 Zhonghua Tu
Zhonghua Tu Yufang Shen
Yufang Shen Shaoying Wen
Shaoying Wen Huanhuan Liu1,2
Huanhuan Liu1,2 Huogen Li
Huogen Li- 1Key Laboratory of Forest Genetics & Biotechnology of Ministry of Education, Nanjing Forestry University, Nanjing, China
- 2Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing, China
Liriodendron chinense is an economically and ecologically important deciduous tree species. Although the reference genome has been revealed, alternative polyadenylation (APA), transcription factors (TFs), long non-coding RNAs (lncRNAs), and co-expression networks of tissue-specific genes remain incompletely annotated. In this study, we used the bracts, petals, sepals, stamens, pistils, leaves, and shoot apex of L. chinense as materials for hybrid sequencing. On the one hand, we improved the annotation of the genome. We detected 13,139 novel genes, 7,527 lncRNAs, 1,791 TFs, and 6,721 genes with APA sites. On the other hand, we found that tissue-specific genes play a significant role in maintaining tissue characteristics. In total, 2,040 tissue-specific genes were identified, among which 9.2% of tissue-specific genes were affected by APA, and 1,809 tissue-specific genes were represented in seven specific co-expression modules. We also found that bract-specific hub genes were associated plant defense, leaf-specific hub genes were involved in energy metabolism. Moreover, we also found that a stamen-specific hub TF Lchi25777 may be involved in the determination of stamen identity, and a shoot-apex-specific hub TF Lchi05072 may participate in maintaining meristem characteristic. Our study provides a landscape of APA, lncRNAs, TFs, and tissue-specific gene co-expression networks in L. chinense that will improve genome annotation, strengthen our understanding of transcriptome complexity, and drive further research into the regulatory mechanisms of tissue-specific genes.
Introduction
Liriodendron chinense (L. chinense) is a deciduous tree species in the magnolia family (Magnoliaceae), which has desirable flowers and leaves with a unique shape. L. chinense is distributed throughout southern China and northern Vietnam and plays increasingly important economic and ecological roles because of its value in landscaping, furniture manufacturing and pharmaceutical manufacturing (Hao et al., 1995; Zhang et al., 2011; Xu et al., 2017). The whole-genome data of L. chinense have been released and contain 38 (2n = 38) chromosomes and 35,269 genes (Chen et al., 2018). However, the transcriptome of L. chinense has not been fully annotated, and information is lacking concerning alternative polyadenylation (APA), transcription factors (TFs), long non-coding RNAs (lncRNAs), and co-expression modules of tissue-specific genes.
Alternative polyadenylation is an important post-transcriptional regulatory mechanism that increases the complexity of the transcriptome and proteome (Sadek et al., 2019). APA causes a single gene to produce more than one transcriptome by changing the length of untranslated regions (UTRs) or coding regions (Zhang et al., 2020). APA in the UTR allows genes to produce different isoforms that have distinct UTR lengths but encode the same protein. Changes in the UTR length may affect mRNA stability, translation efficiency, or subcellular localization because UTRs usually harbor the target sites of microRNAs (miRNAs), RNA binding proteins, and other functional non-coding RNAs, and changes in the UTR length may lead to the gain or loss of these target sites (Chen et al., 2017b; Mayya and Duchaine, 2019; Zhang et al., 2020). These changes eventually affect the physiological and biochemical processes of plants. Ye et al. (2019) reported that genes with APA sites participate in cell wall formation and DNA repair under Cd stress. Cao et al. (2019) found that APA events affected gene expression and played roles in cell wall modification and root hair development. Yan et al. (2021) found that APA was involved in gene regulation in response to temperature stress. Although APA is a crucial post-transcriptional regulatory mechanism, it has not been unveiled in L. chinense.
lncRNAs are non-coding RNAs with lengths greater than 200 nucleotides. lncRNAs participate in transcription silencing and activation, chromosome modification, intranuclear transport, and protein modification by interacting with macromolecules (Quinn and Chang, 2016; Wu et al., 2020). Wang and Chang (2011) summarized four mechanisms of lncRNA regulation: signaling, decoying, guiding, and scaffolding. According to their location in the genome, lncRNAs can be divided into four categories: sense intronic lncRNAs, antisense lncRNAs, large intergenic lncRNAs (lincRNAs), and sense overlapping lncRNAs (Harrow et al., 2012; Wu et al., 2020). These lncRNAs participate in multiple biological processes, including plant growth and development, the stress response, disease resistance, the immune response, and vernalization response (Liu et al., 2020; Wu et al., 2020); for example, ASCO and COLDAIR participate in Arabidopsis thaliana root development and the vernalization response, respectively (Heo and Sung, 2011; Bardou et al., 2014), ALEX1 enhances rice (Oryza sativa) disease resistance (Yu et al., 2020), and PILNCR1 improves the tolerance of maize (Zea mays) under phosphate deficiency conditions (Du et al., 2018). In our previous study, we identified only 183 lncRNAs that had undergone alternative splicing (AS), and this value was far lower than the number of lncRNAs found in other species (Liu et al., 2018; Guo et al., 2020; Tu et al., 2020). Therefore, it is necessary to comprehensively study lncRNAs in L. chinense.
Tissue-specific genes can be regulated by APA and lncRNAs, which affect the biological traits and phenotypic characteristics of plants. Additionally, tissue-specific genes play fundamental roles in tissue differentiation and maintenance (Pattison et al., 2015). The divergence of tissue-specific gene expression patterns was found to modulate ion maintenance in tissues and improve salinity tolerance in Populus euphratica (Yu et al., 2017). In pineapple (Ananas comosus), 273 tissue-specific genes were detected, providing a basis to study the genetic mechanisms that regulate fruit phenotypes (Mao et al., 2018). Additionally, tissue-specific genes are involved in gibberellin metabolism in banana (Musa spp.) (Chen et al., 2016). Although studies of L. chinense tissues (leaves and petals) have been reported, the patterns of tissue-specific genes remain unknown, and this lack of knowledge impedes our understanding of tissue traits in this species (Yang et al., 2014; Ma et al., 2018).
Next-generation sequencing (NGS) is a high-throughput technique with high accuracy and a low cost and has been widely applied in plant transcriptome and genome research. However, it has weaknesses, such as difficulties in sequence assembly, the possibility of producing low-quality transcripts, and the inability to effectively capture full-length transcripts (Abdel-Ghany et al., 2016; Wang et al., 2016; Chen et al., 2017a; Wang et al., 2017; Chao et al., 2018, 2019; Zhang et al., 2019). Single-molecule long-read sequencing technology (SMRT) has been used in animals and plants to identify lncRNAs, comprehensively analyze APA, correct misannotated gene models and uncover novel genes (Xu et al., 2015; Abdel-Ghany et al., 2016; Chen et al., 2017a; Li et al., 2018; Liu et al., 2018; van Dijk et al., 2018; Wang et al., 2018a, b; Chao et al., 2019; Zhang et al., 2019). However, SMRT has the shortcomings of a high error rate (∼13%) and low throughput (van Dijk et al., 2018; Zhang et al., 2019). To overcome these obstacles, the use of hybrid sequencing (combining NGS with SMRT) to study the transcriptome has become an effective strategy that has been successfully applied to study various plants, such as moso bamboo (Phyllostachys edulis) (Zhao et al., 2018), sugarcane (Saccharum officinarum) (Hoang et al., 2017), and Medicago sativa (Chao et al., 2019). In our previous study, hybrid sequencing was applied to study AS, but these hybrid sequencing data could provide other important information in addition to AS, such as APA, TF families, the number of lncRNAs in the genome, and co-expression modules of tissue-specific genes.
Although the reference genome and AS in L. chinense have been reported, the annotation of APA, TFs and lncRNAs is incomplete, and the co-expression network of tissue-specific genes remains unknown. Therefore, to obtain information on APA, TFs, lncRNAs, and tissue-specific gene co-expression networks, we used hybrid sequencing data from bracts, sepals, petals, pistils, stamens, shoot apices, and leaves. Our study revealed the effects of APA on L. chinense, providing a landscape of lncRNAs and TFs, and uncovering the co-expression networks of tissue-specific genes. Additionally, we identified twenty hub tissue-specific genes, including two TFs, which are involved in different pathways, such as stamen development, shoot apical meristem function regulation, and plant defense. Taken together, this work will improve genome annotation and drive further research into tissue-specific functions in L. chinense.
Materials and Methods
Plant Materials
Twenty-six-year-old L. chinense trees located in a provenance trial plantation in Xiashu, Jurong County, Jiangsu Province (119°13′E, 32°7′N) were used as materials for sequencing. Plant samples were collected from the shoot apexes, expanded flower buds and mature leaves on April 15, 2019. The flower bud samples were divided into bracts, sepals, petals, stamens, and pistils. Each sample included three biological replicates. Twenty-one samples were frozen in liquid nitrogen and then stored at –80°C.
SMRT and Illumina Sequencing
Total RNA isolation, library construction, sequencing, and row data processing were fully described in our previous study (Tu et al., 2020). In total, 21 Illumina sequencing libraries and 1 mixed SMRT library were constructed, sequenced, and analyzed.
PacBio Iso-Seq Data Analysis
GMAP software was applied to align the polished consensus reads (long reads) to the reference genome (Wu and Watanabe, 2005). The polished reads were classified into five categories: (a) unmapped, (b) multiply mapped, (c) uniquely mapped, (d) mapped to ‘+’ sequences (sense sequences of the genome), and (e) mapped to ‘–’ sequences (antisense sequences of the genome). According to the mapping results, reads that were aligned to the unannotated regions of the reference genome were defined as novel genes. Seven databases (the NT, GO, Pfam, KEGG, NR, KOG, and Swiss-Prot databases) were used to annotate the functions of unmapped transcripts and novel genes (Kanehisa and Goto, 1999; Tatusov et al., 2003; Pruitt et al., 2007; Young et al., 2010; Finn et al., 2016; The UniProt, 2017).
Four additional tools (PLEK, CNCI, CPC, and the Pfam database) were used to predict lncRNAs, all with default parameters (Kong et al., 2007; Sun et al., 2013; Li et al., 2014; Finn et al., 2016). Transcripts with no potential coding sequence were retained as our set of candidate lncRNAs.
We used the TAPIS pipeline to predict APA sites (Abdel-Ghany et al., 2016) and MEME to analyze the nucleotide composition of the sequences upstream (–50 nt) and downstream (+50 nt) of all APA sites for nucleotide bias (Alamancos et al., 2015).
TFs were identified using iTAK software with the default parameters (Zheng et al., 2016), and expression trend analysis was performed using the OmicStudio tools (STEM package) at https://www.omicstudio.cn/tool.
Identification of Differentially Expressed Genes and Tissue-Specific Genes and GO/KEGG Enrichment Analyses
The number of reads that mapped to each gene was calculated by Cuffdiff (Sreya and Chon-Kit Kenneth, 2016). The fragments per kilobase of transcript sequence per million mapped reads (FPKM) value of each gene was calculated based on the read count mapped to the gene and length of the gene.
DEseq2 software1 was used for differential expression analysis. Genes with adjusted p-values that were less than 0.05 and | log2fold change| ≥ 1 were defined as differentially expressed genes (DEGs). Based on the differential expression analysis results, tissue-specific genes were determined according to the following conditions: the FPKM value must be greater than 1 in a specific tissue, and the expression level of tissue-specific genes must be at least fourfold higher than that in other tissues (Mortazavi et al., 2008; Wang et al., 2019). GO enrichment analysis was then performed using GOseq software2. KOBAS software3 was used to perform KEGG pathway enrichment analysis.
Gene Co-expression Network Analysis and Identification of Hub Genes
Gene co-expression analysis was performed via weighted correlation network analysis (WGCNA) in R software (Langfelder and Horvath, 2008). Of the 21,944 DEGs between the seven tissues, we selected genes that ranked in the top 75% according to the ranking of the FPKM variance values. The most suitable power was then determined (14 in this study) according to the calculation results of the soft threshold power using the pickSoftThreshold function (the soft threshold power ranged from 1 to 30). Next, the blockwiseModules function was applied to construct gene co-expression networks using the following parameters: a power of 14, an unsigned TOM type, a min module size of 200, a mergeCutHeight of 0.2, and default values of the other parameters. Based on the WGCNA results, we filtered edges with weight values less than 0.3. The degree, MCC, EPC, closeness, and radiality algorithms of the CytoHubba package in Cytoscape software4 were then used to select hub genes; genes that ranked in the top 2% by all algorithms were identified as hub genes (Smoot et al., 2011). The gene co-expression networks were visualized using Gephi software5.
Validation of Hybrid Sequencing
RT-qPCR was used to validate the expression levels of five tissue-specific genes and 16 DEGs. In each sample, 1 μg of total RNA was used to synthesize cDNA using PrimeScriptTM RT Master Mix (TaKaRa, Dalian, China). The Oligo 7 algorithm6 was used to design RT-qPCR primers for the 16 selected DEGs (Supplementary Table 1). RT-qPCR was performed using a StepOnePlusTM system (Applied Biosystems) with SYBR® Premix Ex TaqTM (TaKaRa, Dalian, China), as directed by the instructions. Eukaryotic translation initiation factor 3 (eIF3) was designated as an internal control gene (Tu et al., 2019).
Results
Processing of the PacBio Sequencing Data
Using PacBio SMRT, we obtained 618,876 polymerase reads from 21 pooled samples (Supplementary Table 2). From the polymerase reads, 10,437,029 subreads were extracted, with an average length of 2,177 bp. After further processing, 498,059 circular consensus sequences (CCSs) were obtained, each containing a poly(A) tail and 5′ and 3′ adaptors, with an average length of 2,755 bp. Among the CCSs, we detected 430,554 full-length non-chimeric (FLNC) reads, with an average length of 2,568 bp. The reads were polished with both iterative clustering for error correction (ICE), resulting in 227,276 polished consensus reads whose average length was 2,697 bp. By mapping the short reads generated from Illumina sequencing via LoRDEC, we corrected up to 100% of the sequencing errors (Salmela and Rivals, 2014). We ultimately obtained 227,276 high-quality polished consensus reads with an average length of 2,696 bp for further analysis (Supplementary Table 3).
GMAP software was used to align the high-quality polished consensus reads to the reference genome, and 1.13% (2,572/227,276) of the reads were found to be unmapped (Wu and Watanabe, 2005). Among the remaining 224,704 alignments, 9.96% (22,628/227,276) were mapped to multiple positions, and 52.73% (119,840/227,276) and 36.18% (82,236/227,276) were mapped to the sense sequences and antisense sequences in the genome, respectively (Figure 1A). After removing consensus reads with incomplete 5′ and 3′ ends and reads from fusion genes, 144,006 complete consensus reads could be mapped to the reference gene model (Supplementary Table 4).
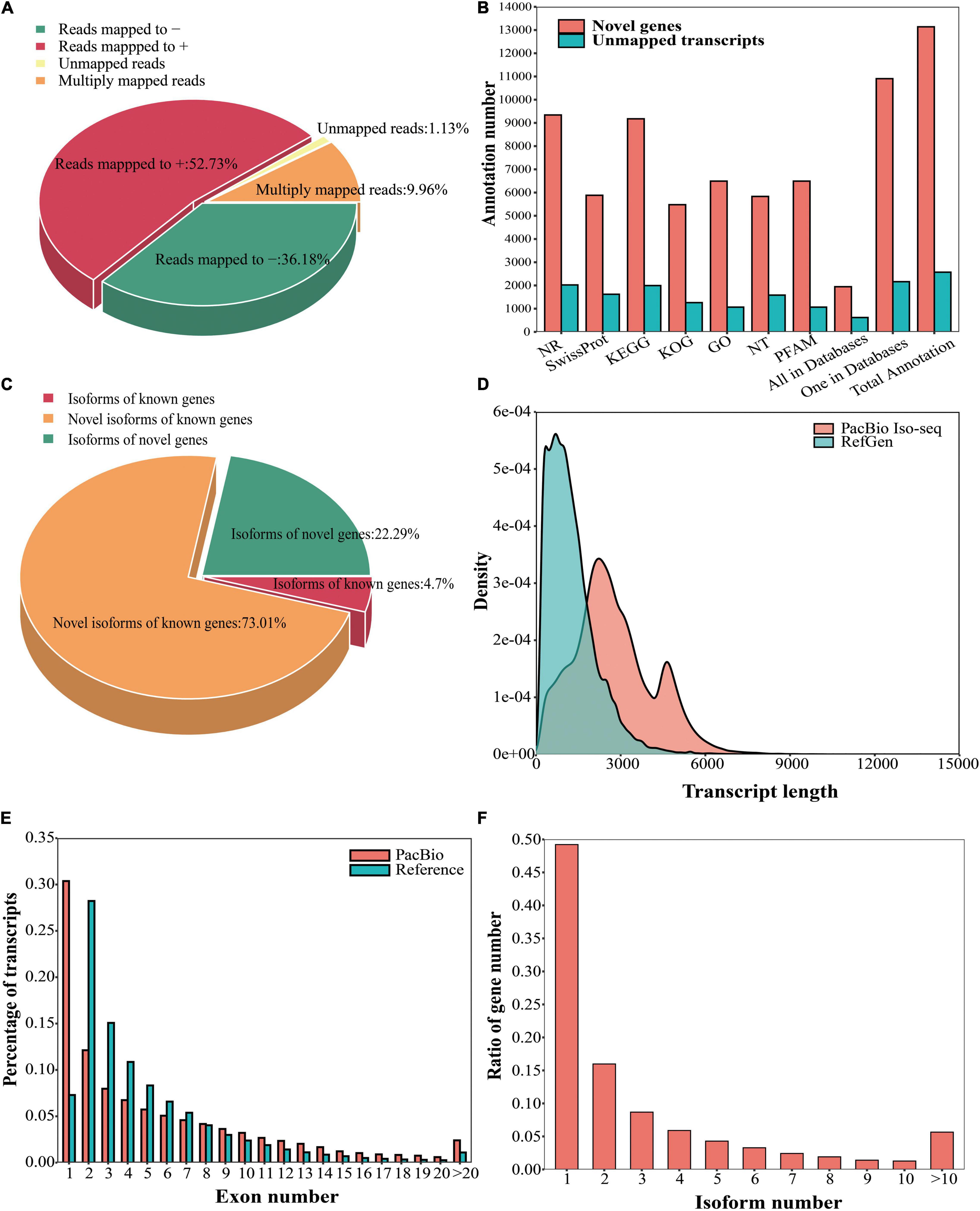
Figure 1. Analysis of the Iso-Seq isoforms. (A) GMAP mapping results (‘+’ represents the sense sequence of the genome, and ‘–’ represents the antisense sequence of the genome). (B) Functional annotation of the unmapped transcripts and novel genes in seven databases. (C) Classification of transcripts based on the reference genome. (D) Comparison of the isoform lengths between the PacBio data and reference genome annotation. (E) Comparison of the number of exons in the transcripts between the PacBio data and reference genome annotation. (F) Number of transcripts produced by each gene according to PacBio Iso-Seq.
In the PacBio sequencing data set, 2,572 unique clusters did not align to the reference genome. These unmapped transcripts were then mapped to 7 databases: NR, NT, Pfam, KOG, Swiss-Prot, KEGG, and GO. In these databases, 2,022 (NR), 1,584 (NT), 1,066 (Pfam), 1,260 (KOG), 1,621 (Swiss-Prot), 1,995 (KEGG), and 1,066 (GO) unmapped transcripts were annotated (Figure 1B). Among these unmapped transcripts, 619 were found in all seven databases, and 2,163 were annotated in only one database (Figure 1B).
We defined reads that mapped to unannotated regions of the reference genome as reads from novel genes, and 13,139 novel genes covered by 31,792 complete consensus reads were detected (Supplementary Table 3). To better understand the novel genes, they were functionally annotated. In total, 9,343 (NR), 5,833 (NT), 6,494 (Pfam), 5,478 (KOG), 5,883 (Swiss-Prot), 9,177 (KEGG), and 6,494 (GO) novel genes were annotated in the seven databases (Figure 1B). Additionally, 1,945 novel genes were annotated across all seven databases, and 10,905 novel genes were annotated in only one database (Figure 1B).
Isoform Detection and Characterization
TAPIS software was used to classify and characterize the full-length isoforms (Abdel-Ghany et al., 2016). A total of 14,488 known genes covered by 69,658 isoforms, including 4,210 (4.70%) isoforms of known genes, 65,448 (73.01%) novel isoforms of known genes, and 13,139 novel genes covered by 19,982 (22.29%) isoforms (Figure 1C). We compared the lengths of the transcripts between the PacBio data and reference genome annotation and found that the transcripts described in the reference genome annotation were shorter than those detected by PacBio Iso-Seq (Figure 1D). Additionally, we compared the numbers of exons in the transcripts between the PacBio data and reference genome annotation. Single-exon genes represented a large proportion (30.37%) of the PacBio data; however, in the reference genome, single-exon genes accounted for only 7.29% (Figure 1E). The proportion of multiple-exon genes (≥2 exons) in the reference genome annotation was greater than that in the PacBio data (Figure 1E). Furthermore, single-isoform genes constituted a large proportion of the PacBio data (49.22%), followed by two-isoform genes, which accounted for 15.98% (Figure 1F).
Comparison of lncRNAs and mRNAs
lncRNAs are RNA molecules that are longer than 200 nt that do not encode proteins. We used four tools, PLEK, CNCI, CPC, and Pfam, to identify lncRNAs in the PacBio Iso-Seq data (Kong et al., 2007; Sun et al., 2013; Li et al., 2014; Finn et al., 2016). The PLEK, CNCI, CPC, and Pfam tools predicted 44,108 lncRNAs, 15,500 lncRNAs, 21,549 lncRNAs, and 43,103 lncRNAs, respectively (Figure 2A). To improve the accuracy of the lncRNA prediction, each lncRNA was predicted by four tools, and 7,527 lncRNAs were detected by all four tools (Figure 2A). Most of the lncRNAs were less than 1,000 bp (Figure 2B). Among these, lincRNAs accounted for the largest proportion (36.44%), followed by sense overlapping lncRNAs (28.19%), sense intronic lncRNAs (18.37%), and antisense lncRNAs (16.99%) (Figure 2C; Harrow et al., 2012). Compared with mRNAs, lncRNAs generally had fewer exons and were shorter, and single-exon lncRNAs accounted for most of the lncRNAs (72.34%) (Figures 2D,E).
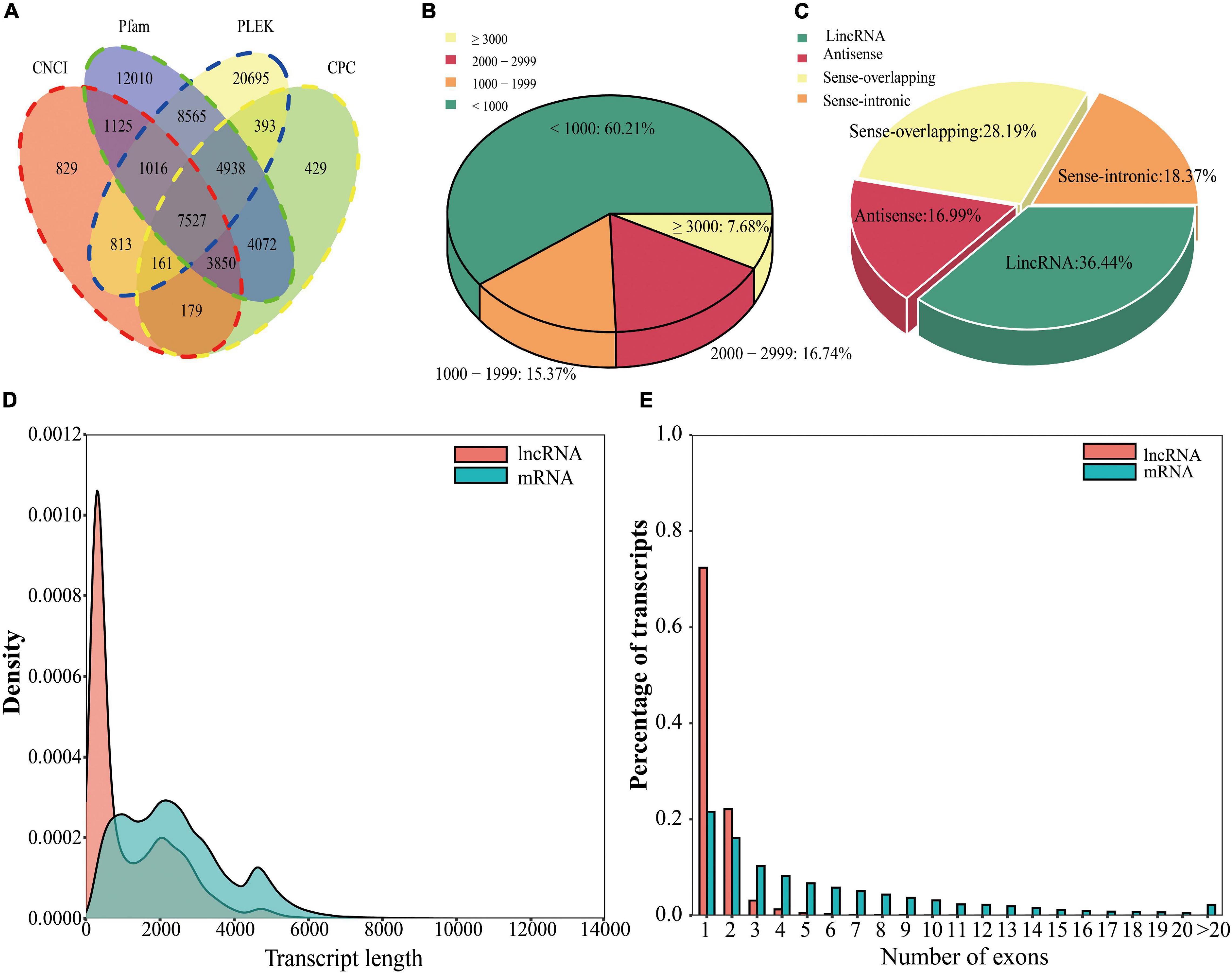
Figure 2. Identification, classification, and analysis of lncRNAs in L. chinense. (A) Venn diagram of lncRNAs predicted by the CNCI, Pfam, PLEK, and CPC tools. (B) Percentage of lncRNAs with different length. (C) Classification of lncRNAs. (D) Comparison of the lengths of mRNAs and lncRNAs. (E) Comparison of the numbers of exons of mRNAs and lncRNAs.
Identification of DEGs and Tissue-Specific Genes
To investigate gene expression patterns in the seven evaluated tissues of L. chinense, we used FPKM values to normalize the reads from Illumina sequencing. We identified 3,720 DEGs between floral tissues (bracts, sepals, petals, stamens, and pistils) and vegetative tissues (leaves and the shoot apex); 1,293 of these DEGs were upregulated, and the other 2,427 were downregulated (Figure 3A). The Euclidean distance method was used to perform clustering analysis of all the genes to identify their clustering patterns in the different tissues (Yang et al., 2018). Hierarchical clustering analysis showed that these DEGs were clustered into three subclusters, one of which was highly expressed in leaves, one in the shoot apex and the other in floral tissues (Figure 3C).
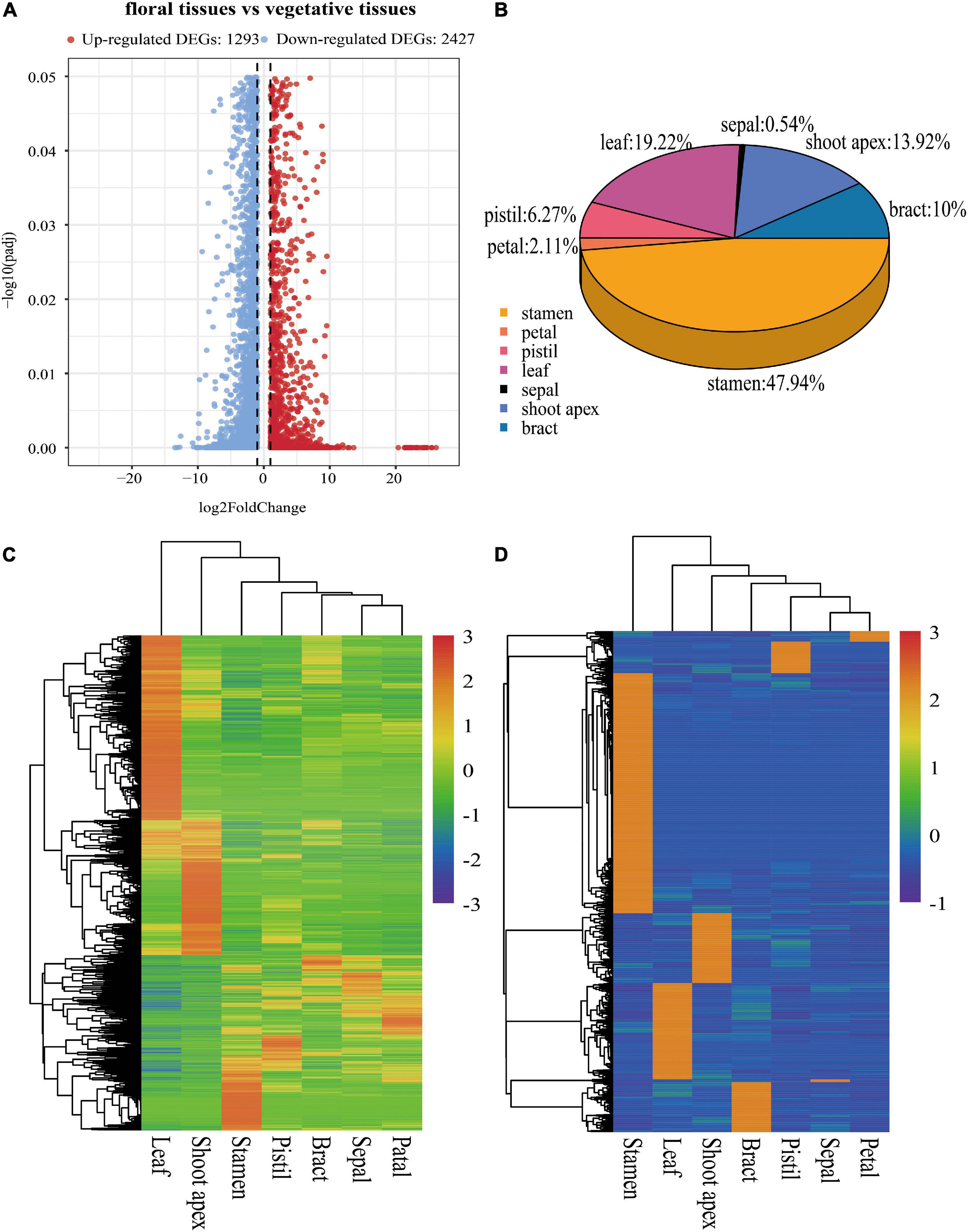
Figure 3. Information about DEGs and tissue-specific genes. (A) Volcano plot of DEGs between floral tissues and vegetative tissues. (B) Distribution of tissue-specific genes in seven tissues. (C) Hierarchical clustering analysis of DEGs based on the normalized log10(FPKM) values. (D) Hierarchical clustering analysis of tissue-specific genes.
To better understand tissue-specific genes in L. chinense, we investigated their distribution and pathways with which these genes were associated. Overall, 2,040 tissue-specific genes in seven tissues were detected. Stamen tissue contained the highest number of tissue-specific genes (978, 47.94%), followed by the leaf (392, 19.22%), shoot apex (284, 13.92%), bract (204, 10.00%), pistil (128, 6.27%), and petal (43, 2.11%) tissues, while the sepal tissue contained the least number of tissue-specific genes (11, 0.54%) (Figure 3B). Hierarchical clustering analysis showed that these tissue-specific genes were obviously clustered into seven subclusters (Figure 3D). GO enrichment analysis results showed that 29 GO terms were significantly enriched, and the largest number of tissue-specific genes were significantly enriched in catalytic activity, followed by transferase activity and hydrolase activity (Supplementary Figure 1A). KEGG pathway enrichment analysis revealed that these tissue-specific genes were significantly enriched in nine pathways, and the largest number of tissue-specific genes were significantly enriched in the biosynthesis of secondary metabolites pathway (Supplementary Figure 1B).
Identification and Analysis of TFs in L. chinense
We identified 1,791 TFs from 94 families, among which the bHLH family had the largest number of TFs (106, 5.92%), followed by MYB (98, 5.47%), and NAC (88, 4.91%) (Figure 4A). These 1,791 TFs showed the highest expression level in the shoot apex and the lowest expression level in petals (Figure 4B). Expression trend analysis identified only six significant expression patterns, including 613 TFs (Figure 4C). Moreover, we found that 244 TFs were differentially expressed between floral tissues and vegetative tissues, and hierarchical clustering analysis showed that these 244 TFs could be clustered into two subclusters (vegetative tissue subcluster and floral tissue subcluster) (Figure 4D). These differentially expressed TFs were significantly enriched in plant hormone signal transduction and circadian rhythm pathways (Figure 4E).

Figure 4. Analysis of TFs in L. chinense. (A) Proportion of different TF families (only the top 9 TF families are shown). (B) Expression trend analysis of TFs. (C) Expression levels of TFs in seven tissues; the red rectangles represent the average values. (D) Hierarchical clustering analysis of differentially expressed TFs. (E) KEGG pathway enrichment analysis of differentially expressed TFs.
Polyadenylation Analysis and Identification of Alternatively Spliced Tissue-Specific Genes
Most genes in eukaryotes can generate various mRNA 3′ ends through APA, markedly increasing the complexity of the transcriptome (Shen et al., 2011; Wu et al., 2011). Using the TAPIS pipeline (Abdel-Ghany et al., 2016), we identified 11,108 annotated genes containing at least one poly(A) site [poly(A) gene], among which 4,387 annotated genes comprised a single poly(A) site and 6,721 annotated genes contained more than one poly(A) site (Figure 5A). Poly(A) genes showed more exons, shorter mean exon lengths, and longer mean intron lengths than genes without poly(A) sites [non-poly(A) genes] (Figures 5B–D). To identify the potential cis-elements involved in polyadenylation, motif enrichment analysis was performed to analyze the 50 nucleotides upstream from the poly(A) sites (Abdel-Ghany et al., 2016). We discovered two conserved motifs (AUAAA and UGUA) upstream of the poly(A) cleavage site (Figure 5E); these motifs were also present in maize, red clover (Trifolium pratense L.), sorghum (Sorghum bicolor), Brassica napus, and poplar (Populus alba var. pyramidalis) (Abdel-Ghany et al., 2016; Wang et al., 2016; Chao et al., 2018; Hu et al., 2020; Yao et al., 2020). Additionally, to investigate the preferential nucleotides at the poly(A) cleavage sites, we analyzed the nucleotide composition of 50 downstream and 50 upstream nucleotides at all APA cleavage sites. Uracil (U) was found to be enriched upstream of the APA cleavage sites, while adenine (A) was enriched downstream of the cleavage sites (Figure 5F). The same findings were reported in red clover and sorghum (Abdel-Ghany et al., 2016; Chao et al., 2018). Additionally, we found that 990 APA genes were differentially expressed between floral tissues and vegetative tissues, among which 394 upregulated APA genes were significantly enriched in glycerophospholipid metabolism, cyanoamino acid metabolism, and ether lipid metabolism pathways, and 596 downregulated APA genes were significantly enriched in glycine, serine and threonine metabolism, circadian rhythm, and glyoxylate and dicarboxylate metabolism pathways (Figures 6A,B).
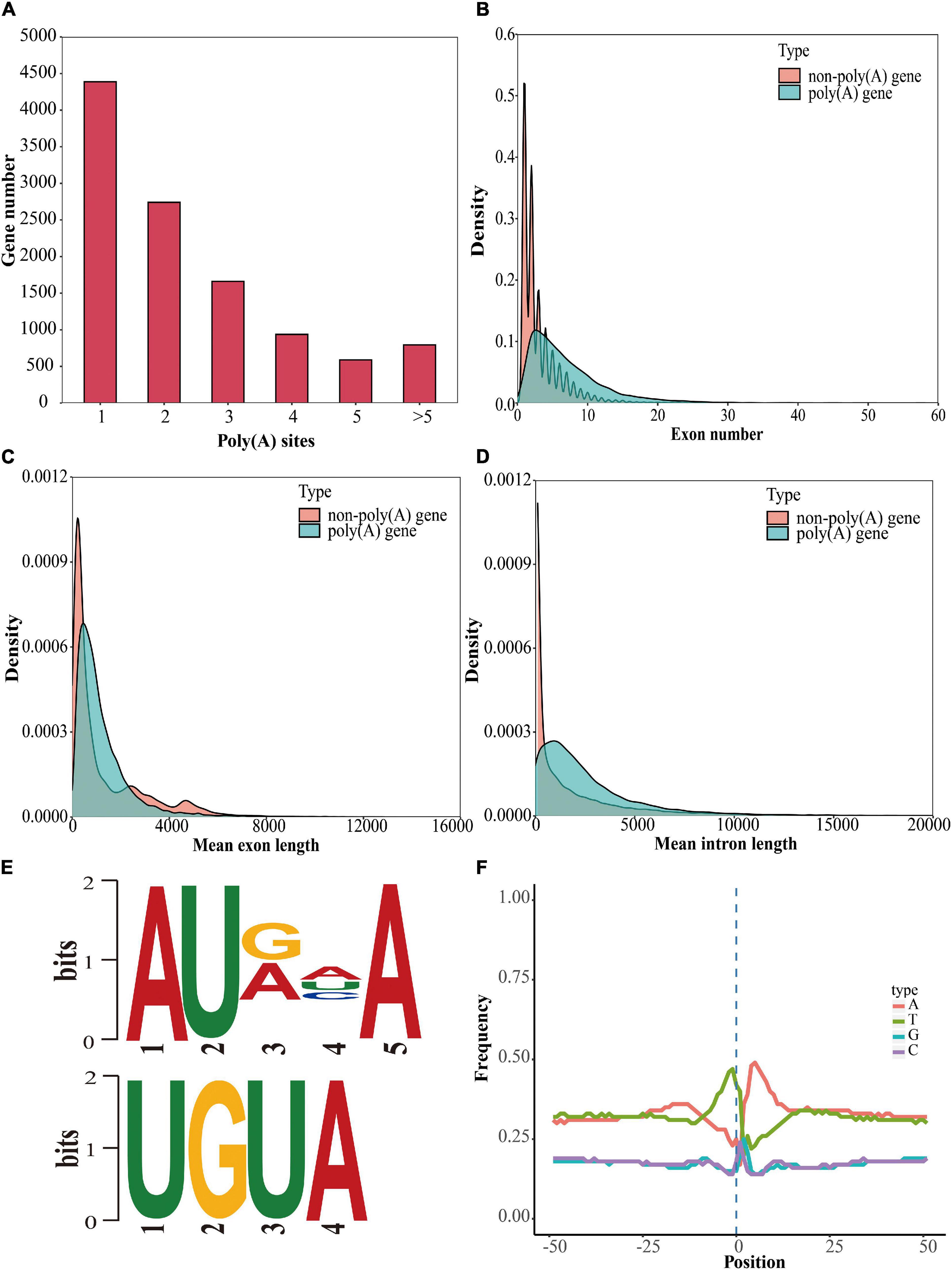
Figure 5. Identification and analysis of genes with poly(A) sites. (A) Analysis of the gene numbers according to the number of poly(A) sites. (B) Comparison of the exon numbers of poly(A) genes and non-poly(A) genes. (C) Comparison of the mean exon lengths of poly(A) genes and non-poly(A) genes. (D) Comparison of the mean intron lengths of poly(A) genes and non-poly(A) genes. (E) Motif analysis around poly(A) cleavage sites. (F) Nucleotide distribution around poly(A) cleavage sites.
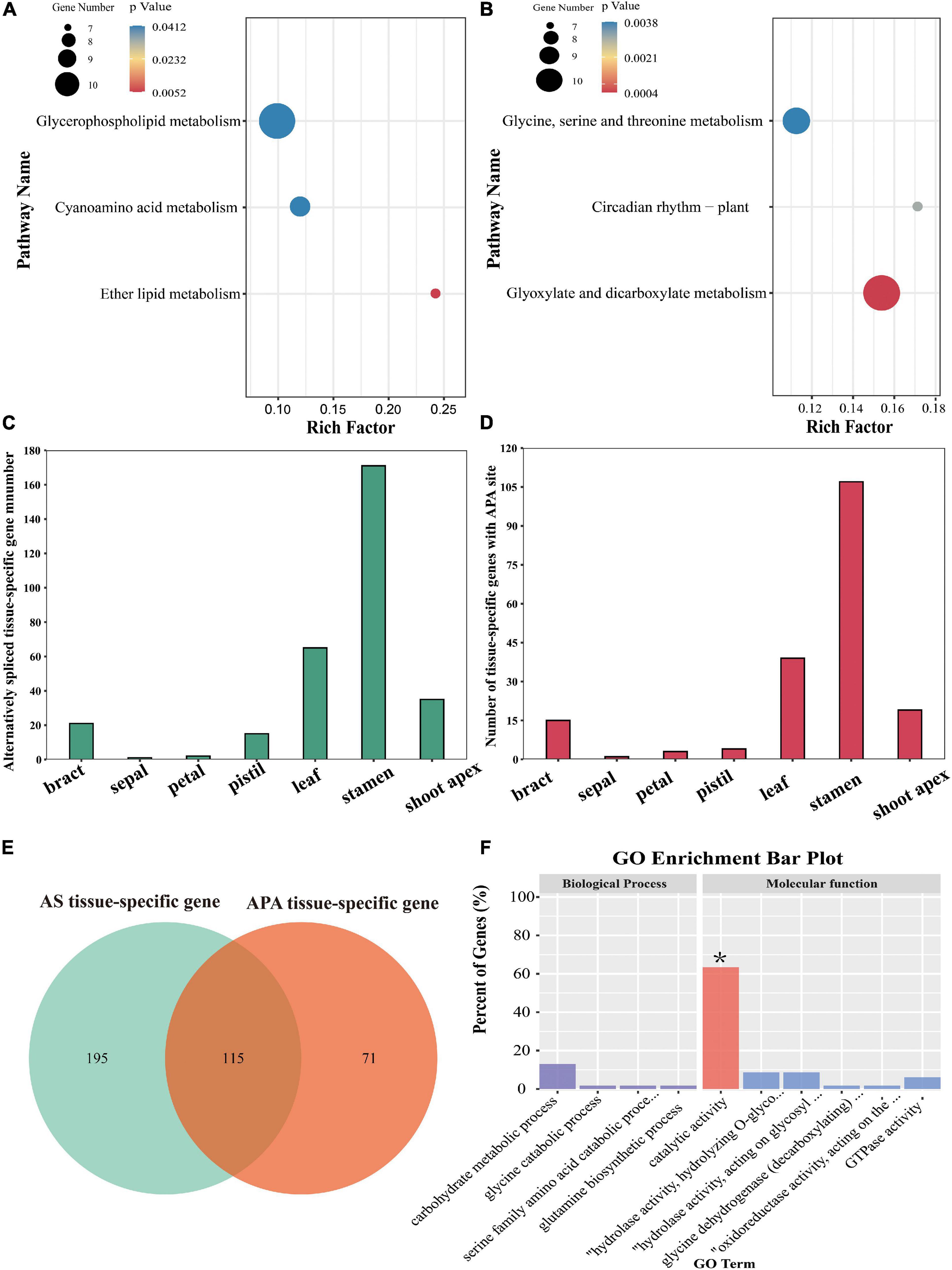
Figure 6. KEGG pathway enrichment analysis of the DEGs with APA sites and analysis of the tissue-specific genes affected by AS or APA. (A,B) KEGG pathway enrichment analysis of up-regulated and down-regulated genes affected by APA, respectively. (C,D) Numbers of tissue-specific genes affected by AS and APA, respectively. (E) Venn plot of AS tissue-specific genes and APA tissue-specific genes. (F) GO enrichment analyses of 115 genes affected by both AS and APA.
In our previous study, we identified 8,503 AS genes (Tu et al., 2020). Actually, 310 tissue-specific genes had undergone AS; stamen-specific genes accounted for most of these (171), followed by leaf-specific genes (65), shoot-apex-specific genes (35), bract-specific genes (21), pistil-specific genes (15), petal-specific genes (2), and a sepal-specific gene (1) (Figure 6C). Moreover, 187 tissue-specific genes were identified to have APA sites, including 107 stamen-specific genes, 39 leaf-specific genes, 18 shoot-apex-specific genes, 15 bract-specific genes, 4 pistil-specific genes, 3 petal-specific genes, and 1 sepal-specific gene (Figure 6D). We found that 115 tissue-specific genes were affected by both AS and APA, among which 73 genes were significantly enriched in catalytic activity GO (Figures 6E,F).
WGCNA Analyses of DEGs and Tissue-Specific Genes
Weighted correlation network analysis was used to analyze gene co-expression (Langfelder and Horvath, 2008). In total, 21,944 DEGs between seven tissues were identified, and only the DEGs ranked in the top 75% based on the ranking of the variance of FPKM values could be used for WGCNA. First, we identified 22 gene co-expression modules (genes that could not be clustered because of their large difference from other genes were divided into a gray module) (Figure 7A). However, some modules were highly correlated. Thus, we merged the modules with correlations higher than 0.8 (mergeCutHeight = 0.2). Finally, 16 co-expression modules were identified. The number of genes in each co-expression module ranged from 3 (gray module) to 2,946 (blue module) (Figure 7C). In total, 1,809 tissue-specific genes were classified into seven co-expression modules as follows: the brown module contained 343 leaf-specific genes, the green module contained 36 petal-specific genes, the midnight-blue module contained 10 sepal-specific genes, the red module contained 167 bract-specific genes, the salmon module contained 117 pistil-specific genes, the turquoise module contained 884 stamen-specific genes, and the yellow module contained 252 shoot-apex-specific genes (Figure 7D).
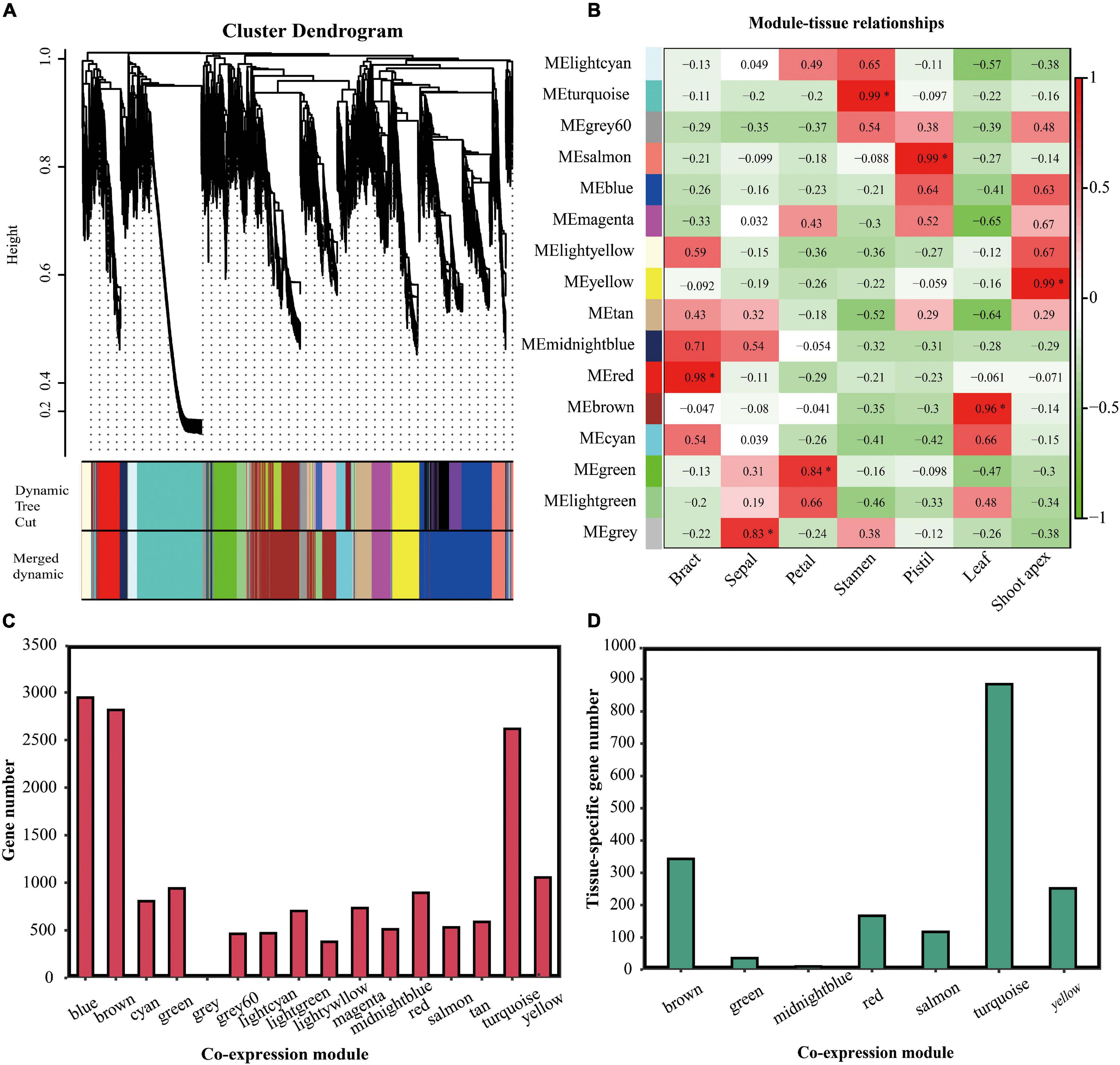
Figure 7. Analysis of the gene co-expression modules. (A) Dendrogram of the gene co-expression modules in L. chinense. The genes are represented by vertical lines in the upper panel. In the bottom panel, rectangles of thirteen colors represent the 16 gene co-expression modules. (B) Relationships between tissues and gene co-expression modules. The asterisk indicates that the co-expression module is significantly correlated to the tissue. (C) Analysis of the gene numbers in the 16 co-expression modules. (D) Analysis of the tissue-specific genes in the 16 co-expression modules.
Because tissue-specific genes tended to appear in specific modules, we investigated whether there were specific co-expression modules that were strongly related to specific tissues. We analyzed the correlation between the 16 co-expression modules and 7 tissues. We found that the red, green, turquoise, salmon, brown, and yellow modules were significantly correlated with bract, petal, stamen, pistil, leaf, and shoot apex tissue, respectively (P < 0.05) (Figure 7B). Although other modules had relationships with certain tissues, these correlations were not significant.
Hierarchical clustering analysis showed that these 1,809 tissue-specific genes represented obvious specificity (Supplementary Figure 2). Next, we performed KEGG pathway enrichment analyses for these 1,809 tissue-specific genes in specific co-expression modules. Tissue-specific genes in specific modules were significantly enriched in the biosynthesis of secondary metabolite pathways, except for bract-specific genes in the red module and petal-specific genes in the green module (Supplementary Figure 3). Shoot-apex-specific genes, leaf-specific genes, pistil-specific genes, and stamen-specific genes were also significantly enriched in the phenylpropanoid biosynthesis pathway (Supplementary Figures 3D–G).
Identification of Hub Genes in Specific Co-expression Modules
As mentioned above, most of the tissue-specific genes were specifically expressed in seven co-expression modules: red module (bract-specific genes), brown module (leaf-specific genes), green module (petal-specific genes), salmon module (pistil-specific genes), midnight blue module (sepal-specific genes), yellow module (shoot-apex-specific genes), and turquoise module (stamen-specific genes). Therefore, we analyzed the co-expression networks of these tissue-specific genes in these specific modules and identified hub genes in the co-expression networks (Supplementary Figure 4). We identified 20 hub genes from seven specific modules that included two TFs, Lchi05072 [AINTEGUMENTA-LIKE 5 (AIL5), shoot-apex-specific] and Lchi25777 (SPL/NZZ, stamen-specific) (Supplementary Table 5). Lchi05072 was co-expressed with 672 genes, and Lchi25777 was co-expressed with 578 genes (Supplementary Figure 4). We performed KEGG and GO enrichment analyses for them. Genes co-expressed with Lchi05072 were significantly enriched in nine pathways, including the biosynthesis of secondary metabolites pathway and plant hormone signal transduction pathway, and four GO terms, including terpene synthase activity (Figures 8A,C). Genes co-expressed with Lchi25777 were significantly enriched in three pathways, one of which was phenylpropanoid biosynthesis; however, they were not significantly enriched in any GO terms (Figures 8B,D). Additionally, we found that most of the bract-specific hub genes were associated with plant resistance, such as Lchi00262 (pathogenesis-related protein, PR), Lchi04173 (L-type lectin receptor-like kinase, LecRLK), and Lchi05264 (leucine-rich repeat receptor-like protein kinase, LRR-RLK). Additionally, we found two leaf-specific hub genes, Lchi03373 (glyceraldehyde-3-phosphate dehydrogenase, GAPDH) and Lchi05497 (ribulose bisphosphate carboxylase/oxygenase, Rubisco), which were involved in energy metabolism (Supplementary Table 5). These hub tissue-specific genes have important research significance.
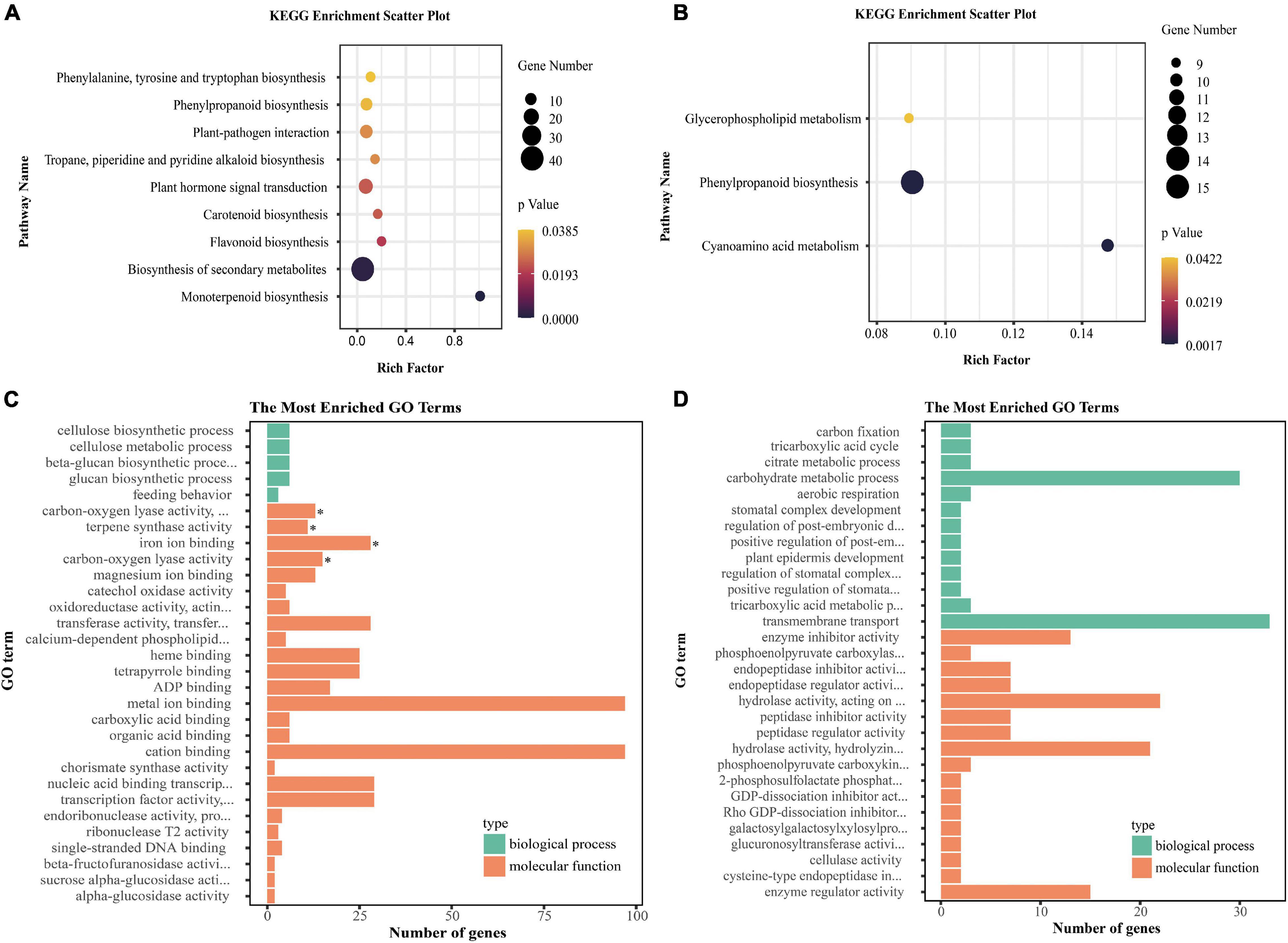
Figure 8. KEGG and GO enrichment analysis of genes co-expressed with two hub TFs. (A,B) KEGG pathway enrichment analysis of genes co-expressed with Lchi05072 and Lchi25777, respectively (only significantly enriched pathways are shown). (C,D) GO enrichment analysis of genes co-expressed with Lchi05072 and Lchi25777, respectively (only the top 30 terms according to P-values are shown). The asterisks represent significantly enriched terms.
RT-qPCR Validation for RNA Sequencing Data
To validate the accuracy of the gene expression levels detected by RNA sequencing, we performed RT-qPCR of 16 randomly selected DEGs. The expression levels of the 16 DEGs measured by Illumina sequencing were consistent with the RT-qPCR analysis results (Figures 9A–P). Correlation analysis revealed that the RNA sequencing data were highly correlated with the RT-qPCR analysis results (R2 = 0.947; P < 0.01) (Figure 9Q). These results indicate that the gene expression levels detected by Illumina sequencing are reliable.

Figure 9. RT-qPCR validation of RNA sequencing. BR, bract; LE, leaf; PE, petal; PI, pistil; SA, shoot apex; SE, sepal; ST, stamen. (A–P) Relative expression analysis and FPKM values of 16 DEGs in different tissues. (Q) Correlation analysis of RT-qPCR data and RNA sequencing data.
Discussion
Liriodendron chinense, an economically and ecologically important tree species, has received consistent research attention. Since the genome of L. chinense was released, the understanding of its evolutionary history has improved (Chen et al., 2018). However, the reference genome has not been fully annotated, and critical information, such as AS, APA, TFs, and lncRNAs, remains to be uncovered. In our previous study, we used hybrid sequencing to reveal the effect of AS in L. chinense, but this approach did not fully exploit these data to obtain more useful information, such as APA, lncRNAs, TFs, and tissue-specific genes. Thus, we conducted in-depth research and analysis of the L. chinense full-length transcriptome from different perspectives.
lncRNAs are essential regulators involved in plant growth and development, the stress response, disease resistance, the immune response, and the vernalization response (Wu et al., 2020). In our previous study, we identified only 183 lncRNAs affected by AS (Tu et al., 2020). However, in other studies, the number of lncRNAs was much more than 183. For example, Liu et al. (2018, 2019) detected 5,952 and 13,099 lncRNAs in poplar, Guo et al. (2020) identified 4,460 lncRNAs, and Tian et al. (2016) found 7,655 lncRNAs; thus, we speculated that 183 AS lncRNAs represented only a proportion of lncRNAs in L. chinense. Through in-depth analysis, we detected 7,527 lncRNAs, much more than the 183 AS lncRNAs. Compared with protein-coding genes, the effect of AS in lncRNAs was much weaker (Tu et al., 2020). Among these lncRNAs, lincRNA accounted for the largest proportion, a finding that was consistent with reports in poplar and hickory (Carya cathayensis) (Liu et al., 2018; Fan et al., 2020; Guo et al., 2020). Studies have revealed that lincRNAs are involved in transcriptional regulation and signal transduction in soybean (Glycine max) and regulate root development in A. thaliana (Li et al., 2016; Golicz et al., 2018). These L. chinense lincRNAs have potential research significance. Additionally, we found that the lncRNAs were shorter and smaller than the mRNAs in terms of the exon length and number. This phenomenon was also observed in hickory, rice, and poplar (Liu et al., 2018; Fan et al., 2020; Zhao et al., 2020). These findings provide a genome-wide landscape of lncRNAs in L. chinense.
Alternative polyadenylation is a critical post-transcriptional regulatory mechanism involved in maintaining RNA stability, ensuring accurate RNA localization and translation; these stabilities are crucial in plant development and flowering (Liu et al., 2010; Shen et al., 2011; Zhang et al., 2018). In A. thaliana, approximately 60% of the genes have multiple poly(A) sites (Shen et al., 2011). In L. chinense, 13.8% (6,721/48,408) of the genes had APA sites. The number and source of sequencing samples, sequencing depth, and analysis methods strongly affect the APA analysis results; thus, the proportion of detected APA genes in L. chinense is lower than that in other studies, likely because of the above reasons (Shen et al., 2011; Wu et al., 2011; Ruan et al., 2018). However, 26.6% of DEGs were affected by APA, indicating that APA may participate in gene expression regulation. Additionally, we found that the distribution of nucleotides upstream and downstream of the APA cleavage sites was consistent with that in previous reports of poly(A) analyses in sorghum and red clover (Abdel-Ghany et al., 2016; Chao et al., 2018). Furthermore, two motifs (AUAAA and UGUA) upstream of the poly(A) cleavage sites were detected. AUAAA is a canonical cis element that plays a role as a polyadenylation signal (PAS) for 3′ end processing and has been reported in red clover, sorghum and other plant species (Abdel-Ghany et al., 2016; Wang et al., 2016; Chao et al., 2018; Turner et al., 2018). The UGUA motif is a U-rich element (USE) that cooperates with auxiliary elements to affect 3′ end processing (Turner et al., 2018). In the absence of PAS, the cleavage factors bind to the 3′ end processing machinery with the help of USE or auxiliary elements (Venkataraman et al., 2005). These findings demonstrate that APA contributes substantially to the complexity and flexibility of the L. chinense transcriptome.
As a special category of DEGs, tissue-specific genes play important roles in plant defense, stress responses, plant development and material metabolism (Pattison et al., 2015; Celedon et al., 2017; Villarino et al., 2017; Yu et al., 2017; Alonso-Serra et al., 2019; Amini et al., 2019). In Ferula asafetida, genes involved in terpenoid and phenylpropanoid metabolism are expressed in flowers (Amini et al., 2019). In tomato (Solanum pennellii), tissue-specific genes play important regulatory roles during fruit development (Pattison et al., 2015). In the present study, we identified 2,040 tissue-specific genes in L. chinense. Enrichment analysis revealed that tissue-specific genes participated in multiple metabolism and synthesis pathways, such as starch and sucrose metabolism, phenylpropanoid biosynthesis, and flavonoid biosynthesis. Previous studies have found that stamen development is accompanied by starch accumulation and degradation, and phenylpropanoid biosynthesis is essential for pollen development (Julian et al., 2011; Xue et al., 2020). Flavonoids are important products of secondary metabolism and participate in multiple functions in plants, such as developmental regulation, flower pigmentation, and stress responses (Mathesius, 2018). Moreover, most of the tissue-specific genes co-expressed in specific modules, providing a landscape of co-expression networks of tissue-specific genes in L. chinense.
Analysis of the co-expression network of tissue-specific genes in specific co-expression modules identified two tissue-specific TFs (AIL5 and SPL/NZZ) with important research value that were also hub genes in the co-expression modules. A previous study revealed that AIL genes synergistically regulate the function of the shoot apical meristem (Mudunkothge and Krizek, 2012). However, other studies showed that AIL5 contributes to flower development (Nole-Wilson et al., 2005; Krizek, 2015). As a shoot-apex-specific hub gene, the function of LcAIL5 need be further studied. Additionally, studies revealed that SPL/NZZ participates in stamen identity determination and is essential for anther development in A. thaliana (Liu et al., 2009; Zhao et al., 2017; Li et al., 2019). SPL/NZZ was reported to be expressed in stamen primordia and developing stamens (Zhao et al., 2017); in our study, SPL/NZZ was specifically expressed in the stamen. Additionally, genes co-expressed with SPL/NZZ were significantly enriched in phenylpropanoid biosynthesis. Phenylpropanoid derivatives are important components of sporopollenin, and the pollen exine comprises sporopollenin, which protects the pollen spore (Xue et al., 2020). Thus, we speculate that LcSPL/NZZ may be involved in stamen identity determination and anther development regulation, similar to AtSPL/NZZ, but this hypothesis required further verification. Furthermore, we found that bracts may be an important component of the L. chinense defense system to protect other floral tissues because most of the bract-specific hub genes were related to plant defense, such as PR, LecRLK, and LRR-RLK. Previous studies have shown that PR, LecRLK, and LRR-RLK play roles in plant immunity and pathogen defense (Parrott et al., 2016; Wang and Bouwmeester, 2017; Farrakh et al., 2018). These findings indicate that tissue-specific genes play a significant role in maintaining tissue characteristics in L. chinense.
Overall, this work enhanced our knowledge of the effect of APA on L. chinense transcriptome complexity and provided a landscape of lncRNAs and tissue-specific gene co-expression modules. This study also identified two hub TFs and 18 tissue-specific hub genes with important research value that were essential for maintaining tissue characteristics.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
ZT and HL: experimental design. ZT, YS, HL, and SW: plant material collection and performing the experiments. ZT, YS, and LW: data analysis. ZT and HL: manuscript writing. All authors read and approved the final manuscript.
Funding
The design of the study, sample collection, NGS and PacBio sequencing, data analysis, and interpretation were supported by the National Natural Science Foundation of China (31770718, 31470660) and by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We wish to thank Prof. Meng Xu (Nanjing Forestry University, Nanjing, China) and master Tengfei Shen (Nanjing Forestry University, Nanjing, China) for suggestions concerning the data analysis and writing. We would also like to thank Novogene Company (Tianjin, China) for RNA sequencing. In addition, we thank their laboratory colleagues for their help in data analysis and writing, including Ziyuan Hao, Weiping Zhong, Lichun Yang, Xinyu Zhai, Xujia Wu, Shan Hu, Kang Xu, Shenghua Zhu, and Longjie Ni.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2021.705321/full#supplementary-material
Supplementary Figure 1 | GO and KEGG pathway enrichment analysis of tissue-specific genes. (A) GO enrichment analysis of tissue-specific genes (only the top 30 terms are shown based on corrected P-value, and terms with asterisks indicate significantly enriched terms). (B) KEGG pathway enrichment analysis of tissue-specific genes.
Supplementary Figure 2 | Hierarchical clustering analysis of tissue-specific genes in different modules. (A–G) Hierarchical clustering analysis of bract-specific genes in the red module, leaf-specific genes in the brown module, petal-specific genes in the green module, pistil-specific genes in the salmon module, sepal-specific genes in the midnight blue module, stamen-specific genes in the turquoise module, and shoot-apex-specific genes in the yellow module, respectively.
Supplementary Figure 3 | KEGG pathway enrichment analysis of tissue-specific genes in different co-expression modules (only significantly enriched pathways are shown). (A–G) KEGG pathway enrichment analysis of bract-specific genes in the red module, petal-specific genes in the green module, sepal-specific genes in the midnight blue module, shoot-apex-specific genes in the yellow module, leaf-specific genes in the brown module, pistil-specific genes in the salmon module, and stamen-specific genes in the turquoise module.
Supplementary Figure 4 | Co-expression networks of tissue-specific genes in specific modules. Yellow, green, salmon, brown, red, turquoise, and midnight blue dots represent hub genes in the yellow, green, salmon, brown, red, turquoise, and midnight blue modules, respectively.
Supplementary Table 1 | Primers used for RT-qPCR validation.
Supplementary Table 2 | Information about the PacBio sequencing data.
Supplementary Table 3 | Transcript lengths before and after correction by Illumina reads.
Supplementary Table 4 | Statistics of the GMAP mapping results.
Supplementary Table 5 | Information of twenty hub genes in specific co-expression modules.
Footnotes
- ^ http://bioconductor.org/packages/release/bioc/html/DESeq2.html
- ^ http://www.bioconductor.org/packages/release/bioc/html/goseq.html
- ^ http://kobas.cbi.pku.edu.cn/download.php
- ^ http://www.cytoscape.org/download.html
- ^ https://gephi.org/users/download/
- ^ https://en.freedownloadmanager.org/Windows-PC/OLIGO.html
References
Abdel-Ghany, S. E., Hamilton, M., Jacobi, J. L., Ngam, P., Devitt, N., Schilkey, F., et al. (2016). A survey of the sorghum transcriptome using single-molecule long reads. Nat. Commun. 7:11706. doi: 10.1038/ncomms11706
Alamancos, G. P., Pages, A., Trincado, J. L., Bellora, N., and Eyras, E. (2015). Leveraging transcript quantification for fast computation of alternative splicing profiles. RNA 21, 1521–1531. doi: 10.1261/rna.051557.115
Alonso-Serra, J., Safronov, O., Lim, K. J., Fraser-Miller, S. J., Blokhina, O. B., Campilho, A., et al. (2019). Tissue-specific study across the stem reveals the chemistry and transcriptome dynamics of birch bark. New Phytol. 222, 1816–1831. doi: 10.1111/nph.15725
Amini, H., Naghavi, M. R., Shen, T., Wang, Y., Nasiri, J., Khan, I. A., et al. (2019). Tissue-specific transcriptome analysis reveals candidate genes for terpenoid and phenylpropanoid metabolism in the medicinal plant ferula assafoetida. G3 (Bethesda) 9, 807–816. doi: 10.1534/g3.118.200852
Bardou, F., Ariel, F., Simpson, C. G., Romero-Barrios, N., Laporte, P., Balzergue, S., et al. (2014). Long noncoding RNA modulates alternative splicing regulators in Arabidopsis. Dev Cell 30, 166–176. doi: 10.1016/j.devcel.2014.06.017
Cao, J., Ye, C., Hao, G., Dabney-Smith, C., Hunt, A. G., and Li, Q. Q. (2019). Root hair single cell type specific profiles of gene expression and alternative polyadenylation under cadmium stress. Front. Plant Sci. 10:589. doi: 10.3389/fpls.2019.00589
Celedon, J. M., Yuen, M. M. S., Chiang, A., Henderson, H., Reid, K. E., and Bohlmann, J. (2017). Cell-type- and tissue-specific transcriptomes of the white spruce (Picea glauca) bark unmask fine-scale spatial patterns of constitutive and induced conifer defense. Plant J. 92, 710–726. doi: 10.1111/tpj.13673
Chao, Y., Yuan, J., Guo, T., Xu, L., Mu, Z., and Han, L. (2019). Analysis of transcripts and splice isoforms in Medicago sativa L. by single-molecule long-read sequencing. Plant Mol. Biol. 99, 219–235. doi: 10.1007/s11103-018-0813-y
Chao, Y., Yuan, J., Li, S., Jia, S., Han, L., and Xu, L. (2018). Analysis of transcripts and splice isoforms in red clover (Trifolium pratense L.) by single-molecule long-read sequencing. BMC Plant Biol. 18:300. doi: 10.1186/s12870-018-1534-8
Chen, J., Hao, Z., Guang, X., Zhao, C., Wang, P., Xue, L., et al. (2018). Liriodendron genome sheds light on angiosperm phylogeny and species-pair differentiation. Nat. Plants 5, 18–25. doi: 10.1038/s41477-018-0323-6
Chen, J., Xie, J., Duan, Y., Hu, H., Hu, Y., and Li, W. (2016). Genome-wide identification and expression profiling reveal tissue-specific expression and differentially-regulated genes involved in gibberellin metabolism between Williams banana and its dwarf mutant. BMC Plant Biol. 16:123. doi: 10.1186/s12870-016-0809-1
Chen, S. Y., Deng, F., Jia, X., Li, C., and Lai, S. J. (2017a). A transcriptome atlas of rabbit revealed by PacBio single-molecule long-read sequencing. Sci. Rep. 7, 7648–7657. doi: 10.1038/s41598-017-08138-z
Chen, W., Jia, Q., Song, Y., Fu, H., Wei, G., and Ni, T. (2017b). Alternative polyadenylation: methods, findings, and impacts. Genomics Proteomics Bioinformatics 15, 287–300. doi: 10.1016/j.gpb.2017.06.001
Du, Q., Wang, K., Zou, C., Xu, C., and Li, W. X. (2018). The PILNCR1-miR399 regulatory module is important for low phosphate tolerance in maize. Plant Physiol. 177, 1743–1753. doi: 10.1104/pp.18.00034
Fan, T., Zhang, Q., Hu, Y., Wang, Z., and Huang, Y. (2020). Genome-wide identification of lncRNAs during hickory (Carya cathayensis) flowering. Funct. Integr. Genomics. 20, 591–607. doi: 10.1007/s10142-020-00737-w
Farrakh, S., Wang, M., and Chen, X. (2018). Pathogenesis-related protein genes involved in race-specific all-stage resistance and non-race specific high-temperature adult-plant resistance to Puccinia striiformis f. sp. tritici in wheat. J. Integr. Agric. 17, 2478–2491. doi: 10.1016/s2095-3119(17)61853-7
Finn, R. D., Coggill, P., Eberhardt, R. Y., Eddy, S. R., Mistry, J., Mitchell, A. L., et al. (2016). The Pfam protein families database: towards a more sustainable future. Nucleic Acids Res. 44(D1), D279–D285. doi: 10.1093/nar/gkv1344
Golicz, A. A., Singh, M. B., and Bhalla, P. L. (2018). The Long Intergenic Noncoding RNA (LincRNA) Landscape of the Soybean Genome. Plant Physiol. 176, 2133–2147. doi: 10.1104/pp.17.01657
Guo, Z., Qin, S., Li, C., Bao, L., Yan, W., Wu, H., et al. (2020). Genome-wide identification and functional prediction of long Non-coding RNAs in the Genesis of Populus euphratica Oliv. Heteromorphic Leaves. J. Plant Biol. 63, 131–140. doi: 10.1007/s12374-020-09229-7
Hao, R., He, S., Tang, S., and Wu, S. (1995). Geographical distribution of Liriodendron chinense in China and its significance. J. Plant Resour. Environ. 4, 1–6.
Harrow, J., Frankish, A., Gonzalez, J. M., Tapanari, E., Diekhans, M., Kokocinski, F., et al. (2012). GENCODE: the reference human genome annotation for The ENCODE Project. Genome Res. 22, 1760–1774. doi: 10.1101/gr.135350.111
Heo, J. B., and Sung, S. (2011). Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science 331, 76–79. doi: 10.1126/science.1197349
Hoang, N. V., Furtado, A., Mason, P. J., Marquardt, A., Kasirajan, L., Thirugnanasambandam, P. P., et al. (2017). A survey of the complex transcriptome from the highly polyploid sugarcane genome using full-length isoform sequencing and de novo assembly from short read sequencing. BMC Genomics 18:395. doi: 10.1186/s12864-017-3757-8
Hu, H., Yang, W., Zheng, Z., Niu, Z., Yang, Y., Wan, D., et al. (2020). Analysis of alternative splicing and alternative polyadenylation in populus alba var. pyramidalis by single-molecular long-read sequencing. Front. Genet. 11:48. doi: 10.3389/fgene.2020.00048
Julian, C., Rodrigo, J., and Herrero, M. (2011). Stamen development and winter dormancy in apricot (Prunus armeniaca). Ann. Bot. 108, 617–625. doi: 10.1093/aob/mcr056
Kanehisa, M., and Goto, S. (1999). KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30.
Kong, L., Zhang, Y., Ye, Z. Q., Liu, X. Q., Zhao, S. Q., Wei, L., et al. (2007). CPC: assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 35(Web Server issue), W345–W349. doi: 10.1093/nar/gkm391
Krizek, B. A. (2015). AINTEGUMENTA-LIKE genes have partly overlapping functions with AINTEGUMENTA but make distinct contributions to Arabidopsis thaliana flower development. J. Exp. Bot. 66, 4537–4549. doi: 10.1093/jxb/erv224
Langfelder, P., and Horvath, S. (2008). WGCNA: an R package for weighted correlation network analysis. BMC Bioinf. 9:559. doi: 10.1186/1471-2105-9-559
Li, A., Zhang, J., and Zhou, Z. (2014). PLEK: a tool for predicting long non-coding RNAs and messenger RNAs based on an improved k-mer scheme. BMC Bioinf. 15:311. doi: 10.1186/1471-2105-15-311
Li, S., Yamada, M., Han, X., Ohler, U., and Benfey, P. N. (2016). High-resolution expression map of the arabidopsis root reveals alternative splicing and lincRNA Regulation. Dev. Cell 39, 508–522. doi: 10.1016/j.devcel.2016.10.012
Li, X., Lian, H., Zhao, Q., and He, Y. (2019). MicroRNA166 Monitors SPOROCYTELESS/NOZZLE for building of the anther internal boundary. Plant Physiol. 181, 208–220. doi: 10.1104/pp.19.00336
Li, Y., Fang, C., Fu, Y., Hu, A., Li, C., Zou, C., et al. (2018). A survey of transcriptome complexity in Sus scrofa using single-molecule long-read sequencing. DNA Res. 25, 421–437. doi: 10.1093/dnares/dsy014
Liu, F., Marquardt, S., Lister, C., Swiezewski, S., and Dean, C. (2010). Targeted 3′ processing of antisense transcripts triggers Arabidopsis FLC Chromatin silencing. Science 327, 94–97.
Liu, S., Sun, Z., and Xu, M. (2018). Identification and characterization of long non-coding RNAs involved in the formation and development of poplar adventitious roots. Ind. Crops Prod. 118, 334–346. doi: 10.1016/j.indcrop.2018.03.071
Liu, S., Wang, L., Cao, M., Pang, S., Li, W., Kato-Noguchi, H., et al. (2020). Identification and characterization of long non-coding RNAs regulating flavonoid biosynthesis in Ginkgo biloba leaves. Ind. Crops Prod. 158:112980. doi: 10.1016/j.indcrop.2020.112980
Liu, S., Wu, L., Qi, H., and Xu, M. (2019). LncRNA/circRNA–miRNA–mRNA networks regulate the development of root and shoot meristems of Populus. Ind. Crops Prod. 133, 333–347. doi: 10.1016/j.indcrop.2019.03.048
Liu, X., Huang, J., Parameswaran, S., Ito, T., Seubert, B., Auer, M., et al. (2009). The SPOROCYTELESS/NOZZLE gene is involved in controlling stamen identity in Arabidopsis. Plant Physiol. 151, 1401–1411. doi: 10.1104/pp.109.145896
Ma, J., Wei, L., Li, J., and Li, H. (2018). The analysis of genes and phytohormone metabolic pathways associated with leaf shape development in liriodendron chinense via de novo transcriptome sequencing. Genes (Basel) 9, 577–592. doi: 10.3390/genes9120577
Mao, Q., Chen, C., Xie, T., Luan, A., Liu, C., and He, Y. (2018). Comprehensive tissue-specific transcriptome profiling of pineapple (Ananas comosus) and building an eFP-browser for further study. PeerJ. 6, e6028–e6043. doi: 10.7717/peerj.6028
Mathesius, U. (2018). Flavonoid functions in plants and their interactions with other organisms. Plants (Basel) 7:30. doi: 10.3390/plants7020030
Mayya, V. K., and Duchaine, T. F. (2019). Ciphers and executioners: how 3′-untranslated regions determine the fate of messenger RNAs. Front. Genet. 10:6. doi: 10.3389/fgene.2019.00006
Mortazavi, A., Williams, B. A., McCue, K., Schaeffer, L., and Wold, B. (2008). Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 5, 621–628. doi: 10.1038/nmeth.1226
Mudunkothge, J. S., and Krizek, B. A. (2012). Three Arabidopsis AIL/PLT genes act in combination to regulate shoot apical meristem function. Plant J. 71, 108–121. doi: 10.1111/j.1365-313X.2012.04975.x
Nole-Wilson, S., Tranby, T. L., and Krizek, B. A. (2005). AINTEGUMENTA-like (AIL) genes are expressed in young tissues and may specify meristematic or division-competent states. Plant Mol. Biol. 57, 613–628. doi: 10.1007/s11103-005-0955-6
Parrott, D. L., Huang, L., and Fischer, A. M. (2016). Downregulation of a barley (Hordeum vulgare) leucine-rich repeat, non-arginine-aspartate receptor-like protein kinase reduces expression of numerous genes involved in plant pathogen defense. Plant Physiol. Biochem. 100, 130–140. doi: 10.1016/j.plaphy.2016.01.005
Pattison, R. J., Csukasi, F., Zheng, Y., Fei, Z., van der Knaap, E., and Catala, C. (2015). Comprehensive tissue-specific transcriptome analysis reveals distinct regulatory programs during early tomato fruit development. Plant Physiol. 168, 1684–1701. doi: 10.1104/pp.15.00287
Pruitt, K. D., Tatusova, T., and Maglott, D. R. (2007). NCBI reference sequences (RefSeq): a curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 35(Database issue), D61–D65. doi: 10.1093/nar/gkl842
Quinn, J. J., and Chang, H. Y. (2016). Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 17, 47–62. doi: 10.1038/nrg.2015.10
Ruan, J., Guo, F., Wang, Y., Li, X., Wan, S., Shan, L., et al. (2018). Transcriptome analysis of alternative splicing in peanut (Arachis hypogaea L.). BMC Plant Biol. 18:139. doi: 10.1186/s12870-018-1339-9
Sadek, J., Omer, A., Hall, D., Ashour, K., and Gallouzi, I. E. (2019). Alternative polyadenylation and the stress response. Wiley Interdiscip. Rev. RNA 10:e1540. doi: 10.1002/wrna.1540
Salmela, L., and Rivals, E. (2014). LoRDEC: accurate and efficient long read error correction. Bioinformatics 30, 3506–3514. doi: 10.1093/bioinformatics/btu538
Shen, Y., Venu, R. C., Nobuta, K., Wu, X., Notibala, V., Demirci, C., et al. (2011). Transcriptome dynamics through alternative polyadenylation in developmental and environmental responses in plants revealed by deep sequencing. Genome Res. 21, 1478–1486. doi: 10.1101/gr.114744.110
Smoot, M. E., Ono, K., Ruscheinski, J., Wang, P. L., and Ideker, T. (2011). Cytoscape 2.8: new features for data integration and network visualization. Bioinformatics 27, 431–432. doi: 10.1093/bioinformatics/btq675
Sreya, G., and Chon-Kit Kenneth, C. (2016). Analysis of RNA-Seq data using tophat and cufflinks. Methods Mol. Biol. 1374, 339–361.
Sun, L., Luo, H., Bu, D., Zhao, G., Yu, K., Zhang, C., et al. (2013). Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 41, e166–e173. doi: 10.1093/nar/gkt646
Tatusov, R. L., Fedorova, N. D., Jackson, J. D., Jacobs, A. R., Kiryutin, B., Koonin, E. V., et al. (2003). The COG database: an updated version includes eukaryotes. BMC Bioinf. 4:41. doi: 10.1186/1471-2105-4-41
The UniProt, C. (2017). UniProt: the universal protein knowledgebase. Nucleic Acids Res. 45(D1), D158–D169. doi: 10.1093/nar/gkw1099
Tian, J., Song, Y., Du, Q., Yang, X., Ci, D., Chen, J., et al. (2016). Population genomic analysis of gibberellin-responsive long non-coding RNAs in Populus. J. Exp. Bot. 67, 2467–2482. doi: 10.1093/jxb/erw057
Tu, Z., Hao, Z., Zhong, W., and Li, H. (2019). Identification of Suitable Reference Genes for RT-qPCR Assays in Liriodendron chinense (Hemsl.) Sarg. Forests 10:441. doi: 10.3390/f10050441
Tu, Z., Shen, Y., Wen, S., Zong, Y., and Li, H. (2020). Alternative splicing enhances the transcriptome complexity of liriodendron chinense. Front. Plant Sci. 11:578100. doi: 10.3389/fpls.2020.578100
Turner, R. E., Pattison, A. D., and Beilharz, T. H. (2018). Alternative polyadenylation in the regulation and dysregulation of gene expression. Semin. Cell Dev. Biol. 75, 61–69. doi: 10.1016/j.semcdb.2017.08.056
van Dijk, E. L., Jaszczyszyn, Y., Naquin, D., and Thermes, C. (2018). The third revolution in sequencing technology. Trends Genet. 34, 666–681. doi: 10.1016/j.tig.2018.05.008
Venkataraman, K., Brown, K. M., and Gilmartin, G. M. (2005). Analysis of a noncanonical poly(A) site reveals a tripartite mechanism for vertebrate poly(A) site recognition. Genes Dev. 19, 1315–1327. doi: 10.1101/gad.1298605
Villarino, G. H., Hu, Q., Scanlon, M. J., Mueller, L., Bombarely, A., and Mattson, N. S. (2017). Dissecting tissue-specific transcriptomic responses from leaf and roots under salt stress in petunia hybrida Mitchell. Genes (Basel) 8, 195–214. doi: 10.3390/genes8080195
Wang, B., Regulski, M., Tseng, E., Olson, A., Goodwin, S., McCombie, W. R., et al. (2018a). A comparative transcriptional landscape of maize and sorghum obtained by single-molecule sequencing. Genome Res. 28, 921–932. doi: 10.1101/gr.227462.117
Wang, B., Tseng, E., Regulski, M., Clark, T. A., Hon, T., Jiao, Y., et al. (2016). Unveiling the complexity of the maize transcriptome by single-molecule long-read sequencing. Nat. Commun. 7, 11708–11720. doi: 10.1038/ncomms11708
Wang, K. C., and Chang, H. Y. (2011). Molecular mechanisms of long noncoding RNAs. Mol. Cell. 43, 904–914. doi: 10.1016/j.molcel.2011.08.018
Wang, M., Wang, P., Liang, F., Ye, Z., Li, J., Shen, C., et al. (2018b). A global survey of alternative splicing in allopolyploid cotton: landscape, complexity and regulation. New Phytol. 217, 163–178. doi: 10.1111/nph.14762
Wang, R., Yang, Z., Fei, Y., Feng, J., Zhu, H., Huang, F., et al. (2019). Construction and analysis of degradome-dependent microRNA regulatory networks in soybean. BMC Genomics 20:534. doi: 10.1186/s12864-019-5879-7
Wang, T., Wang, H., Cai, D., Gao, Y., Zhang, H., Wang, Y., et al. (2017). Comprehensive profiling of rhizome-associated alternative splicing and alternative polyadenylation in moso bamboo (Phyllostachys edulis). Plant J. 91, 684–699. doi: 10.1111/tpj.13597
Wang, Y., and Bouwmeester, K. (2017). L-type lectin receptor kinases: new forces in plant immunity. PLoS Pathog. 13:e1006433. doi: 10.1371/journal.ppat.1006433
Wu, L., Liu, S., Qi, H., Cai, H., and Xu, M. (2020). Research progress on plant long non-coding RNA. Plants (Basel) 9:408. doi: 10.3390/plants9040408
Wu, T. D., and Watanabe, C. K. (2005). GMAP: a genomic mapping and alignment program for mRNA and EST sequences. Bioinformatics 21, 1859–1875. doi: 10.1093/bioinformatics/bti310
Wu, X., Liu, M., Downie, B., Liang, C., Ji, G., Li, Q. Q., et al. (2011). Genome-wide landscape of polyadenylation in Arabidopsis provides evidence for extensive alternative polyadenylation. Proc. Natl. Acad. Sci. U.S.A. 108, 12533–12538. doi: 10.1073/pnas.1019732108
Xu, X., Zhang, H., Xie, T., Xu, Y., Zhao, L., and Tian, W. (2017). Effects of climate change on the potentially suitable climatic geographical range of liriodendron chinense. Forests 8, 399–412. doi: 10.3390/f8100399
Xu, Z., Peters, R. J., Weirather, J., Luo, H., Liao, B., Zhang, X., et al. (2015). Full-length transcriptome sequences and splice variants obtained by a combination of sequencing platforms applied to different root tissues of Salvia miltiorrhiza and tanshinone biosynthesis. Plant J. 82, 951–961. doi: 10.1111/tpj.12865
Xue, J. S., Zhang, B., Zhan, H., Lv, Y. L., Jia, X. L., Wang, T., et al. (2020). Phenylpropanoid derivatives are essential components of Sporopollenin in vascular plants. Mol. Plant. 13, 1644–1653. doi: 10.1016/j.molp.2020.08.005
Yan, C., Wang, Y., Lyu, T., Hu, Z., Ye, N., Liu, W., et al. (2021). Alternative Polyadenylation in response to temperature stress contributes to gene regulation in Populus trichocarpa. BMC Genomics 22:53. doi: 10.1186/s12864-020-07353-9
Yang, L., Jin, Y., Huang, W., Sun, Q., Liu, F., and Huang, X. (2018). Full-length transcriptome sequences of ephemeral plant Arabidopsis pumila provides insight into gene expression dynamics during continuous salt stress. BMC Genomics 19:717. doi: 10.1186/s12864-018-5106-y
Yang, Y., Xu, M., Luo, Q., Wang, J., and Li, H. (2014). De novo transcriptome analysis of Liriodendron chinense petals and leaves by Illumina sequencing. Gene 534, 155–162. doi: 10.1016/j.gene.2013.10.073
Yao, S., Liang, F., Gill, R. A., Huang, J., Cheng, X., Liu, Y., et al. (2020). A global survey of the transcriptome of allopolyploid Brassica napus based on single-molecule long-read isoform sequencing and Illumina-based RNA sequencing data. Plant J. 103, 843–857. doi: 10.1111/tpj.14754
Ye, C., Zhou, Q., Wu, X., Ji, G., and Li, Q. Q. (2019). Genome-wide alternative polyadenylation dynamics in response to biotic and abiotic stresses in rice. Ecotoxicol. Environ. Saf. 183:109485. doi: 10.1016/j.ecoenv.2019.109485
Young, M. D., Wakefield, M. J., Smyth, G. K., and Oshlack, A. (2010). Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol. 11, R14–R25.
Yu, L., Ma, J., Niu, Z., Bai, X., Lei, W., Shao, X., et al. (2017). Tissue-Specific Transcriptome Analysis Reveals Multiple Responses to Salt Stress in Populus euphratica Seedlings. Genes (Basel) 8, 372–385. doi: 10.3390/genes8120372
Yu, Y., Zhou, Y. F., Feng, Y. Z., He, H., Lian, J. P., Yang, Y. W., et al. (2020). Transcriptional landscape of pathogen-responsive lncRNAs in rice unveils the role of ALEX1 in jasmonate pathway and disease resistance. Plant Biotechnol. J. 18, 679–690. doi: 10.1111/pbi.13234
Zhang, J., Gu, H., Dai, H., Zhang, Z., and Miao, M. (2020). Alternative polyadenylation of the stacyose synthase gene mediates source-sink regulation in cucumber. J. Plant Physiol. 245, 153111. doi: 10.1016/j.jplph.2019.153111
Zhang, J., Liu, C., He, M., Xiang, Z., Yin, Y., Liu, S., et al. (2019). A full-length transcriptome of Sepia esculenta using a combination of single-molecule long-read (SMRT) and Illumina sequencing. Mar. Genomics 43, 54–57. doi: 10.1016/j.margen.2018.08.008
Zhang, W. W., Niu, J. F., Wang, X. K., Tian, Y., Yao, F. F., and Feng, Z. Z. (2011). Effects of ozone exposure on growth and photosynthesis of the seedlings of Liriodendron chinense (Hemsl.) Sarg, a native tree species of subtropical China. Photosynthetica 49, 29–36. doi: 10.1007/s11099-011-0003-5
Zhang, Y., Carrion, S. A., Zhang, Y., Zhang, X., Zinski, A. L., Michal, J. J., et al. (2018). Alternative polyadenylation analysis in animals and plants: newly developed strategies for profiling, processing and validation. Int. J. Biol. Sci. 14, 1709–1714. doi: 10.7150/ijbs.27168
Zhao, F., Zheng, Y. F., Zeng, T., Sun, R., Yang, J. Y., Li, Y., et al. (2017). Phosphorylation of SPOROCYTELESS/NOZZLE by the MPK3/6 Kinase Is required for anther development. Plant Physiol. 173, 2265–2277. doi: 10.1104/pp.16.01765
Zhao, H., Gao, Z., Wang, L., Wang, J., Wang, S., Fei, B., et al. (2018). Chromosome-level reference genome and alternative splicing atlas of moso bamboo (Phyllostachys edulis). Gigascience 7:giy115. doi: 10.1093/gigascience/giy115
Zhao, J., Ajadi, A. A., Wang, Y., Tong, X., Wang, H., Tang, L., et al. (2020). Genome-wide identification of lncRNAs during rice seed development. Genes (Basel) 11, 243. doi: 10.3390/genes11030243
Keywords: L. chinense, hybrid sequencing, alternative polyadenylation, transcription factors, tissue-specific genes, gene co-expression modules, lncRNAs
Citation: Tu Z, Shen Y, Wen S, Liu H, Wei L and Li H (2021) A Tissue-Specific Landscape of Alternative Polyadenylation, lncRNAs, TFs, and Gene Co-expression Networks in Liriodendron chinense. Front. Plant Sci. 12:705321. doi: 10.3389/fpls.2021.705321
Received: 05 May 2021; Accepted: 28 June 2021;
Published: 23 July 2021.
Edited by:
Agnieszka Zmienko, Institute of Bioorganic Chemistry (PAS), PolandReviewed by:
Anna Philips, Institute of Bioorganic Chemistry (PAS), PolandBiao Jin, Yangzhou University, China
Copyright © 2021 Tu, Shen, Wen, Liu, Wei and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Huogen Li, hgli@njfu.edu.cn