 Guangshuai Cui1,2
Guangshuai Cui1,2 Francisco I. Pugnaire2
Francisco I. Pugnaire2 Liu Yang1,3
Liu Yang1,3 Wanglin Zhao1
Wanglin Zhao1 Rita Ale1
Rita Ale1 Wei Shen1
Wei Shen1 Tianxiang Luo1
Tianxiang Luo1 Eryuan Liang1
Eryuan Liang1 Lin Zhang1,4*
Lin Zhang1,4*- 1State Key Laboratory of Tibetan Plateau Earth System Science, Resources and Environment (TPESRE), Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing, China
- 2Estación Experimental de Zonas Áridas, Consejo Superior de Investigaciones Científicas, Almería, Spain
- 3University of Chinese Academy of Sciences, Beijing, China
- 4Institute of Science and Technology Information of Tibet Autonomous Region, Lhasa, China
Introduction: Shrub promotes the survival, growth and reproduction of understory species by buffering the environmental extremes and improving limited resources (i.e., facilitation effect) in arid and semiarid regions. However, the importance of soil water and nutrient availability on shrub facilitation, and its trend along a drought gradient have been relatively less addressed in water-limited systems.
Methods: We investigated species richness, plant size, soil total nitrogen and dominant grass leaf δ13C within and outside the dominant leguminous cushion-like shrub Caragana versicolor along a water deficit gradient in drylands of Tibetan Plateau.
Results: We found that C. versicolor increased grass species richness but had a negative effect on annual and perennial forbs. Along the water deficit gradient, plant interaction assessed by species richness (RIIspecies) showed a unimodal pattern with shift from increase to decrease, while plant interaction assessed by plant size (RIIsize) did not vary significantly. The effect of C. versicolor on soil nitrogen, rather than water availability, determined its overall effect on understory species richness. Neither the effect of C. versicolor on soil nitrogen nor water availability affected plant size.
Discussion: Our study suggests that the drying tendency in association with the recent warming trends observed in drylands of Tibetan Plateau, will likely hinder the facilitation effect of nurse leguminous shrub on understories if moisture availability crosses a critical minimum threshold.
1 Introduction
Positive and negative plant-plant interactions co-occur in plant communities (Callaway and Walker, 1997). The negative effect (i.e., competition) always happens between intra- or inter-species when they sharing limited living space or resources. The positive effect, facilitation, has strong impact in most harsh environments, where the mitigation of extreme climate by facilitator species can benefit many other species (Bertness and Callaway, 1994; Pugnaire et al., 1996; Brooker et al., 2008). Facilitation acts as a sort of insurance policy for plant communities in alpine regions (Cavieres et al., 2014; Cavieres et al., 2016; Pistón et al., 2016) and water-limited systems (Armas et al., 2011; Pugnaire et al., 2011; Parajuli et al., 2021; Pugnaire et al., 2021). Improving our knowledge of facilitation is, therefore, crucial to understand plant community dynamics and to predict plant community responses to a changing climate (Anthelme et al., 2014; Pugnaire et al., 2021).
Shrubs often act as nurse plants in arid habitats where water availability and soil nutrients are limiting factors (Butterfield et al., 2016), increasing species diversity (O'Brien et al., 2019) through buffering environmental conditions, increasing resource availability and/or protecting against herbivores (Maestre et al., 2009; Tirado et al., 2015; Zhang et al., 2018a). Shrubs could increase soil water availability through hydraulic lift (Richards and Caldwell, 1987; Prieto et al., 2011) or shading effect (Liu et al., 2021). However, shrubs can also have negative effects on soil water content mostly through interception loss in shrub canopies and enhanced utilization of soil water (Tielbörger and Kadmon, 2000; Darrouzet-Nardi et al., 2006; Hamerlynck et al., 2011). Nevertheless, in arid environments the effects of shrubs on soil moisture depends on the relationship between precipitation and differential evapo-transpiration rates under plants and in gaps (Butterfield et al., 2016). Formation of fertility islands underneath shrubs canopy represents another mechanism involved in the positive shrub-herbaceous interaction in drylands, mainly due to enhanced soil nutrient availability (Zhang et al., 2018b; Cai et al., 2020). In particular, leguminous shrubs have a significant effect on soil nitrogen accruement (Pugnaire et al., 1996; Zhang et al., 2011; Saixiyala et al., 2017). Accumulation of soil organic matter can also buffer soil water and thermal oscillations in the understory (Wezel et al., 2000; Maestre et al., 2001; Noumi et al., 2016). However, the relative contribution of shrub mediated effect on soil water and nutrient availability in the interaction between shrub and understory species have been less addressed in drylands.
Plant-plant interactions changes with resource availability and abiotic conditions. The stress gradient hypothesis (SGH) predicts an increase in the importance of facilitation with increasing environmental severity (Bertness and Callaway, 1994). But other models suggest that facilitation is not always the case, that would tend to be neutral interactions or even a switch from facilitation to competition with increasing severity in arid and semi-arid systems (Tielbörger and Kadmon, 2000; Maestre and Cortina, 2004; Michalet et al., 2006; Holmgren and Scheffer, 2010; O'Brien et al., 2017). Inconsistent plant interactions patterns along biotic gradient has been reported when different measurements (e.g., species richness, plant height, or reproduction) or statistical levels (e.g., paired vs. community scale) are involved (He et al., 2013; Soliveres et al., 2015; Liancourt et al., 2017). In addition, species turnover along gradients may blur predictions (Choler et al., 2001; Liancourt et al., 2005; Le Bagousse-Pinguet et al., 2012; Liancourt et al., 2017).
We addressed the differential effects of facilitation within and outside the canopy of a shrub, Caragana versicolor, along a water availability gradient in west drylands of Tibetan Plateau. West Tibet, with a mean elevation above 4,500 m a.s.l., is characteristic of a low rainfall, high radiation, and barren soil environment. As the dominant shrub species in the western Himalayan regions, C. versicolor is adapted to these habitats through its cushion canopy, deep roots, and N-fixing capacity (Kumar et al., 2016). In this study, we aimed to evaluate the relative effects of soil moisture and soil nutrients on shrub-understories interactions in a water-limited system. Considering that different functional groups show different adaptive strategies which may lead to inconsistent patterns of facilitator-understories interactions (Iyengar et al., 2017; Liancourt et al., 2017), we also compared the different effects of shrub facilitation on different functional groups. Our aims are to test: (1) whether the facilitation effects of C. versicolor on associated species increase with decreasing water availability; (2) whether soil nutrients play a stronger role than soil moisture in driving shrub-understory interactions in this arid region, and (3) whether grasses show stronger responses to facilitation than annual and perennial forbs because of their higher competitive ability. According to the SGH, we expected that facilitation intensity would increase as water availability decreases and that nutrients are more important than water in driving plant-plant interactions. Finally, we expected that shrubs have stronger facilitation on grasses than on other functional groups.
2 Materials and methods
2.1 Study area and sampling design
The study was conducted in Shigatse and Ngari areas in the Tibet Autonomous Region, China where the climate is cold and arid with a mean annual temperature ranging -8~2°C and annual precipitation of 50~500 mm which falls mainly in summer. The plant community is dominated by alpine shrubs (shrub-steppe) and alpine pastures (desert steppe), dominated by species such as C. versicolor, Elymus nutans, Leymus spp., Stipa caucasica, and S. purpurea. C. versicolor is the dominant shrub species, generally spanning 4250~5100 m in elevation. It is a Fabaceae deciduous species that usually grows in well-drained and loose sandy soils, with a 20 cm mean canopy height and a cover ranging 10%~50%.
In August 2019, we selected 5 sites along a longitudinal transect within a naturally-occurring water availability gradient. The locations and elevations of the sampling sites were measured by GPS (eTrex Venture, Garmin, USA) (Figure 1). At each site, we investigated 30 shrubs and 30 paired open areas equal in size to that of shrubs within an area of approximately 1 ha. Shrub understory area was calculated as the area of an ellipse by measuring the largest diameter and its perpendicular. The shape of every shrub was simulated with wire, and the wire was randomly placed 1 meter near the shrub edge to investigate the paired open area. All vascular plants growing within the paired plots were identified, and the mean size of each species was measured. For the erect plants, which size were measured as individual height, the length of stem was measured as plant size instead for the prostrate species. A total of 150 paired plots were explored using the protocol described above.
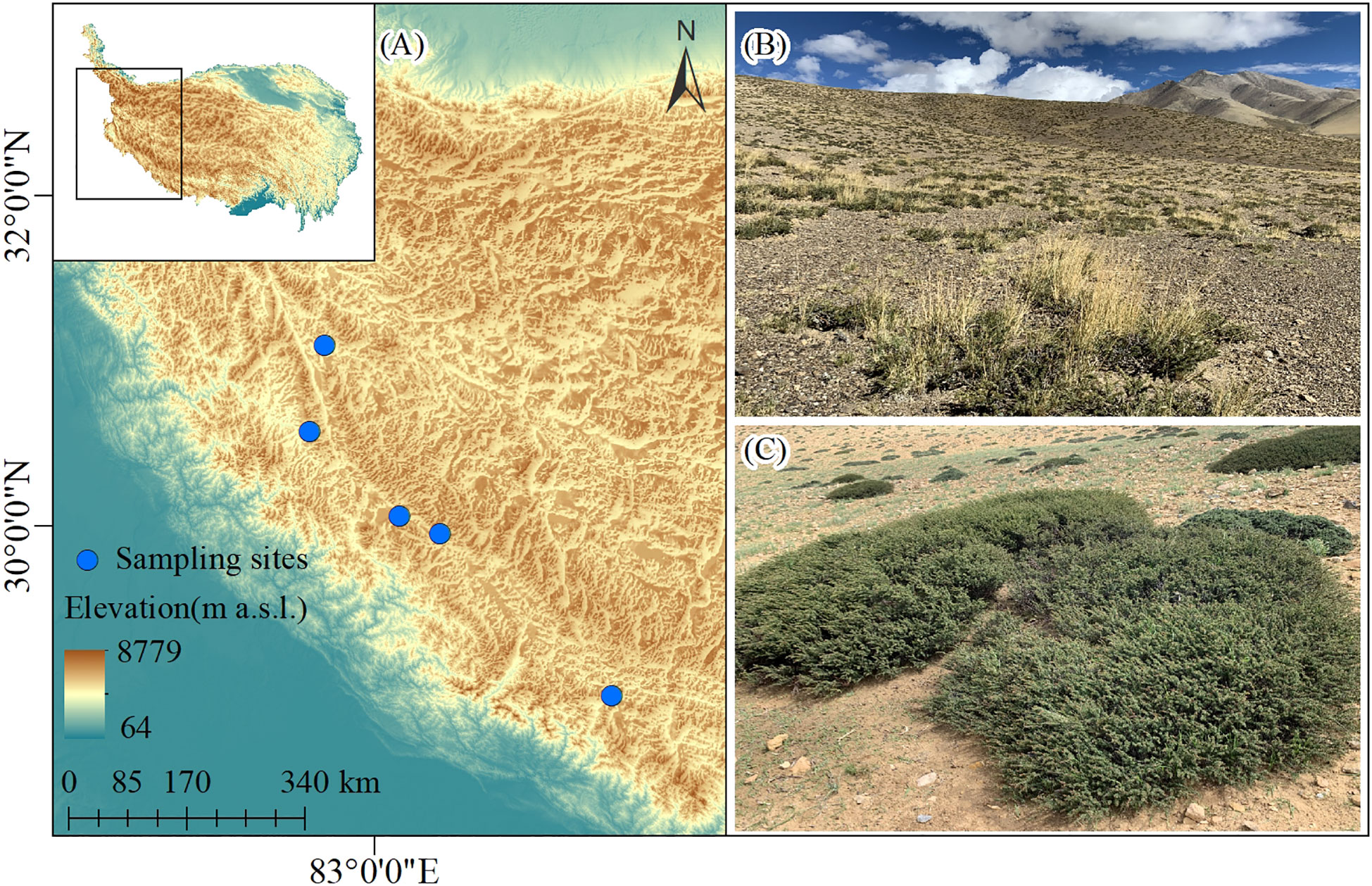
Figure 1 (A) A map showing the study area and sampling sites. Photos of the (B) Caragana versicolor community landscape and (C) shrub individuals in drylands of Tibetan Plateau.
In each site, the first 5 paired plots were selected for sampling soil (0~20 cm) and the same dominant grass leaves under C. versicolor and in the paired open area (Supplementary Table 1). Soil total nitrogen content was determined using an elemental analyzer (2400 II CHNS; Perkin Elmer, Boston, MA, USA). We failed to measure soil moisture across the whole water availability gradient, only one site located in the westernmost gained the soil moisture. Volumetric soil moisture at -10 cm underneath the shrub canopy and in the open area were measured with HOBO weather stations (Onset Inc., Bourne, MA, USA), which were recorded since Aguste 8th 2015 to Journey 1st 2020 with 1 hour interval, the missing data is due to equipment failure (Supplementary Figure 1). In C3 plants, stable carbon isotope composition in leaves (δ13C) can provide an integrated measurement of internal-physiological and external-environmental conditions during the growing season (Farquhar et al., 1989; Dawson et al., 2002). In dry habitats, leaf δ13C is highly sensitive to water availability, and Ale et al. (2018) found that widespread species leaf δ13C would be used as an indicator of water availability in dry mountainous regions like the Himalayas. In this study, leaf δ13C of dominant grass species occurring within and outside the shrub canopy was used to characterize soil water availability. The δ13C ratio in grasses leaves, relative to a Pee Dee Belemnite (PDB) standard, was determined with an elemental analyzer coupled to a stable isotope mass spectrometer (Flash EA + Delta V, Thermo Fisher Scientific Inc., Waltham, MA, USA) at the State Key Laboratory of Tibetan Plateau Earth System Science, Resources and Environment (TPESRE), Chinese Academy of Sciences. The overall precision of the δ13C analysis was 0.1‰.
2.2 Data analyses
The intensity of interactions between C. versicolor and associated species was assessed by species richness and plant size using the relative interaction index, RIIspecies and RIIsize, respectively (Armas et al., 2004) as , where X is either species richness or plant size under the shrub (Xshrub) or in its paired open area (Xopen). We grouped plant species as grasses, annual forbs, and perennial forbs. RII was calculated at community and functional group levels, respectively.
In order to assess whether our selection of 30 plots in each site is good enough to catch the regional species pool, the rarefaction analysis was run for each site (Supplementary Figure 2). We constructed species accumulation curves to quantify species richness under C. versicolor shrubs (Sshrub), open areas (Sopen), and at the community level (Stotal) in each site (Badano et al., 2006). To estimate Stotal per site, we generated datasets combining data taken under C. versicolor and in open areas. For each rarefaction curve, 500 resamples were randomly drawn without replacement for each sample size (from one sample to the maximum number of samples). In such analyses, the point along the sampling effort axis where the species accumulation curve reaches an asymptote indicates the number of samples needed to successfully sample the full assemblage of species (Gotelli and Colwell, 2001). All rarefaction analyses were carried out using EstimateS 9.1 software (Colwell et al., 2012). To assess the magnitude of the increase in species richness at the community level due to the presence of C. versicolor, we calculated the proportion of increase in herbaceous species richness as (Cavieres et al., 2016). This index provides a qualitative assessment of the magnitude of the effect of shrubs on species richness at the scale of the entire local community.
To assess the difference in species compositions along the gradient, we performed a Principal Coordinate Analysis (PCoA) on a Bray-Curtis dissimilarity matrix. This analysis was performed for the quadrats located away from the shrub only (open areas). The significance of differences between sites was tested by ANOSIM with 9999 permutations. PCoA and screening were performed using the R version 4.1.2 with ape package (https://cran.r-project.org/web/packages/ape/index.html) and vegan package (https://cran.r-project.org/web/packages/vegan/index.html).
There is a scarcity of weather stations in the study area, so that, to obtain comparable long-term climate data, monthly mean temperature and precipitation data were obtained from the WorldClim 2.1 dataset with a spatial resolution of 1 km2 (https://worldclim.org/data/worldclim21.html) (Hijmans et al., 2005) using the geographical coordinates of the sampling sites. Temperature data were corrected by a lapse rate of 6°C km-1 (Kattel et al., 2015). WorldClim data have been widely used for calculating moisture indices to determine habitat severity in different mountain systems where long-term climate records are rarely available. Based on monthly WorldClim data, the annual Thornthwaite water deficit (WD) was calculated for each sampling site as follows: WD = P - PET, where P is the mean annual precipitation and PET is the mean annual potential evapotranspiration (Thornthwaite and Mather, 1955).
We used a one-sample t-test to assess whether RII values differed significantly from 0. Site differences in RII were tested by one-way analysis of variance followed by post hoc Tukey/Duncan test or Dunnett’s T3 test (for homogeneity of variances). The effects of shrub on micro-environment (soil nitrogen and soil water availability) and understory species (species richness and plant size) were assessed by the difference in understory canopy (minuend) and in the open area (subtrahend). General linear regressions of the effects of shrub on understory plants against the effects of shrub on micro-environment were performed at the plot level. All statistical analyses were performed using SPSS 18.0 for Windows, and all significant differences were at P < 0.05. The sampling map was drawn by ArcGIS version 10.8 for Desktop (Environmental Systems Research Institute, Inc.). The elevation generated for the map was downloaded from WorldClim 2.1 with 30s resolution (https://worldclim.org/data/worldclim21.html).
3 Results
3.1 Species composition along the water availability gradient
We recorded 68 associated species in our five sampling sites across the water availability gradient (Supplementary Table 2). Species richness reflected nonlinear patterns in different microhabitats, including under C. versicolor, open areas, and at the community level (Figure 2A). Species richness in open areas was higher than under C. versicolor at all sites, and species richness in the whole community (i.e., understory and open areas combined) was higher than under C. versicolor and open areas (Figure 2A). Principal Coordinate Analysis (PCoA) indicated that the species composition in each of our five sampling sites was significantly different from the others (Figure 2B, R = 0.876, P = 0.001).
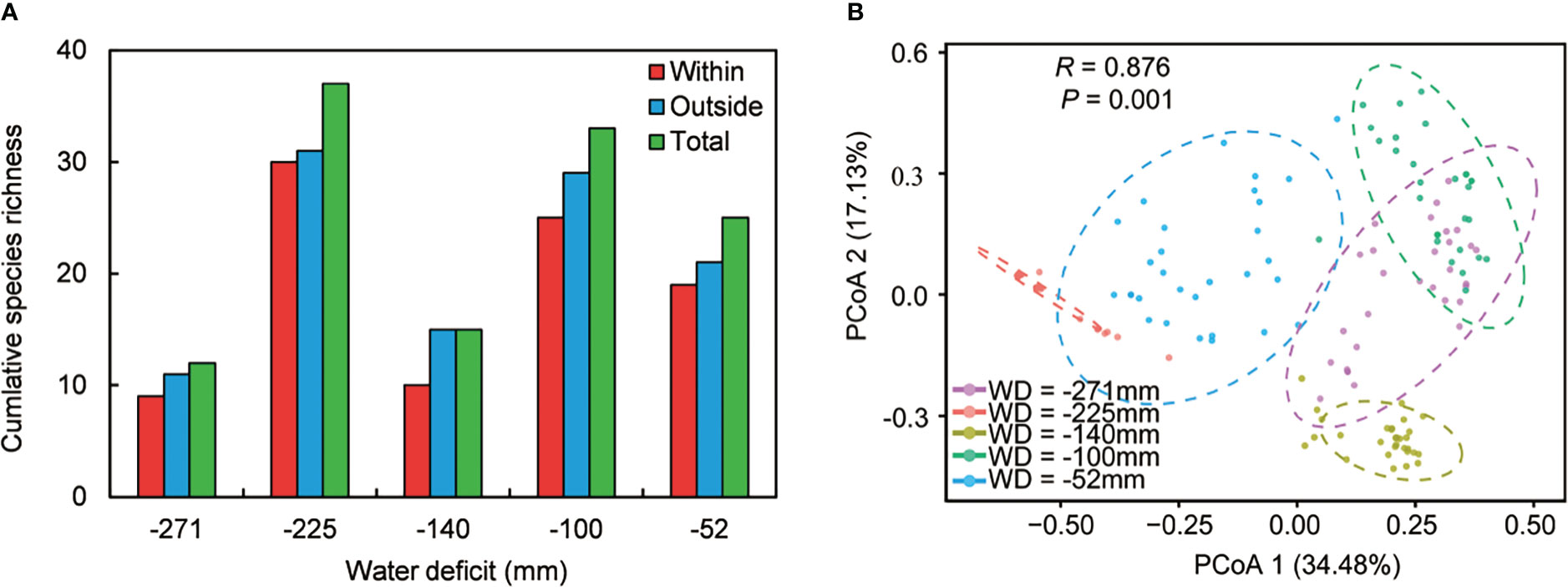
Figure 2 (A) Species richness under Caragana versicolor (within), in open areas (outside) and at the community level (total) along the water deficit gradient in drylands of Tibetan Plateau. (B) Ordination diagram showing Principal Coordinates Analysis (PCoA) performed for species composition in open areas.
We found positive ISR values across sites except at site with WD = -100 mm, indicating that, overall, shrubs enhance species richness at the community level (Table 1). Some specific species were recorded either under C. versicolor or in open areas across the water availability gradient (Table 1), suggesting that some species had strong habitat preferences.
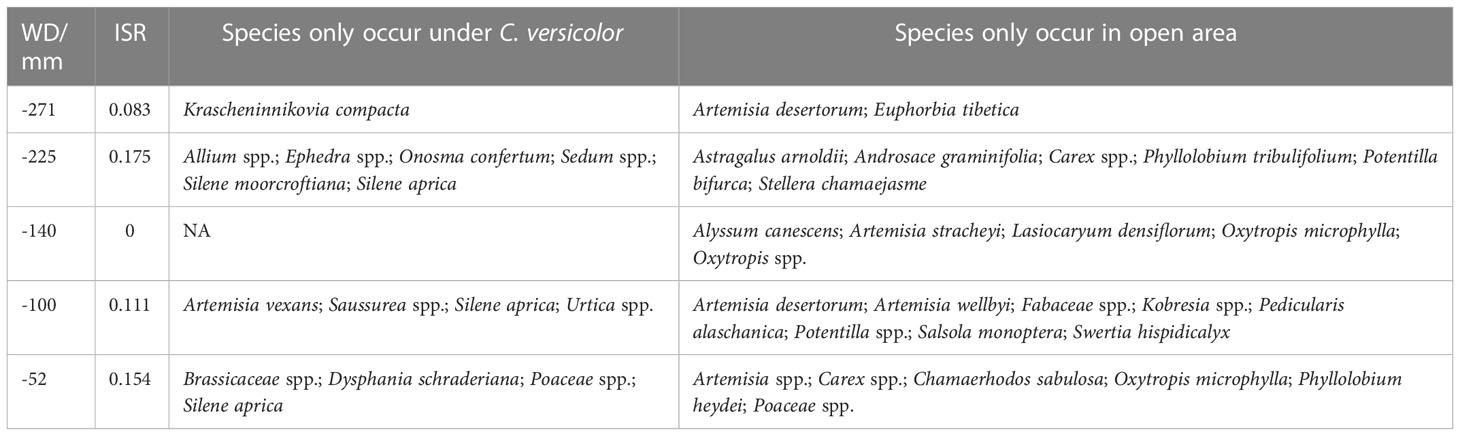
Table 1 Water deficit (WD), the proportion of increase in herbaceous species richness (ISR) and species occurrence under Caragana versicolor or in open areas across 5 sampling sites in drylands of Tibetan Plateau.
3.2 Effects of C. versicolor on species richness and plant size of its understory community
Shrub size, was assessed by the product of the long and minor axes of shrub canopy, as a soft index of shrub age, had no significant effect on understory species richness nor plant size (Supplementary Figure 3). Using all species combined, RIIspecies was mostly negative across the water availability gradient, showing a unimodal trend that suggested a rather strong competition between C. versicolor and its associated understory community (Figure 3A). For the different functional groups, RIIspecies showed different patterns. RIIspecies for grasses was significantly positive except at the driest site (Figure 3B). By contrast, RIIspecies for annual and perennial forbs was generally negative, suggesting competition between shrubs and these two functional groups (Figure 3B). Similarly, RIIspecies including all functional groups also showed a unimodal, negative trend along the water availability gradient (Figure 3B).
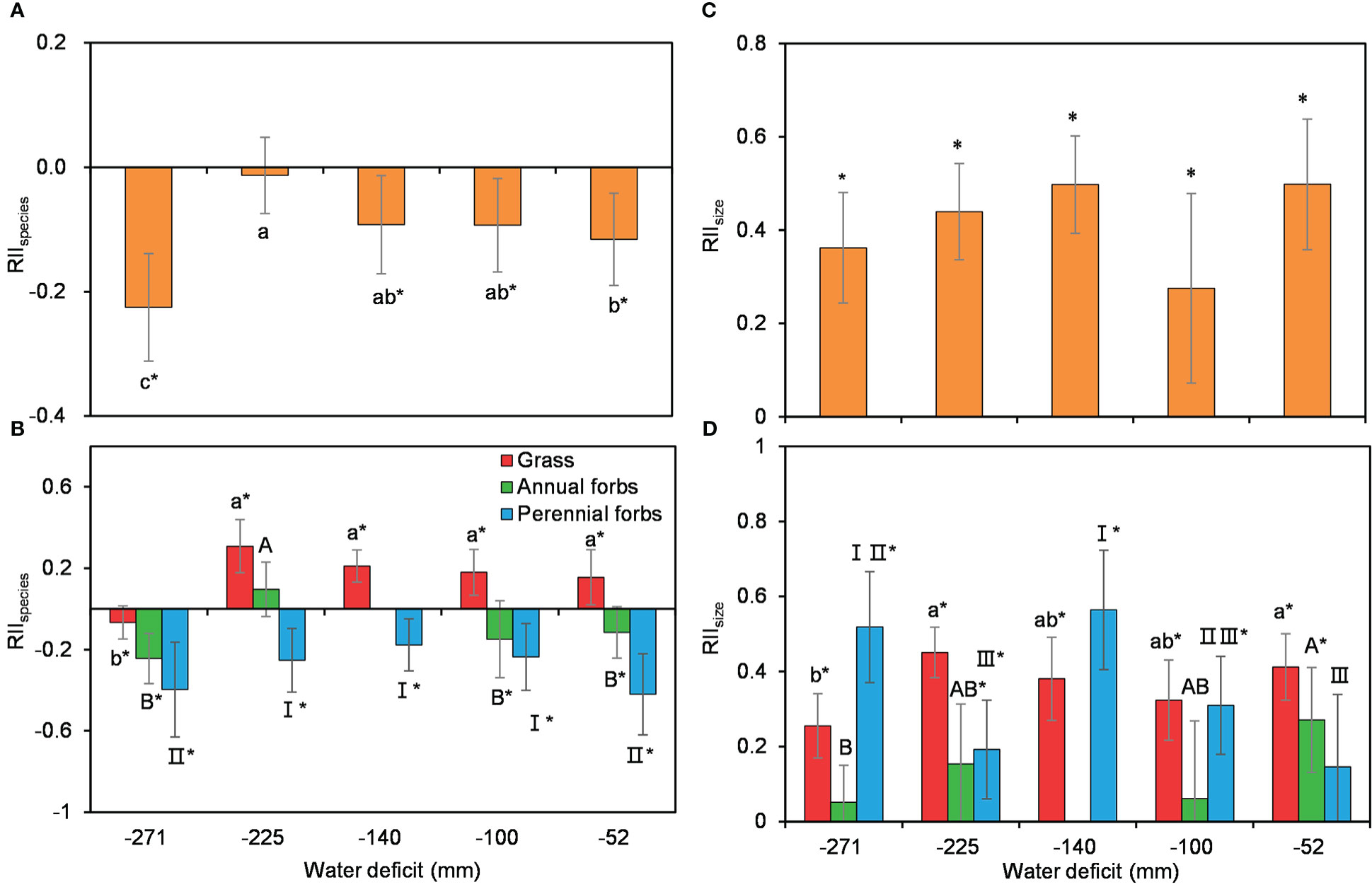
Figure 3 Variations in the interaction effects between Caragana versicolor and understory species assessed by species richness RIIspecies (A, B) and plant size RIIsize (C, D) for total associated species (a and c) and across different functional groups (b and d) along the water deficit gradient in drylands of Tibetan Plateau. Different letters and roman numbers indicate significant differences in RIIspecies or RIIsize between sites; asterisks show that RIIspecies or RIIsize is significantly different from 0.
The effects of C. versicolor on forbs growth, measured as RIIsize, were significantly positive for all species combined and species in different functional groups, indicating a positive effect on plant growth (Figures 3C, D). RIIsize for all species combined showed a similar trend, with no significant changes along the water availability gradient (Figures 3C, D).
3.3 Variations in soil total nitrogen and grass leaf δ13C and their effects on RIIspecies and RIIsize
Soil nitrogen was generally higher under C. versicolor than in open areas except at the driest site (Figure 4A). Soil nitrogen in the understory showed a similar unimodal trend along the water deficit gradient, while there was no significant trend in open areas (Figure 4A). Shrub size had nothing to do with the mediated effect of shrubs on soil total nitrogen (Supplementary Figure 4). There were no differences in grass leaf δ13C across the water gradient, neither within nor outside C. versicolor canopies (Figure 4B).
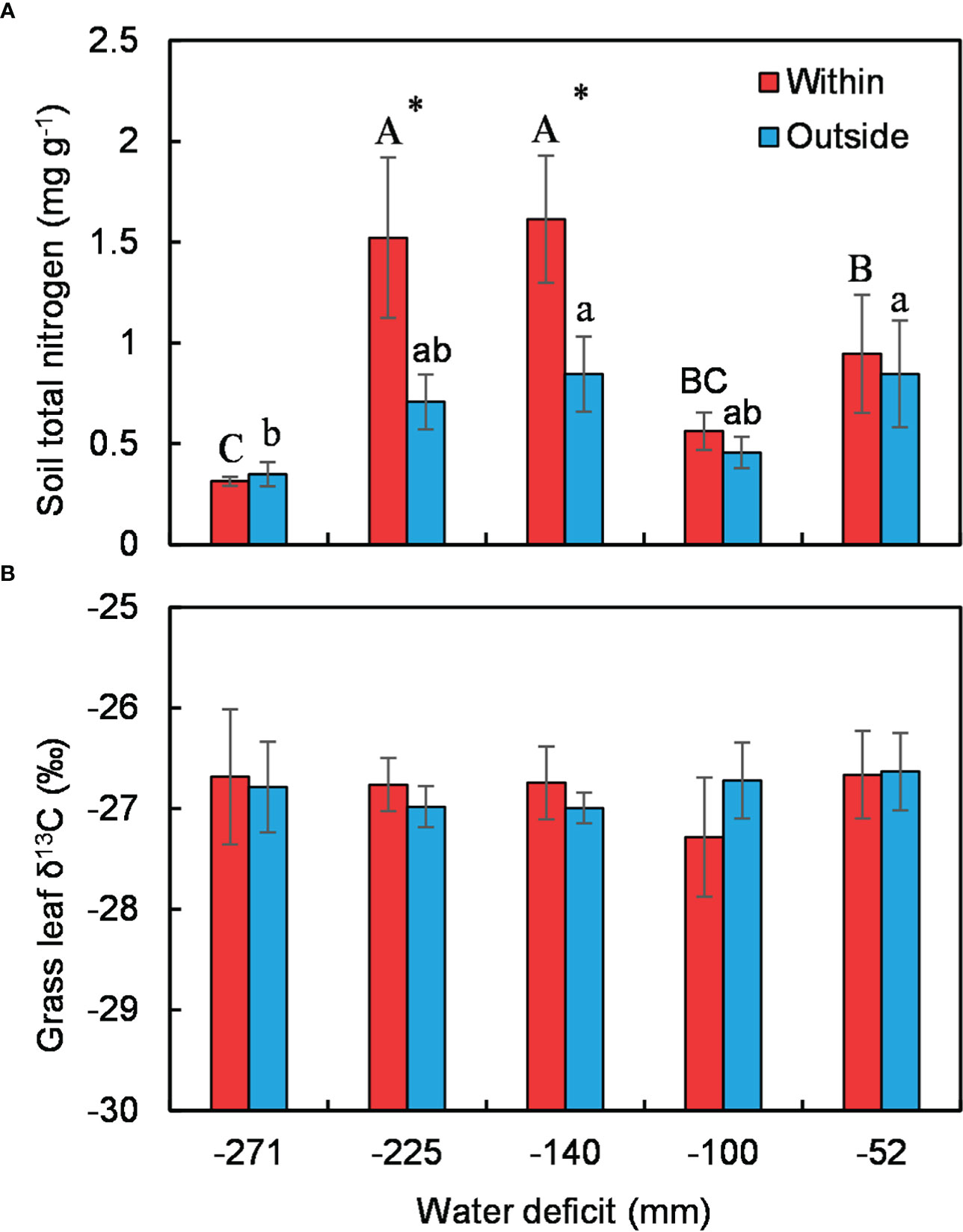
Figure 4 Variations in soil total nitrogen (A) and grass leaf δ13C (B) under Caragana versicolor and in open areas along the water deficit gradient in drylands of Tibetan Plateau. Different letters indicate significant differences in soil total N or δ13C between sites; asterisks show significant differences between the shrub understory and open areas.
The effect of the shrub on species richness compared to open spaces (Δ species richness) was related to its effects on soil N (Δ soil nitrogen; Figure 5A, R2 = 0.892, P = 0.016) and had no significant relationship with Δ grass leaf δ13C (Figure 5B, R2 = 0.114, P = 0.579). The effect of shrubs on understory plant size (Δ plant size) did not show significant relationships with Δ soil nitrogen (Figure 5C, R2 = 0.022, P = 0.814) or Δ grass leaf δ13C (Figure 5D, R2 = 0.141, P = 0.533), suggesting that the effect of C. versicolor on soil nitrogen, rather than water availability, determines its overall effect on understory species richness. Neither the effect of C. versicolor on soil nitrogen nor water availability affected plant size.
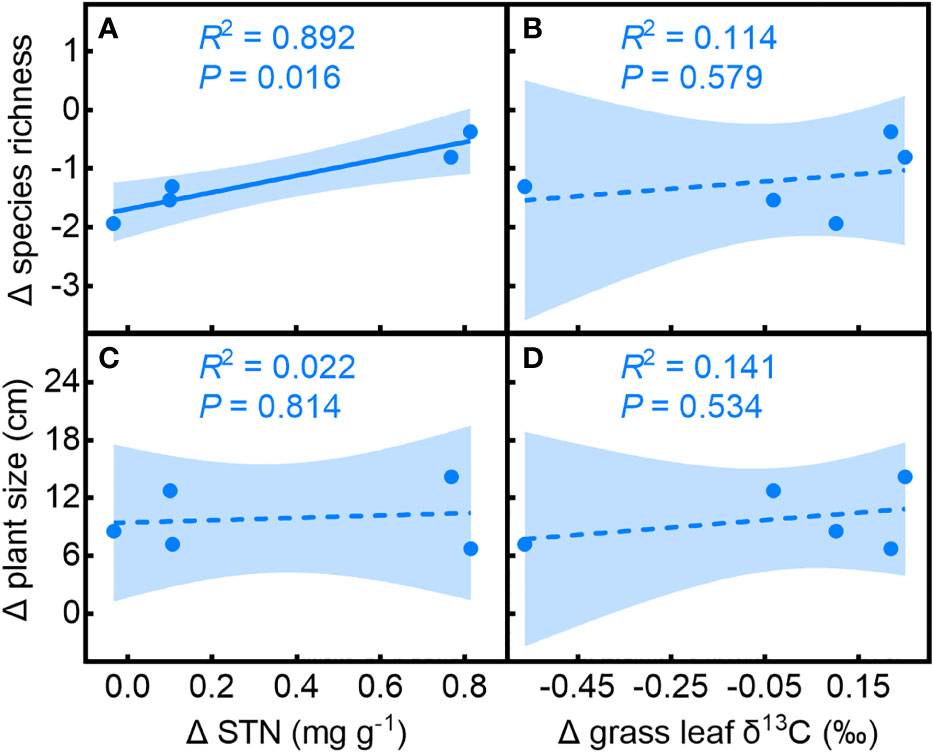
Figure 5 Relationships of the effects of Caragana versicolor on (A, B) species richness (Δ species richness) and (C, D) plant size (Δ plant size) to the effect on soil N (Δ soil N) and grass leaf δ13C (Δ grass leaf δ13C) in drylands of Tibetan Plateau.
4 Discussion
We recorded inconsistent patterns of plant interaction assessed by species richness and plant size with decreasing water availability in drylands of Tibetan Plateau (Figures 3A, C), the unimodal trend of the effect of C. versicolor on species richness departure from the prediction of the SGH (Bertness and Callaway, 1994), but was consistent with reports in other dry systems (Liancourt et al., 2017; O'Brien et al., 2017; Zhang et al., 2018a). We found that C. versicolor significantly increased understory soil nitrogen but had a negligible effect on soil water availability (Figures 4A, B). The effect of C. versicolor on understory species richness was positively related to its effect on soil nitrogen (Figure 5), suggesting that the effect of shrub on soil N, rather than on soil moisture, determines the positive effect of C. versicolor on species richness. As we expected, grass gained more facilitation from C. versicolor than annual and perennial forbs (Figure 3A), likely due to its higher competitive capacity for limited resources compared with other functional groups.
4.1 Effects of arid shrub on associated understory community
Nurse plants have positive effects on the germination, survival, and growth of other species under their canopies in harsh environments such as alpine and arid habitats (Brooker et al., 2008; Michalet and Pugnaire, 2016; Liancourt and Dolezal, 2021). However, for C. versicolor in drylands of Tibetan Plateau, we found opposite trends regarding species richness, which were lower under the shrub canopy. The lower species richness under the canopy than in open spaces between shrubs has been reported also by Liancourt et al. (2017) in the same species in arid Trans-Himalayas but contrasts with reports of positive effects of shrubs on understory species richness in high mountains elsewhere (Ballantyne and Pickering, 2015; Pistón et al., 2016; Iyengar et al., 2017; Parajuli et al., 2021), being most likely an effect of species-specific traits (Callaway, 2007).
We found soil moisture underneath the shrub canopy was generally lower than that in the open area (Supplementary Figure 1), also the water availability indicator, leaf δ13C of grasses, was slightly higher within C. versicolor canopies than outside at most study sites across the water gradient, suggesting that shrubs do not increase soil moisture as do in other facilitator species (Prieto et al., 2011; Tirado et al., 2015; Liu et al., 2021). Water availability is the most important factor limiting plant survival in dry regions, where increasing water availability translates into increased plant survival (Liancourt et al., 2005). In fact, the negative effect of C. versicolor on its understory species is most likely a consequence of competition for soil water. In addition, the negative effect of C. versicolor on the plant survival of associated species would related to its canopy architecture, which branches are dense, forming a tight, hemispheric canopy that provides shade to understory species in this dry environment. Similar results were also reported by Al Hayek et al. (2015) for the spiny cushion shrub, Onobrychis cornuta, which showed strong facilitation in phenotypes with loose canopy and no significant or negative effects in phenotypes with tight canopy. A similar effect was reported for Cytisus galianoi in the Sierra Nevada mountains in Spain (Pistón et al., 2018).
We found that C. versicolor contributed to increasing community-scale species richness at most sites (ISR > 0), since some species were only found under shrub canopies while others preferred open habitats. For instance, Allium spp., Onosma confertum, Silene aprica, and S. moorcroftiana are tall enough to overtop the C. versicolor canopy and therefore are not completely shaded by the shrub, taking advantage of soil nutrients improvement by the shrub. On the contrary, some small Cyperaceae species such as Carex spp. and Kobresia spp., and Fabaceae species with symbiotic nitrogen fixation such as Astragalus arnoldii, Oxytropis microphylla, Phyllolobium heydei and Phyllolobium tribulifolium, only occur in open areas, suggesting they are excluded from the understory by shading. Similarly, some light-demanding species such as Artemisia desertorum, A. stracheyi, and A. wellbyi, which are also unpalatable, were found only in open areas, most likely because they do not stand competition by the shrub (Haase et al., 1996). These cases illustrate how the microenvironmental heterogeneity induced by shrubs affects patterns of species distribution in the community (Armas et al., 2011; Ballantyne and Pickering, 2015; Madrigal-González et al., 2016; Pistón et al., 2016).
In contrast to species richness, we found that RIIsize was generally positive, indicating that C. versicolor increases plant size of associated species. The effect of C. versicolor on plant size may be a consequence of positive shading effect by the shrub. Shrub canopies reduce radiation and wind speed (Padilla and Pugnaire, 2009; Holmgren et al., 2012; Pistón et al., 2016), improving survival of understory species by preventing desiccation. This mechanism should become important in a high-elevation environments such as the Tibetan Plateau, with high radiation and strong winds, where low CO2 levels increase stomatal conductance and pose high risk of desiccation. Although shading may lead to taller plants through etiolation, the benefits of living in the understory may exceed the drawbacks of shade in this otherwise well-lit environment. Additionally, the facilitative effect of C. versicolor on plant size is likely related to the protection against livestock grazing, as plants growing under shrub patches are less accessible to herbivores, therefore experiencing lower grazing pressure (Tirado et al., 2015; Xie et al., 2017; Parajuli et al., 2021).
Considering different functional groups, we found that C. versicolor increased grass species richness but had a negative effect on forbs. The higher competitive ability of grasses compared to other functional groups has been shown in several experiments (Maestre and Cortina, 2004; Noumi et al., 2015; Liu et al., 2021) due to its higher root investment and resources uptake abilities (Gómez-Aparicio, 2009; Gross et al., 2009; Michalet et al., 2015). In addition, exclusion of forbs by C. versicolor could be also due to the generally high N demand of forbs, resulting in strong competition between forbs and shrub for N.
4.2 Shrub facilitation along a water availability gradient
The net balance of benefactor-beneficiary species interaction is species-specific and depends on resource availability (Soliveres et al., 2015; Michalet and Pugnaire, 2016). The SGH predicts a higher frequency of facilitative interactions as resource limitation increases, since facilitator species buffer severe environmental conditions (Bertness and Callaway, 1994). We found that the interaction between C. versicolor and associated species, assessed by species richness and plant size (RIIspecies and RIIsize, respectively), showed different trends along the water deficit gradient. RIIspecies showed a unimodal trend with decreasing water availability, while RIIsize did not change significantly. The effect of C. versicolor on understory species richness (Δ species richness) was positively related to its effect on soil nitrogen (Δ soil nitrogen), but had no significant relationship with Δ leaf δ13C in grasses, indicating that the effect of shrub on soil N, rather than on soil moisture, determines the positive effect of C. versicolor on species richness. Neither the effect of C. versicolor on soil nitrogen nor on water availability affected plant size. This result highlighted the importance of the interaction mechanism to accurately predict the intensity and direction of facilitator-beneficiaries in water-limited systems.
The shift from facilitation to competition under drought conditions may occur when the facilitator species could not take up enough resources for its own subsistence (Maestre and Cortina, 2004; O'Brien et al., 2017; Zhang et al., 2018a). Several studies found that the unimodal trends of plant-plant interactions in water-limited systems were associated with the inability of the facilitator to provide resources (usually, soil water) at the very end of a stress gradient (Michalet et al., 2006; Michalet, 2007; Michalet et al., 2014; Zhang et al., 2016; O'Brien et al., 2017). However, we found that the decrease of C. versicolor on facilitation on species richness at the extremely dry site was related to the decreasing effect on soil nitrogen. Previous studies have proved that soil-mediated effect on soil play an important role on shrub-understories interactions in drylands. For instance, a manipulative experiment found that the roots of Dasiphora fruticosa had stronger effect on understory species composition than the shrub canopy in eastern Tibetan Plateau (Wang et al., 2017). Similar results have also been shown by other studies, which found that understory soil had positive effect on associated species, while shrub canopy had competitive or neutral effects in arid Mediterranean systems (Noumi et al., 2016; Lozano et al., 2020; Chaieb et al., 2021). Soil contribution to facilitation is associated with increased nutrient content in shrub patches or indirectly mediated by soil microorganisms (Rodríguez-Echeverría et al., 2016; Pugnaire et al., 2019). Above mechanism is particular important for leguminous nurse shrub, because their symbiotic N fixation is highly sensitive to water availability. Nitrogenase activities rely on the amount of carbohydrates supplied to nodules, and the supply decreases under water-limited conditions (Marino et al., 2007; Arfin Khan et al., 2014).
The functional strategy (i.e., stress tolerance and competitive ability) of the species involved in the interactions may also affect plant interactions (Michalet et al., 2014). Understory species along the water availability gradient were not always the same and environmental stress should select species that are more drought-tolerant. The turnover of beneficiary species with increasing stress often determines the plant interactions along stress gradients (Choler et al., 2001; Liancourt et al., 2005; Le Bagousse-Pinguet et al., 2012; Liancourt et al., 2017). When both interacting species are stress-tolerant, facilitation can be expected at moderate stress while competition may occur at high-stress levels (Maestre and Cortina, 2004). This may lead to a collapse of facilitation at the extreme end of the stress gradient (Maestre et al, 2009). Our data show that understory species composition in each sampling site significantly differed from other sites. Species are replaced by more drought-tolerant species at the driest site, where they are less likely to obtain a net benefit from the nurse shrub.
Multiple evidences indicate that recent climate change exerts large impacts on the structure and dynamics of plant terrestrial ecosystems, especially in hash systems such as alpine and arid regions (Peñuelas et al., 2013; Piao et al., 2014). In high elevation environments of the Himalaya mountains, climate warming in recent decades along with lower precipitations has led to an increase in water deficit (Yao et al., 2012; Sigdel et al., 2018). Our study found that the facilitation of dominant nurse shrub on soil nitrogen and understory species richness decreased at the driest site in drylands of the Tibetan Plateau, suggesting that the facilitation effect likely decrease if warming-induced water availability crosses a critical minimum threshold. Indeed, the negative effect of climate warming on plant growth and community dynamic has been evidenced, showing that recent warming caused soil moisture deficit and exceeded the thermal optimum of shrub recruitment in high-elevation and high-latitude regions (Francon et al., 2021; Sigdel et al., 2021; Wang et al., 2021; Lu et al., 2022).
5 Conclusion
The dominant leguminous shrub C. versicolor in drylands of the Tibetan Plateau had positive effect on grass species richness, but shown a negative impact on species richness of annual and perennial forbs. Also, C. versicolor exerted a positive effect on plant size of associated species that did not change along the water deficit gradient. The effect of C. versicolor on species richness showed a unimodal trend with decreasing water availability, closely related to the decreasing amelioration of soil nitrogen under extreme dry conditions. Our results suggest that the tendency of drought associated with warming in drylands on the Tibetan Plateau will likely hinder the facilitation effect of nurse leguminous shrub on understories if moisture availability crosses a critical minimum threshold.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
LZ and EL designed the study; LZ, GC, and LY collected and measured the samples. All authors contributed to data analyses, interpretation and writing. All authors contributed to the article and approved the submitted version.
Funding
This research is funded by the Second Tibetan Plateau Scientific Expedition and Research (STEP) program (2019QZKK0301-1) and the Key Technology research and development project in Xizang Autonomous Regions (XZ202101ZY005G). GC was financially supported by the Chinese Scholarship Council (joint PhD student, 201904910331) to conduct ecological research at Estación Experimental de Zonas Áridas, Consejo Superior de Investigaciones Científicas.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2023.1137365/full#supplementary-material
References
Ale, R., Zhang, L., Li, X., Raskoti, B. B., Pugnaire, F. I., Luo, T. X. (2018). Leaf δ13C as an indicator of water availability along elevation gradients in the dry Himalayas. Ecol. Indic. 94, 266–273. doi: 10.1016/j.ecolind.2018.07.002
Al Hayek, P., Maalouf, J. P., Baumel, A., Dagher-Kharrat, M. B., Medail, F., Touzard, B., et al. (2015). Differential effects of contrasting phenotypes of a foundation legume shrub drive plant-plant interactions in a Mediterranean mountain. J. Veg. Sci. 26, 373–384. doi: 10.1111/jvs.12246
Anthelme, F., Cavieres, L. A., Dangles, O. (2014). Facilitation among plants in alpine environments in the face of climate change. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00387
Arfin Khan, M. A. S., Grant, K., Beierkuhnlein, C., Kreyling, J., Jentsch, A. (2014). Climatic extremes lead to species-specific legume facilitation in an experimental temperate grassland. Plant Soil. 379, 161–175. doi: 10.1007/s11104-014-2050-8
Armas, C., Ordiales, R., Pugnaire, F. I. (2004). Measuring plant interactions: a new comparative index. Ecology 85, 2682–2686. doi: 10.1890/03-0650
Armas, C., Rodríguez-Echeverría, S., Pugnaire, F. I. (2011). A field test of the stress-gradient hypothesis along an aridity gradient. J. Veg. Sci. 22, 818–827. doi: 10.1111/j.1654-1103.2011.01301.x
Badano, E. I., Jones, C. G., Cavieres, L. A., Wright, J. P. (2006). Assessing impacts of ecosystem engineers on community organization: A general approach illustrated by effects of a high-Andean cushion plant. Oikos 115, 369–385. doi: 10.1111/j.2006.0030-1299.15132.x
Ballantyne, M., Pickering, C. M. (2015). Shrub facilitation is an important driver of alpine plant community diversity and functional composition. Biodivers. Conserv. 24, 1859–1875. doi: 10.1007/s10531-015-0910-z
Bertness, M. D., Callaway, R. (1994). Positive interactions in communities. Trends Ecol. Evol. 9, 191–193. doi: 10.1016/0169-5347(94)90088-4
Brooker, R. W., Maestre, F. T., Callaway, R. M., Lortie, C. L., Cavieres, L. A., Kunstler, G., et al. (2008). Facilitation in plant communities: The past, the present, and the future. J. Ecol. 96, 18–34. doi: 10.1111/j.1365-2745.2007.01295.x
Butterfield, B. J., Bradford, J. B., Armas, C., Prieto, I., Pugnaire, F. I. (2016). Does the stress-gradient hypothesis hold water? disentangling spatial and temporal variation in plant effects on soil moisture in dryland systems. Funct. Ecol. 30, 10–19. doi: 10.1111/1365-2435.12592
Cai, Y., Yan, Y., Xu, D., Xu, X., Wang, C., Wang, X. (2020). The fertile island effect collapses under extreme overgrazing: Evidence from a shrub-encroached grassland. Plant Soil 448, 201–212. doi: 1007/s11104-020-04426-2
Callaway, R. M. (2007). Positive interactions and interdependence in plant communities (Netherlands: Springer Press).
Callaway, R. M., Walker, L. R. (1997). Competition and facilitation: A synthetic approach to interactions in plant communities. Ecology 78, 1958–1965. doi: 10.1890/0012-9658(1997)078[1958:Cafasa]2.0.Co;2
Cavieres, L. A., Brooker, R. W., Butterfield, B. J., Cook, B. J., Kikvidze, Z., Lortie, C. J., et al. (2014). Facilitative plant interactions and climate simultaneously drive alpine plant diversity. Ecol. Lett. 17, 193–202. doi: 10.1111/ele.12217
Cavieres, L. A., Hernández-Fuentes, C., Sierra-Almeida, A., Kikvidze, Z. (2016). Facilitation among plants as an insurance policy for diversity in alpine communities. Funct. Ecol. 30, 52–59. doi: 10.1111/1365-2435.12545
Chaieb, G., Wang, X., Abdelly, C., Michalet, R. (2021). Shift from short-term competition to facilitation with drought stress is due to a decrease in long-term facilitation. Oikos 130, 29–40. doi: 10.1111/oik.07528
Choler, P., Michalet, R., Callaway, R. M. (2001). Facilitation and competition on gradients in alpine plant communities. Ecology 82, 3295–3308. doi: 10.1890/0012-9658(2001)082[3295:Facogi]2.0.Co;2
Colwell, R. K., Chao, A., Gotelli, N. J., Lin, S. Y., Mao, C. X., Chazdon, R. L., et al. (2012). Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 5, 3–21. doi: 10.1093/jpe/rtr044
Darrouzet-Nardi, A., D’antonio, C. M., Dawson, T. E. (2006). Depth of water acquisition by invading shrubs and resident herbs in a Sierra Nevada meadow. Plant Soil 285, 31–43. doi: 10.1007/s11104-005-4453-z
Dawson, T. E., Mambelli, S., Plamboeck, A. H., Templer, P. H., Tu, K. P. (2002). Stable isotopes in plant ecology. Annu. Rev. Ecol. Evol. Syst. 33, 507–559. doi: 10.1146/annurev.ecolsys.33.020602.095451
Farquhar, G. D., Ehleringer, J. R., Hubick, K. T. (1989). Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Biol. 40, 503–537. doi: 10.1146/annurev.pp.40.060189.002443
Francon, L., Corona, C., Till-Bottraud, I., Choler, P., Roussel, E., Carlson, B. Z., et al. (2021). Shrub growth in the Alps diverges from air temperature since the 1990s. Environ. Res. Lett. 16, 074026. doi: 10.1088/1748-9326/ac0b67
Gómez-Aparicio, L. (2009). The role of plant interactions in the restoration of degraded ecosystems: A meta-analysis across life-forms and ecosystems. J. Ecol. 97, 1202–1214. doi: 10.1111/j.1365-2745.2009.01573.x
Gotelli, N. J., Colwell, R.K. (2001). Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 4, 379–391. doi: 10.1046/j.1461-0248.2001.00230.x
Gross, N., Kunstler, G., Liancourt, P., De Bello, F., Suding, K. N., Lavorel, S. (2009). Linking individual response to biotic interactions with community structure: A trait-based framework. Funct. Ecol. 23, 1167–1178. doi: 10.1111/j.1365-2435.2009.01591.x
Haase, P., Pugnaire, F. I., Clark, S., Incoll, L. (1996). Spatial patterns in a two-tiered semi-arid shrubland in southeastern Spain. J. Veg. Sci. 7, 527–534. doi: 10.2307/3236301
Hamerlynck, E. P., Scott, R. L., Susan Moran, M., Schwander, A. M., Connor, E., Huxman, T. E. (2011). Inter- and under-canopy soil water, leaf-level and whole-plant gas exchange dynamics of a semi-arid perennial C4 grass. Oecologia 165, 17–29. doi: 10.1007/s00442-010-1757-3
He, Q., Bertness, M. D., Altieri, A. H. (2013). Global shifts towards positive species interactions with increasing environmental stress. Ecol. Lett. 16, 695–706. doi: 10.1111/ele.12080
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G., Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25, 1965–1978. doi: 10.1002/joc.1276
Holmgren, M., Gomez-Aparicio, L., Luis Quero, J., Valladares, F. (2012). Non-linear effects of drought under shade: Reconciling physiological and ecological models in plant communities. Oecologia 169, 293–305. doi: 10.1007/s00442-011-2196-5
Holmgren, M., Scheffer, M. (2010). Strong facilitation in mild environments: The stress gradient hypothesis revisited. J. Ecol. 98, 1269–1275. doi: 10.1111/j.1365-2745.2010.01709.x
Iyengar, S. B., Bagchi, S., Barua, D., Mishra, C., Sankaran, M. (2017). A dominant dwarf shrub increases diversity of herbaceous plant communities in a trans-Himalayan rangeland. Plant Ecol. 218, 843–854. doi: 10.1007/s11258-017-0734-x
Kattel, D. B., Yao, T. D., Yang, W., Gao, Y., Tian, L. D. (2015). Comparison of temperature lapse rates from the northern to the southern slopes of the Himalayas. Int. J. Climatol. 35, 4431–4443. doi: 10.1002/joc.4297
Kumar, A., Adhikari, B. S., Rawat, G. S. (2016). Caragana versicolor benth. (Fabaceae), a keystone species of high conservation concern in the Hindu kush Himalayan region. Curr. Sci. 111, 985–987.
Le Bagousse-Pinguet, Y., Liancourt, P., Gross, N., Straile, D. (2012). Indirect facilitation promotes macrophyte survival and growth in freshwater ecosystems threatened by eutrophication. J. Ecol. 100, 530–538. doi: 10.1111/j.1365-2745.2011.01931.x
Liancourt, P., Callaway, R. M., Michalet, R. (2005). Stress tolerance and competitive-response ability determine the outcome of biotic interactions. Ecology 86, 1611–1618. doi: 10.1890/04-1398
Liancourt, P., Dolezal, J. (2021). Community-scale effects and strain: Facilitation beyond conspicuous patterns. J. Ecol. 109, 19–25. doi: 10.1111/1365-2745.13458
Liancourt, P., Le Bagousse-Pinguet, Y., Rixen, C., Dolezal, J. (2017). SGH: stress or strain gradient hypothesis? insights from an elevation gradient on the roof of the world. Ann. Bot. 120, 29–38. doi: 10.1093/aob/mcx037
Liu, L., Bai, Y. X., She, W. W., Qiao, Y. G., Qin, S. G., Zhang, Y. Q. (2021). A nurse shrub species helps associated herbaceous plants by preventing shade-induced evaporation in a desert ecosystem. Land Degrad. Dev. 32, 1796–1808. doi: 10.1002/ldr.3831
Lozano, Y. M., Hortal, S., Armas, C., Pugnaire, F. I. (2020). Complementarity in nurse plant systems: Soil drives community composition while microclimate enhances productivity and diversity. Plant Soil 450, 385–396. doi: 10.1007/s11104-020-04503-6
Lu, X., Liang, E., Babst, F., Camarero, J. J., Büntgen, U. (2022). Warming-induced tipping points of Arctic and alpine shrub recruitment. Proc. Natl. Acad. Sci. U.S.A 119, e2118120119. doi: 10.1073/pnas.2118120119
Madrigal-González, J., Kelt, D. A., Meserve, P. L., Squeo, F. A., Gutiérrez, J. R. (2016). Shrub-ephemeral plants interactions in semiarid north-central Chile: Is the nurse plant syndrome manifested at the community level? J. Arid Environ. 126, 47–53. doi: 10.1016/j.jaridenv.2015.08.001
Maestre, F. T., Bautista, S., Cortina, J., Bellot, J. (2001). Potential for using facilitation by grasses to establish shrubs on a semiarid degraded steppe. Ecol. Appl. 11, 1641–1655. doi: 10.1890/1051-0761(2001)011[1641:Pfufbg]2.0.Co;2
Maestre, F. T., Bowker, M. A., Puche, M. D., Hinojosa M, B. H., Martínez, I., García-Palacios, P., et al. (2009). Shrub encroachment can reverse desertification in semi-arid Mediterranean grasslands. Ecol. Lett. 12, 930–941. doi: 10.1111/j.1461-0248.2009.01352.x
Maestre, F. T., Cortina, J. (2004). Do positive interactions increase with abiotic stress? a test from a semi-arid steppe. Proc. R. Soc B: Biol. Sci. 271 (Suppl 5), S331–S333. doi: 10.1098/rsbl.2004.0181
Marino, D., Frendo, P., Ladrera, R., Zabalza, A., Puppo, A., Arrese-Igor, C. (2007). Nitrogen fixation control under drought stress. Localized or systemic? Plant physiology 143, 1968–1974. doi: 10.1104/pp.107.097139
Michalet, R. (2007). Highlighting the multiple drivers of change in interactions along stress gradients. New Phytol. 173, 3–6. doi: 10.1111/j.1469-8137.2006.01949.x
Michalet, R., Brooker, R. W., Cavieres, L. A., Kikvidze, Z., Lortie, C. J., Pugnaire, F. I., et al. (2006). Do biotic interactions shape both sides of the humped-back model of species richness in plant communities? Ecol. Lett. 9, 767–773. doi: 10.1111/j.1461-0248.2006.00935.x
Michalet, R., Brooker, R. W., Lortie, C. J., Maalouf, J. P., Pugnaire, F. I. (2015). Disentangling direct and indirect effects of a legume shrub on its understorey community. Oikos 124, 1251–1262. doi: 10.1111/oik.01819
Michalet, R., Le Bagousse-Pinguet, Y., Maalouf, J. P., Lortie, C. J. (2014). Two alternatives to the stress-gradient hypothesis at the edge of life: the collapse of facilitation and the switch from facilitation to competition. J. Veg. Sci. 25, 609–613. doi: 10.1111/jvs.12123
Michalet, R., Pugnaire, F. I. (2016). Facilitation in communities: Underlying mechanisms, community and ecosystem implications. Funct. Ecol. 30, 3–9. doi: 10.1111/1365-2435.12602
Noumi, Z., Chaieb, M., Le Bagousse-Pinguet, Y., Michalet, R. (2016). The relative contribution of short-term versus long-term effects in shrub-understory species interactions under arid conditions. Oecologia 180, 529–542. doi: 10.1007/s00442-015-3482-4
Noumi, Z., Chaieb, M., Michalet, R., Touzard, B. (2015). Limitations to the use of facilitation as a restoration tool in arid grazed savanna: A case study. Appl. Veg. Sci. 18, 391–401. doi: 10.1111/avsc.12158
O'Brien, M. J., Pugnaire, F. I., Armas, C., Rodríguez-Echeverría, S., Schöb, C. (2017). The shift from plant-plant facilitation to competition under severe water deficit is spatially explicit. Ecol. Evol. 7, 2441–2448. doi: 10.1002/ece3.2875
O'Brien, M. J., De Menezes, L. F. T., Bråthen, K. A., Losapio, G., Pugnaire, F. I. (2019). Facilitation mediates species presence beyond their environmental optimum. Perspect. Plant Ecol. Evol. Syst. 38, 24–30. doi: 10.1016/j.ppees.2019.03.004
Padilla, F. M., Pugnaire, F. I. (2009). Species identity and water availability determine establishment success under the canopy of retama sphaerocarpa shrubs in a dry environment. Restor. Ecol. 17, 900–907. doi: 10.1111/j.1526-100X.2008.00460.x
Parajuli, R., O'brien, M. J., Timilsina, B., Pugnaire, F. I., Schöeb, C., Ghimire, S. K. (2021). Facilitation by a dwarf shrub enhances plant diversity of human-valued species at high elevations in the Himalayas of Nepal. Basic Appl. Ecol. 54, 23–36. doi: 10.1016/j.baae.2021.04.004
Peñuelas, J., Sardans, J., Estiarte, M., Ogaya, R., Carnicer, J., Coll, M., et al. (2013). Evidence of current impact of climate change on life: A walk from genes to the biosphere. Glob. Change Biol. 19, 2303–2338. doi: 10.1111/gcb.12143
Piao, S., Nan, H., Huntingford, C., Ciais, P., Friedlingstein, P., Sitch, S., et al. (2014). Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nat. Commun. 5, 1–7. doi: 10.1038/ncomms6018
Pistón, N., Michalet, R., Schöb, C., Macek, P., Armas, C., Pugnaire, F. I. (2018). The balance of canopy and soil effects determines intraspecific differences in foundation species’ effects on associated plants. Funct. Ecol. 32, 2253–2263. doi: 10.1111/1365-2435.13139
Pistón, N., Schöb, C., Armas, C., Prieto, I., Pugnaire, F. I. (2016). Contribution of co-occurring shrub species to community richness and phylogenetic diversity along an environmental gradient. Perspect. Plant Ecol. Evol. Syst. 19, 30–39. doi: 10.1016/j.ppees.2016.02.002
Prieto, I., Padilla, F. M., Armas, C., Pugnaire, F. I. (2011). The role of hydraulic lift on seedling establishment under a nurse plant species in a semi-arid environment. Perspect. Plant Ecol. Evol. Syst. 13, 181–187. doi: 10.1016/j.ppees.2011.05.002
Pugnaire, F. I., Armas, C., Maestre, F. T. (2011). Positive plant interactions in the Iberian southeast: Mechanisms, environmental gradients, and ecosystem function. J. Arid Environ. 75, 1310–1320. doi: 10.1016/j.jaridenv.2011.01.016
Pugnaire, F. I., Haase, P., Puigdefábregas, J., Cueto, M., Clark, S. C., Incoll, L. D. (1996). Facilitation and succession under the canopy of a leguminous shrub, Retama sphaerocarpa, in a semi-arid environment in south-east Spain. Oikos 76, 455–464. doi: 10.2307/3546339
Pugnaire, F. I., Losapio, G., Schöb, C. (2021). Species interactions involving cushion plants in high-elevation environments under a changing climate. Ecosistemas 30, 2186. doi: 10.7818/ECOS.2186
Pugnaire, F. I., Morillo, J. A., Peñuelas, J., Reich, P. B., Bardgett, R. D., Gaxiola, A., et al. (2019). Climate change effects on plant-soil feedbacks and consequences for biodiversity and functioning of terrestrial ecosystems. Sci. Adv. 5, eaaz1834. doi: 10.1126/sciadv.aaz1834
Richards, J. H., Caldwell, M. M. (1987). Hydraulic lift: Substantial nocturnal water transport between soil layers by Artemisia tridentata roots. Oecologia 73, 486–489. doi: 10.1007/bf00379405
Rodríguez-Echeverría, S., Lozano, Y. M., Bardgett, R. D. (2016). Influence of soil microbiota in nurse plant systems. Funct. Ecol. 30, 30–40. doi: 10.1111/1365-2435.12594
Saixiyala, Y., Yang, D., Zhang, S. D., Liu, G. F., Yang, X. J., et al. (2017). Facilitation by a spiny shrub on a rhizomatous clonal herbaceous in thicketization-grassland in northern China: Increased soil resources or shelter from herbivores. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.00809
Sigdel, S. R., Pandey, J., Liang, E., Muhammad, S., Babst, F., Leavitt, S. W., et al. (2021). No benefits from warming even for subnival vegetation in the central Himalayas. Sci. Bull. 66, 1825–1829. doi: 10.1016/j.scib.2021.06.005
Sigdel, S. R., Wang, Y., Camarero, J. J., Zhu, H., Liang, E., Peñuelas, J. (2018). Moisture-mediated responsiveness of treeline shifts to global warming in the Himalayas. Glob. Change Biol. 24, 5549–5559. doi: 10.1111/gcb.14428
Soliveres, S., Smit, C., Maestre, F. T. (2015). Moving forward on facilitation research: Response to changing environments and effects on the diversity, functioning and evolution of plant communities. Biol. Rev. 90, 297–313. doi: 10.1111/brv.12110
Thornthwaite, C., Mather, J. (1955). The water balance. centerton, Drexel institute of technology-laboratory of climatology. Publications climatology 8, 104.
Tielbörger, K., Kadmon, R. (2000). Temporal environmental variation tips the balance between facilitation and interference in desert plants. Ecology 81, 1544–1553. doi: 10.2307/177305
Tirado, R., Bråthen, K. A., Pugnaire, F. I. (2015). Mutual positive effects between shrubs in an arid ecosystem. Sci. Rep. 5, 14710. doi: 10.1038/srep14710
Wang, Y., Liang, E., Lu, X., Camarero, J. J., Babst, F., Shen, M., et al. (2021). Warming-induced shrubline advance stalled by moisture limitation on the Tibetan plateau. Ecography 44, 1631–1641. doi: 10.1111/ecog.05845
Wang, X. T., Michalet, R., Chen, S. Y., Zhao, L., An, L. Z., Du, G. Z., et al. (2017). Contrasting understorey species responses to the canopy and root effects of a dominant shrub drive community composition. J. Veg. Sci. 28, 1118–1127. doi: 10.1111/jvs.12565
Wezel, A., Rajot, J. L., Herbrig, C. (2000). Influence of shrubs on soil characteristics and their function in sahelian agro-ecosystems in semi-arid Niger. J. Arid Environ. 44, 383–398. doi: 10.1006/jare.1999.0609
Xie, L. N., Guo, H. Y., Liu, Z., Gabler, C. A., Chen, W. Z., Gu, S., et al. (2017). Shrubs facilitate recruitment of Caragana stenophylla pojark: Microhabitat amelioration and protection against herbivory. Ann. For. Sci. 74, 1–12. doi: 10.1007/s13595-017-0668-4
Yao, T., Thompson, L., Yang, W., Yu, W., Gao, Y., Guo, X., et al. (2012). Different glacier status with atmospheric circulations in Tibetan plateau and surroundings. Nat. Clim. Change 2, 663–667. doi: 10.1038/nclimate1580
Zhang, H. Y., Lü, X. T., Knapp, A. K., Hartmann, H., Bai, E., Wang, X. B., et al. (2018b). Facilitation by leguminous shrubs increases along a precipitation gradient. Funct. Ecol. 32, 203–213. doi: 10.1111/1365-2435.12941
Zhang, P., Yang, J., Zhao, L., Bao, S., Song, B. (2011). Effect of Caragana tibetica nebkhas on sand entrapment and fertile islands in steppe-desert ecotones on the inner Mongolia plateau, China. Plant Soil 347, 79–90. doi: 10.1007/s11104-011-0813-z
Zhang, G., Zhao, L., Yang, Q., Zhao, W., Wang, X. (2016). Effect of desert shrubs on fine-scale spatial patterns of understory vegetation in a dry-land. Plant Ecol. 217, 1141–1155. doi: 10.1007/s11258-016-0639-0
Keywords: stress gradient hypothesis, water availability, soil nitrogen, arid shrub, facilitation
Citation: Cui G, Pugnaire FI, Yang L, Zhao W, Ale R, Shen W, Luo T, Liang E and Zhang L (2023) Shrub-mediated effects on soil nitrogen determines shrub-herbaceous interactions in drylands of the Tibetan Plateau. Front. Plant Sci. 14:1137365. doi: 10.3389/fpls.2023.1137365
Received: 04 January 2023; Accepted: 30 January 2023;
Published: 10 February 2023.
Edited by:
Shiliang Liu, Beijing Normal University, ChinaReviewed by:
Zhishan Zhang, Northwest Institute of Eco-Environment and Resources (CAS), ChinaSa Xiao, Lanzhou University, China
Biying Liu, School of Life Sciences, Sun Yat-sen University, China
Chongchong Ye, Beijing Normal University, China
Copyright © 2023 Cui, Pugnaire, Yang, Zhao, Ale, Shen, Luo, Liang and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lin Zhang, zhanglin@itpcas.ac.cn