
Extracellular Vesicles: Investigating the Pathophysiology of Diabetes-Associated Hypertension and Diabetic Nephropathy


1
Department of Physiology and Aging, College of Medicine, University of Florida, Gainesville, FL 32610, USA
2
Department of Medicine, Division of Nephrology, Hypertension, and Renal Transplantation, University of Florida, Gainesville, FL 32610, USA
3
Department of Pediatrics, College of Medicine, University of Florida, Gainesville, FL 32610, USA
Biology 2023, 12(8), 1138; https://doi.org/10.3390/biology12081138
Submission received: 21 June 2023
/
Revised: 3 August 2023
/
Accepted: 10 August 2023
/
Published: 16 August 2023
(This article belongs to the Special Issue Extracellular Vesicles and Pathophysiology)
Abstract
:Simple Summary
Hypertension may present before or after the onset of diabetes, but in either case, it may increase the risk of developing diabetic nephropathy, the leading cause of end-stage kidney disease. Extracellular vesicles allow for communication between different cell types, the alteration of various signaling pathways, and are involved in disease mechanisms. Various omic studies have revealed that circulating extracellular vesicles and urinary extracellular vesicles are a rich source of biomarkers and markers of the prognosis of specific kidney-associated diseases including diabetes-associated hypertension and diabetic nephropathy.
Abstract
Extracellular vesicles (EVs) include exosomes, microvesicles, and apoptotic bodies. EVs are released by all cell types and are found in biological fluids including plasma and urine. Urinary extracellular vesicles (uEVs) are a mixed population of EVs that comprise small EVs that are filtered and excreted, EVs secreted by tubular epithelial cells, and EVs released from the bladder, urethra, and prostate. The packaged cargo within uEVs includes bioactive molecules such as metabolites, lipids, proteins, mRNAs, and miRNAs. These molecules are involved in intercellular communication, elicit changes in intracellular signaling pathways, and play a role in the pathogenesis of various diseases including diabetes-associated hypertension and diabetic nephropathy. uEVs represent a rich source of biomarkers, prognosis markers, and can be loaded with small-molecule drugs as a vehicle for delivery.
1. Introduction
Extracellular vesicles (EVs) are a heterogeneous population of vesicles that are generally classified according to their size and biogenesis. Exosomes are the smallest type of EV and originate from the formation of multivesicular bodes [1]. Microvesicles (previously called microparticles) are generally less than 1000 nm in diameter and originate from the budding of the plasma membrane [1]. Apopotic bodies are among the largest type of EV and also originate from the budding of the plasma membrane [1]. EVs were once thought to function as cellular garbage disposals [2], but it is now accepted that EVs play an important role in cellular communication [3,4,5], intracellular signaling [6,7,8], and cellular differentiation [9,10,11]. In addition, EVs are used in multiple applications [12,13,14]. EVs isolated from biological fluids including urine [15], plasma [16], and cerebrospinal fluid [17] represent a rich source of biomarkers. Additionally, the profiling of the EV cargo can be used as markers of diagnosis and prognosis [18,19]. Engineered EVs can be loaded or fused with small-molecule drugs and used as a vehicle for targeted delivery and combination drug therapy [20,21]. One study demonstrated the renoprotective potential of uEVs after showing the loss of miRs through EVs may be associated with a decrease in the levels in the kidney and promote renal fibrosis in diabetic nephropathy [22]. Various applications of EVs are depicted in Figure 1. EVs derived from plasma or urine samples contain important molecules that have been used in the diagnostics of various diseases and in some cases reveal information about different stages of disease [23,24,25,26]. Engineered EVs have been shown to have therapeutic potential [27,28]. In addition, EVs have been studied in the context of computational or molecular modeling [29,30]. The cargo enriched in EVs has also been shown to serve as a rich source of biomarkers for various diseases [31,32,33].
2. EVs in Cellular Communication and Alterations of Intracellular Signaling Pathways in Physiology and Pathophysiology
EVs have been shown to allow for crosstalk between different cell types, including those within different segments of the nephron (Figure 2). A study by Wu et al. showed that EVs from high-glucose-treated glomerular endothelial cells activate glomerular mesangial cells by promoting proliferation and increasing extracellular-matrix protein production and alpha smooth muscle actin expression [34]. A study by Jella et al. showed that EVs from proximal tubule cells regulate ENaC activity in recipient Xenopus distal tubule cells [3]. Borges et al. showed EVs released from injured tubular epithelial cells shuttle TGF-β1 mRNA and initiate tissue repair and regenerative responses [35]. A study by Jeon et al. showed that EVs released from damaged podocytes are enriched in various microRNAs that promote apoptosis of renal tubular epithelial cells [36]. It is evident that EVs can mediate changes in cellular function in an autocrine- or paracrine-dependent manner. In addition, EVs can modulate gene expression, control biological pathways, and change the phenotype of cells.
The pathophysiology of hypertension and diabetes-associated hypertension is thought to involve crosstalk between different organ systems and cell types, and also changes in intracellular signaling pathways. The crosstalk between various cell types that are affected during the pathophysiology of diabetes and hypertension can be mediated by bioactive and/or signaling molecules enriched in EVs.
3. Salt-Sensitive Hypertension and Enrichment of Blood-Pressure-Regulating Proteins in uEVs
The development of salt-sensitive hypertension is thought to be multifactorial with complicated genetic influences. Accordingly, the exact mechanisms that are responsible for the underlying pathogenesis are still not completely understood. However, it is generally accepted that kidney dysfunction plays an important role in the imbalance of fluid and electrolytes, and may contribute to the development of salt-sensitive hypertension. Several renal proteins that play an important role in electrolyte balance and blood pressure regulation including NCC, NKCC2, pendrin, and ENaC were shown to be enriched in human uEVs [37,38,39,40].
Several epithelial transport mechanisms have been studied in the context of the induction and maintenance of salt-sensitive hypertension. Multiple studies have shown db/db mice develop hypertension upon salt-loading [41,42,43,44]. A study by Scindia et al. showed that metformin mitigated high blood pressure in hypertensive diabetic db/db mice in a mechanism involving the attenuation of renal ENaC [43]. A study by Lugo et al. showed that alpha-1 antitrypsin (AAT) treatment by intraperitoneal injection normalized blood pressure in salt-loaded hypertensive diabetic db/db mice in a mechanism involving the attenuation of renal ENaC and MARCKS protein expression [42].
4. Association between Circulating EVs and Increased Blood Pressure
Published studies have shown that plasma/serum EVs and urinary EVs are associated with high blood pressure. The data from these studies suggest EVs may be a rich source of biomarkers for the pathogenesis and progression of hypertension. A study by Otani et al. showed that the intraperitoneal injection of plasma EVs from spontaneously hypertensive rats significantly increases systemic blood pressure in Wistar-Kyoto rats with normal blood pressure [45]. Conversely, this group showed that EVs from normotensive Wistar-Kyoto rats prevented an increase in systolic blood pressure in spontaneous hypertensive rats [45]. Additionally, this group also showed that plasma EVs from Wistar-Kyoto rats inhibited the increase in the prostaglandin F2α-induced contraction of the mesenteric artery and prevented perivascular fibrosis in spontaneously hypertensive rats [45]. A study by Good et al. showed that circulating EVs may contribute to the pathogenesis of hypertension and end-organ failure through the disruption of normal vascular tone [46]. This group showed that the functions of both resistance arteries and EVs in the systemic circulation are altered in spontaneously hypertensive rats 12 weeks of age. A study by Sun et al. showed that the circulating levels of endothelial microparticles may reflect renal microvascular capillary injury in patients with hypertension [47]. This group showed that the microparticle levels from renovascular hypertensive patients correlated directly with the histological peritubular capillary count, stenotic kidney hypoxia, and fibrosis, but inversely with cortical perfusion [47].
Previous studies have investigated EV surface biomarkers. Rai et al. investigated the surfaceome of small and large EVs [48]. These surface biomarkers may be important in the context of cellular injury and provide clinical insights into EV function. Moreover, the proteins enriched on the surface of EVs may provide clues to barrier dysfunction. For example, an increase in endothelial microparticles may reflect damage to the endothelium. Also, an increase in small EVs positive for platelet or endothelial cell markers in the urine may indicate a dysfunction of the glomerular filtration barrier.
5. Urinary Excretion of Extracellular Vesicles in Diabetes and Diabetic Nephropathy
uEVs are enriched in various lipids, metabolites, and proteins. These vesicles have been used to identify novel biomarkers in the pathophysiology of diabetes. The hydrophilic core and lipid bilayer of EVs protect genetic information from degradation and protects proteins from proteolysis. The packaged cargo of uEVs has been reported to change in conditions of hyperglycemia or high-glucose-induced injury. Multiple studies have identified specific miRNAs, mRNAs, and proteins in uEVs that can be useful in diagnosing diabetic nephropathy. Several studies have profiled various miRNAs in different diabetic patient subgroups or glucose conditions (Table 1). Jia et al. showed that the levels of miR-192, miR-194, and miR-215 are increased in type 2 diabetic patients with microalbuminuria compared to patients with normoalbuminuria and control subjects, but are decreased in patients with macroalbuminuria [49]. Mohan et al. reported that an elevation in renal miR-451-5p and miR-16 may have a protective effect in diabetes-induced renal fibrosis [50]. Prabu et al. demonstrated that miR-30a-5p concentrations were greater in uEVs isolated from macroalbuminuria patients but not patients with T2DM and normoalbuminuria or T2DM and microalbuminuria [51]. Delić et al. showed that miR-320c and miR-6068 are upregulated in uEVs from diabetic nephropathy patients [52]. Barutta et al. showed that miR-145 is increased in mesangial cells and exosomes derived from mesangial cells after high-glucose treatment [53]. Also, the proteins enriched in uEVs in diabetic patients or from diabetic db/db mice treated with various therapeutic agents have been studied. Kalani et al. showed that Wilms’ tumor 1 protein was enriched in uEVs from patients with proteinuria compared to those without proteinuria [54]. In a study by Zubiri et al., regucalcin protein was found to be downregulated in uEVs from rats with diabetic nephropathy [55]. A study by Musante et al. investigated the alterations in protease profiles in uEVs in diabetic nephropathy patients [56]. Lugo et al. showed that multiple hexosylceramides and other classes of lipids are increased in uEVs released from hypertensive diabetic db/db mice compared to control animals [42]. A study by Scindia et al. showed that uEVs from diabetic db/db mice are enriched in the cysteine protease cathepsin B, and the amount is attenuated after treating the mice with metformin compared to the vehicle [43]. In another study by Gholam et al., the concentration of EVs and their size were shown to be less in dapagliflozin-treated diabetic mice compared to vehicle-treated mice [57]. While it has been reported that the concentration of EVs is greater in urine from db/db mice with hyperglycemia or from conditioned media of various cell types [34,58] treated under high-glucose conditions, it appears this is not the case for all cell types. Wen et al. reported that high-glucose conditions reduced EV secretion from mouse kidney proximal tubule BUMPT cells [59]. Interestingly, this study showed that EVs from high-glucose-treated proximal tubule cells activate fibroblasts [59]. This group showed that several proteins were upregulated in the group of cells treated with high glucose compared to normal glucose.
Numerous animal models have been used to study mechanisms associated with the pathogenesis of diabetic nephropathy. The db/db mouse model is perhaps the most characterized and utilized model to study the many aspects that recapitulate human pathophysiology. Also, this mouse model exhibits similar functional and histological features associated with human metabolic dysfunction. These mice present with substantial albuminuria and mesangial matrix expansion. A study by Cohen et al. showed that diabetic nephropathy in db/db mice is ameliorated after inhibiting albumin glycation [60].
6. uEVs and Diabetes-Associated Hypertension
There is a high incidence of hypertension in patients with human type 2 diabetes. The presence of hypertension is known to worsen the complications associated with diabetes [61]. Db/db mice have become a popular model to study the various pathways and signaling cascades associated with diabetes-associated hypertension.
The db/db mouse model was established in 1966 by Jackson Labs. In the db/db mouse, there is point mutation in the leptin receptor gene. Mice homozygous for the db mutation are morbidly obese and diabetic within 8 weeks of age. Db/db mice recapitulate many of the symptoms seen in type 2 diabetic patients, including hyperglycemia, insulin resistance, compensatory hyperinsulinemia, and obesity. This animal model has provided important insights into the mechanisms of diabetes and obesity. There are multiple strains of the db/db mouse model, and the course of disease is greatly influenced by the genetic background.
The pathogenesis of disease is most aggressive in the C57BLKS/J background. Multiple studies have investigated blood pressure in db/db mice, and there are some differences between those studies. For example, some studies show that db/db mice have significantly elevated systolic and mean arterial pressures [62,63], while another study showed a reduced arterial pressure in db/db mice compared to wild-type mice [64]. This could be attributed to the different genetic backgrounds of db/db mice, various diets, and the range of ages that were used in those studies. For example, db/db mice used in a study by Su et al. were of a C57BL/KsJ background with a plasma glucose of 537.3 ± 33.3 mg/dL and elevated systolic, diastolic, and mean arterial pressures, while mice used in a study by Bodary et al. [64] had a plasma glucose of 340 mg/dL.
Db/db mice have become an appreciated model to investigate the mechanisms associated with the development of hypertension in the diabetic kidney. These mice develop profound hypertension after salt loading and have elevated ENaC protein expression and proteolysis [42,43]. Also, this model can be useful in investigating the crosstalk between multiple organ systems, including the cardiovascular and renal systems. For example, future studies may investigate how cardiac hormones regulate epithelial transport mechanisms in the kidney.
7. Relevance of Protease and Protease Inhibitor Enrichment in EVs to Diabetic Kidney Disease
Various proteases including cathepsins [43,65,66] and prostasin [37] and the versatile human alpha-1 antitrypsin (AAT) protein [67] have been found in EVs. Cathepsin B expression is known to increase with obesity and modulate inflammatory markers [68]. Therefore, it is not surprising that cathepsin B expression was found to be elevated in the kidney of hypertensive diabetic db/db mice that were strikingly obese. Cathepsin S levels are elevated in individuals with type 2 diabetes and obesity, but are reduced after weight loss [69]. Consistent with cathepsin S levels being increased in type 2 diabetes and obesity, the results from a study by Karimkhanloo et al. showed increased cathepsin S plasma levels in db/db mice compared to lean normoglycemic db/+ littermates [70]. In addition, this group showed that cathepsin S regulated glucose metabolism in cultured primary hepatocytes that were isolated from db/db mice but found that acute cathepsin S administration did not affect the plasma insulin levels or glucose tolerance of diabetic db/db mice [70].
A study by Hadler-Olsen showed there was an increased amount of gelatin-degrading serine proteinases in the plasma of db/db mice compared to db/+ control mice [71]. This study also showed that there was a greater amount of MMP-9 (gelatinase B) in the kidney extracts of db/+ control mice compared to db/db mice. Conversely, the expression of MMP-9 in the kidneys of KKay mice that developed nephropathy was augmented when compared to its expression in control mice.
The efficacy of exogenous kallikrein in ameliorating the effects of diabetic nephropathy in db/db mice was reported by Liu et al. [72]. In that study, inflammation, renal fibrosis, and oxidative stress were ameliorated in diabetic mice treated with kallikrein compared to diabetic controls [72]. In addition, exogenous kallikrein resulted in a decreased glomerular basement membrane thickness and protected against the loss of endothelial fenestrae, loss of podocytes, the effacement of foot processes in diabetic mice, and ultimately mitigated diabetic nephropathy [72].
8. Contribution of EVs in the Development of Insulin Resistance and Diabetes and Associated Complications
A study by Kumar et al. showed exosomes isolated from obese mice fed a high-fat diet or from type 2 diabetic patients caused lean mice to develop insulin resistance [73]. This group showed that CD63+A33+ exosomes are released from the intestinal epithelial cells of high-fat mice, and the uptake of these exosomes by liver cells is mediated by exosomal lipid composition. Moreover, they showed that the uptake of exosomes from high-fat-diet-fed mice by liver macrophages leads to the release of IL-6 and TNF-α. The results from this study showed that the CD63+A33+ exosomes induce insulin resistance and glucose intolerance in a mechanism involving the ligand-activated transcription factor AhR. A study by Abdelsaid et al. showed that plasma exosomes isolated from diabetic mice reduced angiogenic responses, while plasma exosomes from exercise trained T2DM mice restored the angiogenic effects in endothelial cells and in wound-healing models [74]. This group showed the mechanism by which exosomes mediated their effects involved an increase in the copper transporter ATP7A and the superoxide dismutase SOD3 protein in the plasma exosomes [74]. A study by Pillai et al. showed that the oxidation of LDL results in an increase in exosome secretion by adipocytes allowing for crosstalk with macrophages and the contribution of atherogenesis [75].
9. Circadian Regulation of EV Cargo Relevant to Diabetes Research
The mammalian circadian clock regulates many physiological and pathophysiological processes during light and dark cycles. Circadian rhythmicity is maintained by the positive regulators, circadian locomotor output cycles kaput (CLOCK) and brain and muscle aryl hydrocarbon receptor nuclear translocator-like 1 (BMAL1), and their co-repressors, period (PER) and cryptochrome (CRY). Importantly, the cargo enriched in EVs released by various cell types appears to vary over a 24 h cycle. Multiple studies have shown EV marker proteins are differentially enriched in uEVs isolated from urine samples during the inactive and active cycles of adult mice. A study by Lugo et al. showed differences in the amount of annexin A2, GAPDH, TSG101, and caveolin-1 proteins, as well as lipids including specific hexosylceramides, monoacylglycerols, phosphatidylcholines, lysophosphatidylethanolamine, phosphatidylethanolamine, and phosphatidylglycerol enriched in uEVs from male and female diabetic db/db mice treated with human AAT (hAAT) or the vehicle [42]. A study by Lopez et al. showed differences in the amounts of syntein and flotillin proteins, as well as lipids including specific phosphatidylethanolamines, triacylglycerols, and phosphatidylcholines enriched in uEVs from aged male mice with spontaneous hypertension [15]. Yeung et al. performed a functional enrichment analysis of circadian proteins and showed actin-binding and intermediate filament proteins associated with the cytoskeleton had peak abundances in small EVs that were temporally separated at 8 h and ~23 h after synchronization [76]. This group also showed that the various proteins that contribute to vesicle formation had peak abundances between 12 and 24 h after synchronization. Taken together, these studies suggest the biogenesis and packaging of cargo in uEVs are regulated in a circadian-dependent manner. Multiple groups have investigated the cargo enriched in EVs in a time-of-day-dependent manner. A study by Koritzinsky et al. suggested that the circadian variation in EV release can be normalized to TSG101 enrichment or by vesicle number [77]. Su et al. showed that hypertensive diabetic db/db mice have a disrupted heart rate, blood pressure, and locomotor activity that is associated with dampened oscillations of clock genes [78]. Grosbellet et al. showed a lengthened endogenous period altered photic integration in db/db mice [79]. Time-of-day variations in the vasocontractility of db/db mice have been previously investigated. Su et al. showed that normal time-of-day variations in vasocontractility to angiotensin II, phenylephrine, and high-potassium depolarization are lost in the aorta of db/db mice, and this is associated with alterations in the expression of clock genes or their oscillation phases [63]. Multiple groups have shown db/db mice display a nondipping phenotype [78,80,81]. It was suggested that this nondipping blood pressure phenotype in db/db mice is associated with the variations in vasocontractility and the disruption in vascular clock oscillation in these mice [63].
10. Renal Handling of EVs
Although there have been significant advances in the treatment of diabetic nephropathy in the past decade, it remains the leading cause of end-stage kidney disease. Alterations in cell types within the glomerulus is important in the pathogenesis of diabetic nephropathy [82].
The glomerulus consists of a glomerular basement membrane, fenestrated endothelium, and podocytes with distinct foot processes that allow for highly selective filtration. Various studies suggest that the cutoff for passive glomerular filtration is about 4.4–7.6 nm [83] or 30–50 kDa [84]. Due to the composition of these layers, large molecules and negatively charged molecules are not readily filterable. Molecular composition, size, and charge are important determinants of glomerular filtration. Interestingly, it has been shown that the renal elimination of specific molecules that are relatively large and have negative charges can be filtered and excreted [84]. Although most EVs that are present in the systemic circulation, including larger exosomes and microvesicles, are generally not filterable, smaller EVs may be filtered. The amount of EVs from the systemic circulation that is filtered likely increases when the glomerular filtration barrier is compromised as in diabetic nephropathy, as depicted in Figure 3. Also, larger EVs are filtered, and are either taken up by various cell types within the nephron or excreted in diabetic nephropathy, where there is a dysfunctional glomerular filtration barrier due to podocyte injury, foot process effacement, and/or podocyte loss, which contributes to the increased filtration of circulating EVs, as illustrated in Figure 3. EVs that are present in the tubule lumen can be taken up by any cell type in the nephron leading to cellular crosstalk and changes in intracellular signaling.
Polarized epithelial cells of the kidney that form the different segments of the nephron have an apical plasma membrane that faces the tubular lumen and a basolateral plasma membrane that faces the peritubular capillaries. All cell types in the nephron release EVs across the luminal/apical plasma membrane. However, it is unclear whether EVs are taken up by the peritubular capillaries after being released across the basolateral plasma membrane. In vitro studies imply EVs can be released from the luminal/apical plasma membrane or basolateral plasma membrane. For example, mouse cortical collecting duct cells grown on permeable supports were shown to release EVs across the luminal plasma membrane and the basolateral plasma membrane, and these two populations of EVs were found to be remarkably different in their lipid and protein composition [85]. These two populations of EVs that were released from the same mouse cortical collecting duct cells showed differential enrichment of proteins and lipids. Presumably, other cell types in the nephron release EVs with unique proteomic and lipidomic profiles. The sorting of urinary EVs and plasma EVs, and the characterization of the molecular cargo enriched in these EVs, from an animal model used to study pathophysiology, such as diabetic db/db mice, may allow for the identification of novel biomarkers important in disease mechanisms.
11. Proteomics of uEVs and EVs Released from Renal Cell Types
Specific proteins have been shown to be enriched in EVs isolated from glomerular cells and tubule cells (Table 2). Also, the phosphoproteomic profiling of uEVs has been reported [86]. Several proteins associated with renal diseases and hypertension have been identified and discussed in a review by Pisitkun et al. [40]. These proteins include aquaporin-2, epithelial sodium channel subunits, sodium chloride co-transporter, sodium potassium chloride co-transporter 2, carbonic anhydrase II and IV, polycystin-1, uromodulin, podocin, adenine phosphoribosyltransferase, nonmuscle myosin heavy chain IIA, FXYD domain-containing ion transport regulator-2, dimethylarginine dimethylaminohydrolase-1, neprilysin, hydroxyprostaglandin dehydrogenase, angiotensin I-converting enzyme isoform-1, and aminopeptidase N, P, and A. As summarized in Table 2, several studies have identified proteins enriched in EVs released from specific cell types including podocytes, proximal tubule cells, and principal cells [59,85,87,88,89].
12. uEVs as Biomarkers for Various Human Diseases
Interestingly, the quantification of specific apical membrane proteins in uEVs is not limited to renal diseases and hypertension. A study by Hayakawa et al. suggested that the quantification of ENaC gamma normalized to CD9 may be a biomarker for mineralocorticoid receptor (MR) activity in primary aldosteronism patients and perhaps other MR-associated diseases [90]. A study by Hu et al. suggested proteolytically cleaved products of ENaC alpha and gamma subunits normalized to CD9 were significantly enriched in uEVs from women with pre-eclampsia compared to those with a normal pregnancy [91], Additionally, the same study showed a phosphorylated form of NCC, at a T60 SPAK/OSR1 site, was significantly reduced in uEVs normalized to CD9 in the pre-eclampsia group compared to the normal pregnancy group [91]. A study by Wu et al. showed that mannan-binding lectin-associated serine protease 2 (MASP2) enrichment in uEVs from immunoglobulin A (IgA) nephropathy (IgAN) patients correlated with the level of urinary microalbumin and suggested MASP2 enrichment in uEVs may be a valuable biomarker for assessing the severity of renal injury as well as the prognosis of IgAN [92]. Taken together, these studies suggest uEVs may serve as noninvasive sources of novel biomarkers that can complement existing methods for diagnosing various diseases in humans.
13. Conclusions
The profiling of EVs isolated from the conditioned media of specific renal cell types cultured under high-glucose conditions or from the urine of animals or patients with hypertension coupled to diabetes or diabetic nephropathy may offer clues to the pathophysiology of these diseases. EVs released from various cell types in response to different stimuli or during the progression of diabetes-associated hypertension or diabetic nephropathy are packaged with bioactive cargo including metabolites, lipids, proteins, mRNAs, and miRNAs that contribute to cellular communication and changes in intracellular signaling. The isolation of plasma EVs and urinary EVs represent a rich source of biomarkers and prognosis markers, while engineered EVs may allow for the delivery of small drugs. The growing number of studies in EV research related to diabetes suggest these vesicles have great potential as tools to help elucidate the complex mechanisms associated with this disease.
Funding
The research reported in this publication was supported by a National Institute of Diabetes and Digestive and Kidney Diseases grant, R01 DK123078 (to AAA), and Centers for Disease Control and Prevention, National Institute for Occupational Safety and Health of the U.S. Department of Health and Human Services under Cooperative Agreement Award Number 6 U54OH011230-06-02. The content is solely the responsibility of the author and does not necessarily represent the official views of the Centers for Disease Control and Prevention or the Department of Health and Human Services.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No unpublished data are referenced in this review.
Conflicts of Interest
The author declares no conflict of interest.
References
- Merchant, M.L.; Rood, I.M.; Deegens, J.K.J.; Klein, J.B. Isolation and characterization of urinary extracellular vesicles: Implications for biomarker discovery. Nat. Rev. Nephrol. 2017, 13, 731–749. [Google Scholar] [CrossRef] [PubMed]
- Madison, R.D.; McGee, C.; Rawson, R.; Robinson, G.A. Extracellular vesicles from a muscle cell line (C2C12) enhance cell survival and neurite outgrowth of a motor neuron cell line (NSC-34). J. Extracell. Vesicles 2014, 3, 22865. [Google Scholar] [CrossRef] [PubMed]
- Jella, K.K.; Yu, L.; Yue, Q.; Friedman, D.; Duke, B.J.; Alli, A.A. Exosomal GAPDH from Proximal Tubule Cells Regulate ENaC Activity. PLoS ONE 2016, 11, e0165763. [Google Scholar] [CrossRef] [PubMed]
- Pais, V.; Pais, E.S. Intercellular communication by extracellular vesicles with emphasis on the roles of cordocytes in the human brain. An ultrastructural study. Ultrastruct. Pathol. 2015, 39, 177–186. [Google Scholar] [CrossRef]
- Zieske, J.D.; Hutcheon, A.E.K.; Guo, X. Extracellular Vesicles and Cell-Cell Communication in the Cornea. Anat. Rec. (Hoboken) 2020, 303, 1727–1734. [Google Scholar] [CrossRef]
- Chen, G.; Xu, C.; Gillette, T.G.; Huang, T.; Huang, P.; Li, Q.; Li, X.; Li, Q.; Ning, Y.; Tang, R.; et al. Cardiomyocyte-derived small extracellular vesicles can signal eNOS activation in cardiac microvascular endothelial cells to protect against Ischemia/Reperfusion injury. Theranostics 2020, 10, 11754–11774. [Google Scholar] [CrossRef]
- Pekkucuksen, N.T.; Liu, L.P.; Aly, R.; Shoemaker, L.R.; Alli, A.A. Extracellular vesicles from focal segmental glomerulosclerosis pediatric patients induce STAT3 activation and mesangial cell proliferation. PLoS ONE 2022, 17, e0274598. [Google Scholar] [CrossRef]
- Wei, X.; Ye, J.; Pei, Y.; Wang, C.; Yang, H.; Tian, J.; Si, G.; Ma, Y.; Wang, K.; Liu, G. Extracellular vesicles from colorectal cancer cells promote metastasis via the NOD1 signalling pathway. J. Extracell. Vesicles 2022, 11, e12264. [Google Scholar] [CrossRef]
- Ditte, Z.; Silbern, I.; Ditte, P.; Urlaub, H.; Eichele, G. Extracellular vesicles derived from the choroid plexus trigger the differentiation of neural stem cells. J. Extracell. Vesicles 2022, 11, e12276. [Google Scholar] [CrossRef]
- Ito, A.; Kagawa, S.; Sakamoto, S.; Kuwada, K.; Kajioka, H.; Yoshimoto, M.; Kikuchi, S.; Kuroda, S.; Yoshida, R.; Tazawa, H.; et al. Extracellular vesicles shed from gastric cancer mediate protumor macrophage differentiation. BMC Cancer 2021, 21, 102. [Google Scholar] [CrossRef]
- Ma, Q.; Beal, J.R.; Bhurke, A.; Kannan, A.; Yu, J.; Taylor, R.N.; Bagchi, I.C.; Bagchi, M.K. Extracellular vesicles secreted by human uterine stromal cells regulate decidualization, angiogenesis, and trophoblast differentiation. Proc. Natl. Acad. Sci. USA 2022, 119, e2200252119. [Google Scholar] [CrossRef] [PubMed]
- Alter, C.L.; Detampel, P.; Schefer, R.B.; Lotter, C.; Hauswirth, P.; Puligilla, R.D.; Weibel, V.J.; Schenk, S.H.; Heusermann, W.; Schurz, M.; et al. High efficiency preparation of monodisperse plasma membrane derived extracellular vesicles for therapeutic applications. Commun. Biol. 2023, 6, 478. [Google Scholar] [CrossRef]
- Antich-Rossello, M.; Munar-Bestard, M.; Forteza-Genestra, M.A.; Calvo, J.; Gaya, A.; Monjo, M.; Ramis, J.M. Evaluation of Platelet-Derived Extracellular Vesicles in Gingival Fibroblasts and Keratinocytes for Periodontal Applications. Int. J. Mol. Sci. 2022, 23, 7668. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Li, Y.J.; Hu, X.B.; Huang, S.; Xiang, D.X. Preservation of small extracellular vesicles for functional analysis and therapeutic applications: A comparative evaluation of storage conditions. Drug Deliv. 2021, 28, 162–170. [Google Scholar] [CrossRef]
- Lopez, J.P.; Nouri, M.Z.; Ebrahim, A.; Chacko, K.M.; Schramm, W.C.; Gholam, M.F.; Ozrazgat-Baslanti, T.; Denslow, N.D.; Alli, A.A. Lipid Profiles of Urinary Extracellular Vesicles Released during the Inactive and Active Phases of Aged Male Mice with Spontaneous Hypertension. Int. J. Mol. Sci. 2022, 23, 15397. [Google Scholar] [CrossRef]
- Darwish, S.; Liu, L.P.; Robinson, T.O.; Tarugu, S.; Owings, A.H.; Glover, S.C.; Alli, A.A. COVID-19 Plasma Extracellular Vesicles Increase the Density of Lipid Rafts in Human Small Airway Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 1654. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, S.; Jedrychowski, M.P.; Yanamandra, K.; Ikezu, S.; Gygi, S.P.; Ikezu, T. Proteomic Profiling of Extracellular Vesicles Derived from Cerebrospinal Fluid of Alzheimer’s Disease Patients: A Pilot Study. Cells 2020, 9, 1959. [Google Scholar] [CrossRef]
- Sequeiros, T.; Rigau, M.; Chiva, C.; Montes, M.; Garcia-Grau, I.; Garcia, M.; Diaz, S.; Celma, A.; Bijnsdorp, I.; Campos, A.; et al. Targeted proteomics in urinary extracellular vesicles identifies biomarkers for diagnosis and prognosis of prostate cancer. Oncotarget 2017, 8, 4960–4976. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, X.; Yang, X.; Jiang, Y.; Li, A.; Cong, J.; Li, Y.; Xie, Q.; Xu, C.; Liu, D. Identification of faecal extracellular vesicles as novel biomarkers for the non-invasive diagnosis and prognosis of colorectal cancer. J. Extracell. Vesicles 2023, 12, e12300. [Google Scholar] [CrossRef]
- Kim, S.; Kang, J.H.; Nguyen Cao, T.G.; Kang, S.J.; Jeong, K.; Kang, H.C.; Kwon, Y.J.; Rhee, W.J.; Ko, Y.T.; Shim, M.S. Extracellular vesicles with high dual drug loading for safe and efficient combination chemo-phototherapy. Biomater. Sci. 2022, 10, 2817–2830. [Google Scholar] [CrossRef]
- Niu, W.; Xiao, Q.; Wang, X.; Zhu, J.; Li, J.; Liang, X.; Peng, Y.; Wu, C.; Lu, R.; Pan, Y.; et al. A Biomimetic Drug Delivery System by Integrating Grapefruit Extracellular Vesicles and Doxorubicin-Loaded Heparin-Based Nanoparticles for Glioma Therapy. Nano Lett. 2021, 21, 1484–1492. [Google Scholar] [CrossRef]
- Mishra, D.D.; Sahoo, B.; Maurya, P.K.; Sharma, R.; Varughese, S.; Prasad, N.; Tiwari, S. Therapeutic potential of urine exosomes derived from rats with diabetic kidney disease. Front. Endocrinol. 2023, 14, 1157194. [Google Scholar] [CrossRef]
- Kontopoulou, E.; Strachan, S.; Reinhardt, K.; Kunz, F.; Walter, C.; Walkenfort, B.; Jastrow, H.; Hasenberg, M.; Giebel, B.; von Neuhoff, N.; et al. Evaluation of dsDNA from extracellular vesicles (EVs) in pediatric AML diagnostics. Ann. Hematol. 2020, 99, 459–475. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Yang, H.; Chen, T.; Jin, J.; Ruan, L.; Hu, L.; Chen, L. Extracellular vesicles metabolic changes reveals plasma signature in stage-dependent diabetic kidney disease. Ren. Fail. 2022, 44, 1840–1849. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Jo, H.; Park, S.; Kim, H.; Kim, S.I.; Han, Y.; Lee, J.; Seol, A.; Kim, J.; Lee, M.; et al. Integrated analysis of ascites and plasma extracellular vesicles identifies a miRNA-based diagnostic signature in ovarian cancer. Cancer Lett. 2022, 542, 215735. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Pan, B.; Zeng, F.; He, B.; Gao, Y.; Liu, X.; Song, Y. Magnetic Colloid Antibodies Accelerate Small Extracellular Vesicles Isolation for Point-of-Care Diagnostics. Nano Lett. 2021, 21, 2001–2009. [Google Scholar] [CrossRef]
- Dooley, K.; McConnell, R.E.; Xu, K.; Lewis, N.D.; Haupt, S.; Youniss, M.R.; Martin, S.; Sia, C.L.; McCoy, C.; Moniz, R.J.; et al. A versatile platform for generating engineered extracellular vesicles with defined therapeutic properties. Mol. Ther. 2021, 29, 1729–1743. [Google Scholar] [CrossRef]
- Esteves, M.; Abreu, R.; Fernandes, H.; Serra-Almeida, C.; Martins, P.A.T.; Barao, M.; Cristovao, A.C.; Saraiva, C.; Ferreira, R.; Ferreira, L.; et al. MicroRNA-124-3p-enriched small extracellular vesicles as a therapeutic approach for Parkinson’s disease. Mol. Ther. 2022, 30, 3176–3192. [Google Scholar] [CrossRef]
- Lu, M.; Huang, B.; Hanash, S.M.; Onuchic, J.N.; Ben-Jacob, E. Modeling putative therapeutic implications of exosome exchange between tumor and immune cells. Proc. Natl. Acad. Sci. USA 2014, 111, E4165–E4174. [Google Scholar] [CrossRef] [PubMed]
- Trac, D.; Hoffman, J.R.; Bheri, S.; Maxwell, J.T.; Platt, M.O.; Davis, M.E. Predicting Functional Responses of Progenitor Cell Exosome Potential with Computational Modeling. Stem Cells Transl. Med. 2019, 8, 1212–1221. [Google Scholar] [CrossRef]
- Lennon, K.M.; Saftics, A.; Abuelreich, S.; Sahu, P.; Lehmann, H.I.; Maddox, A.L.; Bagabas, R.; Januzzi, J.L.; Van Keuren-Jensen, K.; Shah, R.; et al. Cardiac troponin T in extracellular vesicles as a novel biomarker in human cardiovascular disease. Clin. Transl. Med. 2022, 12, e979. [Google Scholar] [CrossRef] [PubMed]
- Paluschinski, M.; Loosen, S.; Kordes, C.; Keitel, V.; Kuebart, A.; Brandenburger, T.; Scholer, D.; Wammers, M.; Neumann, U.P.; Luedde, T.; et al. Extracellular Vesicles as Markers of Liver Function: Optimized Workflow for Biomarker Identification in Liver Disease. Int. J. Mol. Sci. 2023, 24, 9631. [Google Scholar] [CrossRef]
- Takizawa, K.; Ueda, K.; Sekiguchi, M.; Nakano, E.; Nishimura, T.; Kajiho, Y.; Kanda, S.; Miura, K.; Hattori, M.; Hashimoto, J.; et al. Urinary extracellular vesicles signature for diagnosis of kidney disease. iScience 2022, 25, 105416. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.M.; Gao, Y.B.; Cui, F.Q.; Zhang, N. Exosomes from high glucose-treated glomerular endothelial cells activate mesangial cells to promote renal fibrosis. Biol. Open 2016, 5, 484–491. [Google Scholar] [CrossRef]
- Borges, F.T.; Melo, S.A.; Ozdemir, B.C.; Kato, N.; Revuelta, I.; Miller, C.A.; Gattone, V.H., 2nd; LeBleu, V.S.; Kalluri, R. TGF-beta1-containing exosomes from injured epithelial cells activate fibroblasts to initiate tissue regenerative responses and fibrosis. J. Am. Soc. Nephrol. 2013, 24, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.S.; Kim, E.; Bae, Y.U.; Yang, W.M.; Lee, H.; Kim, H.; Noh, H.; Han, D.C.; Ryu, S.; Kwon, S.H. microRNA in Extracellular Vesicles Released by Damaged Podocytes Promote Apoptosis of Renal Tubular Epithelial Cells. Cells 2020, 9, 1409. [Google Scholar] [CrossRef] [PubMed]
- Castagna, A.; Pizzolo, F.; Chiecchi, L.; Morandini, F.; Channavajjhala, S.K.; Guarini, P.; Salvagno, G.; Olivieri, O. Circadian exosomal expression of renal thiazide-sensitive NaCl cotransporter (NCC) and prostasin in healthy individuals. Proteomics Clin. Appl. 2015, 9, 623–629. [Google Scholar] [CrossRef]
- Ochiai-Homma, F.; Kuribayashi-Okuma, E.; Tsurutani, Y.; Ishizawa, K.; Fujii, W.; Odajima, K.; Kawagoe, M.; Tomomitsu, Y.; Murakawa, M.; Asakawa, S.; et al. Characterization of pendrin in urinary extracellular vesicles in a rat model of aldosterone excess and in human primary aldosteronism. Hypertens. Res. 2021, 44, 1557–1567. [Google Scholar] [CrossRef]
- Pathare, G.; Dhayat, N.; Mohebbi, N.; Wagner, C.A.; Cheval, L.; Neuhaus, T.J.; Fuster, D.G. Acute regulated expression of pendrin in human urinary exosomes. Pflugers Arch. 2018, 470, 427–438. [Google Scholar] [CrossRef]
- Pisitkun, T.; Johnstone, R.; Knepper, M.A. Discovery of urinary biomarkers. Mol. Cell. Proteom. 2006, 5, 1760–1771. [Google Scholar] [CrossRef]
- Hirohama, D.; Nishimoto, M.; Ayuzawa, N.; Kawarazaki, W.; Fujii, W.; Oba, S.; Shibata, S.; Marumo, T.; Fujita, T. Activation of Rac1-Mineralocorticoid Receptor Pathway Contributes to Renal Injury in Salt-Loaded db/db Mice. Hypertension 2021, 78, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Lugo, C.I.; Liu, L.P.; Bala, N.; Morales, A.G.; Gholam, M.F.; Abchee, J.C.; Elmoujahid, N.; Elshikha, A.S.; Avdiaj, R.; Searcy, L.A.; et al. Human Alpha-1 Antitrypsin Attenuates ENaC and MARCKS and Lowers Blood Pressure in Hypertensive Diabetic db/db Mice. Biomolecules 2022, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Scindia, Y.M.; Gholam, M.F.; Waleed, A.; Liu, L.P.; Chacko, K.M.; Desai, D.; Lopez, J.P.; Malik, Z.; Schramm, W.C.; Morales, A.G.; et al. Metformin Alleviates Diabetes-Associated Hypertension by Attenuating the Renal Epithelial Sodium Channel. Biomedicines 2023, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Veiras, L.C.; Shen, J.Z.Y.; Bernstein, E.A.; Regis, G.C.; Cao, D.; Okwan-Duodu, D.; Khan, Z.; Gibb, D.R.; Dominici, F.P.; Bernstein, K.E.; et al. Renal Inflammation Induces Salt Sensitivity in Male db/db Mice through Dysregulation of ENaC. J. Am. Soc. Nephrol. 2021, 32, 1131–1149. [Google Scholar] [CrossRef]
- Otani, K.; Yokoya, M.; Kodama, T.; Hori, K.; Matsumoto, K.; Okada, M.; Yamawaki, H. Plasma exosomes regulate systemic blood pressure in rats. Biochem. Biophys. Res. Commun. 2018, 503, 776–783. [Google Scholar] [CrossRef]
- Good, M.E.; Musante, L.; La Salvia, S.; Howell, N.L.; Carey, R.M.; Le, T.H.; Isakson, B.E.; Erdbrugger, U. Circulating Extracellular Vesicles in Normotension Restrain Vasodilation in Resistance Arteries. Hypertension 2020, 75, 218–228. [Google Scholar] [CrossRef]
- Sun, I.O.; Santelli, A.; Abumoawad, A.; Eirin, A.; Ferguson, C.M.; Woollard, J.R.; Lerman, A.; Textor, S.C.; Puranik, A.S.; Lerman, L.O. Loss of Renal Peritubular Capillaries in Hypertensive Patients Is Detectable by Urinary Endothelial Microparticle Levels. Hypertension 2018, 72, 1180–1188. [Google Scholar] [CrossRef]
- Rai, A.; Fang, H.; Claridge, B.; Simpson, R.J.; Greening, D.W. Proteomic dissection of large extracellular vesicle surfaceome unravels interactive surface platform. J. Extracell. Vesicles 2021, 10, e12164. [Google Scholar] [CrossRef]
- Jia, Y.; Guan, M.; Zheng, Z.; Zhang, Q.; Tang, C.; Xu, W.; Xiao, Z.; Wang, L.; Xue, Y. miRNAs in Urine Extracellular Vesicles as Predictors of Early-Stage Diabetic Nephropathy. J. Diabetes Res. 2016, 2016, 7932765. [Google Scholar] [CrossRef]
- Mohan, A.; Singh, R.S.; Kumari, M.; Garg, D.; Upadhyay, A.; Ecelbarger, C.M.; Tripathy, S.; Tiwari, S. Urinary Exosomal microRNA-451-5p Is a Potential Early Biomarker of Diabetic Nephropathy in Rats. PLoS ONE 2016, 11, e0154055. [Google Scholar] [CrossRef]
- Prabu, P.; Rome, S.; Sathishkumar, C.; Gastebois, C.; Meugnier, E.; Mohan, V.; Balasubramanyam, M. MicroRNAs from urinary extracellular vesicles are non-invasive early biomarkers of diabetic nephropathy in type 2 diabetes patients with the ‘Asian Indian phenotype’. Diabetes Metab. 2019, 45, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Delic, D.; Eisele, C.; Schmid, R.; Baum, P.; Wiech, F.; Gerl, M.; Zimdahl, H.; Pullen, S.S.; Urquhart, R. Urinary Exosomal miRNA Signature in Type II Diabetic Nephropathy Patients. PLoS ONE 2016, 11, e0150154. [Google Scholar] [CrossRef] [PubMed]
- Barutta, F.; Tricarico, M.; Corbelli, A.; Annaratone, L.; Pinach, S.; Grimaldi, S.; Bruno, G.; Cimino, D.; Taverna, D.; Deregibus, M.C.; et al. Urinary exosomal microRNAs in incipient diabetic nephropathy. PLoS ONE 2013, 8, e73798. [Google Scholar] [CrossRef]
- Kalani, A.; Mohan, A.; Godbole, M.M.; Bhatia, E.; Gupta, A.; Sharma, R.K.; Tiwari, S. Wilm’s tumor-1 protein levels in urinary exosomes from diabetic patients with or without proteinuria. PLoS ONE 2013, 8, e60177. [Google Scholar] [CrossRef]
- Zubiri, I.; Posada-Ayala, M.; Benito-Martin, A.; Maroto, A.S.; Martin-Lorenzo, M.; Cannata-Ortiz, P.; de la Cuesta, F.; Gonzalez-Calero, L.; Barderas, M.G.; Fernandez-Fernandez, B.; et al. Kidney tissue proteomics reveals regucalcin downregulation in response to diabetic nephropathy with reflection in urinary exosomes. Transl. Res. 2015, 166, 474–484.e4. [Google Scholar] [CrossRef]
- Musante, L.; Tataruch, D.; Gu, D.; Liu, X.; Forsblom, C.; Groop, P.H.; Holthofer, H. Proteases and protease inhibitors of urinary extracellular vesicles in diabetic nephropathy. J. Diabetes Res. 2015, 2015, 289734. [Google Scholar] [CrossRef]
- Gholam, M.F.; Liu, L.P.; Searcy, L.A.; Denslow, N.D.; Alli, A.A. Dapagliflozin Treatment Augments Bioactive Phosphatidylethanolamine Concentrations in Kidney Cortex Membrane Fractions of Hypertensive Diabetic db/db Mice and Alters the Density of Lipid Rafts in Mouse Proximal Tubule Cells. Int. J. Mol. Sci. 2023, 24, 1408. [Google Scholar] [CrossRef]
- Rice, G.E.; Scholz-Romero, K.; Sweeney, E.; Peiris, H.; Kobayashi, M.; Duncombe, G.; Mitchell, M.D.; Salomon, C. The Effect of Glucose on the Release and Bioactivity of Exosomes From First Trimester Trophoblast Cells. J. Clin. Endocrinol. Metab. 2015, 100, E1280–E1288. [Google Scholar] [CrossRef]
- Wen, J.; Ma, Z.; Livingston, M.J.; Zhang, W.; Yuan, Y.; Guo, C.; Liu, Y.; Fu, P.; Dong, Z. Decreased secretion and profibrotic activity of tubular exosomes in diabetic kidney disease. Am. J. Physiol. Renal Physiol. 2020, 319, F664–F673. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.P.; Masson, N.; Hud, E.; Ziyadeh, F.; Han, D.C.; Clements, R.S. Inhibiting albumin glycation ameliorates diabetic nephropathy in the db/db mouse. Exp. Nephrol. 2000, 8, 135–143. [Google Scholar] [CrossRef]
- Stas, S.N.; El-Atat, F.A.; Sowers, J.R. Pathogenesis of hypertension in diabetes. Rev. Endocr. Metab. Disord. 2004, 5, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Bagi, Z.; Erdei, N.; Toth, A.; Li, W.; Hintze, T.H.; Koller, A.; Kaley, G. Type 2 diabetic mice have increased arteriolar tone and blood pressure: Enhanced release of COX-2-derived constrictor prostaglandins. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1610–1616. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Xie, Z.; Guo, Z.; Duncan, M.J.; Lutshumba, J.; Gong, M.C. Altered clock gene expression and vascular smooth muscle diurnal contractile variations in type 2 diabetic db/db mice. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H621–H633. [Google Scholar] [CrossRef] [PubMed]
- Bodary, P.F.; Shen, Y.; Ohman, M.; Bahrou, K.L.; Vargas, F.B.; Cudney, S.S.; Wickenheiser, K.J.; Myers, M.G., Jr.; Eitzman, D.T. Leptin regulates neointima formation after arterial injury through mechanisms independent of blood pressure and the leptin receptor/STAT3 signaling pathways involved in energy balance. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 70–76. [Google Scholar] [CrossRef]
- Bellini, S.; Saraceno, C.; Benussi, L.; Geviti, A.; Longobardi, A.; Nicsanu, R.; Cimini, S.; Ricci, M.; Canafoglia, L.; Coppola, C.; et al. Plasma Small Extracellular Vesicle Cathepsin D Dysregulation in GRN/C9orf72 and Sporadic Frontotemporal Lobar Degeneration. Int. J. Mol. Sci. 2022, 23, 10693. [Google Scholar] [CrossRef]
- Downs, C.A.; Dang, V.D.; Johnson, N.M.; Denslow, N.D.; Alli, A.A. Hydrogen Peroxide Stimulates Exosomal Cathepsin B Regulation of the Receptor for Advanced Glycation End-Products (RAGE). J. Cell. Biochem. 2018, 119, 599–606. [Google Scholar] [CrossRef]
- Khodayari, N.; Oshins, R.; Alli, A.A.; Tuna, K.M.; Holliday, L.S.; Krotova, K.; Brantly, M. Modulation of calreticulin expression reveals a novel exosome-mediated mechanism of Z variant alpha(1)-antitrypsin disposal. J. Biol. Chem. 2019, 294, 6240–6252. [Google Scholar] [CrossRef]
- Araujo, T.F.; Cordeiro, A.V.; Vasconcelos, D.A.A.; Vitzel, K.F.; Silva, V.R.R. The role of cathepsin B in autophagy during obesity: A systematic review. Life Sci. 2018, 209, 274–281. [Google Scholar] [CrossRef]
- Taleb, S.; Cancello, R.; Poitou, C.; Rouault, C.; Sellam, P.; Levy, P.; Bouillot, J.L.; Coussieu, C.; Basdevant, A.; Guerre-Millo, M.; et al. Weight loss reduces adipose tissue cathepsin S and its circulating levels in morbidly obese women. J. Clin. Endocrinol. Metab. 2006, 91, 1042–1047. [Google Scholar] [CrossRef]
- Karimkhanloo, H.; Keenan, S.N.; Sun, E.W.; Wattchow, D.A.; Keating, D.J.; Montgomery, M.K.; Watt, M.J. Circulating cathepsin S improves glycaemic control in mice. J. Endocrinol. 2021, 248, 167–179. [Google Scholar] [CrossRef]
- Hadler-Olsen, E.; Winberg, J.O.; Reinholt, F.P.; Larsen, T.; Uhlin-Hansen, L.; Jenssen, T.; Berg, E.; Kolset, S.O. Proteases in Plasma and Kidney of db/db Mice as Markers of Diabetes-Induced Nephropathy. ISRN Endocrinol. 2011, 2011, 832642. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yang, Y.; Liu, Y.; Lu, X.; Guo, S.; Wu, M.; Wang, M.; Yan, L.; Wang, Q.; Zhao, X.; et al. Exogenous kallikrein protects against diabetic nephropathy. Kidney Int. 2016, 90, 1023–1036. [Google Scholar] [CrossRef]
- Kumar, A.; Sundaram, K.; Mu, J.; Dryden, G.W.; Sriwastva, M.K.; Lei, C.; Zhang, L.; Qiu, X.; Xu, F.; Yan, J.; et al. High-fat diet-induced upregulation of exosomal phosphatidylcholine contributes to insulin resistance. Nat. Commun. 2021, 12, 213. [Google Scholar] [CrossRef] [PubMed]
- Abdelsaid, K.; Sudhahar, V.; Harris, R.A.; Das, A.; Youn, S.W.; Liu, Y.; McMenamin, M.; Hou, Y.; Fulton, D.; Hamrick, M.W.; et al. Exercise improves angiogenic function of circulating exosomes in type 2 diabetes: Role of exosomal SOD3. FASEB J. 2022, 36, e22177. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.S.; Pereira, D.G.; Zhang, J.; Huang, W.; Beg, M.A.; Knaack, D.A.; de Souza Goncalves, B.; Sahoo, D.; Silverstein, R.L.; Shapiro, J.I.; et al. Contribution of adipocyte Na/K-ATPase alpha1/CD36 signaling induced exosome secretion in response to oxidized LDL. Front. Cardiovasc. Med. 2023, 10, 1046495. [Google Scholar] [CrossRef]
- Yeung, C.C.; Dondelinger, F.; Schoof, E.M.; Georg, B.; Lu, Y.; Zheng, Z.; Zhang, J.; Hannibal, J.; Fahrenkrug, J.; Kjaer, M. Circadian regulation of protein cargo in extracellular vesicles. Sci. Adv. 2022, 8, eabc9061. [Google Scholar] [CrossRef]
- Koritzinsky, E.H.; Street, J.M.; Chari, R.R.; Glispie, D.M.; Bellomo, T.R.; Aponte, A.M.; Star, R.A.; Yuen, P.S.T. Circadian variation in the release of small extracellular vesicles can be normalized by vesicle number or TSG101. Am. J. Physiol. Renal Physiol. 2019, 317, F1098–F1110. [Google Scholar] [CrossRef]
- Su, W.; Guo, Z.; Randall, D.C.; Cassis, L.; Brown, D.R.; Gong, M.C. Hypertension and disrupted blood pressure circadian rhythm in type 2 diabetic db/db mice. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1634–H1641. [Google Scholar] [CrossRef]
- Grosbellet, E.; Dumont, S.; Schuster-Klein, C.; Guardiola-Lemaitre, B.; Pevet, P.; Criscuolo, F.; Challet, E. Circadian phenotyping of obese and diabetic db/db mice. Biochimie 2016, 124, 198–206. [Google Scholar] [CrossRef]
- Hou, T.; Su, W.; Guo, Z.; Gong, M.C. A Novel Diabetic Mouse Model for Real-Time Monitoring of Clock Gene Oscillation and Blood Pressure Circadian Rhythm. J. Biol. Rhythms 2019, 34, 51–68. [Google Scholar] [CrossRef]
- Senador, D.; Kanakamedala, K.; Irigoyen, M.C.; Morris, M.; Elased, K.M. Cardiovascular and autonomic phenotype of db/db diabetic mice. Exp. Physiol. 2009, 94, 648–658. [Google Scholar] [CrossRef]
- Tung, C.W.; Hsu, Y.C.; Shih, Y.H.; Chang, P.J.; Lin, C.L. Glomerular mesangial cell and podocyte injuries in diabetic nephropathy. Nephrology 2018, 23 (Suppl. S4), 32–37. [Google Scholar] [CrossRef] [PubMed]
- Bolten, J.S.; Pratsinis, A.; Alter, C.L.; Fricker, G.; Huwyler, J. Zebrafish (Danio rerio) larva as an in vivo vertebrate model to study renal function. Am. J. Physiol. Renal Physiol. 2022, 322, F280–F294. [Google Scholar] [CrossRef]
- Ruggiero, A.; Villa, C.H.; Bander, E.; Rey, D.A.; Bergkvist, M.; Batt, C.A.; Manova-Todorova, K.; Deen, W.M.; Scheinberg, D.A.; McDevitt, M.R. Paradoxical glomerular filtration of carbon nanotubes. Proc. Natl. Acad. Sci. USA 2010, 107, 12369–12374. [Google Scholar] [CrossRef]
- Dang, V.D.; Jella, K.K.; Ragheb, R.R.T.; Denslow, N.D.; Alli, A.A. Lipidomic and proteomic analysis of exosomes from mouse cortical collecting duct cells. FASEB J. 2017, 31, 5399–5408. [Google Scholar] [CrossRef]
- Gonzales, P.A.; Pisitkun, T.; Hoffert, J.D.; Tchapyjnikov, D.; Star, R.A.; Kleta, R.; Wang, N.S.; Knepper, M.A. Large-scale proteomics and phosphoproteomics of urinary exosomes. J. Am. Soc. Nephrol. 2009, 20, 363–379. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, K.; Lay, A.C.; Leparc, G.; Tran, V.D.T.; Rosler, M.; Dayalan, L.; Burdet, F.; Ibberson, M.; Coward, R.J.M.; Huber, T.B.; et al. An in vitro approach to understand contribution of kidney cells to human urinary extracellular vesicles. J. Extracell. Vesicles 2023, 12, e12304. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wilkinson, R.; Kildey, K.; Ungerer, J.P.J.; Hill, M.M.; Shah, A.K.; Mohamed, A.; Dutt, M.; Molendijk, J.; Healy, H.; et al. Molecular and functional profiling of apical versus basolateral small extracellular vesicles derived from primary human proximal tubular epithelial cells under inflammatory conditions. J. Extracell. Vesicles 2021, 10, e12064. [Google Scholar] [CrossRef]
- Prunotto, M.; Farina, A.; Lane, L.; Pernin, A.; Schifferli, J.; Hochstrasser, D.F.; Lescuyer, P.; Moll, S. Proteomic analysis of podocyte exosome-enriched fraction from normal human urine. J. Proteom. 2013, 82, 193–229. [Google Scholar] [CrossRef]
- Hayakawa, T.; Fukuhara, A.; Saiki, A.; Otsuki, M.; Shimomura, I. gammaENaC/CD9 in urinary extracellular vesicles as a potential biomarker of MR activity. J. Endocrinol. 2021, 252, 81–90. [Google Scholar] [CrossRef]
- Hu, C.C.; Katerelos, M.; Choy, S.W.; Crossthwaite, A.; Walker, S.P.; Pell, G.; Lee, M.; Cook, N.; Mount, P.F.; Paizis, K.; et al. Pre-eclampsia is associated with altered expression of the renal sodium transporters NKCC2, NCC and ENaC in urinary extracellular vesicles. PLoS ONE 2018, 13, e0204514. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.W.B.; Chen, Y.Y.; Huang, J.X.; Wang, K.Y.; Xu, H.S.; Lin, D. Significance of Mannan Binding Lectin-Associated Serine Protease 2 in Urinary Extracellular Vesicles in IgA Nephropathy. Clin. Investig. Med. 2022, 45, E47–E54. [Google Scholar] [CrossRef] [PubMed]
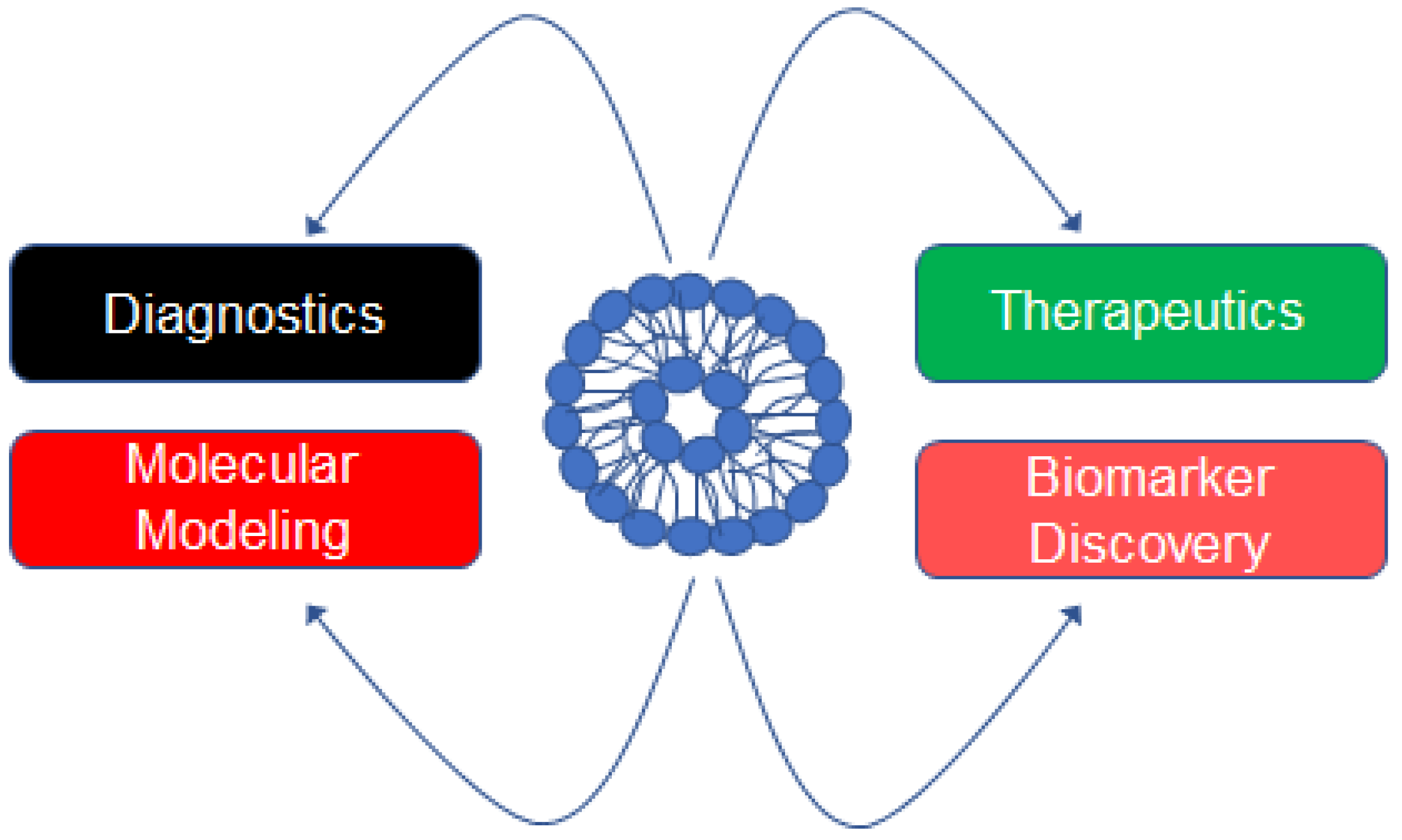
Figure 1.
Applications of EVs isolated from biological fluids or cell-conditioned media. EVs can be purified from biological fluids including plasma, serum, urine, or cell-conditioned media. EVs can be used as diagnostic markers and indicate the stage of disease. The molecules enriched in EVs can be subject to bioinformatic studies and used for computational or molecular modeling. Engineered EVs can be loaded with various molecules or drugs for targeted delivery to treat various diseases. EVs are a rich source of biomarkers in physiology and pathophysiology.
Figure 1.
Applications of EVs isolated from biological fluids or cell-conditioned media. EVs can be purified from biological fluids including plasma, serum, urine, or cell-conditioned media. EVs can be used as diagnostic markers and indicate the stage of disease. The molecules enriched in EVs can be subject to bioinformatic studies and used for computational or molecular modeling. Engineered EVs can be loaded with various molecules or drugs for targeted delivery to treat various diseases. EVs are a rich source of biomarkers in physiology and pathophysiology.

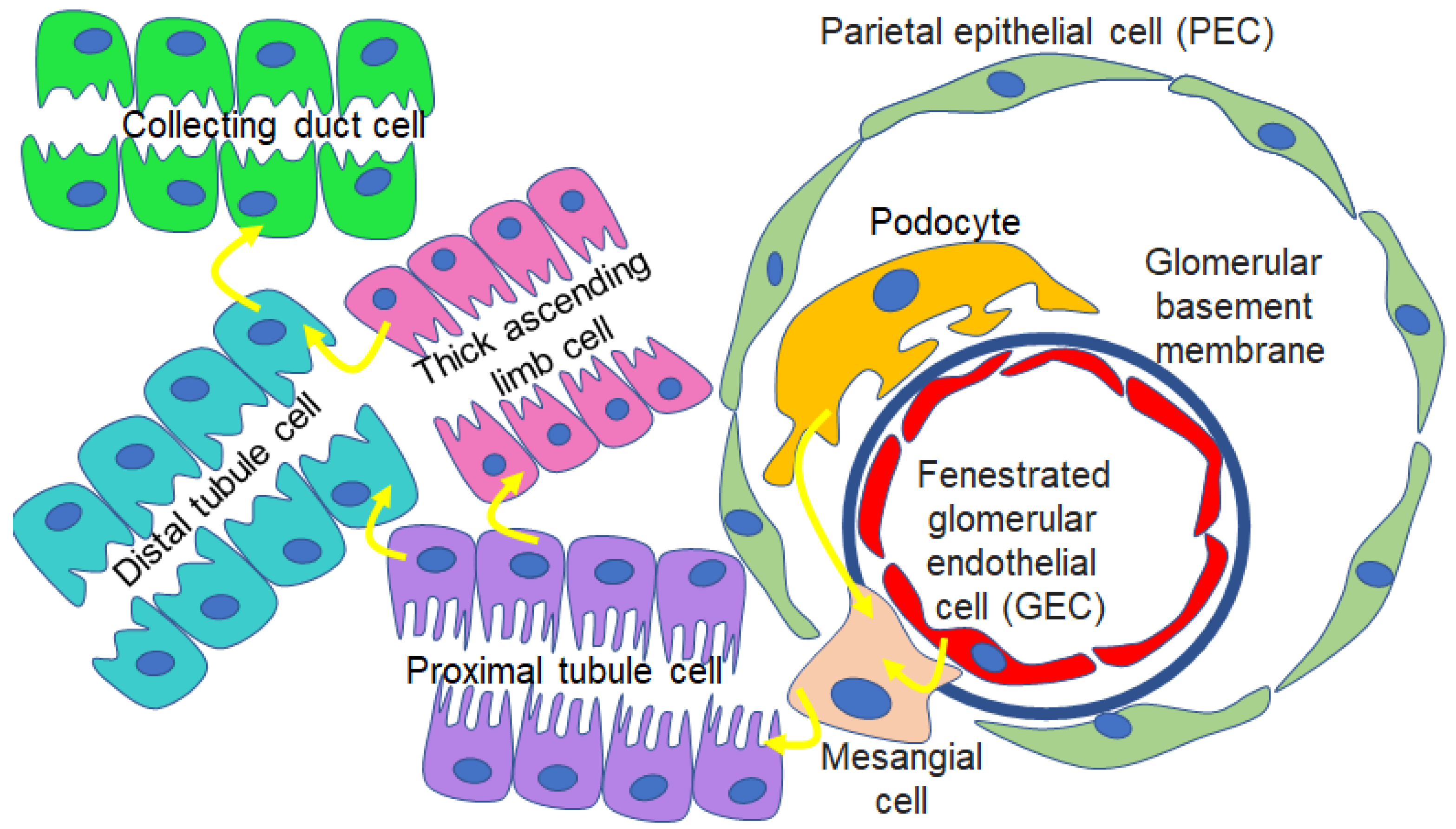
Figure 2.
EV-mediated crosstalk between various kidney cell types. EVs mediate intercellular communication between podocytes and mesangial cells, endothelial cells and mesangial cells, glomerular cells and tubular cells, and between other cell types within the segments of the nephron.
Figure 2.
EV-mediated crosstalk between various kidney cell types. EVs mediate intercellular communication between podocytes and mesangial cells, endothelial cells and mesangial cells, glomerular cells and tubular cells, and between other cell types within the segments of the nephron.

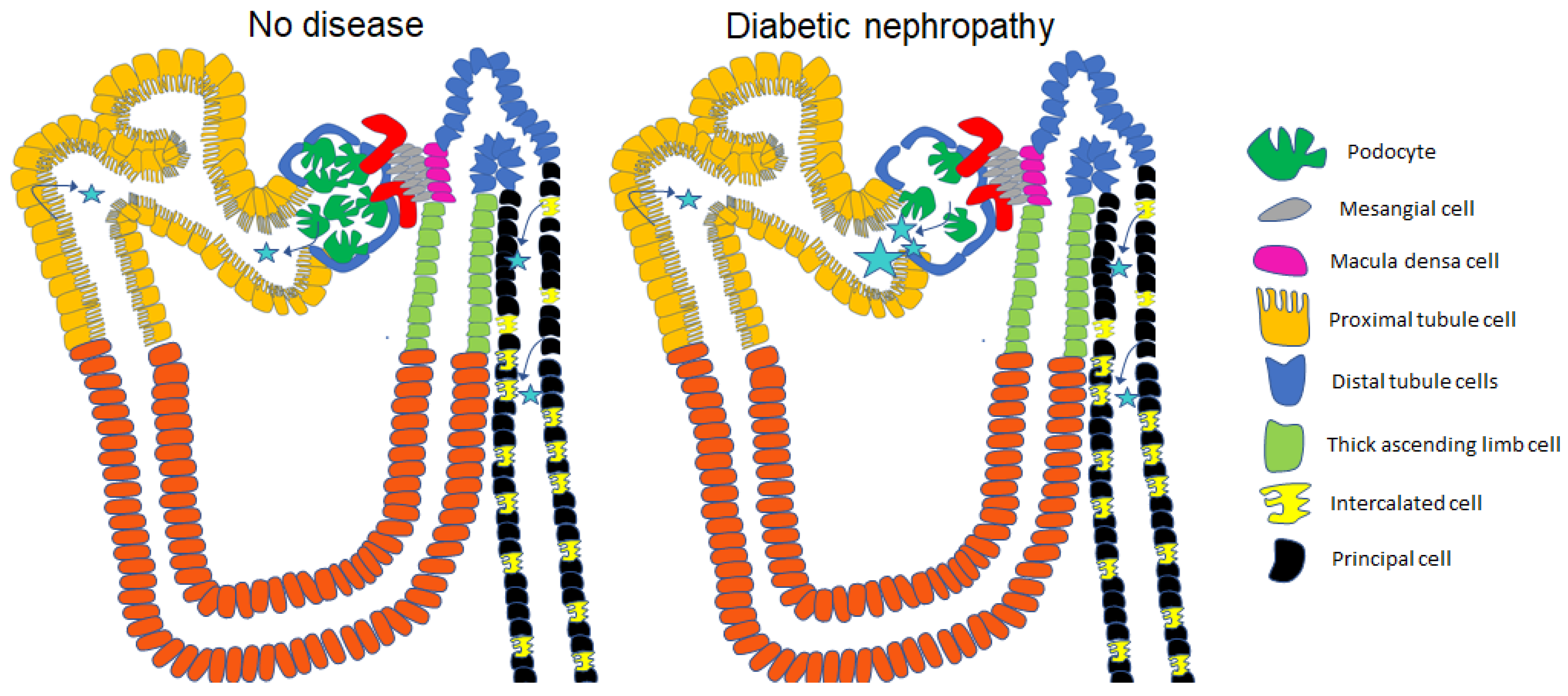
Figure 3.
A greater number of EVs from the systemic circulation is filtered across the glomerular filtration barrier with the progression of diabetic nephropathy compared to the health kidney in the absence of disease. Stars represent EVs. Arrows represent the cell types that release EVs into the lumen.
Figure 3.
A greater number of EVs from the systemic circulation is filtered across the glomerular filtration barrier with the progression of diabetic nephropathy compared to the health kidney in the absence of disease. Stars represent EVs. Arrows represent the cell types that release EVs into the lumen.


{kind=link}
{kind=link}
{kind=link}
Table 1.
miRs in diabetes and diabetic nephropathy.
| mIR | Finding | Reference |
|---|---|---|
| miR-192, miR-194, miR-215 | increased in type 2 diabetic patients with microalbuminuria compared to patients with normoalbuminuria | [49] |
| miR-451-5p, miR-16 | protective effect in diabetes-induced renal fibrosis | [50] |
| miR-30a-5p | greater in uEVs isolated from macroalbuminuria patients but not patients with T2DM and normoalbuminuria or T2DM and microalbuminuria | [51] |
| miR-320c, miR-6068 | upregulated in uEVs from diabetic nephropathy patients | [52] |
| miR-145 | increased in mesangial cells and exosomes derived from mesangial cells after high-glucose treatment | [53] |
Table 2.
Enrichment of various proteins in EVs released from specific kidney cell types.
| Cell Type | Podocytes | Mesangial Cells | Glomerular Endothelial Cells | Proximal Tubule Cells | Principal Cells |
|---|---|---|---|---|---|
| Podocalyxin, Nephrin (Prunotto et al. [89]) Proteasome subunit beta type-2, Cullin-2, Complement C3, Cystatin-A, Keratin, type I cytoskeletal 14, Putative phospholipase B-like 2, Endoplasmic reticulum resident protein 44 (Barreiro et al. [87]) | Creatine kinase, Cysteine-rich motor neuron 1 protein (Barreiro et al. [87]) | EGF containing fibulin-like extracellular matrix protein 1 isoform 1, Prosaposin, Pentraxin-related protein, Latenttransforming growth factor beta-binding protein 2, Thioredoxin reductase 1, cytoplasmic, Chondroitin sulphate proteoglycan 4 (Barreiro et al. [87]) | CD63, CD9, CD81, TSG101, ALIX, VPS4B, HSP70, HSP84, 60S ribosomal protein L27, V-set domain-containing T-cell activation inhibitor 1, Protein lin-7 homolog C, HLA class I histocompatibility antigen, A-24 alpha chain, TNFAIP3-interacting protein 1, Laminin subunit alpha-3, Beta (β)-2-macroglobulin, Interferon-induced transmembrane protein 1 TNF alpha-induced protein 3, Myosin-10, Serum amyloid A-1 protein, Intercellular adhesion molecule 1, C-X-C motif chemokine 10, Alcohol dehydrogenase, Hyaluronan and proteoglycan link protein 3, Guanylate-binding protein 5, Indoleamine 2,3-dioxygenase 1, Tryptophan–tRNA ligase, cytoplasmic 1, Interferon-induced guanylate-binding protein 1, Tumour necrosis factor receptor superfamily member 5, ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 1, Tumour necrosis factor alpha-induced protein (Wang et al. [88]) Annexin A5, Annexin A11, Adenylyl cyclase 7, GTPase-activating protein subunit alpha-2, Cofilin-2, Heat shock cognate 71 kDa protein, Phosphoglycerate kinase 1, 14-3-3 protein theta, 14-3-3 protein gamma, 14-3-3 protein zeta/delta, retinoic acid-induced protein 3 (Wen et al. [59]) Betahexosaminidase subunit beta, Elongation factor 1-gamma, Serine/threonineprotein phosphatase PP1-alpha catalytic subunit, Serine/threonineprotein phosphatase PP1-beta catalytic subunit, Brain acid soluble protein 1 (Barreiro et al. [87]) | α-actinin-1, moesin, 14-3-3 protein ζ/δ, annexin A1/A3/A4/A5/A6, clathrin heavy chain 1, GAPDH, α-enolase, filamin-A, HSP 90 (Dang et al. [85]) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alli, A.A. Extracellular Vesicles: Investigating the Pathophysiology of Diabetes-Associated Hypertension and Diabetic Nephropathy. Biology 2023, 12, 1138. https://doi.org/10.3390/biology12081138
AMA Style
Alli AA. Extracellular Vesicles: Investigating the Pathophysiology of Diabetes-Associated Hypertension and Diabetic Nephropathy. Biology. 2023; 12(8):1138. https://doi.org/10.3390/biology12081138
Chicago/Turabian StyleAlli, Abdel A. 2023. "Extracellular Vesicles: Investigating the Pathophysiology of Diabetes-Associated Hypertension and Diabetic Nephropathy" Biology 12, no. 8: 1138. https://doi.org/10.3390/biology12081138
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.