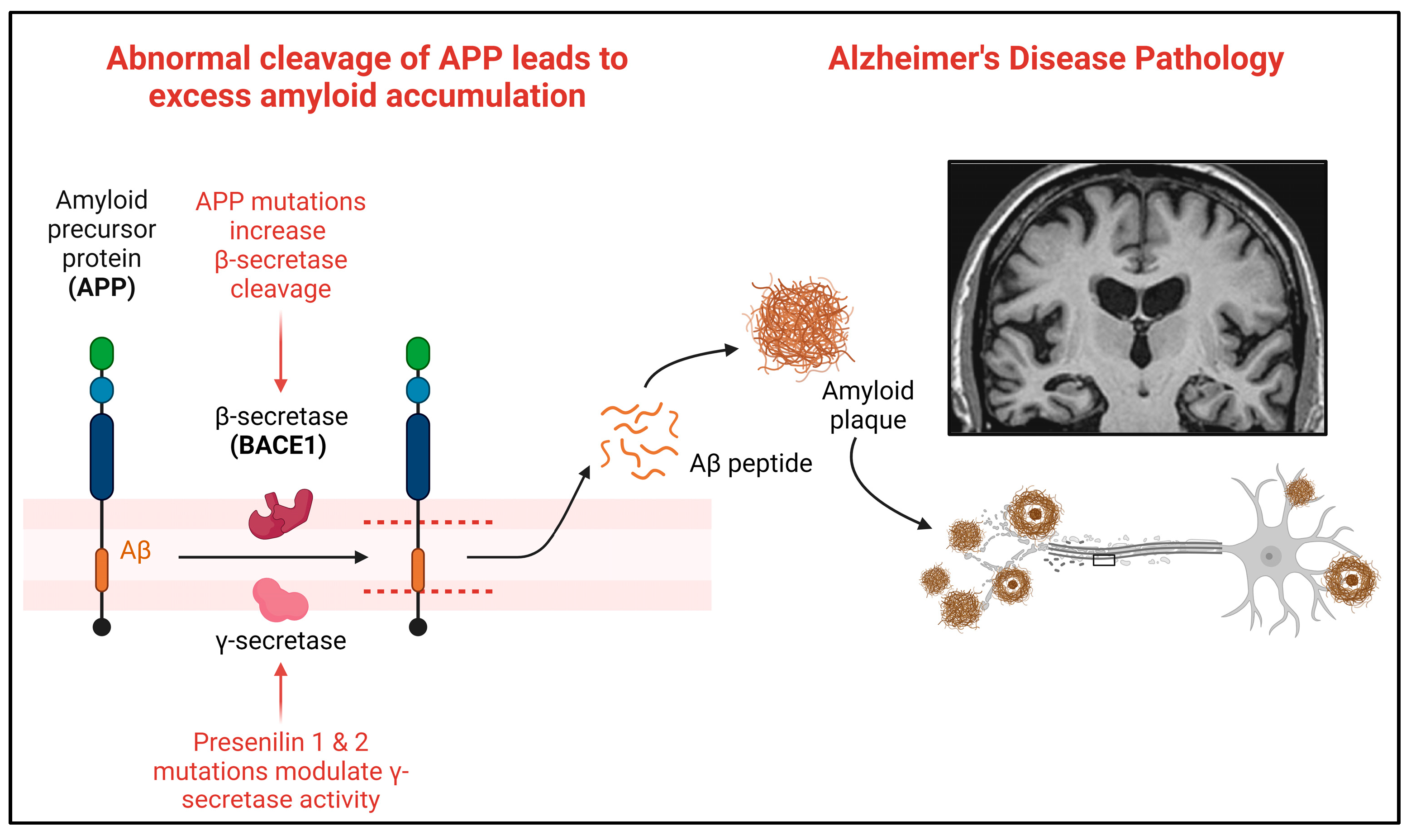
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Identification of Putative Homologous Proteins in Axolotls and Humans
2.3. Putative Protein Alignment
2.4. Antibody Selection
2.5. Preparation of Brain Tissue for Immunoblot Assay
2.6. Immunoblotting
2.7. Immunohistochemistry
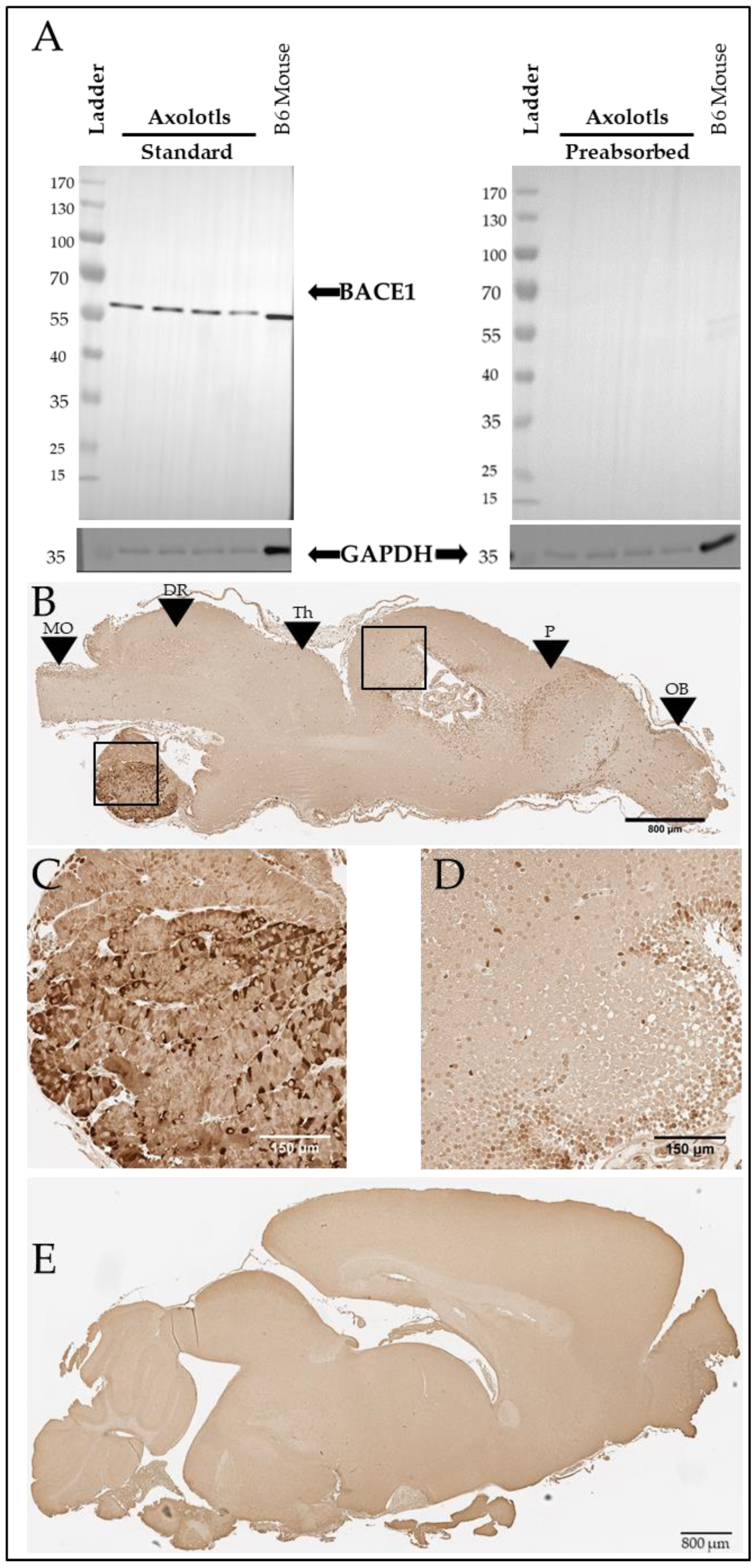
3. Results
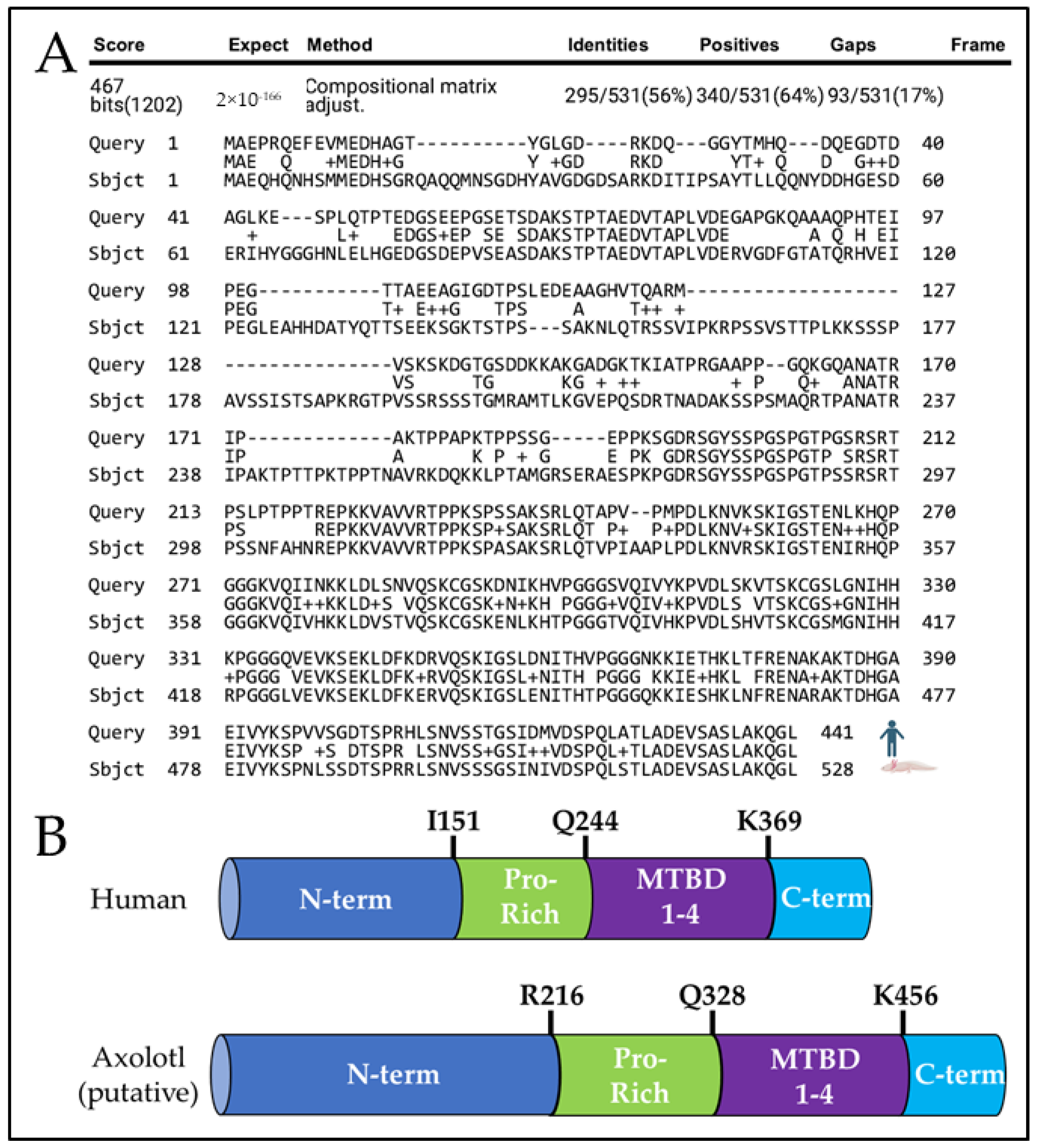
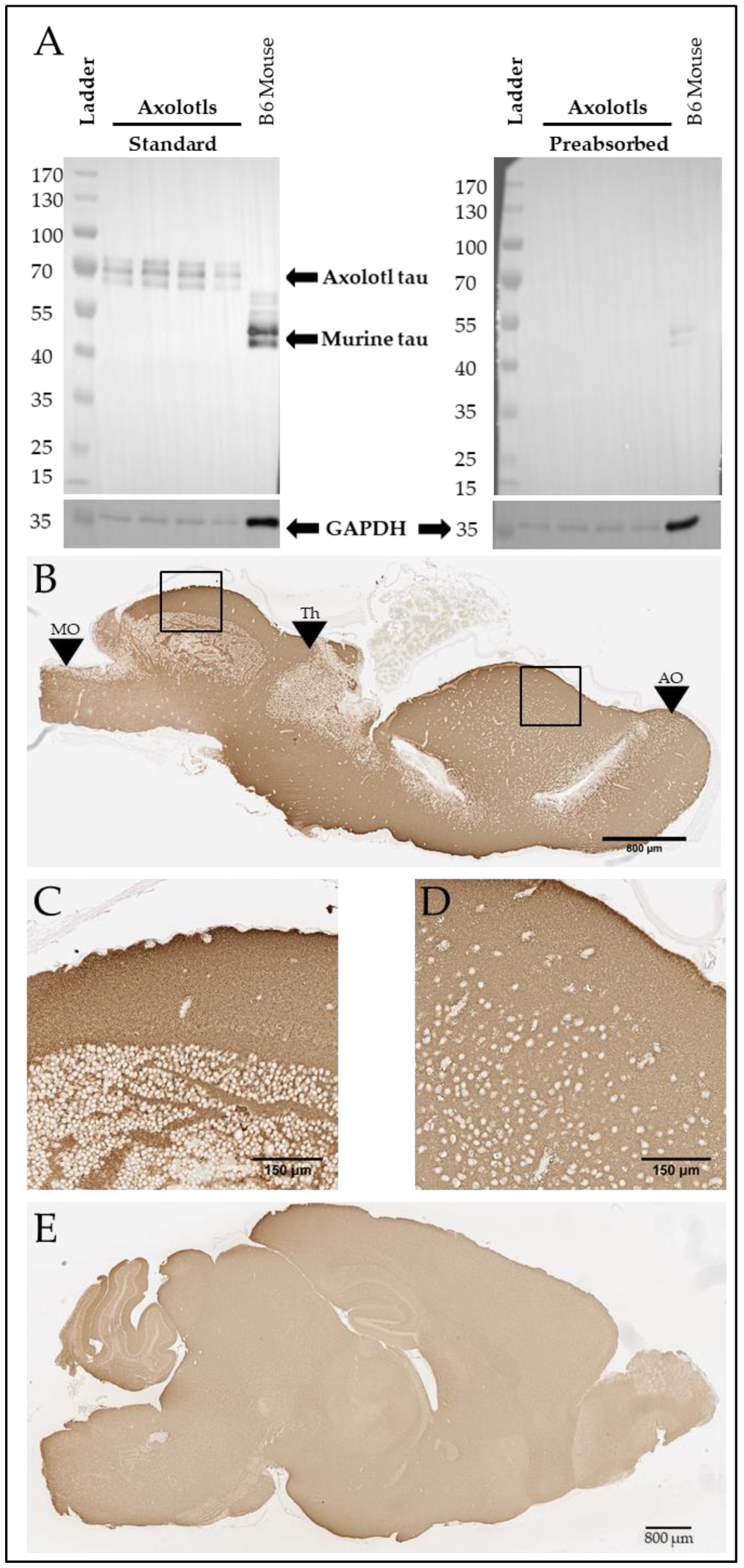
A Tau-like Protein Is Expressed in the Axolotl Brain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Status Report on the Public Health Response to Dementia; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Selkoe, D.J.; Hardy, J. The Amyloid Hypothesis of Alzheimer’s Disease at 25 Years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef]
- Rabbito, A.; Dulewicz, M.; Kulczyńska-Przybik, A.; Mroczko, B. Biochemical Markers in Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 1989. [Google Scholar] [CrossRef]
- Ciechanover, A.; Kwon, Y.T. Degradation of Misfolded Proteins in Neurodegenerative Diseases: Therapeutic Targets and Strategies. Exp. Mol. Med. 2015, 47, e147. [Google Scholar] [CrossRef]
- Anderson, R.M.; Hadjichrysanthou, C.; Evans, S.; Wong, M.M. Why Do so Many Clinical Trials of Therapies for Alzheimer’s Disease Fail? Lancet 2017, 390, 2327–2329. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, H.; Li, R.; Sterling, K.; Song, W. Amyloid β-Based Therapy for Alzheimer’s Disease: Challenges, Successes and Future. Signal Transduct. Target. Ther. 2023, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Jin, Y. Intrinsic Control of Axon Regeneration. Neuron 2016, 90, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Eremichev, R.; Kulebyakina, M.; Alexandrushkina, N.; Nimiritsky, P.; Basalova, N.; Grigorieva, O.; Egiazaryan, M.; Dyikanov, D.; Tkachuk, V.; Makarevich, P. Scar-Free Healing of Endometrium: Tissue-Specific Program of Stromal Cells and Its Induction by Soluble Factors Produced after Damage. Front. Cell Dev. Biol. 2021, 9, 616893. [Google Scholar] [CrossRef]
- Takeo, M.; Lee, W.; Ito, M. Wound Healing and Skin Regeneration. Cold Spring Harb. Perspect. Med. 2015, 5, a023267. [Google Scholar] [CrossRef]
- Angelotti, M.L.; Becherucci, F.; Mazzinghi, B.; Peired, A.; Romagnani, P. Chapter 69—Principles of Kidney Regeneration. In Kidney Transplantation, Bioengineering and Regeneration; Orlando, G., Remuzzi, G., Williams, D.F., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 973–988. ISBN 978-0-12-801734-0. [Google Scholar]
- Navarro Negredo, P.; Yeo, R.W.; Brunet, A. Aging and Rejuvenation of Neural Stem Cells and Their Niches. Cell Stem Cell 2020, 27, 202–223. [Google Scholar] [CrossRef]
- Dennis, C.V.; Suh, L.S.; Rodriguez, M.L.; Kril, J.J.; Sutherland, G.T. Human Adult Neurogenesis across the Ages: An Immunohistochemical Study. Neuropathol. Appl. Neurobiol. 2016, 42, 621–638. [Google Scholar] [CrossRef] [PubMed]
- Maden, M. Salamanders as Key Models for Development and Regeneration Research. In Salamanders; Methods in Molecular Biology; Humana: New York, NY, USA, 2023; Volume 2562, pp. 1–23. [Google Scholar] [CrossRef]
- Amamoto, R.; Huerta, V.G.L.; Takahashi, E.; Dai, G.; Grant, A.K.; Fu, Z.; Arlotta, P. Adult Axolotls Can Regenerate Original Neuronal Diversity in Response to Brain Injury. eLife 2016, 5, e13998. [Google Scholar] [CrossRef]
- Maden, M.; Manwell, L.A.; Ormerod, B.K. Proliferation Zones in the Axolotl Brain and Regeneration of the Telencephalon. Neural Dev. 2013, 8, 1. [Google Scholar] [CrossRef]
- Berg, D.A.; Kirkham, M.; Beljajeva, A.; Knapp, D.; Habermann, B.; Ryge, J.; Tanaka, E.M.; Simon, A. Efficient Regeneration by Activation of Neurogenesis in Homeostatically Quiescent Regions of the Adult Vertebrate Brain. Development 2010, 137, 4127–4134. [Google Scholar] [CrossRef]
- Nowoshilow, S. The Axolotl Genome and the Evolution of Key Tissue Formation Regulators. Nature 2018, 554, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Nowoshilow, S.; Tanaka, E.M. Introducing www.Axolotl-Omics.Org—An Integrated -Omics Data Portal for the Axolotl Research Community. Exp. Cell Res. 2020, 394, 112143. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Wootton, J.C.; Gertz, E.M.; Agarwala, R.; Morgulis, A.; Schäffer, A.A.; Yu, Y.-K. Protein Database Searches Using Compositionally Adjusted Substitution Matrices. FEBS J. 2005, 272, 5101–5109. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Bryant, S.H. CD-Search: Protein Domain Annotations on the Fly. Nucleic Acids Res. 2004, 32, W327–W331. [Google Scholar] [CrossRef] [PubMed]
- Petry, F.R.; Pelletier, J.; Bretteville, A.; Morin, F.; Calon, F.; Hébert, S.S.; Whittington, R.A.; Planel, E. Specificity of Anti-Tau Antibodies When Analyzing Mice Models of Alzheimer’s Disease: Problems and Solutions. PLoS ONE 2014, 9, e94251. [Google Scholar] [CrossRef] [PubMed]
- Murie, M.; Peng, Y.; Rigby, M.J.; Dieterich, I.A.; Farrugia, M.A.; Endresen, A.; Bhattacharyya, A.; Puglielli, L. ATase Inhibition Rescues Age-Associated Proteotoxicity of the Secretory Pathway. Commun. Biol. 2022, 5, 173. [Google Scholar] [CrossRef]
- Zou, H.-Y.; Guo, L.; Zhang, B.; Chen, S.; Wu, X.-R.; Liu, X.-D.; Xu, X.-Y.; Li, B.-Y.; Chen, S.; Xu, N.-J.; et al. Aberrant miR-339-5p/Neuronatin Signaling Causes Prodromal Neuronal Calcium Dyshomeostasis in Mutant Presenilin Mice. J. Clin. Investig. 2022, 132, e149160. [Google Scholar] [CrossRef]
- Bailey, R.M.; Covy, J.P.; Melrose, H.L.; Rousseau, L.; Watkinson, R.; Knight, J.; Miles, S.; Farrer, M.J.; Dickson, D.W.; Giasson, B.I.; et al. LRRK2 Phosphorylates Novel Tau Epitopes and Promotes Tauopathy. Acta Neuropathol. 2013, 126, 809–827. [Google Scholar] [CrossRef]
- Iqbal, K.; Liu, F.; Gong, C.-X.; Grundke-Iqbal, I. Tau in Alzheimer Disease and Related Tauopathies. Curr. Alzheimer Res. 2010, 7, 656–664. [Google Scholar] [CrossRef]
- Xu, G.; Ulm, B.S.; Howard, J.; Fromholt, S.E.; Lu, Q.; Lee, B.B.; Walker, A.; Borchelt, D.R.; Lewis, J. TAPPing into the Potential of Inducible Tau/APP Transgenic Mice. Neuropathol. Appl. Neurobiol. 2022, 48, e12791. [Google Scholar] [CrossRef]
- Giasson, B.I.; Forman, M.S.; Higuchi, M.; Golbe, L.I.; Graves, C.L.; Kotzbauer, P.T.; Trojanowski, J.Q.; Lee, V.M.-Y. Initiation and Synergistic Fibrillization of Tau and α-Synuclein. Science 2003, 300, 636–640. [Google Scholar] [CrossRef]
- Gao, Y.-L.; Wang, N.; Sun, F.-R.; Cao, X.-P.; Zhang, W.; Yu, J.-T. Tau in Neurodegenerative Disease. Ann. Transl. Med. 2018, 6, 175. [Google Scholar] [CrossRef]
- Lee, G.; Leugers, C.J. Tau and Tauopathies. Prog. Mol. Biol. Transl. Sci. 2012, 107, 263–293. [Google Scholar] [CrossRef] [PubMed]
- Gustke, N.; Trinczek, B.; Biernat, J.; Mandelkow, E.-M.; Mandelkow, E. Domains of Tau Protein and Interactions with Microtubules. Biochemistry 1994, 33, 9511–9522. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Rojas, C.; Cabezas-Opazo, F.; Deaton, C.A.; Vergara, E.H.; Johnson, G.V.W.; Quintanilla, R.A. It’s All about Tau. Prog. Neurobiol. 2019, 175, 54–76. [Google Scholar] [CrossRef] [PubMed]
- Mukrasch, M.D.; Bibow, S.; Korukottu, J.; Jeganathan, S.; Biernat, J.; Griesinger, C.; Mandelkow, E.; Zweckstetter, M. Structural Polymorphism of 441-Residue Tau at Single Residue Resolution. PLoS Biol. 2009, 7, e34. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Wischik, C.M.; Crowther, R.A.; Walker, J.E.; Klug, A. Cloning and Sequencing of the cDNA Encoding a Core Protein of the Paired Helical Filament of Alzheimer Disease: Identification as the Microtubule-Associated Protein Tau. Proc. Natl. Acad. Sci. USA 1988, 85, 4051–4055. [Google Scholar] [CrossRef]
- Shoji, M.; Golde, T.E.; Ghiso, J.; Cheung, T.T.; Estus, S.; Shaffer, L.M.; Cai, X.D.; McKay, D.M.; Tintner, R.; Frangione, B. Production of the Alzheimer Amyloid β Protein by Normal Proteolytic Processing. Science 1992, 258, 126–129. [Google Scholar] [CrossRef]
- O’Brien, R.J.; Wong, P.C. Amyloid Precursor Protein Processing and Alzheimer’s Disease. Annu. Rev. Neurosci. 2011, 34, 185–204. [Google Scholar] [CrossRef]
- Weidemann, A.; König, G.; Bunke, D.; Fischer, P.; Salbaum, J.M.; Masters, C.L.; Beyreuther, K. Identification, Biogenesis, and Localization of Precursors of Alzheimer’s Disease A4 Amyloid Protein. Cell 1989, 57, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, E.; Small, D.H. Insights into the Physiological Function of the β-Amyloid Precursor Protein: Beyond Alzheimer’s Disease. J. Neurochem. 2014, 129, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.A.; Przemylska, L.; Clavane, E.M.; Meakin, P.J. BACE1: More than Just a β -secretase. Obes. Rev. 2022, 23, e13430. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P. The Sequence Manipulation Suite: JavaScript Programs for Analyzing and Formatting Protein and DNA Sequences. BioTechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Li, P.; Liu, P.; Xu, S.; Wang, F.; Huang, J.H. Post-Translational Modifications of BACE1 in Alzheimer’s Disease. Curr. Neuropharmacol. 2022, 20, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Prokop, S.; Giasson, B.I. “Don’t Phos Over Tau”: Recent Developments in Clinical Biomarkers and Therapies Targeting Tau Phosphorylation in Alzheimer’s Disease and Other Tauopathies. Mol. Neurodegener. 2021, 16, 37. [Google Scholar] [CrossRef]
- Waxman, E.A.; Giasson, B.I. Induction of Intracellular Tau Aggregation Is Promoted by α-Synuclein Seeds and Provides Novel Insights into the Hyperphosphorylation of Tau. J. Neurosci. 2011, 31, 7604–7618. [Google Scholar] [CrossRef]
- Peeraer, E.; Bottelbergs, A.; Van Kolen, K.; Stancu, I.-C.; Vasconcelos, B.; Mahieu, M.; Duytschaever, H.; Ver Donck, L.; Torremans, A.; Sluydts, E.; et al. Intracerebral Injection of Preformed Synthetic Tau Fibrils Initiates Widespread Tauopathy and Neuronal Loss in the Brains of Tau Transgenic Mice. Neurobiol. Dis. 2015, 73, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Iba, M.; Guo, J.L.; McBride, J.D.; Zhang, B.; Trojanowski, J.Q.; Lee, V.M.-Y. Synthetic Tau Fibrils Mediate Transmission of Neurofibrillary Tangles in a Transgenic Mouse Model of Alzheimer’s-like Tauopathy. J. Neurosci. 2013, 33, 1024–1037. [Google Scholar] [CrossRef] [PubMed]
- Sarasa, M.; Pesini, P. Natural Non-Trasgenic Animal Models for Research in Alzheimer’s Disease. Curr. Alzheimer Res. 2009, 6, 171–178. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
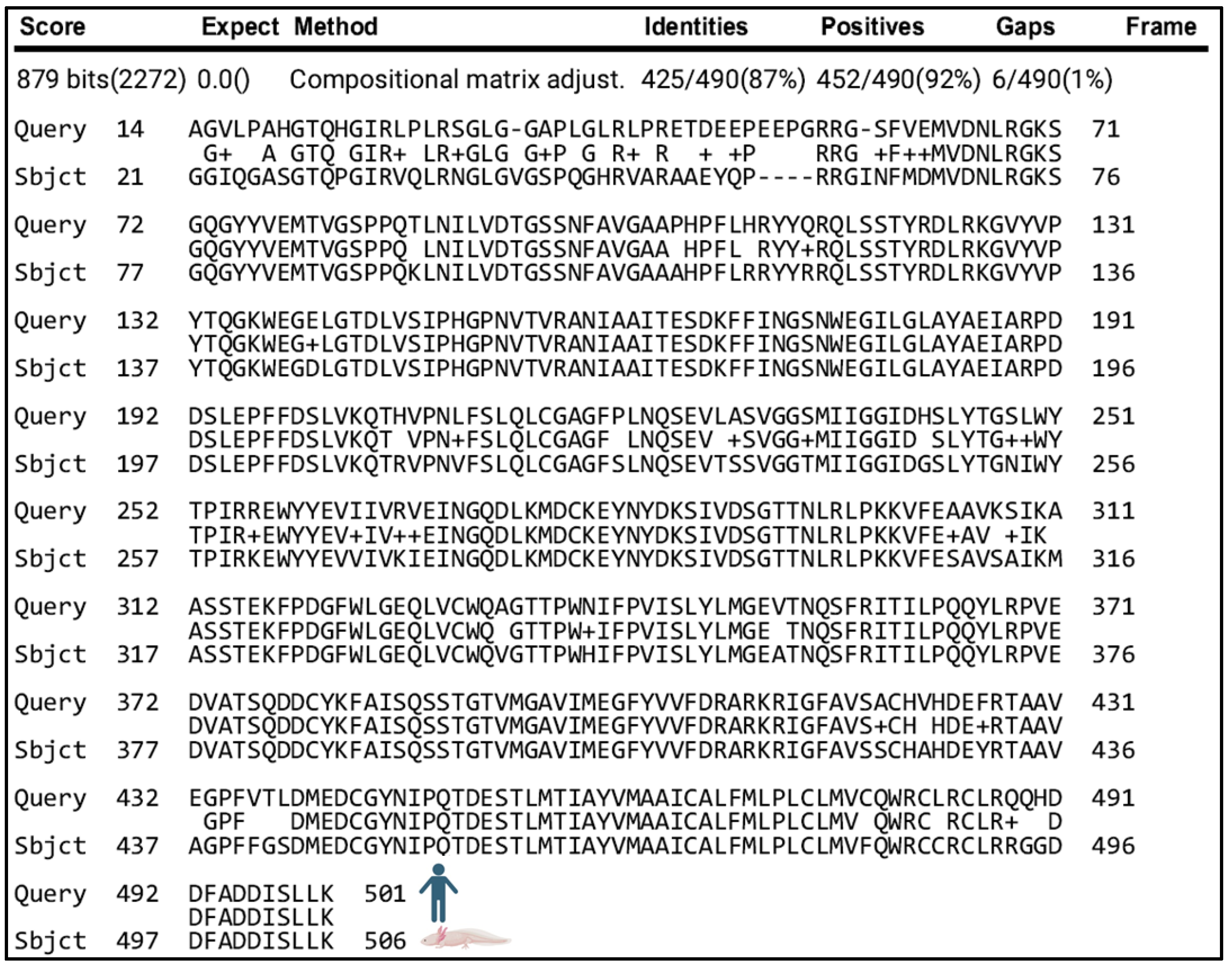
| Human Gene | Protein | Score | E-Value | Percent Identical | Percent Functionally Equivalent | Gaps |
|---|---|---|---|---|---|---|
| MAPT | Microtubule-associated protein tau | 467 | 2 × 10−166 | 56% (295/531) | 64% (340/531) | 17% (93/531) |
| APP | Amyloid precursor protein | 1305 | 0.0 | 88% (658/752) | 92% (696/752) | 2% (22/752) |
| BACE1 | β-secretase 1 | 879 | 0.0 | 87% (425/490) | 92% (452/490) | 1% (6/490) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, L.M.; Strickland, Z.; Lopez, N.; Whited, J.L.; Maden, M.; Lewis, J. Identification and Analysis of Axolotl Homologs for Proteins Implicated in Human Neurodegenerative Proteinopathies. Genes 2024, 15, 310. https://doi.org/10.3390/genes15030310
James LM, Strickland Z, Lopez N, Whited JL, Maden M, Lewis J. Identification and Analysis of Axolotl Homologs for Proteins Implicated in Human Neurodegenerative Proteinopathies. Genes. 2024; 15(3):310. https://doi.org/10.3390/genes15030310
Chicago/Turabian StyleJames, Lucas M., Zachary Strickland, Noah Lopez, Jessica L. Whited, Malcolm Maden, and Jada Lewis. 2024. "Identification and Analysis of Axolotl Homologs for Proteins Implicated in Human Neurodegenerative Proteinopathies" Genes 15, no. 3: 310. https://doi.org/10.3390/genes15030310