 Eva Gfrerer1†
Eva Gfrerer1† Danae Laina1†
Danae Laina1† Marc Gibernau2†
Marc Gibernau2† Roman Fuchs1†
Roman Fuchs1† Martin Happ3†Till Tolasch4Wolfgang Trutschnig5†Anja C. Hörger1†Hans Peter Comes1†
Martin Happ3†Till Tolasch4Wolfgang Trutschnig5†Anja C. Hörger1†Hans Peter Comes1† Stefan Dötterl1*†
Stefan Dötterl1*†- 1Department of Biosciences, Paris Lodron University of Salzburg, Salzburg, Austria
- 2Laboratory of Sciences for the Environment, CNRS – University of Corsica, Ajaccio, France
- 3Lab for Intelligent Data Analytics Salzburg, Paris Lodron University of Salzburg, Salzburg, Austria
- 4FG Tierökologie, University of Hohenheim, Stuttgart, Germany
- 5Department of Mathematics, Paris Lodron University of Salzburg, Salzburg, Austria
Floral scent is a key mediator in plant-pollinator interactions. However, little is known to what extent intraspecific scent variation is shaped by phenotypic selection, with no information yet in deceptive plants. In this study, we collected inflorescence scent and fruit set of the deceptive moth fly-pollinated Arum maculatum L. (Araceae) from six populations north vs. five populations south of the Alps, accumulating to 233 samples in total, and tested for differences in scent, fruit set, and phenotypic selection on scent across this geographic barrier. We recorded 289 scent compounds, the highest number so far reported in a single plant species. Most of the compounds occurred both north and south of the Alps; however, plants of the different regions emitted different absolute and relative amounts of scent. Fruit set was higher north than south of the Alps, and some, but not all differences in scent could be explained by differential phenotypic selection in northern vs. southern populations. This study is the first to provide evidence that floral scents of a deceptive plant are under phenotypic selection and that phenotypic selection is involved in shaping geographic patterns of floral scent in such plants. The hyperdiverse scent of A. maculatum might result from the imitation of various brood substrates of its pollinators.
Introduction
Approximately 88% of angiosperms are cross-pollinated by animals (Ollerton et al., 2011) that are attracted to flowers by multifaceted cues (Chittka and Thomson, 2001). Together with visual cues, the main attractant for pollinators is the floral scent (Knudsen et al., 2006; Raguso, 2008). Therefore, the scent has strong effects on pollinator visitation and frequency and, hence, the reproductive success of the plant (Raguso, 2008; Delle-Vedove et al., 2017). With more than 2,000 floral volatile organic compounds (VOCs) described (Knudsen et al., 2006; El-Sayed, 2019), and an average of 20–60 VOCs per species (Knudsen and Gershenzon, 2020), floral scent blends can tremendously vary among species in terms of composition and quantity. Consequently, they facilitate discrimination by pollinators among host plant species and contribute to reproductive isolation of closely related species (Stökl et al., 2009; Friberg et al., 2014).
In addition to interspecific variation, floral scent is also known to vary intraspecifically, both within and among populations (Delle-Vedove et al., 2017). Such intraspecific variability might result directly from abiotic (e.g., temperature; Farré-Armengol et al., 2014) and/or biotic (e.g., herbivores; Kessler and Halitschke, 2009) factors. Given that scent is heritable (e.g., Zu et al., 2016), intraspecific differences can also result from varying evolutionary forces, such as natural selection and genetic drift (Herrera et al., 2006; Majetic et al., 2009). Although not explicitly demonstrated, genetic drift was suggested to be responsible for strong inter-population differences in floral scents (Delle-Vedove et al., 2017) or to counteract pollinator-mediated selection (in two Yucca species; Svensson et al., 2006). In contrast, natural selection on floral scent emission, both on total scent amount and individual scent components, has been shown by analyses of phenotypic selection, correlating scent phenotypes and fitness measures (e.g., Parachnowitsch et al., 2012; Gross et al., 2016; Chapurlat et al., 2019).
Phenotypic selection on floral scent can vary intraspecifically, potentially leading to variable adaptive responses to spatially variable pollinator assemblages (Gross et al., 2016). Until now, studies examining phenotypic selection on floral scents have been conducted in rewarding but not in deceptive species, although the latter also often rely on luring and deceiving their pollinators with scents (Jürgens et al., 2013; Schiestl and Johnson, 2013). Compared with their rewarding relatives, non-rewarding species often display higher variation in scent and other traits attractive to pollinators (e.g., Ackerman et al., 2011; Dormont et al., 2014). Furthermore, non-rewarding species are frequently more pollen-limited, e.g., Tremblay et al. (2005). In consequence, they might experience stronger selection on floral scent than rewarding species, as shown for floral traits other than scent (Sletvold and Ågren, 2014).
An ideal target for studying phenotypic selection on scent is the moth fly-pollinated and brood-site deceptive Arum maculatum L. (Araceae). This strongly scented (e.g., Kite, 1995; Chartier et al., 2013) plant species attracts its pollinators by olfactory deception (e.g., Kite et al., 1998), shows high variation in fruit and seed sets within and among populations (e.g., Ollerton and Diaz, 1999), and has a geographically variable pollinator spectrum (Espíndola et al., 2011). This perennial herb is widespread in Europe, and the main pollinators are two moth flies, namely Psychoda phalaenoides L. and P. grisescens TONN. (Psychodidae). In Central and much of Western Europe, high abundances of female P. phalaenoides were found (Espíndola et al.,2011). In other regions, such as Mediterranean Europe and Western France, A. maculatum was generally visited by a higher diversity of Diptera (psychodids and non-psychodids) but in much lower abundances, often dominantly by both sexes of P. grisescens and not by P. phalaenoides (Espíndola et al., 2011). This geographic pollinator variation is particularly pronounced north vs. south of the Alps (Espíndola et al., 2011; Laina et al., unpublished data) and matches a genetic subdivision (amplified fragment length polymorphisms; AFLP) of A. maculatum across this geographic barrier (Espíndola and Alvarez, 2011). Arum maculatum occurs only up to the submontane level, thus being absent in the Central Alps (Eggenberg et al., 2018). The insects are attracted by the strong dung-like inflorescence scent and not by visual cues of A. maculatum (Gfrerer et al., unpublished data) while looking for oviposition sites and/or mating partners (Kite et al., 1998; Espíndola and Alvarez, 2011). Previous analyses have shown that the scent profile of A. maculatum consists of up to 60 compounds, also differing among populations in their composition (Diaz and Kite, 2002; Chartier et al., 2013; Marotz-Clausen et al., 2018; Szenteczki et al., 2021; and references therein). At least in part, this scent variation appears to reflect the population variation in pollinator assemblages of A. maculatum across its distribution range (Chartier et al., 2013; Szenteczki et al., 2021). However, it is presently unclear whether the pollinator and genetic differences of A. maculatum north vs. south of the Alps are also reflected in the species' scent patterns. Nonetheless, it is known that the two main pollinating moth fly species have dissimilar floral scent preferences (Chartier et al., 2013; Szenteczki et al., 2021). Hence, we assume that the dissimilar scent preferences of the two fly species, along with their different floral visitation in regions north vs. south of the Alps, could have led to differing selection pressures on scent among respective regional populations of A. maculatum from north vs. south of the Alps.
In this study, we investigated the floral scent characteristics and fruit set (as an indicator for female fitness) of A. maculatum in six populations north of the Alps vs. five populations south of the Alps and tested for phenotypic selection on scent in the largest and most extensively sampled population in each of the two regions. Specifically, this study aimed to answer the following: (1) Do scent and fruit sets differ between north vs. south of the Alps, and among populations within regions? (2) Is there phenotypic selection on floral scent? If so, (3) do compounds, under selection differ between northern and southern populations? This study expects to find pronounced population differences in scent both at the inter-regional level and within the southern region, considering the differences in pollinator abundance and diversity between regions and also among southern, but not northern, populations (Espíndola et al., 2011). Additionally, when taking the different olfactory preferences of pollinator species into account, we expect lower fruit set south than north of the Alps, and different signs of selection in the most extensively sampled northern and southern populations.
Materials and Methods
Study Species and Populations
Brood-site deceptive A. maculatum is a rhizomatous perennial woodland herb (2n = 4x = 56) that is widespread throughout Western and Central Europe, including the British Isles, and reaches as far south as Italy, Northern Spain, and the Balkans (Boyce, 2006; Espíndola et al., 2010). It is thermogenically active, exhibits a sapromyiophilous pollination strategy, and emits a strong dung-like scent for attracting moth fly pollinators during the evening on the first day of anthesis (Kite et al., 1998; Marotz-Clausen et al., 2018). The inflorescence of A. maculatum consists of a spadix (fleshy spike) and a spathe (bract), is protogynous, and the anthesis lasts <2 days (Lack and Diaz, 1991; Marotz-Clausen et al., 2018). The spathe, which completely encloses the spadix during floral development, partially opens during anthesis to reveal the sterile appendix of the apical part of the spadix. This appendix produces and releases the scent for pollinator attraction, e.g., Lack and Diaz (1991) and Scheven (1994). At the base of the spadix, female (fertile and sterile) flowers are situated lowest, followed upwards by male flowers and staminodes (sterile male flowers). All flowers remain enveloped by the spathe during anthesis, forming a chamber that is closed by the staminodes throughout the female stage to prevent trapped insects from leaving. Pollinators are attracted in the evening on the first day of anthesis, during the female stage, slip and fall into the floral chamber, and are trapped overnight (Lack and Diaz, 1991; Gibernau et al., 2004; Espíndola et al., 2011). On the next morning, during the male stage, they are dusted with pollen, before being released at around noon when the staminodes and spathe wither (Lack and Diaz, 1991; Espíndola and Alvarez, 2011). After pollination in spring, red berry-like fruits develop as an infructescence until summer (Lack and Diaz, 1991).
During springtime in 2017–2019, we collected scents from randomly chosen A. maculatum individuals of six populations located north of the Alps (n = 106; Northwestern Austria: JOS, Josefiau; Central/Southern Germany: BUR, Burg Hohenstein; HOH, Hohendilching; MUR, Murnau; NEC, Horb am Neckar; Northern Switzerland: RÜM, Rümikon) and five populations from south of the Alps (n = 127; Northern Italy: DAO, Daone; LIM, Limone-Piemonte; MAH, Santa Maria Hoé; MON, Montese; UDI, Udine) (Figure 1 and Supplementary Table 1). We kept a minimum distance of 1 m between sampled individuals to avoid sampling potential clones, as A. maculatum can also propagate vegetatively by fragmenting rhizomes (Lack and Diaz, 1991). In summer, we harvested fruits from all individuals surveyed for scent. At most sites, we recorded scent and the fruit set of 15 individuals, except for each of the largest population per region (JOS and DAO; n = 70 each), and a northern population (HOH; n = 7) where only a few individuals had flowered at the time of scent sampling (Figure 1 and Supplementary Table 1).
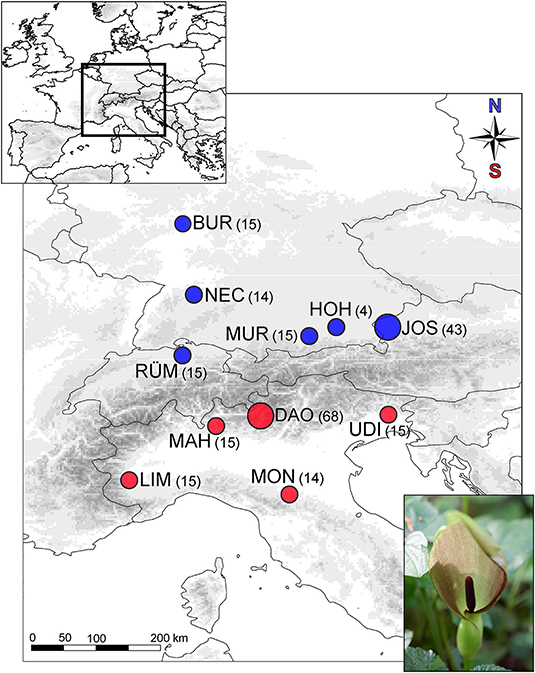
Figure 1. Sampling localities of Arum maculatum from the north (blue) vs. south (red) of the Alps. Numbers in brackets give the number of individuals used for scent (and selection) analyses. The two most extensively sampled populations (JOS, DAO) are indicated by larger circles. North: JOS, Josefiau; BUR, Burg Hohenstein; HOH, Hohendilching; MUR, Murnau; NEC, Horb am Neckar; RÜM, Rümikon; South: DAO, Daone; LIM, Limone-Piemonte; MAH, Santa Maria Hoè; MON, Montese; UDI, Udine.
Plant Volatile Collection and Analysis
Scent sampling took place on the first day of anthesis during the female stage between 6.00 p.m. and 7:30 p.m. which is the period of maximum scent emission (Marotz-Clausen et al., 2018), employing a non-invasive dynamic headspace technique. We enclosed each inflorescence in situ using a plastic oven bag (c. 30 cm × 12 cm; Toppits®, Melitta, Germany) and immediately collected scent for 5 min at 200 ml min−1 on adsorbent tubes (inner diameter: 2 mm) filled with a mixture of Tenax-TA (mesh 60–80) and Carbotrap B (mesh 20–40; 1.5 mg each; both Supelco, Germany), using a battery-operated vacuum pump (rotary vane pump G12/01 EB, Gardner Denver Austria GmbH, Vienna, Austria; Marotz-Clausen et al., 2018). In the same way, we collected scent samples from leaves and ambient air as negative controls in each population.
The dynamic headspace samples were analysed by thermal desorption-gas chromatography/mass spectrometry (TD-GC/MS; Marotz-Clausen et al., 2018), and obtained data were handled using GCMSolution v.4.41 (Shimadzu Corporation, Kyoto, Japan) (see Supplementary Methods 1 for details). Compounds were chemically identified by comparison of Kováts' retention indices (KRIs), based on commercially available n-alkanes (C7-C20; Sigma Aldrich, Vienna, Austria), and mass spectra to data available in the libraries of Adams (Adams, 2007), FFNSC 2, Wiley9, NIST11, and ESSENTIAL OILS (available in MassFinder 3, Hochmuth Scientific Consulting, Hamburg, Germany). We established our own library of mass-spectral and KRIs for semi-automatic analysis (Supplementary Methods 1). Whenever possible, compounds were verified by comparison with authentic reference compounds available in the collection of the Plant Ecology Lab of Salzburg University or with chemically synthesised reference compounds (Supplementary Methods 2). In total, 233 scent samples yielded a sufficiently informative chromatogram and were included in the analyses (Figure 1). Ultimately, a compound was only considered if it occurred in more than three scent samples and did not occur in leaf and air controls.
Fruit Set
Percentage fruit set, i.e., number of fruits by total number of flowers per individual × 100, was determined as a measure of female reproductive success. For selection analyses, we further estimated relative fruit set, i.e., number of fruits per individual divided by mean number of fruits per given population, (e.g., Gross et al., 2016), for the most extensively sampled populations JOS and DAO. In one southern population (MON), a shallow landslide destroyed all plants, with the exception of one; hence, this population was excluded from fruit set analyses.
Statistical Analyses
Geographic Patterns in Scent and Fruit Set Data
In order to test for geographic differences in floral scent, we performed permutational multivariate ANOVA (permANOVAs; Anderson, 2001) as implemented in the R package vegan v.2.6-6 (Oksanen et al., 2019). This was carried out on (1) pairwise Bray–Curtis dissimilarities of either absolute or relative scent data, i.e., the absolute amount of single compounds or the relative amount of single compounds in relation to the total amount of scent in a sample, respectively; (2) pairwise Euclidean distances of both total absolute emission of scent and the total number of floral volatiles per individual. In all these analyses, we used region (north vs. south of the Alps) and population nested in region as explanatory variables (9,999 permutations). Using permANOVA (population as an explanatory variable, 9,999 permutations), we also tested for differences in relative and absolute scent between the two most extensively sampled northern (JOS) and southern (DAO) populations by using either (1) all compounds, (2) only those that were under selection and correlated with relative fruit in the elastic net/Boruta analyses (see below and Supplementary Methods 3), or (3) those that were not under selection and did not correlate with relative fruit set.
The Bray–Curtis dissimilarity matrices, based on absolute and relative scent data across all populations, were further used to conduct canonical analyses of principal coordinates (CAPs; Anderson and Willis, 2003) with population as factor, using the capscale function in vegan (Oksanen et al., 2019) to visualise similarities and dissimilarities in scent among the samples. For each ordination, we also calculated vectors that represent compounds most correlating with the axes (Pearson correlations with capscale scores, r > |0.5|, corrected for false-discovery rate; Benjamini and Hochberg, 1995). Given that CAP is not appropriate to display similarities and dissimilarities in scent between only two populations in a two-dimensional ordination, we used non-metric multidimensional scaling (nDMS) to visualise similarities and dissimilarities in scent among the samples of only JOS and DAO, using only compounds that correlated with relative fruit set or those that did not.
Additionally, we subjected the absolute and relative scent data to random forest analyses (Breiman, 2001) in the R package randomForest v.4.6-14 (Liaw and Wiener, 2002) (ntree = 9,999 bootstrap samples with mtry = 17) to evaluate the distinctness in the scent of northern and southern samples (factor region) and among populations within each region (factor population). Distinctness was quantified as the average out-of-bag (OOB) error estimate (in %), i.e., the more distinct, the lower the OOB error. From the resulting randomForest objects, we further extracted the importance measurements to determine volatiles that are critical for regional distinction.
To test for relationships between the dissimilarity of median absolute and relative scent properties of populations and their geographic distances (in kilometres), we performed Mantel tests with the function mantel in vegan (9,999 permutations, Spearman's rank correlation). To assess whether absolute amounts of single compounds under selection (see below) differ between the two regions, we performed Mann–Whitney U-tests. Differences in fruit set across regions and among populations within regions were assessed by an ANOVA (regions and populations nested within regions as factors).
Analyses of Phenotypic Selection
To estimate the direction and strength of phenotypic selection on scent compounds, we tested for phenotypic selection (Lande and Arnold, 1983) in the northern JOS and southern DAO populations by correlating relative fruit set with z-transformed scent data (standardised to mean = 0, sd = 1; e.g., Parachnowitsch et al., 2012; Gross et al., 2016). These two populations cover a large part of their respective regional scent variation (see Results). As a major challenge, our dataset had a considerably higher number of factors (VOCs) than samples. Previous studies solved this by pre-selecting variables to reduce high dimensionality (e.g., Parachnowitsch et al., 2012) and performed selection analyses only on the most abundant compounds (Knauer and Schiestl, 2017), on principal component scores (e.g., Gross et al., 2016), or physiologically active volatiles (Chapurlat et al., 2019). Due to the very limited knowledge of attractive compounds in the study system (Scheven, 1994; Kite et al., 1998), the fact that the assumptions for principal component analysis were violated, and that also minor volatiles can be under selection (Chapurlat et al., 2019), these solutions were not suitable for our dataset. Instead, we pre-selected volatiles that correlated with relative fruit set via an elastic net, i.e., a penalised multivariate linear regression (Zou and Hastie, 2005), and via the Boruta algorithm (Kursa and Rudnicki, 2010) to identify linear (elastic net) and non-linear (Boruta) relationships between total absolute emission and the absolute emission of individual volatiles and relative fruit set (for details see Supplementary Methods 3). Additionally, the scent matrix contained many zeros (non-detects), as many compounds were quite rare (c. 70% of VOCs in <50% of samples). This zero-inflation can cause severe problems when fitting linear models, as estimates will be biased (Hogg et al., 2019). In consequence, the influence of an individual scent compound on fruit set can be either overestimated or underestimated, leading to potentially wrong conclusions. To quantify the impact of non-detects on elastic net estimates, we performed a simulation study for JOS and DAO separately before the pre-selective analyses (see Supplementary Methods 3, Supplementary Figure 1). Based on the simulation results, we obtained 93 and 81 scent compounds for JOS and DAO, respectively, each of which were then included in both the elastic net regression and the Boruta analyses (Supplementary Methods 3, Supplementary Figure 1). For the JOS population, elastic net and Boruta recovered 19 and four volatiles, respectively, whereby the latter were already among the linear ones (see Results). In the southern DAO population, no volatile correlated with fruit set in the elastic net but three in the Boruta analysis. None of these volatiles was detected for both populations (see Results). Additionally, the total absolute scent amount did not correlate with the fruit set in any of the analyses.
To ultimately test for phenotypic selection (Supplementary Figure 1), we subjected those volatiles selected by the elastic net model (Supplementary Methods 3) to multivariate linear regression (linear β-gradients; Lande and Arnold, 1983) and subjected those volatiles identified by the Boruta analyses (Supplementary Methods 3) to multivariate quadratic regression (non-linear/quadratic γ-gradients; Lande and Arnold, 1983) by squaring the terms and doubling resulting estimates (Stinchcombe et al., 2008). For the multivariate regression model of the southern (DAO) population, we excluded the plant individual “DAO076,” as it was determined by Cook's distance as an outlier influencing the model (DDAO076 = 235.4). Although elastic net handles multicollinearity well, volatiles identified to correlate with fruit set might still correlate with each other (L2 penalty, see Supplementary Methods 3). Therefore, we also tested for multicollinearity within the multivariate regression models by calculating the variance inflation factor (VIF) (R package car v.3.0.8; Fox and Weisberg, 2019) for each scent compound in each model. For the northern (JOS) model, the VIF values of various compounds were high (>5), while for the unknowns UNK1496 and UNK1503 they even exceeded 10, a threshold that indicates strong multicollinearity (Quinn and Keough, 2002). After including these two compounds as an interaction term, the VIF values of most compounds were <5, except for 3-octanol and UNK1279 (VIF>6). After further including the interaction of the latter two volatiles in the model, the VIF values of all volatiles were <4. Based on this, the final northern (JOS) model had an adjusted R2 value of 0.71. For the southern (DAO) model, all VIF values were <2 (adjusted R2 = 0.26). All statistical analyses were performed in R v.4.0.2 (R Core Team, 2020).
Results
Floral Scent
The total absolute amount of scent was highly variable among the 233 sampled individuals of A. maculatum (range of 1–2,052 ng inflorescence−1 h−1; Table 1). When taken together, northern plants released a 3-fold lower amount of scent than those from the South, along with differences among populations within regions (permANOVA: region: pseudo-F(1, 222) = 25.7, population nested within the region: pseudo-F(9, 222) = 5.36, both P < 0.001). For three of the five southern populations (MAH, MON, LIM; Figure 1), we estimated a median scent amount of c. 200 ng inflorescence−1 h−1, while DAO and UDI showed 1.5-fold higher and 5-fold lower amounts, respectively (Table 1 and Supplementary Table 1). For three of the six northern populations (MUR, NEC, RÜM; Figure 1), median estimates ranged between 40 and 81 ng inflorescence−1 h−1, while amounts in the remaining populations were manifold higher (JOS and HOH) or lower (BUR) (Table 1 and Supplementary Table 1).
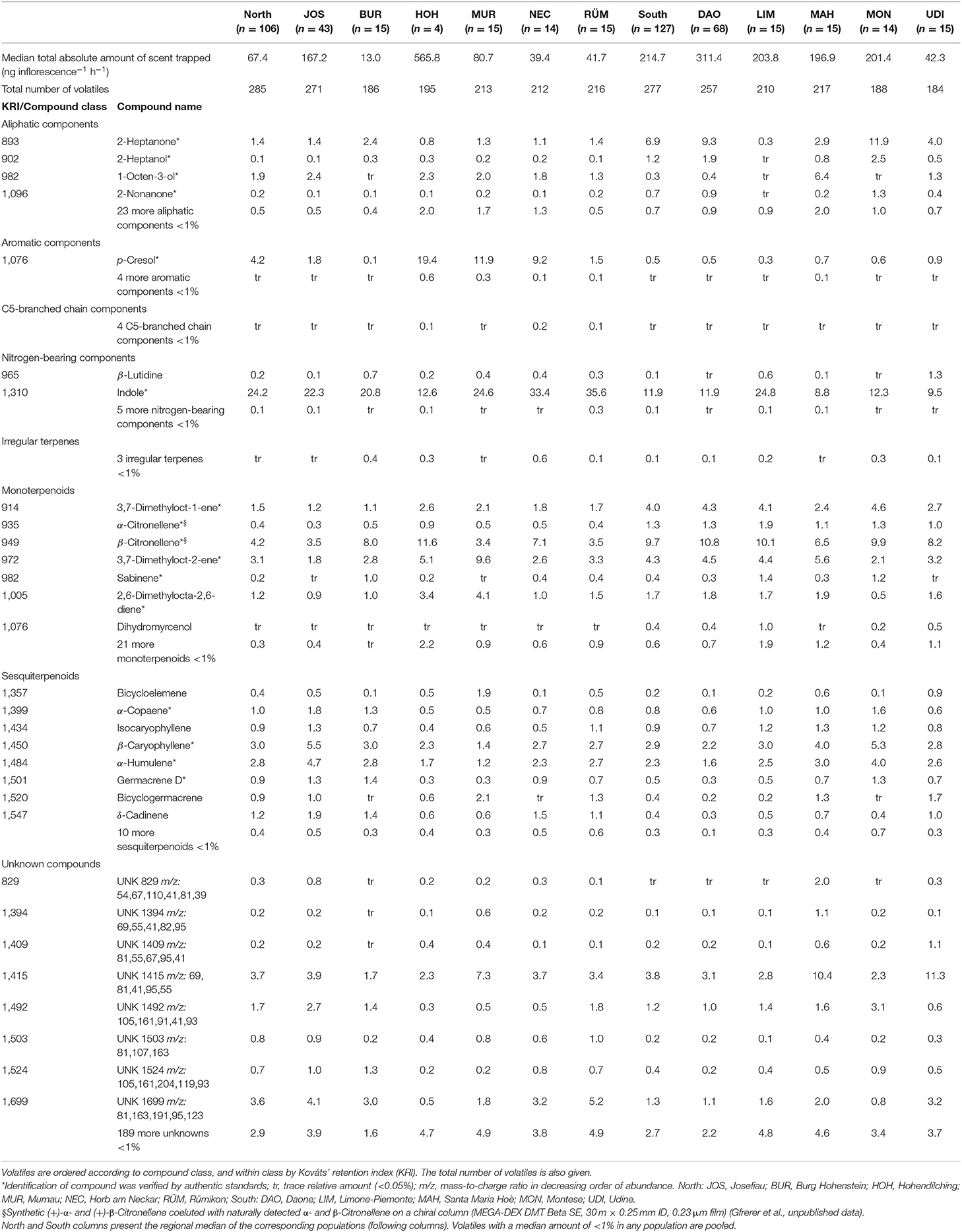
Table 1. Median amounts of total absolute and relative (contribution of single compounds to total scent) inflorescence scent of Arum maculatum surveyed in six and five populations north and south of the Alps, respectively.
Across all scent samples, we detected a total of 289 floral volatiles (285 north vs. 277 south), of which 92 could be chemically identified (Table 1 and Supplementary Table 2). A median of 102 compounds per individual was recorded (Figure 2), and the number of compounds was independent of the region (permANOVA: pseudo-F(1, 222) = 1.98, P = 0.16) but varied among populations within regions (pseudo-F(9, 222) = 4.57, P = 0.001). At the population level, between 186 (BUR) and 271 (JOS) compounds were recorded in the North, and between 188 (MON) and 257 (DAO) in the South (Figure 2). The two most extensively sampled northern (JOS) vs. southern (DAO) populations covered 96 vs. 94% of their respective regional diversity (Figure 2) and together 99% (287/289) of the total number of compounds (Table 1 and Supplementary Table 2). The five most frequent compounds found in more than 99% of the samples were the nitrogen-bearing compound indole, the monoterpenoids 3,7-dimethyloct-1-ene and β-citronellene, the sesquiterpenoid β-caryophyllene, and the unidentified UNK1492 (Table 1).
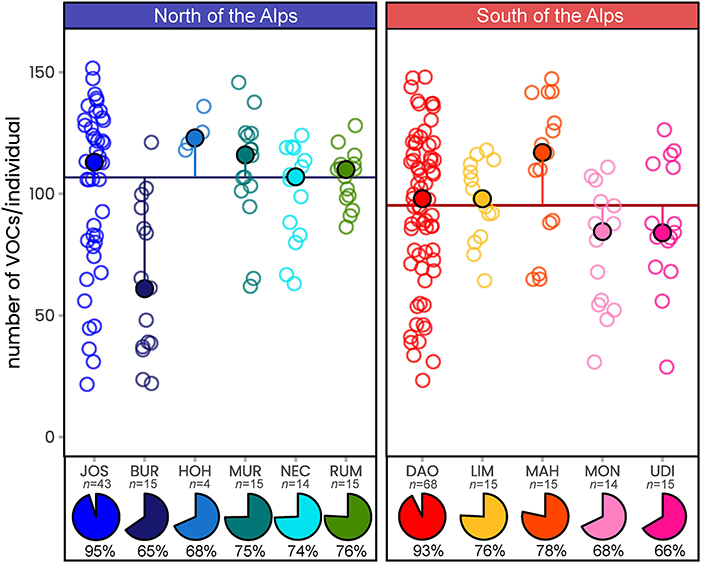
Figure 2. The number of floral scent compounds recorded in Arum maculatum individuals from populations north and south of the Alps. Filled circles denote the population median of the number of volatiles per individual; the vertical lines indicate the distance to the region median (horizontal line); open circles mark the number of volatiles detected in the individual samples. Pie charts indicate the percentage of volatiles detected per population (n, sample size) compared to the number of compounds detected across all samples (289 compounds). See Figure 1 and Supplementary Table 1 for identification of population codes.
The absolute amounts of single compounds significantly differed between regions (permANOVA: pseudo-F(1, 222) = 22.53, P < 0.001), between JOS and DAO only (pseudo-F(1, 109) = 9.95, P < 0.001), and among populations within regions (pseudo-F(9, 222) = 6.44, P < 0.001). However, differences were more pronounced between regions than among populations within regions (north vs. south OOB error: 10.3%; among populations within north OOB error: 27.3%; within south OOB error: 25.2%). Only a few abundant compounds dominated the scent bouquet of A. maculatum, including indole, β-citronellene, the unknown UNK1415, and 3,7-dimethyloct-2-ene (all abundant in both regions), p-cresol (most abundant only north), and 2-heptanone (only south, Table 1).
We also detected differences in the relative amounts of scent compounds between regions (permANOVA: pseudo-F(1, 222) = 30.18, P < 0.001), between JOS and DAO only (pseudo-F(1, 109) = 22.81, P < 0.001), and among populations within regions (pseudo-F(9, 222) = 4.90, P < 0.001; Figure 2). Again, these differences were more pronounced at the inter-regional than within-region levels (north vs. south OOB error: 9%; among populations within north OOB error: 35.8%; among populations within south OOB error: 24.4%; see also Supplementary Figure 2).
Across all populations, variation in absolute or relative amounts of scent could not be explained by their geographic distances (Mantel's Rho = 0.108, P = 0.25 and Rho = −0.154, P = 0.85, respectively).
Among the 25 compounds each that were most responsible for regional differences in the absolute and relative datasets in the randomForest analyses, 20 were common to both datasets (Supplementary Table 3). These 20 compounds included 2-heptanone, 2-heptanol, and α- and β-citronellene, all of which were more abundant (in relative and absolute amounts) south of the Alps, and 1-pentadecanol, the unknown UNK1503, p-cresol, and indole, which occurred in higher amounts north of the Alps (Table 1 and Supplementary Tables 2, 3). Many of these compounds, and some non-overlapping ones (absolute: α-copaene, β-caryophyllene; relative: UNK1409, bicyclogermacrene), explained most of the variation in scent among all samples (for relative data see Figure 3; for absolute data see Supplementary Table 4).
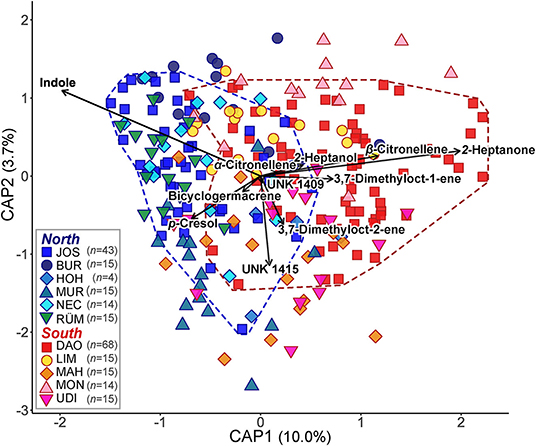
Figure 3. Canonical analysis of principal coordinates (CAP) based on a Bray–Curtis dissimilarity matrix of relative floral scent in Arum maculatum individuals from populations north and south of the Alps. n denotes the sample size per population. The vectors depict the volatiles most correlating with the capscale scores. The coloured dashed lines delineate the individual scent variation of the two most extensively sampled populations JOS (blue) and DAO (red). See Figure 1 and Supplementary Table 1 for identification of population codes.
There was also a considerably high variation in scent within populations, most prominently in the most extensively sampled northern (JOS) and southern (DAO) populations, which harboured almost all of the absolute and relative scent variation of their respective regions (for relative data see Figure 3).
Fruit Set
Among the 233 individuals surveyed for inflorescence scent, 113 set fruit in summer. Percentages of fruit set were significantly higher north of the Alps (42 ± 41% mean ± SD, 0–100% Min–Max) than south of the Alps [26 ± 33% mean ± SD, 0–100% Min–Max; Figure 4; region: F(1, 209) = 10.11, P = 0.002] and differed significantly among populations within regions [population nested within region: F(8, 209) = 2.23, P = 0.03].
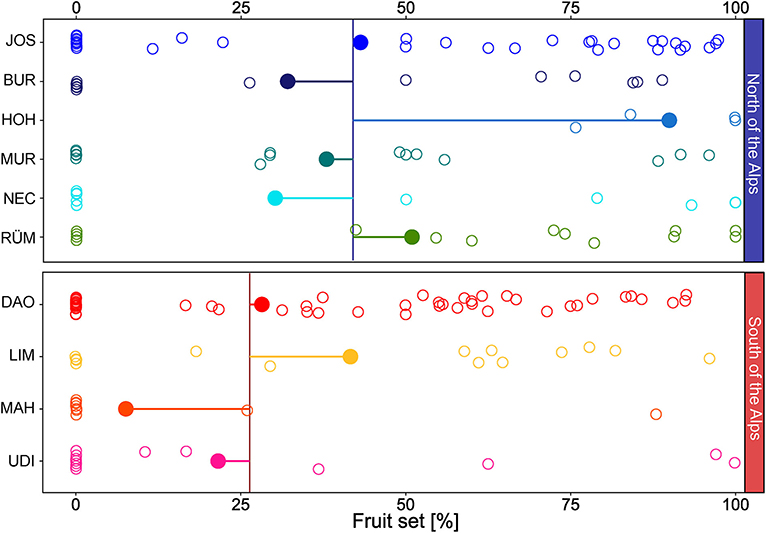
Figure 4. Fruit set (% female flowers that developed into fruits) of Arum maculatum individuals from populations north and south of the Alps. Filled circles denote the population mean of the fruit set; horizontal lines indicate the distance to the region mean (vertical line); the open circles mark the fruit set of each individual. See Figure 1 and Supplementary Table 1 for identification of population codes.
Phenotypic Selection on Scent
In the most extensively sampled northern (JOS) and southern (DAO) populations, we tested 19 and three compounds for phenotypic selection, respectively, as they correlated with relative fruit set in the elastic net and Boruta analyses (see section Materials and Methods; Supplementary Methods 3). Among those 22 compounds, seven showed signals of linear phenotypic selection (two of which as an interaction), all in the north, and two for non-linear (quadratic) phenotypic selection, all in the south (Figure 5). Seven of the overall nine compounds that were under phenotypic selection correlated positively with relative fruit set (linear: 2-heptanol, 2-nonanol, α-terpinene, UNK681, and UNK1496 together with UNK1503; non-linear: sabinene), while two correlated negatively (linear: UNK960; non-linear: 4-terpinenol; Figure 5 and Supplementary Table 5).
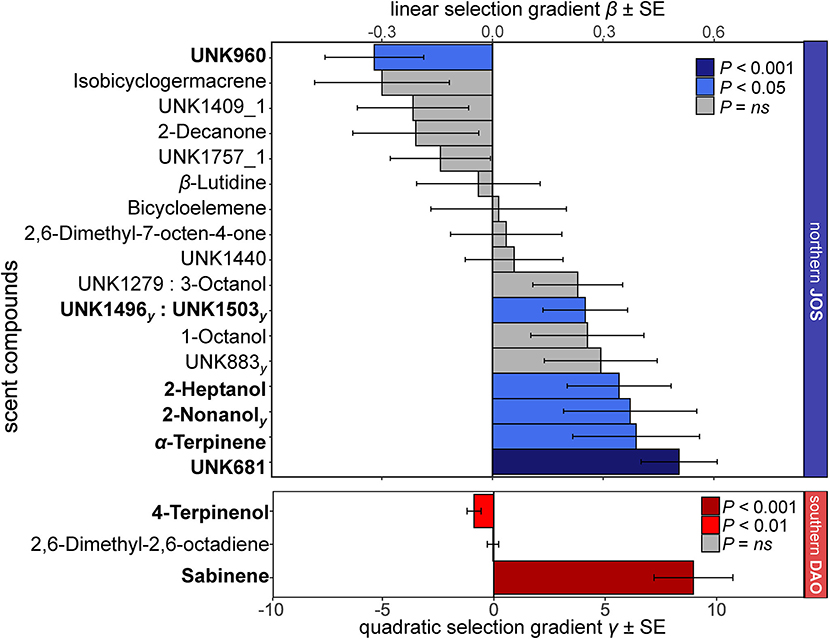
Figure 5. Linear selection gradients β and non-linear quadratic selection gradients γ (and their standard errors, SE) for individual floral scent compounds in the most extensively sampled Arum maculatum populations from north (JOS, blue, n = 43) and south (DAO, red, n = 68) of the Alps. Only compounds that correlated with relative fruit set in the elastic net/Boruta analyses are shown (see Material and Methods). Scent compounds under significant selection (P < 0.05) are in bold and their bars are coloured. Note the different scaling for linear (β) and non-linear (γ) selection. For the northern population, compounds that were also detected by the non-linear Boruta analyses are indicated with a subscript (γ).
Among the 25 compounds that most strongly contributed to the absolute (and relative) differences in scent between the regions, only four were under selection (north: 2-heptanol, 2-nonanol, UNK681, UNK1503), but not others (e.g., 2-heptanone, α- and β-citronellene; Supplementary Table 3). Differences in absolute and relative scent traits between the northern JOS and the southern DAO populations remained significant, regardless of performing permANOVA separately on the nine compounds that correlated with relative fruit set in the elastic net/Boruta analyses and were under selection [absolute vs. relative datasets: pseudo-F(1, 109) = 18.4 vs. 30.8, both P < 0.001], or on the 93 compounds that were not under selection and did not correlate with fruit set [absolute vs. relative datasets: pseudo-F(1, 109) = 9.8 vs. 24.5, both P < 0.001; see Supplementary Figure 3, Supplementary Methods 3].
Discussion
Our study shows that A. maculatum has hyperdiverse inflorescence scents that, in agreement with genetic and pollinator patterns, differ in their composition between populations north vs. south of the Alps. Scent differed not only among southern but also among northern populations, although the pollinator spectrum only differed in populations south of the Alps (Espíndola et al., 2011; Laina et al., unpublished data). As expected, samples from the southern populations had lower fruit set than northern ones, which agrees with lower insect numbers reported in the south (Espíndola et al., 2011; Laina et al., unpublished data), and different signs of phenotypic selection were found in the most extensively sampled northern and southern populations.
Hyperdiversity of Floral Scent
With 289 floral volatiles recorded, the inflorescence scent diversity of A. maculatum is extraordinarily high and not matched by any other plant species to the best of our knowledge. In fact, we are not aware of any species from which more than 200 floral compounds are reported, a number that a single A. maculatum individual can reach by three quarters (max. = 152 VOCs; Figure 2). This difference in the number of scent compounds between A. maculatum and other species cannot just be explained by differences in techniques used for scent analyses, given that scents of a high number of species were analysed using a similar approach as we did (dynamic headspace and thermal desorption of samples; Gottsberger et al., 2013; Borchsenius et al., 2016; Lukas et al., 2019). Species closest to the high number of VOCs in A. maculatum include the sapromyiophilous Sauromatum guttatum (Araceae, with altogether 196 different VOCs; Skubatz et al., 1996; Hadacek and Weber, 2002) and Aristolochia gigantea (Aristolochiaceae, 168 VOCs; Martin et al., 2017), as well as the insect-pollinated and rewarding Geonoma macrostachys (Arecaceae, 176 VOCs; Borchsenius et al., 2016) and Echinopsis ancistrophora (Cactaceae, 145 VOCs; Schlumpberger and Raguso, 2008). Other species for which c. 100 VOCs are described likewise include insect-pollinated and rewarding species [e.g., Philodendron bipinnatifidum (Araceae), Gottsberger et al., 2013; Pyrus communis (Rosaceae), Lukas et al., 2019], but also the sexually deceptive orchid Ophrys sphegodes (Ayasse et al., 2000). Thus, high numbers of compounds are found across a wide range of plant families and are apparently not restricted to a specific pollination system.
One explanation for the high diversity of scent compounds in A. maculatum is that this species likely imitates the various breeding substrates of its moth fly pollinators, all potentially differently scented. The two main pollinators, P. phalaenoides and P. grisescens, breed in a variety of different substrates such as rotting manure from cattle and horse, fungi (P. grisescens), waste pits, mud-flats, plant litter in drainages (P. phalaenoides), and the hygropetric zones of river banks and ponds (Satchell, 1947; Ježek, 1990; Sigsgaard et al., 2020). Arum maculatum emits compounds described from several substrates, such as cattle and horse manure (e.g., indole, p-cresol, skatole), fungi (e.g., 1-octen-3-ol, (E)-2-octen-1-ol, 3-octanone), and general degrading and fermenting plant or animal material (e.g., 2,3-heptanedione, acetoin, butanoic acid) (Dormont et al., 2010; Jürgens et al., 2013). Highly specialised deceptive plant systems frequently rely on only a few volatiles to attract pollinators; they seem to imitate a more specific model, thus releasing less complex scent blends (e.g., Wee et al., 2018).
The number of volatiles detected across the 233 samples (11 populations) of A. maculatum (289 VOCs) is five to 10 times higher than previously reported for this species (18–61, and 143 VOCs in total; Scheven, 1994; Diaz and Kite, 2002; Chartier et al., 2013; Marotz-Clausen et al., 2018; Szenteczki et al., 2021; and references therein). This discrepancy cannot be explained by differences in sample size, as a similar number of individuals were surveyed in those previous studies (n = 222 in total, representing 23 populations). Interestingly, we found a similar number of compounds in some individuals (up to 152 VOCs; median of 102; Figure 2) as overall detected previously (Diaz and Kite, 2002; Chartier et al., 2013; Marotz-Clausen et al., 2018; Szenteczki et al., 2021; and references therein). With the exception of two studies (Marotz-Clausen et al., 2018; Szenteczki et al., 2021), each sharing one of our sampled populations (JOS and MON, respectively), all previous studies sampled scents in other populations across Europe (Kite, 1995; Diaz and Kite, 2002; Chartier et al., 2013). Thus, some of the differences in the number of A. maculatum compounds detected across studies might reflect population-specific scent characteristics (see Figure 2). However, more importantly we believe that the discrepancy in the number of compounds recorded largely reflects differences in methodology between the present and previous studies. For example, these are: higher sensitivity of modern GC/MS systems, usage of less selective adsorbent agents [Carbotrap/Tenax-TA vs. polydimethylsiloxane/ divinylbenzene (Chartier et al., 2013) vs. polydimethylsiloxane (Szenteczki et al., 2021)]; in situ vs. ex situ samplings (Scheven, 1994; Marotz-Clausen et al., 2018), and including all vs. only compounds above a specific threshold in relative amounts (Chartier et al., 2013; Szenteczki et al., 2021). Among the 92 compounds chemically identified in this study, more than half (50) were previously unknown to be released by A. maculatum. Some of these newly described compounds for A. maculatum are known from other species of Araceae (e.g., α-cubebene, β-phellandrene, γ-terpinene, Sauromatum guttatum, Hadacek and Weber, 2002), or other plant families (e.g., methyl anthranilate, isobutyl butyrate, citronellal, Knudsen et al., 2006; El-Sayed, 2019). To the best of our knowledge, this study is, however, the first to identify p-cresyl butyrate as a floral scent compound.
Geographic Patterns of Floral Scent
In the region south of the Alps, the qualitative, absolute, and relative differences in scent among populations of A. maculatum may be related to the pollinator assemblages that are in this region more diverse in terms of abundance, species composition, and sex ratio (Espíndola et al., 2011; Laina et al., unpublished data). In the region north of the Alps, females of P. phalaenoides are the principal pollinators in all studied populations (Espíndola et al., 2011; Laina et al., unpublished data), even though other Psychoda spp. also occur in this region (Chartier et al., 2013; Laina et al., unpublished data). Hence, the scent variation we observed among northern populations is not reflected by variations in pollinator spectra in this region. In this study, all variations in scent were more pronounced between regions than among populations within each region. This strong regional component of scent variation in A. maculatum across the Alps thus accords with strong differences in pollinator spectra (Espíndola et al., 2011; Laina et al., unpublished data) and coincides with a genetic (AFLP) subdivision of A. maculatum across this geographic barrier (Espíndola and Alvarez, 2011). Differing pollinator availability has been linked to different climatic factors (Espíndola et al., 2011), which might also influence scent variation (e.g., Farré-Armengol et al., 2014). However, a preliminary transplant experiment shows that A. maculatum individuals originating from north or south of the Alps keep their population-typic scent after transplantation (Gfrerer et al., unpublished data). This suggests that abiotic factors do not directly influence scent emissions (see also Szenteczki et al., 2021). However, we cannot exclude that they might exert differential selection pressures, thus influencing evolutionary processes that may lead to differences in scent emission between the regions. Previous studies in A. maculatum also found population effects in scent composition, e.g., Chartier et al. (2013) and Szenteczki et al. (2021). Nonetheless, our study is the first to demonstrate such population differentiation in scent across the Alps. Intraspecific variation in floral scent among populations and regions has also been reported for other plant species (e.g., Dötterl et al., 2005; Chapurlat et al., 2019; and Schlumpberger and Raguso, 2008), including sapromyiophilous species (e.g., Chen et al., 2017). In some of those, this variation, as shown in this study, could be linked to pollinator assemblages and/or genetic patterns (e.g., Chapurlat et al., 2019), but not in others (Dötterl et al., 2005; Schlumpberger and Raguso, 2008).
Phenotypic Selection on Floral Scents
The two most extensively sampled northern (JOS) and southern (DAO) populations differed in absolute and relative amounts of scent, regardless of whether the analyses were conducted on all compounds, on only those that correlated with relative fruit set and were under selection, or on those that did not correlate with fruit set (Material and methods, Supplementary Figure 3). Thus, this regional difference in scent could be caused by different selection regimes, as well as other reasons, such as phenotypic plasticity (but see Szenteczki et al., 2021) or genetic drift (Herrera et al., 2006; Majetic et al., 2009). In support of differential selection, we detected population-specific signatures of phenotypic selection on scent in JOS and DAO, possibly due to different olfactory preferences of those Psychoda species that dominate the pollinator spectra of A. maculatum in the northern (female P. phalaenoides) vs. southern (and P. grisescens) regions (Espíndola et al., 2011; Chartier et al., 2013; Szenteczki et al., 2021; Laina et al., unpublished data).
For the five compounds under phenotypic selection that we were able to chemically identify, i.e., 2-nonanol, 2-heptanol, sabinene, 4-terpinenol and α-terpinene, information on their attractiveness to pollinators of A. maculatum is lacking. However, the aliphatic compounds 2-heptanol and 2-nonanol are known, either together or alone, as attractants for bees (Meliponini, Pianaro et al., 2009) and kleptoparasitic flies (Heiduk et al., 2016). They are also known as (sex-)pheromones of female Diptera (Cecidomyiidae, Censier et al., 2014) and female non-Diptera (Trichoptera, Löfstedt et al., 1994). The monoterpenoids sabinene, α-terpinene, and 4-terpinenol are defence substances of some insects (Coleoptera, e.g., Wheeler et al., 2002; Lepidoptera, Ômura et al., 2006) but are used by others (e.g., Lepidoptera, Baur et al., 1993) as oviposition stimulants. The latter two volatiles are also pheromones of fruit flies (Fletcher et al., 1992). In summary, these five compounds, found to be under phenotypic selection, elicit responses in insects other than moth flies. Furthermore, they are known from the floral scent of other sapromyiophilous species (e.g., Hadacek and Weber, 2002; Johnson and Jürgens, 2010), and some of them (α-terpinene and 4-terpinenol) are also known from cattle dung (Dormont et al., 2010; Sládeček et al., 2021), i.e., one of the oviposition substrates of moth flies. Further research is required to establish whether these five compounds, which are all widespread floral scent compounds (Knudsen et al., 2006; El-Sayed, 2019), are attractive to the pollinators of A. maculatum.
Several of the compounds most responsible for regional differences in inflorescence scent, e.g., 2-heptanone, 3,7-dimethyloct-1-ene, UNK966 (Supplementary Table 3), did not show signals of phenotypic selection (see Figure 5). Thus, the different selection regimes cannot explain several of the most obvious differences in scent between A. maculatum from north and south of the Alps (see also Supplementary Figure 2). However, some other compounds that also differed in absolute amounts between regions (2-heptanol, 2-nonanol, UNK681, sabinene; Supplementary Tables 3, 5) were under phenotypic selection, either in northern JOS or southern DAO (Figure 5), and some of the differences between regions could, therefore, be due to differential selection.
Somewhat unexpectedly, we did not find phenotypic selection for the most abundant compounds in the scent of A. maculatum, e.g., indole, β-citronellene, unknown UNK1415, with the exception of 2-heptanol (Figure 5). Even more surprisingly, we also did not find phenotypic selection for those compounds known to attract P. phalaenoides, i.e., indole, 2-heptanone, p-cresol, and α-humulene (Scheven, 1994; Kite et al., 1998), occurring both north and south of the Alps, and also in JOS and DAO (Espíndola et al., 2011). This contrasts with most other studies, where main compounds and/or pollinator attractants showed signals of phenotypic selection (but see Chapurlat et al., 2019). In Penstemon digitalis (Plantaginaceae), one of the main compounds, linalool, was under phenotypic selection and attractive to bumblebees in the laboratory but not in field bioassays (Parachnowitsch et al., 2012; Burdon et al., 2020). Possible explanations for not finding phenotypic selection on the main compounds of A. maculatum include the following: (1) their released amounts are high enough to achieve maximum pollinator attractiveness (see also Chapurlat et al., 2019); (2) there are opposing selection pressures on these compounds by different pollinators or herbivores, resulting in zero ‘net' selection (e.g., Knauer and Schiestl, 2017; Chapurlat et al., 2019); and (3) their relationship with flower visitors is non-linear and non-quadratic (e.g., Galen et al., 2011). Although our multivariate models detected non-linear phenotypic selection by including quadratic terms, such quadratic analyses cannot uncover all potential non-linear relationships (e.g., Stinchcombe et al., 2008). Hence, we cannot exclude the possibility that such abundant and/or attractive compounds are still under phenotypic selection, which in turn calls for future statistical developments that allow testing for any kind of non-linear multivariate relationships.
Deceptive plant species might experience stronger selection than rewarding ones (Sletvold and Ågren, 2014). However, by comparison with rewarding species (Parachnowitsch et al., 2012; Gross et al., 2016; Gervasi and Schiestl, 2017; Chapurlat et al., 2019), we found that deceptive A. maculatum does not release a higher number of volatiles with signatures of phenotypic selection (7 vs. 3–42%), but these volatiles appear to be under slightly stronger positive linear phenotypic selection (−0.3 to 0.5 vs. −0.3 to 0.4, Min to Max; Figure 5; Parachnowitsch et al., 2012; Gross et al., 2016; Chapurlat et al., 2019) and stronger non-linear phenotypic selection (−0.9 to 9 vs. −0.5 to −0.3, Min to Max; Figure 5; Gervasi and Schiestl, 2017). Future studies on other deceptive plant species that also attract specific pollinators by chemical cues, but have lower levels of fruit set than A. maculatum (such as many orchids, e.g., Tremblay et al., 2005), might reveal even stronger signatures of phenotypic selection.
Conclusions
Our study on sapromyiophilous A. maculatum reported the highest number of floral volatiles ever found in a single plant species to date. This chemical hyperdiversity could be due to the fact that A. maculatum imitates the odours of a multitude of differently scented breeding substrates of its moth fly pollinators, e.g., dung, fungi, and rotting plant material. We recorded pronounced scent differences between populations from north vs. south of the Alps, and this geographic pattern in scent agrees with previously described pollinator and genetic patterns across this geographic barrier. For the first time, the results of this study provide evidence that floral scents of a deceptive plant are under phenotypic selection and suggest that populational and regional differences in scent are partly due to differential selection, while other reasons such as phenotypic plasticity and genetic drift cannot be excluded. The biological role of most compounds under selection is unknown and awaits determination in future studies in A. maculatum and other plants where phenotypic selection on scent was demonstrated (Parachnowitsch et al., 2012; Gross et al., 2016; Gervasi and Schiestl, 2017; Knauer and Schiestl, 2017; Chapurlat et al., 2019).
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, and the R code for the simulation and the full scent dataset can be found in the Dryad Digital Repository (https://doi.org/10.5061/dryad.pnvx0k6kn).
Author Contributions
SD, MG, AH, and HC designed the research. EG and DL conducted the fieldwork. RF executed the scent sample laboratory work. EG and SD built the scent library. EG analysed all scent and fruit set data, designed and performed the selection analyses, and wrote the first draft of the manuscript. TT identified and synthesised unknown compounds. MH, WT, RF, SD, and EG discussed statistical approaches for selection analyses. MH designed and performed the simulations. All authors contributed to the final version.
Funding
This study was funded by a grant from the Austrian Science Fund (FWF; P30175-B29) to AH, HC, and SD (PI). All samplings were carried out in compliance with the current laws of the respective countries. MH and WT gratefully acknowledge support from the WISS 2025 project ‘Lab for Intelligent Data Analytics Salzburg’ (20204-WISS/225/197-2019 and 20102-F1901166-KZP).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank Irmgard Schäffler for methodological support, Valerie Scheurecker, Karin Moosbrugger, Bernadette Mükisch, S. Sophie Brandauer, and Christopher Gorofsky for their support in the field, and Florian Schiestl for providing data on populations in Switzerland. We also thank Robert A. Raguso for discussing hyperdiverse scents, and members of the Plant Ecology group of the University of Salzburg, especially Karin Gross and Herbert Braunschmid, for constructive comments on an earlier version of the manuscript. The previous version of this manuscript has appeared online at bioRxiv (10.1101/2021.04.28.441155).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2021.719092/full#supplementary-material
References
Ackerman, J. D., Cuevas, A. A., and Hof, D. (2011). Are deception-pollinated species more variable than those offering a reward? Plant Syst. Evolut. 293, 91–99. doi: 10.1007/s00606-011-0430-6
Adams, R. P. (2007). Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry. Carol Stream, IL: Allured Publishing Corporation.
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26, 32–46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
Anderson, M. J., and Willis, T. J. (2003). Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 84, 511–525. doi: 10.1890/0012-9658(2003)084[0511:CAOPCA]2.0.CO;2
Ayasse, M., Schiestl, F. P., Paulus, H. F., Löfstedt, C., Hansson, B., Ibarra, F., et al. (2000). Evolution of reproductive strategies in the sexually deceptive orchid Ophrys sphegodes: how does flower-specific variation of odor signals influence reproductive success? Evolution 54, 1995–2006. doi: 10.1111/j.0014-3820.2000.tb01243.x
Baur, R., Feeny, P., and Städler, E. (1993). Oviposition stimulants for the black swallowtail butterfly: identification of electrophysiologically active compounds in carrot volatiles. J. Chem. Ecol. 19, 919–937. doi: 10.1007/BF00992528
Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. B 57, 289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x
Borchsenius, F., Lozada, T., and Knudsen, J. T. (2016). Reproductive isolation of sympatric forms of the understorey palm Geonoma macrostachys in western Amazonia. Botanical J. Linnean Soc. 182, 398–410. doi: 10.1111/boj.12428
Burdon, R. C. F., Raguso, R. A., Gegear, R. J., Pierce, E. C., Kessler, A., and Parachnowitsch, A. L. (2020). Scented nectar and the challenge of measuring honest signals in pollination. J. Ecol. 108, 2132–2144. doi: 10.1111/1365-2745.13432
Censier, F., Fischer, C. Y., Chavalle, S., Heuskin, S., Fauconnier, M. L., Bodson, B., et al. (2014). Identification of 1-methyloctyl butanoate as the major sex pheromone component from females of the saddle gall midge, Haplodiplosis marginata (Diptera: Cecidomyiidae). Chemoecology 24, 243–251. doi: 10.1007/s00049-014-0167-0
Chapurlat, E., Ågren, J., Anderson, J, Friberg, M., and Sletvold, N. (2019). Conflicting selection on floral scent emission in the orchid Gymnadenia conopsea. New Phytol. 222, 2009–2022. doi: 10.1111/nph.15747
Chartier, M., Pélozuelo, L., Buatois, B., Bessière, J. M., and Gibernau, M. (2013). Geographical variations of odour and pollinators, and test for local adaptation by reciprocal transplant of two European Arum species. Funct. Ecol. 27, 1367–1381. doi: 10.1111/1365-2435.12122
Chen, G., Gong, W. C., Ge, J., Schinnerl, J., Wang, B., and Sun, W. B. (2017). Variation in floral characters, particularly floral scent, in sapromyophilous Stemona species. J. Integr. Plant Biol. 59, 825–839. doi: 10.1111/jipb.12580
Chittka, L., and Thomson, J. D. (2001). Cognitive Ecology of Pollination: Animal Behavior and Floral Evolution. Cambridge: Cambridge University Press.
Delle-Vedove, R., Schatz, B., and Dufay, M. (2017). Understanding intraspecific variation of floral scent in light of evolutionary ecology. Ann. Bot. 120, 1–20. doi: 10.1093/aob/mcx055
Diaz, A., and Kite, G. C. (2002). A comparison of the pollination ecology of Arum maculatum and Arum italicum in England. Watsonia 24, 171–181.
Dormont, L., Delle-Vedove, R., Bessière, J. M., and Schatz, B. (2014). Floral scent emitted by white and coloured morphs in orchids. Phytochemistry 100, 51–59. doi: 10.1016/j.phytochem.2014.01.009
Dormont, L., Jay-Robert, P., Bessière, J. M., Rapior, S., and Lumaret, J. P. (2010). Innate olfactory preferences in dung beetles. J. Exp. Biol. 213, 3177–3186. doi: 10.1242/jeb.040964
Dötterl, S., Wolfe, L. M., and Jürgens, A. (2005). Qualitative and quantitative analyses of flower scent in Silene latifolia. Phytochemistry 66, 203–213. doi: 10.1016/j.phytochem.2004.12.002
Eggenberg, S., Bornand, C., Juillerat, P., Jutzi, M., Möhl, A., Nyffeler, R., et al. (2018). Flora Helvetica - Exkursionsführer. Bern: Haupt Verlag.
El-Sayed, A. M. (2019). The Pherobase: Database of Pheromones and Semiochemicals. Available online at: http://www.pherobase.com/ (accessed November 10, 2020).
Espíndola, A., and Alvarez, N. (2011). Comparative phylogeography in a specific and obligate pollination antagonism. PLoS ONE 6:e28662. doi: 10.1371/journal.pone.0028662
Espíndola, A., Buerki, S., Bedalov, M., Küpfer, P., and Alvarez, N. (2010). New insights into the phylogenetics and biogeography of Arum (Araceae): unravelling its evolutionary history. Bot. J. Linnean Soc. 163, 14–32. doi: 10.1111/j.1095-8339.2010.01049.x
Espíndola, A., Pellissier, L., and Alvarez, N. (2011). Variation in the proportion of flower visitors of Arum maculatum along its distributional range in relation with community-based climatic niche analyses. Oikos 120, 728–734. doi: 10.1111/j.1600-0706.2010.18937.x
Farré-Armengol, G., Filella, I., Llusi,à, J, Niinemets, Ü., and Peñuelas, J. (2014). Changes in floral bouquets from compound-specific responses to increasing temperatures. Glob. Chang. Biol. 20, 3660–3669. doi: 10.1111/gcb.12628
Fletcher, M. T., Wells, J. A., Jacobs, M. F., Krohn, S., Kitching, W., Drew, R. A. I., et al. (1992). Chemistry of fruit-flies. Spiroacetal-rich secretions in several Bacfrocera species from the South-West Pacific Region. J. Chem. Soc. Perkin Trans. 1, 2827–2831. doi: 10.1039/p19920002827
Fox, J., and Weisberg, S. (2019). An R Companion to Applied Regression, 3rd Edn. Thousand Oaks, CA: Sage Publications.
Friberg, M., Schwind, C., Roark, L. C., Raguso, R. A., and Thompson, J. N. (2014). Floral scent contributes to interaction specificity in coevolving plants and their insect pollinators. J. Chem. Ecol. 40, 955–965. doi: 10.1007/s10886-014-0497-y
Galen, C., Kaczorowski, R., Todd, S. L., Geib, J., and Raguso, R. A. (2011). Dosage-dependent impacts of a floral volatile compound on pollinators, larcenists, and the potential for floral evolution in the Alpine skypilot Polemonium viscosum. Am. Nat. 177, 258–272. doi: 10.1086/657993
Gervasi, D. D. L., and Schiestl, F. P. (2017). Real-time divergent evolution in plants driven by pollinators. Nat. Commun. 8, 1–8. doi: 10.1038/ncomms14691
Gibernau, M., Macquart, D., and Przetak, G. (2004). Pollination in the genus Arum – a review. Aroideana 27, 148–166.
Gottsberger, G., Silberbauer-Gottsberger, I., and Dötterl, S. (2013). Pollination and floral scent differentiation in species of the Philodendron bipinnatifidum complex (Araceae). Plant Syst. Evol. 299, 793–809. doi: 10.1007/s00606-013-0763-4
Gross, K., Sun, M., and Schiestl, F. P. (2016). Why do floral perfumes become different? Region-specific selection on floral scent in a terrestrial orchid. PLoS ONE 11:e147975. doi: 10.1371/journal.pone.0147975
Hadacek, F., and Weber, M. (2002). Club-shaped organs as additional osmophores within the Sauromatum inflorescence: odour analysis, ultrastructural changes and pollination aspects. Plant Biol. 4, 367–383. doi: 10.1055/s-2002-32335
Heiduk, A., Göhl, M., Jürgens, A., Dötterl, S., Brake, I., Meve, U., et al. (2016). Ceropegia sandersonii mimics attacked honeybees to attract kleptoparasitic flies for pollination. Current Biology 26, 2787–2793. doi: 10.1016/j.cub.2016.07.085
Herrera, C. M., Castellanos, M. C., and Medrano, M. (2006). “Geographical context of floral evolution: towards an improved research programme in floral diversification,” in Ecology and Evolution of Flowers, eds L. D. Harder and S. C. H. Barrett (Oxford: Oxford University Press), 278–294.
Hogg, R. V., McKean, J. W., and Craig, A. T. (2019). Introduction to Mathematical Statistics. Boston, MA: Pearson.
Ježek, J. (1990). Redescriptions of nine common Palaearctic and Holarctic species of Psychodini End (Diptera: Psychodidae). Acta Entomol. Muesei Nationalis Pragae 43, 33–83.
Johnson, S. D., and Jürgens, A. (2010). Convergent evolution of carrion and faecal scent mimicry in fly-pollinated angiosperm flowers and a stinkhorn fungus. South Afr. J. Botany 76, 796–807. doi: 10.1016/j.sajb.2010.07.012
Jürgens, A., Wee, S.-L., Shuttleworth, A., and Johnson, S. D. (2013). Chemical mimicry of insect oviposition sites: a global analysis of convergence in angiosperms. Ecol. Lett. 16, 1157–1167. doi: 10.1111/ele.12152
Kessler, A., and Halitschke, R. (2009). Testing the potential for conflicting selection on floral chemical traits by pollinators and herbivores: predictions and case study. Funct. Ecol. 23, 901–912. doi: 10.1111/j.1365-2435.2009.01639.x
Kite, G. C. (1995). The floral odour of Arum maculatum. Biochem. Syst. Ecol. 23, 343–354. doi: 10.1016/0305-1978(95)00026-Q
Kite, G. C., Hetterscheid, W. L. A., Lewis, M. J., Boyce, P. C., Ollerton, J., Cocklin, E., et al. (1998). “Inflorescence odours and pollinators of Arum and Amorphophallus (Araceae),” in Reproductive Biology, eds S. J. Owens and P. J. Rudall (Kew: Royal Botanic Gardens), 295–315.
Knauer, A. C., and Schiestl, F. P. (2017). The effect of pollinators and herbivores on selection for floral signals: a case study in Brassica rapa. Evol. Ecol. 31, 285–304. doi: 10.1007/s10682-016-9878-8
Knudsen, J. T., Eriksson, R., Gershenzon, J., and Ståhl, B. (2006). Diversity and distribution of floral scent. Botanical Rev. 72, 1–120. doi: 10.1663/0006-8101(2006)72[1:DADOFS]2.0.CO;2
Knudsen, J. T., and Gershenzon, J. (2020). “The chemical diversity of floral scent,” in Biology of Plant Volatiles, eds E. Pichersky and N. Dudareva (Boca Raton, FL: CRC Press), 57–78.
Kursa, M. B., and Rudnicki, W. R. (2010). Feature selection with the Boruta package. J. Stat. Softw. 36, 1–13. doi: 10.18637/jss.v036.i11
Lack, A. J., and Diaz, A. (1991). The pollination of Arum maculatum L. – a historical review and new observations. Watsonia 18, 333–342.
Lande, R., and Arnold, S. J. (1983). The measurement of selection on correlated characters. Evolution 37, 1210–1226. doi: 10.1111/j.1558-5646.1983.tb00236.x
Löfstedt, C., Hansson, B. S., Petersson, E., Valeur, P., and Richards, A. (1994). Pheromonal secretions from glands on the 5th abdominal sternite of hydropsychid and rhyacophilid caddisflies (Trichoptera). J. Chem. Ecol. 20, 153–170. doi: 10.1007/BF02065998
Lukas, K., Harig, T., Schulz, S., Hadersdorfer, J., and Dötterl, S. (2019). Flowers of European pear release common and uncommon volatiles that can be detected by honey bee pollinators. Chemoecology 29, 211–223. doi: 10.1007/s00049-019-00289-x
Majetic, C. J., Raguso, R. A., and Ashman, T. L. (2009). Sources of floral scent variation. Plant Signal. Behav. 4, 129–131. doi: 10.4161/psb.4.2.7628
Marotz-Clausen, G., Jürschik, S., Fuchs, R., Schäffler, I., Sulzer, P., Gibernau, M., et al. (2018). Incomplete synchrony of inflorescence scent and temperature patterns in Arum maculatum L. (Araceae). Phytochemistry 154, 77–84. doi: 10.1016/j.phytochem.2018.07.001
Martin, K. R., Mor,é, M, Hipólito, J., Charlemagne, S., Schlumpberger, B. O., and Raguso, R. A. (2017). Spatial and temporal variation in volatile composition suggests olfactory division of labor within the trap flowers of Aristolochia gigantea. Flora 232, 153–168. doi: 10.1016/j.flora.2016.09.005
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., Mcglinn, D., et al (2019). Vegan: Community Ecology Package. CRAN. Availble online at: https://CRAN.R-project.org/package=vegan/ (accessed November 10, 2020).
Ollerton, J., and Diaz, A. (1999). Evidence for stabilising selection acting on flowering time in Arum maculatum (Araceae): the influence of phylogeny on adaptation. Oecologia 119, 340–348. doi: 10.1007/s004420050794
Ollerton, J., Winfree, R., and Tarrant, S. (2011). How many flowering plants are pollinated by animals? Oikos 120, 321–326. doi: 10.1111/j.1600-0706.2010.18644.x
Ômura, H., Honda, K., and Feeny, P. (2006). From terpenoids to aliphatic acids: further evidence for late-instar switch in osmeterial defense as a characteristic trait of swallowtail butterflies in the tribe Papilionini. J. Chem. Ecol. 32, 1999–2012. doi: 10.1007/s10886-006-9124-x
Parachnowitsch, A. L., Raguso, R. A., and Kessler, A. (2012). Phenotypic selection to increase floral scent emission, but not flower size or colour in bee-pollinated Penstemon digitalis. New Phytol. 195, 667–675. doi: 10.1111/j.1469-8137.2012.04188.x
Pianaro, A., Menezes, C., Kerr, W. E., Singer, R. B., Patricio, E. F. L. R. A., and Marsaioli, A. J. (2009). Stingless bees: chemical differences and potential functions in Nannotrigona testaceicornis and Plebeia droryana males and workers. J. Chem. Ecol. 35, 1117–1128. doi: 10.1007/s10886-009-9679-4
Quinn, G. P., and Keough, M. J. (2002). Experimental Design and Data Analysis for Biologists. Cambridge: Cambridge University Press.
R Core Team (2020). A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: http://www.R-project.org/
Raguso, R. A. (2008). Wake up and smell the roses: the ecology and evolution of floral scent. Annu. Rev. Ecol. Evol. Syst. 39, 549–569. doi: 10.1146/annurev.ecolsys.38.091206.095601
Satchell, G. H. (1947). The ecology of the British species of Psychoda (Diptera: Psychodidae). Ann. Appl. Biol. 34, 611–621. doi: 10.1111/j.1744-7348.1947.tb06392.x
Scheven, H. J. (1994). GC/MS Untersuchungen des Appendixduftes blühender Pflanzen von Arum maculatum L. und Arum italicum MILLER; Nachweis der attraktiven Wirkung der Duftbestandteile Indol, Humulen und p-Kresol auf Psychoda phalaenoides L (dissertation thesis). Philipps-Universität Marburg, Marburg, Germany.
Schiestl, F. P., and Johnson, S. D. (2013). Pollinator-mediated evolution of floral signals. Trends Ecol. Evol. 28, 307–315. doi: 10.1016/j.tree.2013.01.019
Schlumpberger, B. O., and Raguso, R. A. (2008). Geographic variation in floral scent of Echinopsis ancistrophora (Cactaceae); evidence for constraints on hawkmoth attraction. Oikos 117, 801–814. doi: 10.1111/j.0030-1299.2008.16211.x
Sigsgaard, E. E., Olsen, K., Hansen, M. D. D., Hansen, O. L. P., Høye, T. T., Svenning, J.-C., et al. (2020). Environmental DNA metabarcoding of cow dung reveals taxonomic and functional diversity of invertebrate assemblages. Mol. Ecol. 30, 3374–3389. doi: 10.1111/mec.15734
Skubatz, H., Kunkel, D. D., Howald, W. N., Trenkle, R., and Mookherjee, B. (1996). The Sauromatum guttatum appendix as an osmophore: excretory pathways, composition of volatiles and attractiveness to insects. New Phytol. 134, 631–640. doi: 10.1111/j.1469-8137.1996.tb04928.x
Sládeček, F. X. J., Dötterl, S., Schäffler, I., Segar, S. T., and Konvicka, M. (2021). Succession of dung-inhabiting beetles and flies reflects the succession of dung-emitted volatile compounds. J. Chem. Ecol. 47, 433–443. doi: 10.1007/s10886-021-01266-x
Sletvold, N., and Ågren, J. (2014). There is more to pollinator-mediated selection than pollen limitation. Evolution 68, 1907–1918. doi: 10.1111/evo.12405
Stinchcombe, J. R., Agrawal, A. F., Hohenlohe, P. A., Arnold, S. J., and Blows, M. W. (2008). Estimating nonlinear selection gradients using quadratic regression coefficients: double or nothing? Evolution 62, 2435–2440. doi: 10.1111/j.1558-5646.2008.00449.x
Stökl, J., Schlüter, P. M., Stuessy, T. F., Paulus, H. F., Fraberger, R., Erdmann, D., et al. (2009). Speciation in sexually deceptive orchids: pollinator-driven selection maintains discrete odour phenotypes in hybridizing species. Biol. J. Linnean Soc. 98, 439–451. doi: 10.1111/j.1095-8312.2009.01279.x
Svensson, G. P., Pellmyr, O., and Raguso, R. A. (2006). Strong conservation of floral scent composition in two allopatric yuccas. J. Chem. Ecol. 32, 2657–2665. doi: 10.1007/s10886-006-9189-6
Szenteczki, M. A., Godschalx, A. L., Galmán, A., Espíndola, A., Gibernau, M., Alvarez, N., et al. (2021). Spatial and temporal heterogeneity in pollinator communities maintains within-species floral odour variation. Oikos. doi: 10.1111/oik.08445
Tremblay, R. L., Ackerman, J. D., Zimmerman, J. K., and Calvo, R. N. (2005). Variation in sexual reproduction in orchids and its evolutionary consequences: a spasmodic journey to diversification. Biol. J. Linnean Soc. 84, 1–54. doi: 10.1111/j.1095-8312.2004.00400.x
Wee, S. L., Tan, S. B., and Jürgens, A. (2018). Pollinator specialization in the enigmatic Rafflesia cantleyi: a true carrion flower with species-specific and sex-biased blow fly pollinators. Phytochemistry 153, 120–128. doi: 10.1016/j.phytochem.2018.06.005
Wheeler, G. S., Massey, L. M., and Southwell, I. A. (2002). Antipredator defense of biological control agent Oxyops vitiosa is mediated by plant volatiles sequestered from the host plant Melaleuca quinquenervia. J. Chem. Ecol. 28, 297–315. doi: 10.1023/A:1017982007812
Zou, H., and Hastie, T. (2005). Regularization and variable selection via the elastic net. J. R. Stat. Soc. Ser. B Stat. Methodol. 67, 301–320. doi: 10.1111/j.1467-9868.2005.00503.x
Keywords: Arum maculatum, brood-site deception, chemical ecology, geographic variation, hyperdiverse floral scents, phenotypic selection, Psychodidae
Citation: Gfrerer E, Laina D, Gibernau M, Fuchs R, Happ M, Tolasch T, Trutschnig W, Hörger AC, Comes HP and Dötterl S (2021) Floral Scents of a Deceptive Plant Are Hyperdiverse and Under Population-Specific Phenotypic Selection. Front. Plant Sci. 12:719092. doi: 10.3389/fpls.2021.719092
Received: 01 June 2021; Accepted: 12 August 2021;
Published: 24 September 2021.
Edited by:
Sergio Rossi, Université du Québec à Chicoutimi, CanadaReviewed by:
Gao Chen, Kunming Institute of Botany, Chinese Academy of Sciences, ChinaAphrodite Kantsa, ETH Zürich, Switzerland
Robert A. Raguso, Cornell University, United States
Copyright © 2021 Gfrerer, Laina, Gibernau, Fuchs, Happ, Tolasch, Trutschnig, Hörger, Comes and Dötterl. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stefan Dötterl, stefan.doetterl@plus.ac.at
†ORCID: Eva Gfrerer orcid.org/0000-0003-0837-1327
Danae Laina orcid.org/0000-0002-1058-5095
Marc Gibernau orcid.org/0000-0003-3866-3099
Roman Fuchs orcid.org/0000-0001-9423-5588
Martin Happ orcid.org/0000-0003-0009-2665
Wolfgang Trutschnig orcid.org/0000-0002-7131-1944
Anja C. Hörger orcid.org/0000-0001-6816-195X
Hans Peter Comes orcid.org/0000-0002-2659-8069
Stefan Dötterl orcid.org/0000-0001-5228-1332