 Weipeng Liu1†
Weipeng Liu1† Yunyun Zhang1†Fengdan Huang2†Qianli Ma3
Yunyun Zhang1†Fengdan Huang2†Qianli Ma3 Chuanyin Li1
Chuanyin Li1 Shuyuan Liu1Yan Liang1
Shuyuan Liu1Yan Liang1 Li Shi1*
Li Shi1* Yufeng Yao1*
Yufeng Yao1*- 1Institute of Medical Biology, Chinese Academy of Medical Sciences and Peking Union Medical College, Kunming, China
- 2Graduate School of Yunnan University, Yunnan University, Kunming, China
- 3Department of Thoracic Surgery, The 3rd Affiliated Hospital of Kunming Medical University, Kunming, China
Previous investigations have reported that microRNA-126 (miR-126) and its host gene, epidermal growth factor-like domain-containing protein 7 (EGFL7) are involved in lung cancer progression, suggesting EGFL7 and miR-126 play a joint role in lung cancer development. In this study, we analyzed the methylation-associated regulation of EGFL7 and miR-126 in non-small cell lung cancer (NSCLC) and further investigated the association between EGFL7/miR-126 polymorphisms and NSCLC susceptibility in the Han Chinese population. Based on our data, relative to those in adjacent normal tissue, both EGFL7 expression and miR-126 expression were decreased significantly in lung cancer tissue (P = 3x10-4 and P < 1x10-4), and the expression of EGFL7 mRNA and miR-126 was significantly correlated in both NSCLC tissue n = 46, r = 0.43, P = 0.003 and adjacent normal tissue n = 46, r = 0.37, P = 0.011. Differential methylation analysis indicated that methylation levels of multiple CG loci in EGFL7 were significantly higher in the lung cancer samples than in the normal samples (P < 0.01). Moreover, EGFL7 mRNA and miR-126 were significantly upregulated after treatment with the DNA demethylating agent 5-aza-2′-deoxycytidine (5-Aza-CdR) in lung cancer cell lines. In addition, the A allele of rs2297538 was significantly associated with a decreased NSCLC risk (OR = 0.68, 95% CI: 0.52~0.88), and the expression of EGFL7 and miR-126 was significantly lower in rs2297538 homozygous G/G tumor tissue than in A/G+A/A tumor tissue (P = 0.01 and P = 0.002). Our findings suggest that the expression of EGFL7 and miR-126 in NSCLC can be concomitantly downregulated through methylation and the EGFL7/miR-126 polymorphism rs2297538 is correlated with NSCLC risk. Together, these results provide new insights into the pathogenesis of NSCLC.
Introduction
Lung cancer is one of the most common cancers in the world; in 2020, there were more than 2.2 million newly reported lung cancer cases and over 1.79 million new lung cancer-related deaths worldwide (1). Non-small cell lung cancer (NSCLC) is the most prevalent lung cancer type, accounting for 85–90% of all lung cancer cases (2).
Recently, studies have demonstrated that genetic factors also play important roles in the occurrence of lung cancer, and twin studies have shown that the heritability of lung cancer is approximately 18% (3, 4). In recent years, a number of differentially expressed genes have been identified in lung cancer (5–7), and many researchers have turned their attention to the study of common single nucleotide polymorphisms (SNPs) in human carcinogenic/anticancer genes (8–10). These studies can not only expand our understanding of the pathogenesis of lung cancer but also provide new clues for the diagnosis and treatment of this disease.
Epidermal growth factor-like domain-containing protein 7 (EGFL7) is a secreted protein that was initially reported to play a key role in angiogenesis (11, 12). Subsequently, the dysregulation of EGFL7 has been found in a variety of tumors, including lung cancer (13), hepatocellular carcinoma (14), acute myeloid leukemia (15) and malignant pleural mesothelioma (16), suggesting that EGFL7 participates in tumorigenesis through a wide range of effects.
MicroRNA−126 (miR−126) is located within intron 7 of EGFL7 on human chromosome 9q34.3 (17). Recent studies have shown that miR-126 is downregulated in various cancer tissues, including breast cancer (18), pancreatic cancer (19) and lung cancer (20). In recent years, increasing evidence has demonstrated the role of miR−126 in lung tumorigenesis via targeting multiple genes including CCR1 (21), PIK3R2 (22), Crk (23) and so forth. In addition, miR−126 has been found to play a role in both diagnosis and prognosis of NSCLC (24, 25), thus indicating that miR−126 could be a promising biomarker in lung cancer.
Previous investigations have shown that miR−126 is not transcribed from its own promoter but is likely transcribed together with its host gene EGFL7 (17), indicating that the expression levels of miR-126 and EGFL7 may be regulated by the same mechanisms during the occurrence of NSCLC. Furthermore, studies have identified CpG islands around the transcription initiation site of EGFL7 (17); thus, it is reasonable that miR-126 and EGFL7 can be silenced by methylation of cytosine residues. In addition, recent studies have revealed that single nucleotide polymorphisms (SNPs) in gene regulatory and coding regions could confer risk of lung cancer by regulating the expression of specific genes (26–28). Thus, SNPs in the transcriptional regulatory region and coding region of EGFL7 might also play a role in the regulation of EGFL7 and miR-126 expression and are further involved in NSCLC susceptibility. Herein, we explored the methylation-associated regulation of miR-126 and EGFL7 in NSCLC and further investigated the association between EGFL7 polymorphisms and NSCLC susceptibility in the Han Chinese population. We showed that the expression of miR-126 and EGFL7 is concomitantly downregulated in NSCLC through methylation and that the eQTL-missense polymorphism of EGFL7 is associated with lung cancer risk in a Han Chinese population.
Materials and Methods
Subjects and Tissue Samples
This study was approved by the Ethics Committee of the Third Affiliated Hospital of Kunming Medical University, and all study protocols were performed in accordance with the Declaration of Helsinki. Written informed consent was acquired from each participant.
We performed a case-control association study, a total of 497 patients with NSCLC treated in the Third Affiliated Hospital of Kunming Medical University in Yunnan were selected as the case group, and 502 healthy people undergoing a physical examination in the hospital during the same period were selected as the control group. The NSCLC patients were diagnosed according to the Chinese Medical Association guidelines for clinical diagnosis and treatment of lung cancer (Edition 2018) at the Third Affiliated Hospital of Kunming Medical University. The lung cancer histological type and pathologic stage were identified according to the International System for Staging Lung Cancer (29). The NSCLC patient inclusion criteria: 1) the patients were histologically and pathologically diagnosed NSCLC (adenocarcinoma and squamous cell carcinoma); 2) the patients had not received chemotherapy and radiotherapy. The criteria for the exclusion was 1) the patients with a prior history of primary cancer other than lung cancer; 2) the patients with small cell lung cancer or unclear pathological diagnosis; 3) the patient with malignant tumors except lung cancer.
A total of 46 matched sets of primary NSCLC tumors and adjacent normal tissues were acquired from NSCLC patients at the Third Affiliated Hospital of Kunming Medical University. All tissues were identified by pathological examination and fresh-frozen at -80°C.
Quantitative RT–PCR
Total RNA was isolated from tissues or cells using TRIzol reagent. The PrimeScript™ RT reagent Kit with gDNA Eraser (TaKaRa Bio Inc, Tokyo, Japan) was used to synthesize cDNA. We reverse transcribed 1 µg of total RNA, and diluted cDNA at a final concentration of 20 µg/µL. Quantitative real-time PCR was carried out under the following conditions using SYBR Green to detect the expression levels of EGFL7 and miR-126 in NSCLC tissues and corresponding adjacent tissues in clinical patients: denaturation at 95°C for 10 min, followed by 40 cycles of denaturation at 95°C for 15 s, annealing at 60°C for 15 s, and extension at 72°C for 15 s. GAPDH was used as the internal control of EGFL7, and U6 was used as the internal control of miR-126. All the primers are listed in Table S1.
Cell Culture
All the cell lines used in this study were originally obtained from the ATCC. Human lung epithelial BEAS-2B cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Gibco) containing 10% fetal bovine serum (FBS, Gibco), SPC-A1 cells were cultured in RPMI-1640 with 10% fetal bovine serum (FBS, Gibco), H1299 cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Gibco) containing 10% fetal bovine serum (FBS, Gibco). Cells were cultured in a cell incubator at 37°C with 5% CO2.
5-Aza-2’-Deoxycytidine (5-Aza-CdR) Treatment
5-Aza-CdR has been widely used to demonstrate the correlation between loss of methylation in specific gene regions and activation of the associated genes (30). In the current study, BEAS-2B, SPC-A1 and H1299 cells were seeded in six-well culture dishes 24 h prior to treatment with 5-Aza-CdR. 5-Aza-CdR was continuously administered by replacing the medium containing 5-Aza-CdR (0 μM and 10 μM) every 24 h for 2 days. The dose of 5-Aza-CdR (10 μM) was chosen based on our preliminary studies. Similar reactivation was shown of EGFL7 and miR-126 expression when cells were treated with varying concentrations (1–10 μM) of 5-Aza-CdR as well as previously published studies (31, 32). Cells were then harvested for total RNA extraction to demonstrate whether treatment with the demethylating agent was able to increase EGFL7 and miR-126 mRNA expression in these cell lines.
SNP Selection
In this study, 1670 bp upstream of the EGFL7/miR-126 transcription start site was chosen as the promoter region according to a previous study (33). JASPAR (http://jaspar.genereg.net/) was used to predict whether the SNPs in the promoter region of EGFL7 are located in the transcription factor binding site and disrupt the binding of specific transcription factors (34). Missense variants with a minor allele frequency (MAF) greater than 0.05 were called and filtered using the Ensembl Variant Effect Predictor (http://www.ensembl.org/vep) (35). Finally, rs1332793, rs9411260 and rs2297538 were chosen as candidate SNPs in the current study.
DNA Extraction and Sequencing
A heparin anticoagulant tube was used to collect 10 ml of venous blood from the study subjects. DNA was extracted by a QIAamp DNA Blood Mini Kit (Qiagen, Hilden, Germany), and the concentration and purity of the genomic DNA were detected by a Multiskan Skyhigh full-wavelength enzyme plate (ND-2000, Thermo Fisher Scientific). After genomic DNA was extracted, it was stored in a -20°C refrigerator for later use.
Three SNPs rs1332793, rs9411260 and rs2297538 were genotyped by TaqMan probe real-time fluorescence quantitative polymerase chain reaction (RTFQ-PCR). The probes and primers were designed and produced by Thermo Fisher Scientific Company (Waltham, MA, USA), and TaqMan Genotyping Master Mix was purchased from ABI. PCR amplification was carried out in 384-well reaction plates with 2.5 μL Master Mix, 0.125 μL primer and probe (FAM and VIC) mix, 1.375 μL ddH2O and 1 μL genomic DNA in each well. Amplification was conducted in a QuantStudio 6 Flex Fast Real-Time PCR system as follows: 95°C preheat denaturing for 10 min, 92°C for 10 s and 60°C for 1 min, all repeated for 40 cycles. We also confirmed the genotyping results of 20 randomly selected individuals using Sanger sequencing, and no genotyping errors were found.
In-Silico Analysis of EGFL7 Expression and Methylation Status
GEPIA was used to explore the relative expression of EGFL7 in lung cancer tissue and normal tissue (http://gepia.cancer-pku.cn/), which provides the expression data of tumors and normal samples from the TCGA and GTEx projects (36). SurvivalMeth was used to explore the methylation status of EGFL7 in lung cancer tumor tissues and normal tissues (http://bio-bigdata.hrbmu.edu.cn/survivalmeth) (37).
Statistical Analysis
GraphPad Prism 8.3.0 software was used for statistical analysis; Hardy-Weinberg equilibrium was used to test sample representativeness. Student’s t-test was used to analyze the age difference between the NSCLC group and the control group. The χ2 test was used to analyze whether there was a sex difference between the NSCLC group and the control group. The differences in allele frequencies and genotype distribution of rs1332793, rs9411260, and rs2297538 in the NSCLC group and control groups were also analyzed via χ2 test, and the significance threshold was set at P<0.017 (0.05/3). SHEsis online software (38) was used to analyze the linkage disequilibrium between SNPs, and D’ > 0.8 indicates strong linkage between SNPs. The genetic pattern of SNPs was analyzed by SnpStats online software (39), obtaining the optimal genetic pattern according to the AIC (akaike information criterion) and BIC (bayesian information criterion) values. Relative expression levels of EGFL7 and miR-126 are presented as the means of 2−ΔΔCt. Statistical tests against different groups were conducted using a two-tailed t-test. Spearman’s correlation analysis was performed to detect the correlation between EGFL7 and miR-126 expression in NSCLC tumor tissue and adjacent normal tissue. The significance threshold was set at P<0.05. Statistical analyses were performed using SPSS 21 (Chicago, IL) and GraphPad Prism 7.00.
Results
Expression of miR-126 and EGFL7 mRNA Is Significantly Reduced in NSCLC Tissues Compared With Adjacent Normal Tissues
Previous studies have reported low miR-126 expression in lung cancer tissue (20). In silico analysis of EGFL7 mRNA expression indicated that the expression of EGFL7 in lung cancer tissue was lower than that in normal lung tissue (P < 0.01, Figure S1). To validate these results, we tested miR-126 and EGFL7 mRNA expression in lung cancer tissue along with matched adjacent normal tissue from 46 NSCLC patients using qRT–PCR. We found that EGFL7 and miR-126 expression decreased significantly in lung cancer tissue compared with adjacent normal tissue (P = 3x10-4 and P < 1x10-4, respectively, Figures 1A, B), which was consistent with in silico analysis (Figure S1) and previous results (20).
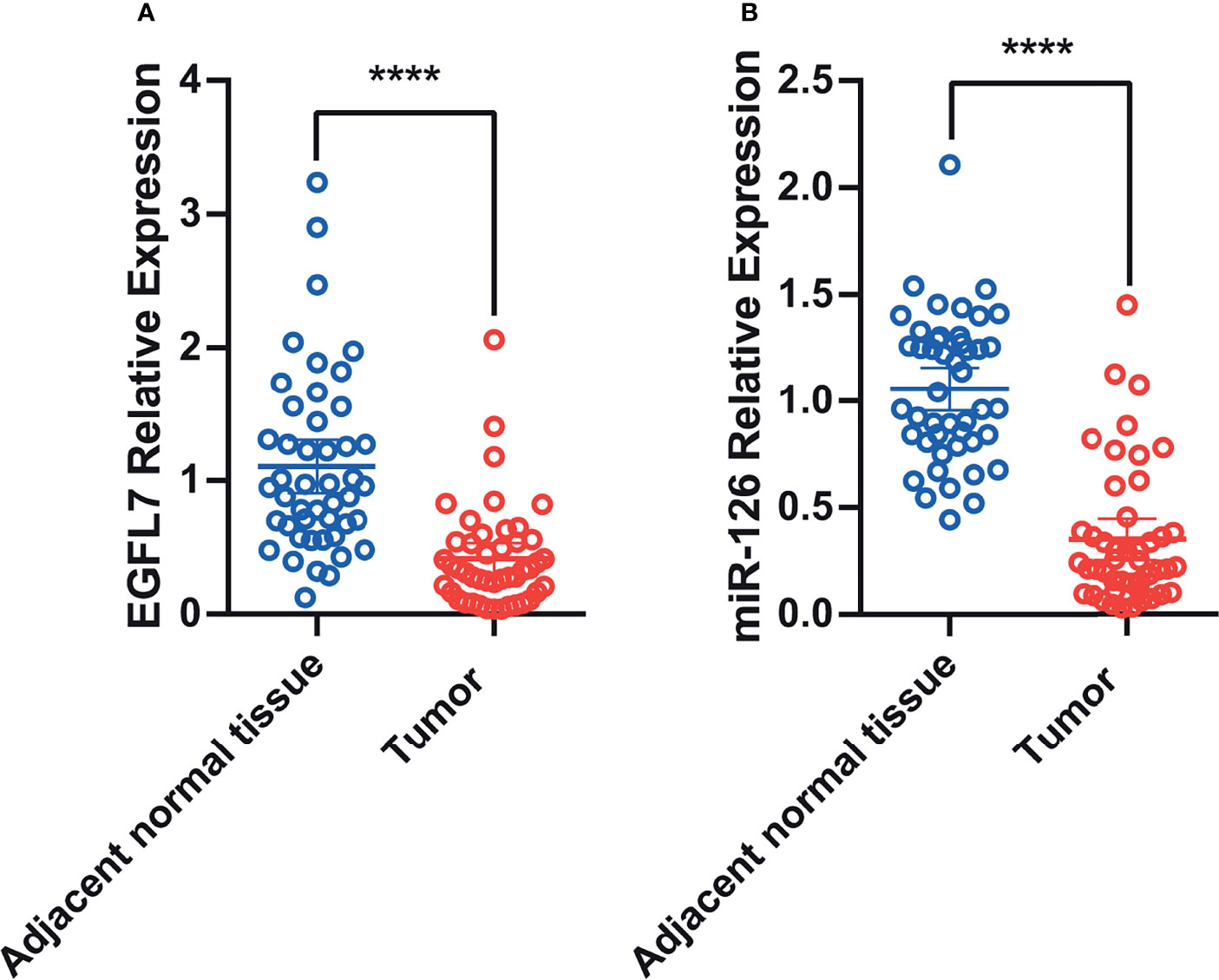
Figure 1 Scatter plot of EGFL7 mRNA (A) and miR-126 (B) expression in lung tumor tissue and adjacent normal tissues. The values on Y-axis were presented as 2–ΔΔCt. **** represent P ≤ 0.0001.
Expression of miR-126 and EGFL7 mRNA Is Correlated in NSCLC Tissues and Adjacent Normal Tissues
To further investigate whether miR-126 and EGFL7 were concomitantly expressed in NSCLC as previously described for prostate cancer (17) and malignant pleural mesothelioma (16), we performed spearman’s correlation analysis to detect the correlation of miR-126 and EGFL7 expression in both NSCLC tissues and adjacent normal tissues. Our results showed that the expression of EGFL7 and miR-126 were significantly correlated in both NSCLC tissues (n = 46, r = 0.43, P = 0.003, Figure 2) and adjacent normal tissues (n = 46, r = 0.37, P = 0.011, Figure 2).
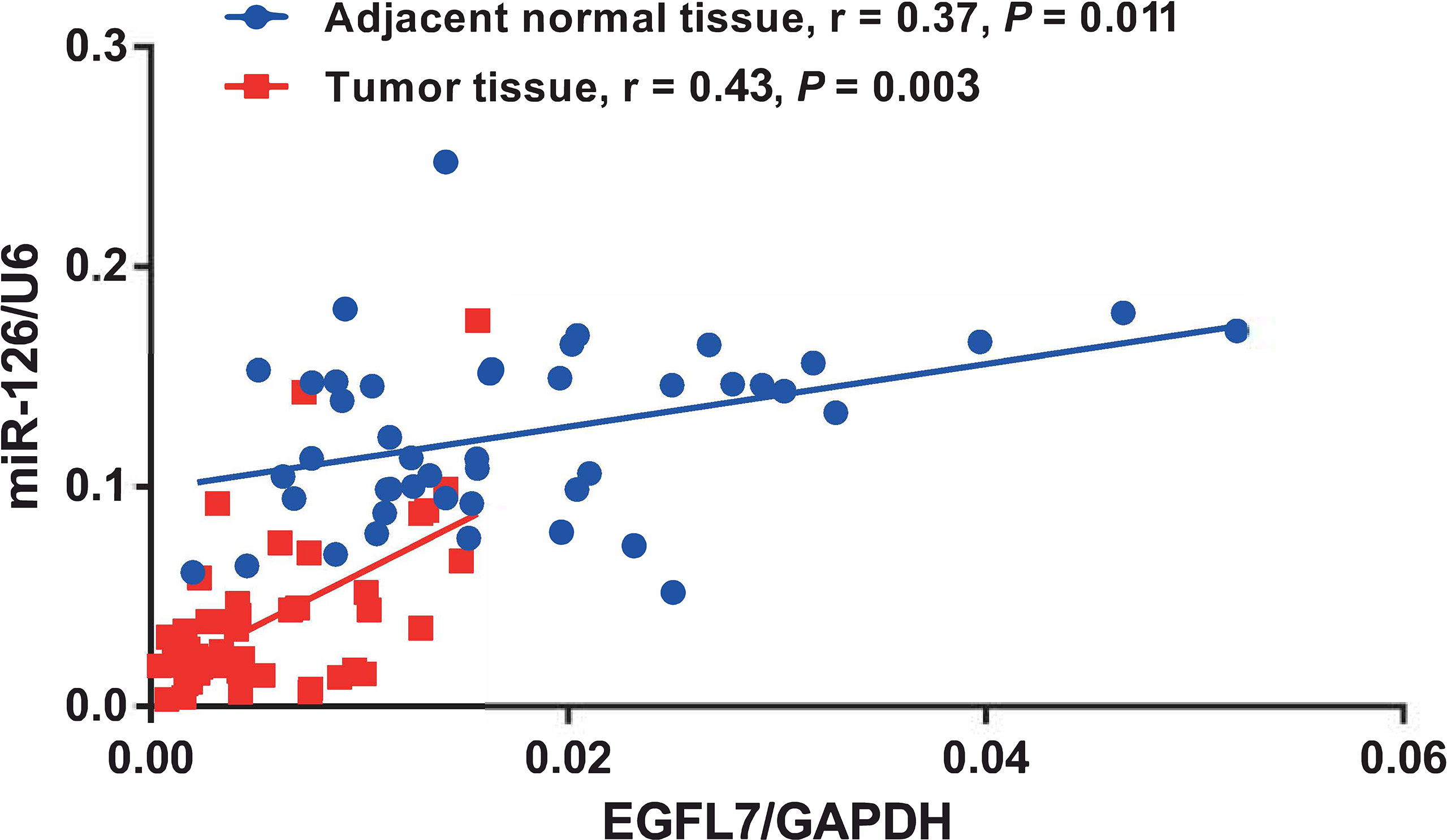
Figure 2 Correlation between EGFL7 and miR-126 expression in NSCLC tumor tissue and adjacent normal tissue. Spearman’s correlation coefficient (r) and P values are shown for each analysis.
EGFL7 and miR-126 Are Upregulated After Treatment With the DNA Demethylating Agent 5-aza-2’-Deoxycytidine (5-Aza-CdR) in Lung Cancer Cell Lines
Previous studies have reported CpG islands in EGFL7 (17). To investigate the methylation status of EGFL7 in lung cancer tumor tissues and normal tissues, we performed the differential methylation analysis of EGFL7 CG loci in patients with adenocarcinoma (AC) and squamous cell carcinoma (SCC) using the SurvivalMeth database (http://bio-bigdata.hrbmu.edu.cn/survivalmeth). Our analysis revealed that the methylation levels of 4 CG loci (cg08529852, cg14548542, cg17443080 and cg20997159) within EGFL7 were significantly higher in the AC samples than in the normal samples (P < 0.01, Figure S2). Furthermore, the methylation levels of 6 CG loci (cg04074066, cg05936059, cg08529852, cg14353956, cg14548542 and cg20997159) were significantly higher in the SCC samples than in the normal samples, while the methylation level of cg21184800 markedly decreased in SCC tumor samples (P < 0.01, Figure S2). These results were consistent with the result that EGFL7 mRNA expression decreased in lung tumor tissue and indicated that the expression of EGFL7 mRNA in NSCLC can be downregulated through methylation.
To verify the expression of miR-126 and EGFL7 can be silenced by methylation of cytosine residues in lung cancer cells. BEAS-2B (non-tumorigenic human bronchial epithelial cells), SPC-A1 and H1299 cells were treated with 5-Aza-CdR (10 μM) for 48 h. After treatment with the DNA demethylating agent 5-Aza-CdR, EGFL7 mRNA and miR-126 were significantly upregulated in both SPC-A1 and H1299 lung cancer cell lines (P < 0.05, Figure 3), while no significant difference of EGFL7 mRNA expression was observed in BEAS-2B (Figure 3). Overall, our results indicated that the expression of miR-126 and EGFL7 mRNA in NSCLC can be concomitantly regulated through methylation.
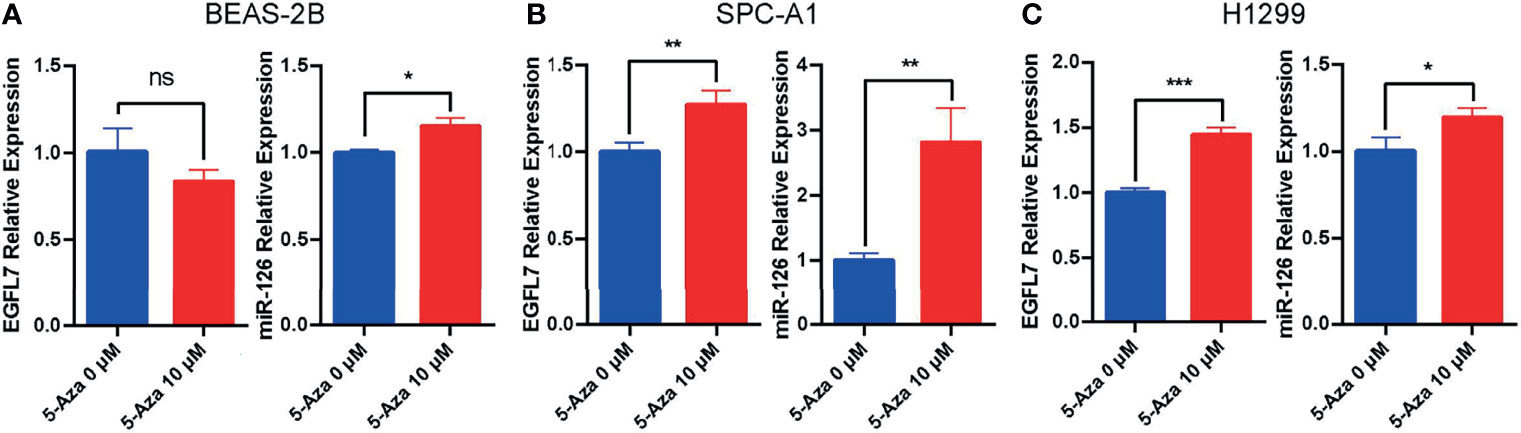
Figure 3 Expression of EGFL7 and miR-126 after demethylating agent 5-Aza treatment in BEAS-2B (A), SPC-A1 (B) and H1299 (C) cell lines. * represents P ≤ 0.05, ** represents P ≤ 0.01, *** represents P ≤ 0.001, ns represents P > 0.05.
Association of EGFL7 and miR-126 Gene SNPs With NSCLC
Recent studies have revealed that single nucleotide polymorphisms (SNPs) in gene regulatory and coding regions could confer risk of lung cancer by regulating the expression of specific genes (26–28), hinting that SNPs in the transcriptional regulatory region and coding region of EGFL7 might also play a role in the regulation of EGFL7 and miR-126 expression, and be further involved in NSCLC susceptibility. Hence, in the current study, we investigated the association between EGFL7 polymorphisms and NSCLC susceptibility in the Han Chinese population.
A total of 502 healthy control samples and 497 NSCLC samples were collected in this study. The clinical characteristics of the study subjects are summarized in Table 1. No significant difference in age or sex was found between the control and NSCLC groups (P = 0.09 and P = 0.13, respectively) (Table 1). In the NSCLC group, 338 patients had adenocarcinoma (AC), 159 patients had squamous cell carcinoma (SCC), 204 patients were Clinical stage I+ II, and 293 patients were Clinical stage III+ IV.

Table 1 Characteristics of the subjects enrolled in the current study.
All three SNPs (rs1332793, rs9411260 and rs2297538) were in Hardy-Weinberg equilibrium (HWE) in the control and NSCLC groups (P > 0.05). The allelic and genotypic distributions of these three SNPs among the healthy control and NSCLC groups are presented in Table 2. Among these SNPs, the allele and genotype distributions of rs2297538 were significantly different between the control and NSCLC groups (P < 0.017). The A allele of rs2297538 might be associated with a decreased risk of NSCLC (P = 0.003; OR = 0.68, 95% CI: 0.52~0.88; Table 2). In addition, the GG genotype frequency of rs2297538 was higher in the NSCLC group than in the control group (P = 0.012; Table 2). We further evaluated the distributions of the alleles and genotypes of these 3 SNPs in different NSCLC pathological types (AC and SCC) and different NSCLC pathological stages (I+II and III+IV). Logistic regression analysis revealed no significant differences in genotypic and allelic distributions of these SNPs among different subgroups (Table S2). The AIC and BIC were calculated to evaluate the best fitting inheritance model (codominant, dominant, recessive, overdominant and log‐additive) for each SNP in this study (39). Only rs2297538 genotypes were found to be associated with NSCLC risk (Table 3). The best fit inheritance model with the lowest AIC and BIC for rs2297538 was dominant, and the A/G and A/A genotypes conferred a greater risk of NSCLC (P = 3.1x10-3; OR = 1.62, 95% CI: 0.65–4.01). These results indicate that EGFL7 and miR-126 play roles in NSCLC pathogenesis.

Table 2 The allelic and genotypic distribution of SNPs in EGFL7 genes among healthy control and NSCLC groups.
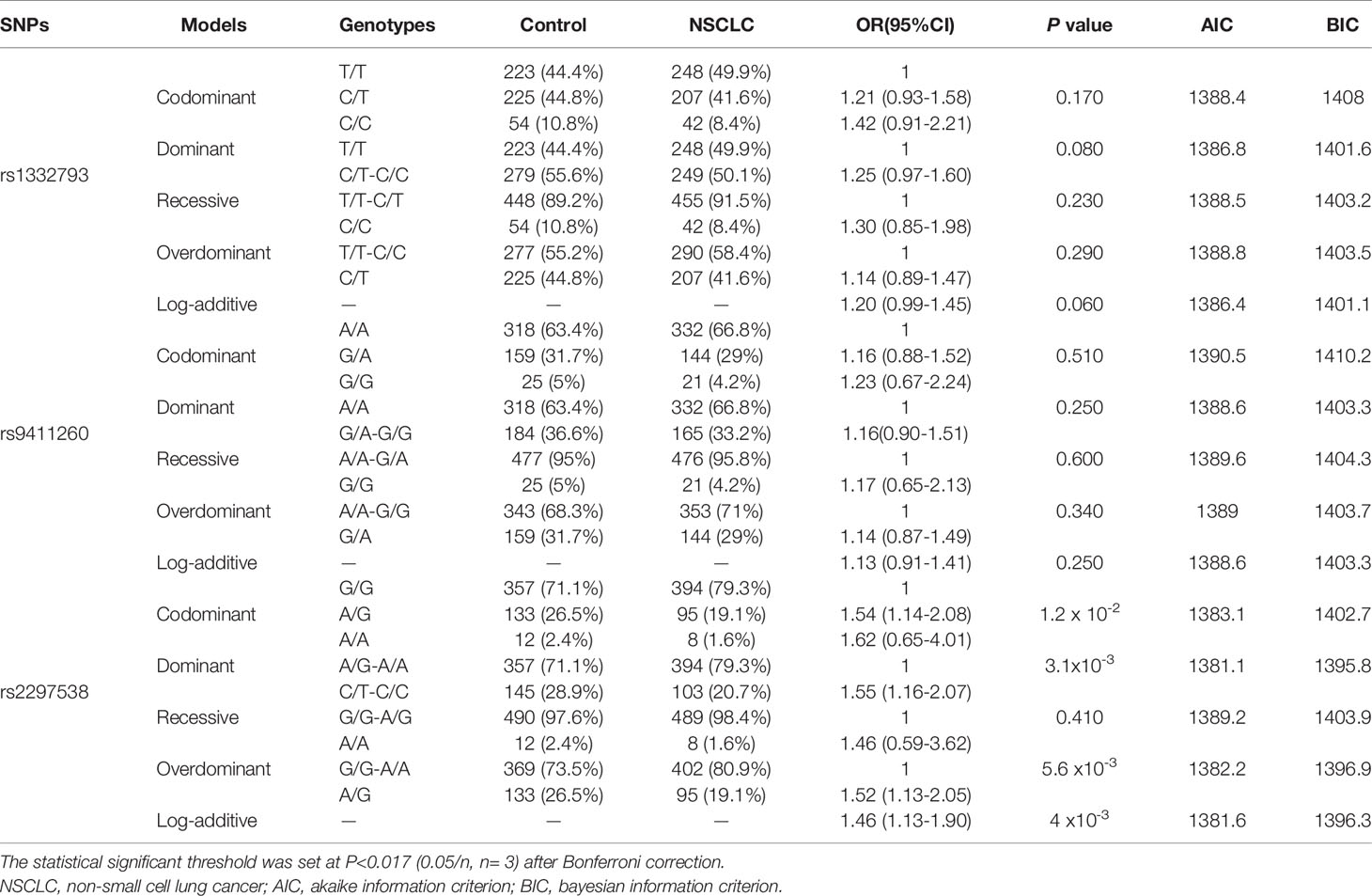
Table 3 Inheritance model analysis of SNPs in the EGFL7 gene between healthy controls and NSCLC patients.
Rs2297538 Is Associated With miR-126 and EGFL7 mRNA Expression
Single nucleotide polymorphisms (SNPs) in gene regulatory and coding regions often confer a risk of lung cancer by affecting gene expression (26–28). To determine whether rs2297538 was related to the expression of nearby genes, we conducted eQTL analysis between this SNP and miR-126 and EGFL7 mRNA expression using qRT–PCR in 46 lung cancer tissues. We compared the mRNA expression of EGFL7 and miR-126 between the risk allele homozygous group [G/G] and the other genotypic groups [A/G + A/A], and we found that the expression of EGFL7 and miR-126 was significantly lower in the G/G group (lung cancer risk allele homozygotes) than in the A/G+A/A group (P = 0.01 and P = 0.002, respectively, Figures 4A, B). These results suggested that reduced expression of EGFL7 and miR-126 might be risk factors for NSCLC and that rs2297538 may confer a risk of NSCLC by regulating EGFL7 and miR-26 expression.
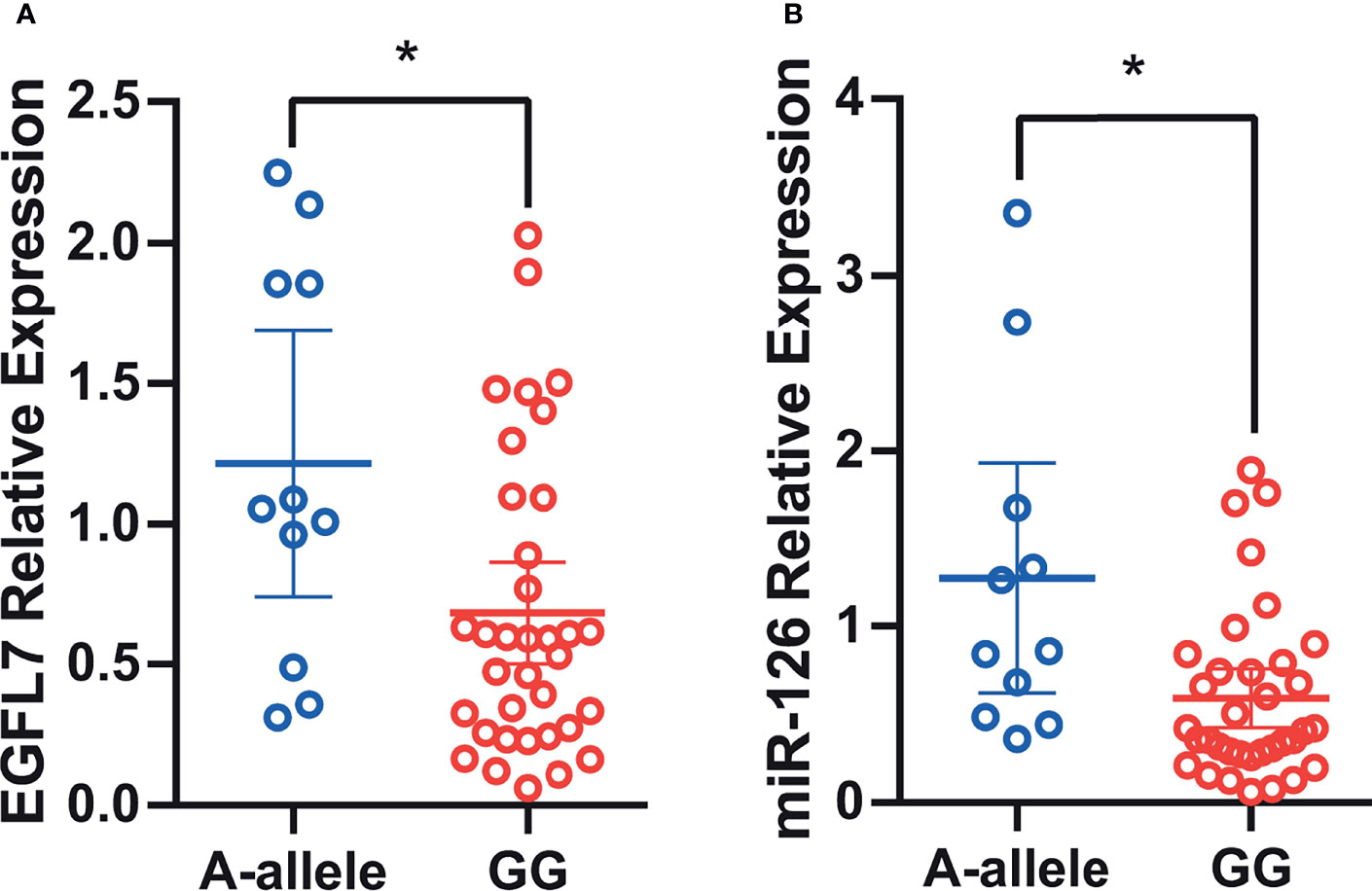
Figure 4 Association of rs2297538 with EGFL7 mRNA (A) and miR-126 (B) expression in lung tumor tissue (N = 46). The values on Y-axis were presented as 2–ΔΔCt. * represents P ≥ 0.05.
Discussion
In the current study, we found that the expression of miR-126 and EGFL7 mRNA was concomitantly downregulated through the methylation of CpG islands in NSCLC and that the eQTL-missense polymorphism of EGFL7, rs2297538 (located at 386 bp 5’ of miR-126), was significantly associated with the risk of NSCLC in the Han Chinese population.
Several studies have reported aberrant expression of EGFL7 and miR-126 in various human cancers (40), suggesting that EGFL7 and miR-126 play a joint role in cancer development. We found that miR-126 and EGFL7 are significantly downregulated in lung cancer tissues, which is consistent with the results of Fan and Yang et al. (13, 20). Our results indicated that aberrant expression of EGFL7 and miR-126 could play an important role in the development of NSCLC. One of the reasons for the association between aberrant expression of EGFL7 and miR-126 and NSCLC could be the change in DNA hypermethylation.
Recently, reduced EGFL7 and miR-126 expression regulated via DNA hypermethylation of the promoter region was observed in ovarian cancer (41), prostate cancer (17) and malignant pleural mesothelioma (16). However, only a few studies have shown the same regulatory mechanism in lung cancer. For example, Watanabe et al. showed miR-126 can be silenced by the DNA methylation of its host gene EGFL7 in lung cancer cell lines (42).Our results also showed that the expression of miR-126 and EGFL7 mRNA in NSCLC is concomitantly regulated, probably by methylation. This is in accordance with findings in other cancer types, including malignant pleural mesothelioma (MPM) (16), breast cancer (13), prostate cancer (17) and lung cancer (42). In 2011, Azhikina et al. reported that the methylation level of the EGFL7 promoter is higher in lung tumors than in healthy lung tissue (43), which further supports our results. We thus further confirmed that the EGFL7 promoter CpG island is highly methylated and thus downregulates EGFL7 and miR-126 in lung tumors.
In addition to hypermethylation of the promoter region, polymorphisms could play an important role in the development of NSCLC. For example, our previous study found that another common SNP, rs4636297, located in the EGFL7 gene region (which is also located in the pri-miR-126 gene) was associated with susceptibility to cervical cancer (44), and a previous study revealed a significant relationship between the EGFL7 3’UTR variant rs1051851 and the overall survival of metastatic colorectal cancer patients (45), suggesting that EGFL7 and miR-126 SNPs might be associated with multiple cancer types. In the current study, we found that rs2297538, which located in exon 7 of EGFL7 and changes valine to isoleucine, was significantly associated with the risk of NSCLC. To our knowledge, this is the first study to demonstrate an association between EGFL7 and miR-126 genetic variants and NSCLC. Notably, in a recent study, Duan et al. found that individuals carrying rs2297538 homozygous GG had lower leukocyte mitochondrial DNA copy numbers in the polycyclic aromatic hydrocarbon exposure group (46). As a lower mitochondrial DNA copy number could predict an increased risk of cancer induced by polycyclic aromatic hydrocarbon exposure (46) and miR-126 has been reported to regulate mitochondrial energy metabolism (47, 48), it is reasonable to hypothesize that rs2297538 may confer a risk of NSCLC by regulating miR-126 expression and further affecting mitochondrial functions. However, because the function of rs2297538 is currently unknown, we need to expand the sample size for further validation and explore its role in miR-126 and EGFL7 mRNA expression as well as mitochondrial function.
In the current study, we firstly found that rs2297538 was associated with the expression of miR-126 and EGFL7 mRNA in lung cancer tissues. As majority of previous studies focus on transcriptional regulation of EGFL7/miR-126 by DNA methylation in multiple tumor cell lines, our brand new finding in this article suggested that EGFL7 and miR-126 mRNA levels could be regulated via DNA methylation as well as single nucleotide polymorphisms. Moreover, the risk rs2297538 G allele might be correlated with lower expression of miR-126 and EGFL7 mRNA in lung cancer tissues, which indicated that reduced expression of miR-126 and EGFL7 might be risk factors for NSCLC. However, increased expression of EGFL7 has been reported in certain epithelial cancers (13). This may be somewhat surprising, given that elevated expression of EGFL7 may play a crucial role in cancer biology by modulating tumor angiogenesis (40). One of the reasons for the discrepancy might be the different types of cancer, as the lung is inherently a highly vascularized tissue and maintains high levels of EGFL7. In addition, decreased expression of EGFL7 may confer a risk of NSCLC via other biological mechanisms, such as altering the mitochondrial function of lung cancer cells. Considering that heterogeneity plays a key role in cancer management, we also performed the EGFL7 expression assay at single cells’ level using the human protein atlas data base (49, 50). Our results revealed that EGFL7 expressed mainly in endothelial cells and alveolar type 2 cells. In 2016, Pinte et al. reported that EGFL7 was able to repress endothelial cells activation (51). Their finding indicated EGFL7 could play a role in tumoral angiogenesis and tumor progression. In addition, alveolar type 2 cells were reported to play an active role in enhancing alveolar fluid clearance and reducing lung inflammation. Thus, the alveolar type 2 cells therapy was used and it was reported to have great potential effects for acute lung injury/acute respiratory distress syndrome in several preclinical studies (52). Based on these evidences, altered expression of EGFL7 in endothelial cells and alveolar type 2 cells could be associated with the lung cancer susceptibility.
Notably, there are several limitations in the current study, and we are cautious in the interpretation of the present results. Our data indicated that methylation levels of several CG loci in EGFL7 were significantly higher in the lung cancer samples than in the normal samples and 5-Aza-CdR treatment of NSCLC cell lines could result in up-regulation of EGFL7 mRNA expression. However, the classic locus-specific methylation experiments to explore the methylation frequency of CpG-islands EGFL7 in NSCLC cell lines and the methylation changes in different pathological types and stages should be carried out in the future. In addition, even though our results indicated that expression of miR-126 and EGFL7 mRNA are significantly reduced in NSCLC tissues compared with adjacent normal tissues, the protein expression difference of EGFL7 also needs to be investigated.
In summary, we confirmed that the expression of EGFL7 and miR-126 in NSCLC can be concomitantly downregulated through methylation of CpG islands. We also report that EGFL7 and miR-126 are correlated with NSCLC risk in the Han Chinese population, and our results suggest that rs2297538 may confer a risk of NSCLC by altering the gene expression of EGFL7 and miR-126. Together, these results provide new insights into the pathogenesis of NSCLC.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics Statement
The studies involving human participants were reviewed and approved by The Ethics Committee of the Third Affiliated Hospital of Kunming Medical University. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
YY, LS, and WL designed the study and interpreted the results. WL, YZ, FH, CL, and SL conducted the SNP genotyping, the primary functional assays, including qRT-PCR, cell line experiments, and analysis of those data. QM and YL contributed to collection of clinical samples. YY and LS drafted the manuscript. All authors contributed to the final version of the paper.
Funding
This work was supported by grants from the Fundamental Research Funds for the Central Universities and the PUMC Youth Fund (3332021071), the National Science Foundation for Young Scientists of China (82103190), the Special Funds for High-level Healthy Talents of Yunnan Province (L-201615, H-2018014 and D-2018037). The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2022.772405/full#supplementary-material
Supplementary Figure 1 | The relative expression of EGFL7 in lung cancer tissue and normal tissue from GEPIA database. N, normal; T, tumor; AC, adenocarcinoma; SCC, squamous.
Supplementary Figure 2 | The methylation status of EGFL7 in lung cancer tumor tissues and normal tissues from SurvivalMeth database. AC, adenocarcinoma; SCC, squamous.
References
1. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin (2021) 71(3):209–49. doi: 10.3322/caac.21660
2. Chang JT, Lee YM, Huang RS. The Impact of the Cancer Genome Atlas on Lung Cancer. Transl Res (2015) 166:568–85. doi: 10.1016/j.trsl.2015.08.001
3. Mucci LA, Hjelmborg JB, Harris JR, Czene K, Havelick DJ, Scheike T, et al. Familial Risk and Heritability of Cancer Among Twins in Nordic Countries. Jama (2016) 315:68–76. doi: 10.1001/jama.2015.17703
4. Wang J, Liu Q, Yuan S, Xie W, Liu Y, Xiang Y, et al. Genetic Predisposition to Lung Cancer: Comprehensive Literature Integration, Meta-Analysis, and Multiple Evidence Assessment of Candidate-Gene Association Studies. Sci Rep (2017) 7:8371. doi: 10.1038/s41598-017-07737-0
5. Chen R, Li WX, Sun Y, Duan Y, Li Q, Zhang AX, et al. Comprehensive Analysis of lncRNA and mRNA Expression Profiles in Lung Cancer. Clin Lab (2017) 63:313–20. doi: 10.7754/Clin.Lab.2016.160812
6. Long T, Liu Z, Zhou X, Yu S, Tian H, Bao Y. Identification of Differentially Expressed Genes and Enriched Pathways in Lung Cancer Using Bioinformatics Analysis. Mol Med Rep (2019) 19:2029–40. doi: 10.3892/mmr.2019.9878
7. Wang K, Chen R, Feng Z, Zhu YM, Sun XX, Huang W, et al. Identification of Differentially Expressed Genes in Non-Small Cell Lung Cancer. Aging (Albany NY) (2019) 11:11170–85. doi: 10.18632/aging.102521
8. Mckay JD, Hung RJ, Han Y, Zong X, Carreras-Torres R, Christiani DC, et al. Large-Scale Association Analysis Identifies New Lung Cancer Susceptibility Loci and Heterogeneity in Genetic Susceptibility Across Histological Subtypes. Nat Genet (2017) 49:1126–32. doi: 10.1038/ng.3892
9. Bosse Y, Amos CI. A Decade of GWAS Results in Lung Cancer. Cancer Epidemiol Biomarkers Prev (2018) 27:363–79. doi: 10.1158/1055-9965.EPI-16-0794
10. Dai J, Lv J, Zhu M, Wang Y, Qin N, Ma H, et al. Identification of Risk Loci and a Polygenic Risk Score for Lung Cancer: A Large-Scale Prospective Cohort Study in Chinese Populations. Lancet Respir Med (2019) 7:881–91. doi: 10.1016/S2213-2600(19)30144-4
11. Nichol D, Stuhlmann H. EGFL7: A Unique Angiogenic Signaling Factor in Vascular Development and Disease. Blood (2012) 119:1345–52. doi: 10.1182/blood-2011-10-322446
12. Bambino K, Lacko LA, Hajjar KA, Stuhlmann H. Epidermal Growth Factor-Like Domain 7 Is a Marker of the Endothelial Lineage and Active Angiogenesis. Genesis (2014) 52:657–70. doi: 10.1002/dvg.22781
13. Fan C, Yang LY, Wu F, Tao YM, Liu LS, Zhang JF, et al. The Expression of Egfl7 in Human Normal Tissues and Epithelial Tumors. Int J Biol Markers (2013) 28:71–83. doi: 10.5301/JBM.2013.10568
14. Campagnolo L, Telesca C, Massimiani M, Stuhlmann H, Angelico M, Lenci I, et al. Different Expression of VEGF and EGFL7 in Human Hepatocellular Carcinoma. Dig Liver Dis (2016) 48:76–80. doi: 10.1016/j.dld.2015.09.019
15. Papaioannou D, Shen C, Nicolet D, Mcneil B, Bill M, Karunasiri M, et al. Prognostic and Biological Significance of the Proangiogenic Factor EGFL7 in Acute Myeloid Leukemia. Proc Natl Acad Sci USA (2017) 114:E4641–7. doi: 10.1073/pnas.1703142114
16. Andersen M, Trapani D, Ravn J, Sørensen JB, Andersen CB, Grauslund M, et al. Methylation-Associated Silencing of microRNA-126 and its Host Gene EGFL7 in Malignant Pleural Mesothelioma. Anticancer Res (2015) 35:6223–9.
17. Saito Y, Friedman JM, Chihara Y, Egger G, Chuang JC, Liang G. Epigenetic Therapy Upregulates the Tumor Suppressor microRNA-126 and its Host Gene EGFL7 in Human Cancer Cells. Biochem Biophys Res Commun (2009) 379:726–31. doi: 10.1016/j.bbrc.2008.12.098
18. Miao Y, Lu J, Fan B, Sun L. MicroRNA-126-5p Inhibits the Migration of Breast Cancer Cells by Directly Targeting Cnot7. Technol Cancer Res Treat (2020) 19:1533033820977545. doi: 10.1177/1533033820977545
19. Wu DM, Wen X, Han XR, Wang S, Wang YJ, Shen M, et al. Bone Marrow Mesenchymal Stem Cell-Derived Exosomal MicroRNA-126-3p Inhibits Pancreatic Cancer Development by Targeting Adam9. Mol Ther Nucleic Acids (2019) 16:229–45. doi: 10.1016/j.omtn.2019.02.022
20. Yang J, Lan H, Huang X, Liu B, Tong Y. MicroRNA-126 Inhibits Tumor Cell Growth and its Expression Level Correlates With Poor Survival in Non-Small Cell Lung Cancer Patients. PloS One (2012) 7:e42978. doi: 10.1371/journal.pone.0042978
21. Liu R, Zhang YS, Zhang S, Cheng ZM, Yu JL, Zhou S, et al. MiR-126-3p Suppresses the Growth, Migration and Invasion of NSCLC via Targeting CCR1. Eur Rev Med Pharmacol Sci (2019) 23:679–89. doi: 10.26355/eurrev_201901_16881
22. Song L, Li D, Gu Y, Wen ZM, Jie J, Zhao D, et al. MicroRNA-126 Targeting PIK3R2 Inhibits NSCLC A549 Cell Proliferation, Migration, and Invasion by Regulation of PTEN/PI3K/AKT Pathway. Clin Lung Cancer (2016) 17:e65–75. doi: 10.1016/j.cllc.2016.03.012
23. Crawford M, Brawner E, Batte K, Yu L, Hunter MG, Otterson GA, et al. MicroRNA-126 Inhibits Invasion in Non-Small Cell Lung Carcinoma Cell Lines. Biochem Biophys Res Commun (2008) 373:607–12. doi: 10.1016/j.bbrc.2008.06.090
24. Zheng W, Zhou Y, Lu J, Xu H, Lei L, Chen C, et al. The Prognostic Value of miR-126 Expression in Non-Small-Cell Lung Cancer: A Meta-Analysis. Cancer Cell Int (2017) 17:71. doi: 10.1186/s12935-017-0440-8
25. Chen Q, Chen S, Zhao J, Zhou Y, Xu L. MicroRNA-126: A New and Promising Player in Lung Cancer. Oncol Lett (2021) 21:35. doi: 10.3892/ol.2020.12296
26. Zhu M, Wang Y, Wang C, Shen W, Liu J, Geng L, et al. The eQTL-Missense Polymorphisms of APOBEC3H Are Associated With Lung Cancer Risk in a Han Chinese Population. Sci Rep (2015) 5:14969. doi: 10.1038/srep14969
27. Shao L, Zuo X, Yang Y, Zhang Y, Yang N, Shen B, et al. The Inherited Variations of a P53-Responsive Enhancer in 13q12.12 Confer Lung Cancer Risk by Attenuating TNFRSF19 Expression. Genome Biol (2019) 20:103. doi: 10.1186/s13059-019-1696-1
28. Wang Y, Ma R, Liu B, Kong J, Lin H, Yu X, et al. SNP Rs17079281 Decreases Lung Cancer Risk Through Creating an YY1-Binding Site to Suppress DCBLD1 Expression. Oncogene (2020) 39:4092–102. doi: 10.1038/s41388-020-1278-4
29. Detterbeck FC, Chansky K, Groome P, Bolejack V, Crowley J, Shemanski L, et al. The IASLC Lung Cancer Staging Project: Methodology and Validation Used in the Development of Proposals for Revision of the Stage Classification of NSCLC in the Forthcoming (Eighth) Edition of the TNM Classification of Lung Cancer. J Thorac Oncol (2016) 11:1433–46. doi: 10.1016/j.jtho.2016.06.028
30. Christman JK. 5-Azacytidine and 5-aza-2’-Deoxycytidine as Inhibitors of DNA Methylation: Mechanistic Studies and Their Implications for Cancer Therapy. Oncogene (2002) 21:5483–95. doi: 10.1038/sj.onc.1205699
31. Li XY, Wu JZ, Cao HX, Ma R, Wu JQ, Zhong YJ, et al. Blockade of DNA Methylation Enhances the Therapeutic Effect of Gefitinib in Non-Small Cell Lung Cancer Cells. Oncol Rep (2013) 29:1975–82. doi: 10.3892/or.2013.2298
32. Nakamura M, Nishikawa J, Saito M, Sakai K, Sasaki S, Hashimoto S, et al. Decitabine Inhibits Tumor Cell Proliferation and Up-Regulates E-Cadherin Expression in Epstein-Barr Virus-Associated Gastric Cancer. J Med Virol (2017) 89:508–17. doi: 10.1002/jmv.24634
33. Harris TA, Yamakuchi M, Kondo M, Oettgen P, Lowenstein CJ. Ets-1 and Ets-2 Regulate the Expression of microRNA-126 in Endothelial Cells. Arterioscler Thromb Vasc Biol (2010) 30:1990–7. doi: 10.1161/ATVBAHA.110.211706
34. Fornes O, Castro-Mondragon JA, Khan A, van der Lee R, Zhang X, Richmond PA, et al. JASPAR 2020: Update of the Open-Access Database of Transcription Factor Binding Profiles. Nucleic Acids Res (2020) 48:D87–D92. doi: 10.1093/nar/gkz1001
35. Mclaren W, Gil L, Hunt SE, Riat HS, Ritchie GR, Thormann A, et al. The Ensembl Variant Effect Predictor. Genome Biol (2016) 17:122. doi: 10.1186/s13059-016-0974-4
36. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: A Web Server for Cancer and Normal Gene Expression Profiling and Interactive Analyses. Nucleic Acids Res (2017) 45:W98–W102. doi: 10.1093/nar/gkx247
37. Zhang C, Zhao N, Zhang X, Xiao J, Li J, Lv D, et al. SurvivalMeth: A Web Server to Investigate the Effect of DNA Methylation-Related Functional Elements on Prognosis. Brief Bioinform (2021) 22:bbaa162. doi: 10.1093/bib/bbaa162
38. Shi YY, He L. SHEsis, a Powerful Software Platform for Analyses of Linkage Disequilibrium, Haplotype Construction, and Genetic Association at Polymorphism Loci. Cell Res (2005) 15:97–8. doi: 10.1038/sj.cr.7290272
39. Solé X, Guinó E, Valls J, Iniesta R, Moreno V. SNPStats: A Web Tool for the Analysis of Association Studies. Bioinformatics (2006) 22:1928–9. doi: 10.1093/bioinformatics/btl268
40. Hong G, Kuek V, Shi J, Zhou L, Han X, He W, et al. EGFL7: Master Regulator of Cancer Pathogenesis, Angiogenesis and an Emerging Mediator of Bone Homeostasis. J Cell Physiol (2018) 233:8526–37. doi: 10.1002/jcp.26792
41. Rattanapan Y, Korkiatsakul V, Kongruang A, Chareonsirisuthigul T, Rerkamnuaychoke B, Wongkularb A, et al. EGFL7 and RASSF1 Promoter Hypermethylation in Epithelial Ovarian Cancer. Cancer Genet (2018) 224-225:37–40. doi: 10.1016/j.cancergen.2018.04.117
42. Watanabe K, Emoto N, Hamano E, Sunohara M, Kawakami M, Kage H, et al. Genome Structure-Based Screening Identified Epigenetically Silenced microRNA Associated With Invasiveness in Non-Small-Cell Lung Cancer. Int J Cancer (2012) 130:2580–90. doi: 10.1002/ijc.26254
43. Azhikina T, Kozlova A, Skvortsov T, Sverdlov E. Heterogeneity and Degree of TIMP4, GATA4, SOX18, and EGFL7 Gene Promoter Methylation in Non-Small Cell Lung Cancer and Surrounding Tissues. Cancer Genet (2011) 204:492–500. doi: 10.1016/j.cancergen.2011.07.010
44. Yan Z, Zhou Z, Li C, Yang X, Yang L, Dai S, et al. Polymorphisms in miRNA Genes Play Roles in the Initiation and Development of Cervical Cancer. J Cancer (2019) 10:4747–53. doi: 10.7150/jca.33486
45. Hansen TF, Andersen RF, Olsen DA, Sørensen FB, Jakobsen A. Prognostic Importance of Circulating Epidermal Growth Factor-Like Domain 7 in Patients With Metastatic Colorectal Cancer Treated With Chemotherapy and Bevacizumab. Sci Rep (2017) 7:2388. doi: 10.1038/s41598-017-02538-x
46. Duan X, Yang Y, Zhang H, Liu B, Wei W, Wang L, et al. Polycyclic Aromatic Hydrocarbon Exposure, miRNA Genetic Variations, and Associated Leukocyte Mitochondrial DNA Copy Number: A Cross-Sectional Study in China. Chemosphere (2020) 246:125773. doi: 10.1016/j.chemosphere.2019.125773
47. Tomasetti M, Nocchi L, Staffolani S, Manzella N, Amati M, Goodwin J, et al. MicroRNA-126 Suppresses Mesothelioma Malignancy by Targeting IRS1 and Interfering With the Mitochondrial Function. Antioxid Redox Signal (2014) 21:2109–25. doi: 10.1089/ars.2013.5215
48. Lima Queiroz A, Zhang B, Comstock DE, Hao Y, Eriksson M, Hydbring P, et al. miR-126-5p Targets Malate Dehydrogenase 1 in Non-Small Cell Lung Carcinomas. Biochem Biophys Res Commun (2018) 499:314–20. doi: 10.1016/j.bbrc.2018.03.154
49. Vieira Braga FA, Kar G, Berg M, Carpaij OA, Polanski K, Simon LM, et al. A Cellular Census of Human Lungs Identifies Novel Cell States in Health and in Asthma. Nat Med (2019) 25:1153–63. doi: 10.1038/s41591-019-0468-5
50. Karlsson M, Zhang C, Méar L, Zhong W, Digre A, Katona B, et al. A Single-Cell Type Transcriptomics Map of Human Tissues. Sci Adv (2021) 7:eabh2169. doi: 10.1126/sciadv.abh2169
51. Pinte S, Caetano B, Le Bras A, Havet C, Villain G, Dernayka R, et al. Endothelial Cell Activation Is Regulated by Epidermal Growth Factor-Like Domain 7 (Egfl7) During Inflammation. J Biol Chem (2016) 291:24017–28. doi: 10.1074/jbc.M116.731331
Keywords: EGFL7, miR-126, NSCLC, DNA methylation, SNPs
Citation: Liu W, Zhang Y, Huang F, Ma Q, Li C, Liu S, Liang Y, Shi L and Yao Y (2022) The Polymorphism and Expression of EGFL7 and miR-126 Are Associated With NSCLC Susceptibility. Front. Oncol. 12:772405. doi: 10.3389/fonc.2022.772405
Received: 08 September 2021; Accepted: 21 March 2022;
Published: 14 April 2022.
Edited by:
Parvin Mehdipour, Tehran University of Medical Sciences, IranReviewed by:
Soudabeh Fallah, Iran University of Medical Sciences, IranMatteo Canale, Scientific Institute of Romagna for the Study and Treatment of Tumors (IRCCS), Italy
Copyright © 2022 Liu, Zhang, Huang, Ma, Li, Liu, Liang, Shi and Yao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Li Shi, shili.imb@gmail.com; Yufeng Yao, leoyyf@gmail.com; yufeng_yao@imbcams.com.cn
†These authors have contributed equally to this work and share first authorship