cFOS expression in the prefrontal cortex correlates with altered cerebral metabolism in developing germ-free mice
 Trinity Pate
Trinity Pate  Daniel C. Anthony
Daniel C. Anthony Daniel E. Radford-Smith
Daniel E. Radford-Smith- Department of Pharmacology, University of Oxford, Oxford, United Kingdom
Introduction: The microbiota plays a critical role in modulating various aspects of host physiology, particularly through the microbiota-gut-brain (MGB) axis. However, the mechanisms that transduce and affect gut-to-brain communication are still not well understood. Recent studies have demonstrated that dysbiosis of the microbiome is associated with anxiety and depressive symptoms, which are common complications of metabolic syndrome. Germ-free (GF) animal models offer a valuable tool for studying the causal effects of microbiota on the host.
Methods: We employed gene expression and nuclear magnetic resonance (NMR)-based metabolomic techniques to investigate the relationships between brain plasticity and immune gene expression, peripheral immunity, and cerebral and liver metabolism in GF and specific pathogen-free (SPF) mice.
Results: Our principal findings revealed that brain acetate (p = 0.012) was significantly reduced in GF relative to SPF mice, whereas glutamate (p = 0.0013), glutamine (p = 0.0006), and N-acetyl aspartate (p = 0.0046) metabolites were increased. Notably, cFOS mRNA expression, which was significantly decreased in the prefrontal cortex of GF mice relative to SPF mice (p = 0.044), correlated with the abundance of a number of key brain metabolites altered by the GF phenotype, including glutamate and glutamine.
Discussion: These results highlight the connection between the GF phenotype, altered brain metabolism, and immediate-early gene expression. The study provides insight into potential mechanisms by which microbiota can regulate neurotransmission through modulation of the host’s brain and liver metabolome, which may have implications for stress-related psychiatric disorders such as anxiety.
Introduction
The gut microbiome is a complex and dense microbial network that regulates many aspects of human physiology and behavior. In adulthood, it is estimated that there are over 1,000 different species comprising approximately 1014 organisms, although inter-individual variation is significant (Qin et al., 2010).
This postnatal colonization and persistence of microbiota throughout life is a normal part of mammalian development, with microbiota metabolism contributing to the function of several host systems and processes including host inflammatory responses, energy metabolism, and gastrointestinal absorption (Cebra, 1999; Velagapudi et al., 2010; Semova et al., 2012). The gut microbiota digest host-indigestible dietary fibers, producing metabolic by-products which are absorbed and utilized by the host such as short-chain fatty acids (SCFAs), which regulate several aspects of host physiology (van der Hee and Wells, 2021). This exemplifies the symbiotic relationship between bacteria and their host organisms, and highlights the critical role of microbiota in host metabolism.
Gut microbiota metabolism has also been shown to modify brain chemistry and behavior via the bidirectional MGB axis (Cryan et al., 2019). For instance, it has been demonstrated that microbiota are required for normal social development in mice (Barreau et al., 2004; Zheng et al., 2016). Metabolic profiling studies have even shown that the maternal microbiome can modulate fetal neurodevelopment in mice (Bravo et al., 2011). Rodent models of maternal separation demonstrate how early life stress modifies the composition of the gut flora (Sobko et al., 2006) and can result in long-term changes in gut physiology, including the permeability of the epithelial barrier which can trigger exaggerated immune responses (Nicolas et al., 2022). Gut dysbiosis also affects brain chemistry and behavior. Germ-free mice colonized by faucal microbiota transplant with “major depressive disorder (MDD) microbiota,” but not healthy control microbiota, had disturbed serum and hippocampal metabolomes and begin displaying anxiety- and depressive-like behaviors (Schéle et al., 2013).
Germ-free (GF) mice lack microbiota from birth and have proved essential in understanding the casual impact of microbiota on host physiology. In the brain, GF mice display an immature microglial phenotype with altered morphological characteristics and gene expression profiles, which can be restored by engraftment of a complex gut microbiota, or short-chain fatty acid (SCFA) supplementation (Erny et al., 2015). Microglia are the resident macrophages of the central nervous system (CNS), responding to brain infections and coordinating downstream immune reactions (Borst et al., 2021), whilst also regulating synaptic pruning and shaping neural circuits in the developing brain (Schafer et al., 2012). Their deficiency in GF mice may partly explain their altered brain chemistry and immune activity.
Recent reviews have introduced the concept of the “gut-liver-brain axis,” which emphasizes the central role of the liver in peripheral metabolism and immune regulation (Tripathi et al., 2018; Brescia and Rescigno, 2021; Kronsten et al., 2022). Extensive inter-organ communication exists between the liver and the gut, particularly at the level of circulating metabolites, through the portal vein. Moreover, hepatic encephalopathy is a severe yet common complication of end-stage liver disease, whereby neurotoxic metabolites (some gut-derived) are not able to metabolized by the liver. Despite the increasing awareness of the gut-liver-brain axis in health and disease, the liver and brain metabolome have not been jointly analyzed in GF and SPF mice.
Current understanding of GF physiology is still relatively limited. Most studies have focused on gene expression, far fewer have identified metabolomic changes, and to our knowledge, no study has combined both techniques in the same animal and drawn inferences between them. Global metabolite profiling, otherwise known as untargeted metabolomics, can be used to explore metabolic differences in phenotypes as a consequence of the microbiome. Exploring the gene expression changes in conjunction with metabolomic changes could identify novel pathways relating to the microbial modulation of brain function. Therefore, the aim of the current study was to interrogate the contribution of the gut microbiota to shaping the relationship between neuroplasticity gene expression in the prefrontal cortex (PFC) and cerebral metabolite profiling. This is supported by previous reports of altered metabolism in the cerebral cortex of GF mice (Lai et al., 2021), and the relationship between early life probiotic supplementation, cerebral metabolite levels, and altered PFC gene expression (Radford-Smith et al., 2022). Accordingly, we hypothesized that GF status would affect cerebral metabolite levels through altered neurodevelopmental trajectories, with the understanding that the maternal generation was also germ free and microbial metabolites are thought to play a role in normal brain development (Vuong et al., 2020). We also expected to discover mRNA-metabolite correlations in SPF mice that dissociated in GF mice which would aid our understanding of the relationship between microbiota, metabolites, and brain function. An examination of the biochemical pathways perturbed in GF mice will provide further insights into the role of the gut microbiota in brain metabolism and may be indicative of future therapeutic avenues to pursue.
Materials and methods
Animals
C57Bl/6 mice were killed by cervical dislocation and the whole brain and liver were immediately dissected from juvenile (3-week-old) female GF and SPF was animals and snap frozen in isopentane. Mice were interrogated for liver and brain qPCR and metabolomic analyses (Figure 1). A total of 2 cohorts of mice were used in this experiment. For the liver gene expression analysis, 5 SPF and 7 GF mice were used. For the liver and brain metabolomics analysis, and brain qPCR analysis, a separate cohort of 6 SPF and 6 GF mice were used.
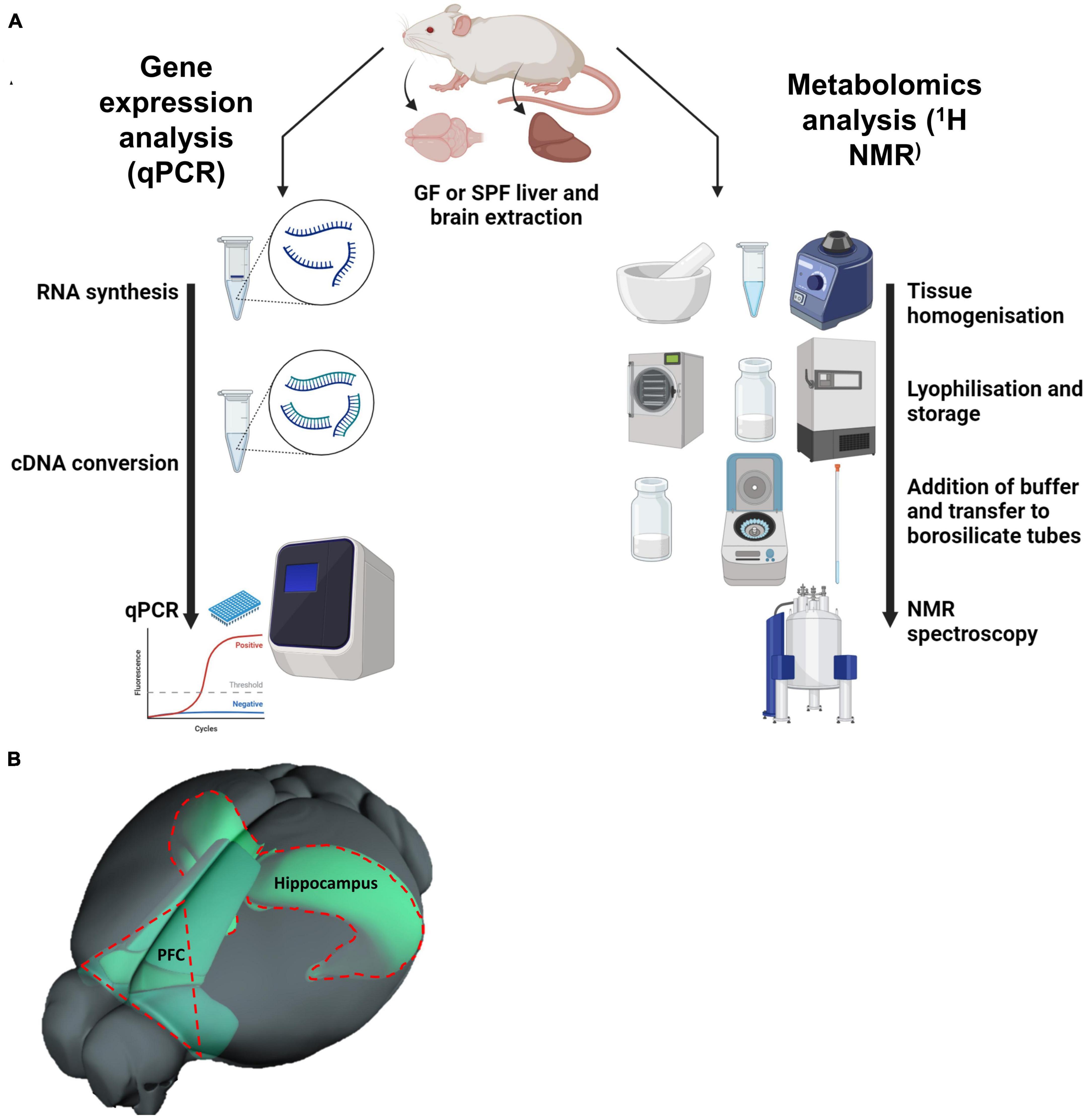
Figure 1. (A) Diagram depicting the methodology of the project: quantitative PCR (qPCR) and 1D 1H nuclear magnetic resonance (NMR) spectroscopy. (B) Illustration of prefrontal cortex within the mouse brain, relative to the hippocampus. (B) Is adapted from the Allen Mouse Brain Connectivity Atlas.
Gene expression
RNA extraction and cDNA conversion
The prefrontal cortex was dissected from the whole brain by making a V-shaped incision at the most anterior part of the brain, avoiding the olfactory bulb and white matter. The Qiagen RNeasy Mini Kit (Qiagen Ltd., Manchester, UK) was utilized for RNA extraction, using methods described in detail elsewhere (Radford-Smith et al., 2022). A total of 1,000 nanograms of RNA was converted to cDNA using the high-capacity cDNA reverse transcription kit (Thermo Fisher, Loughborough, UK). To assess suitability for cDNA conversion, both the total RNA and RNA purity were assessed using a NanoDrop1000 (Thermo Fisher, Loughborough, UK).
Quantitative (q)PCR
Quantitative PCR was conducted using a LightCycler® 480 instrument (Roche Diagnostics, West Sussex, UK) with SYBR green master mix (Primerdesign, Hampshire, UK). 25 ng cDNA was used per 10 uL reaction. Primers (Table 1) were obtained from Primerdesign, Merck Life Science LTD (Dorset, UK), or Bio-Rad Laboratories (Hertfordshire, UK) and used at a working concentration of 300 nM. GAPDH was used as the reference gene in both the brain and liver tissue. Melt curves were analyzed to confirm that a singular PCR product had been generated for each reaction. Samples were run in duplicate and normalized using 2–Δ Ct for each sample, where ΔCt = (Cttarget gene–Ctreference gene), and then analyzed data as fold change in gene expression relative to the control group.

Table 1. List of primers for qPCR.
Metabolic profiling
Sample preparation
Samples were prepared in small batches to minimize variation, using an optimized protocol based on previously published methods (Waters et al., 2001). Between 115 and 135 mg of fresh liver or brain (left hemisphere of cerebrum) were homogenized with a pestle and mortar on dry ice; equipment was dry-wiped between samples to prevent cross-contamination. Samples were diluted eight-fold (μL/mg) in 50% acetonitrile in pure water and further homogenized by being briefly vortexed. Samples were then centrifuged at 5,060 × g for 5 min at 4°C and 750 μL of each sample was aspirated into a fresh Eppendorf. Samples were then lyophilized and stored at −80°C until the day of NMR analysis where they were resuspended by vortex in 600 μL of 75 mM sodium phosphate buffer (5:1 disodium phosphate [Na2HPO4] and monosodium phosphate [NaH2PO4] in 100% D2O, pH = 7.4) and centrifuged at 2,500 × g for 5 min at 4°C to remove any particulate matter. Lastly, samples were transferred to a 5 mm borosilicate tube using a glass Pasteur pipette, ready for NMR analysis.
1D 1H nuclear magnetic resonance (NMR) spectroscopy and data processing
1H NMR spectra were obtained using a 700-MHz Bruker AVII spectrometer, operating at a 16.4T equipped with a 1H (13C/15N) TCI cryoprobe. Sample temperature was maintained at 298K, and spectra were acquired using a 1D NOESY experiment (brain) and CPMG experiment (liver) as described previously (Radford-Smith et al., 2022). Topspin 2.1 (Bruker, Germany) was used for data processing: to manually phase the spectra, correct baselines, and reference the chemical shifts to the lactate CH3 doublet resonance at δ = 1.33 ppm. All spectra of the same tissue were overlayed and automatically binned with 0.02 ppm-sized bins, although some compounds were manually binned. Each bin was normalized to the sum of all integrals in the spectrum of each sample and data exported to R v3.3.1 and Microsoft Excel for statistical analysis and metabolite identification. Metabolites were assigned by comparing spectral peaks to the Human Metabolome Database (Wishart et al., 2018) and by spiking in known compounds.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 6 and R v3.3.1 (R Foundation for Statistical Computing). An in-house R script was used for principal component analysis (PCA) of pareto-scaled data to initially visualize and inspect the data. PCA loadings were plotted to identify key metabolites in the liver and brain. Individual metabolites were then run through the following analyses.
Initially, Grubb’s test for outliers was performed on each group for all qPCR and metabolomic data and statistically significant outliers (p < 0.05) were removed from downstream analyses. Normality of data was assessed using the Kolmogorov–Smirnov (KS) test: if assumed (p < 0.1), a parametric independent samples t-test was conducted, if not assumed (p < 0.1, or the test concluded N was too small due to exclusion of anomalous results) then the non-parametric Mann–Whitney U test was conducted. For parametric data, the independent samples t-test was followed by an F-test to compare variances (p < 0.05) and if significant, the t-test was repeated using Welch’s correction to account for the unequal variances between the populations. Results from t-tests and Mann–Whitney U tests were considered statistically significant if p < 0.05 and trending if p < 0.1. Data were then visualized as boxplots where the upper and lower bounds of the box denote the interquartile range, the line within the box as the median, and whiskers encompassing the full range of values.
Correlations between brain qPCR and all metabolome data were performed using the Spearman rank correlation. Since normality could not be assumed due to too low an N number for correlation analysis, a non-parametric test was used. False discovery rate (FDR) analysis was carried out to correct for multiple testing and significant correlations (q < 0.05) were visualized as scatterplots.
Results
Germ-free mice have altered cerebral and liver metabolite profiles
To study the impact of the microbiome on mouse metabolite profiles, untargeted proton NMR spectroscopy was performed on liver and cerebrum tissue extracts of female, 3-week-old GF and SPF mice. In both tissues, there was significant variation between the two groups, visualized in the PCA scores plots (Figures 2A, C) and NMR spectra (Figure 3). PCA loadings plots (Figures 2B, D) were then used to determine which metabolites were contributing most to the group variation (Table 2).

Figure 2. PCA score plots comparing the GF and SPF liver and brain metabolome. Purple data points are from GF (n = 6) and green from SPF (n = 6) mice. (A) Brain PCA plot; P1 = 71%, P2 = 11% and (B) corresponding loadings plot. (C) Liver PCA plot; P1 = 43%, P2 = 22% and (D) corresponding loadings plot.
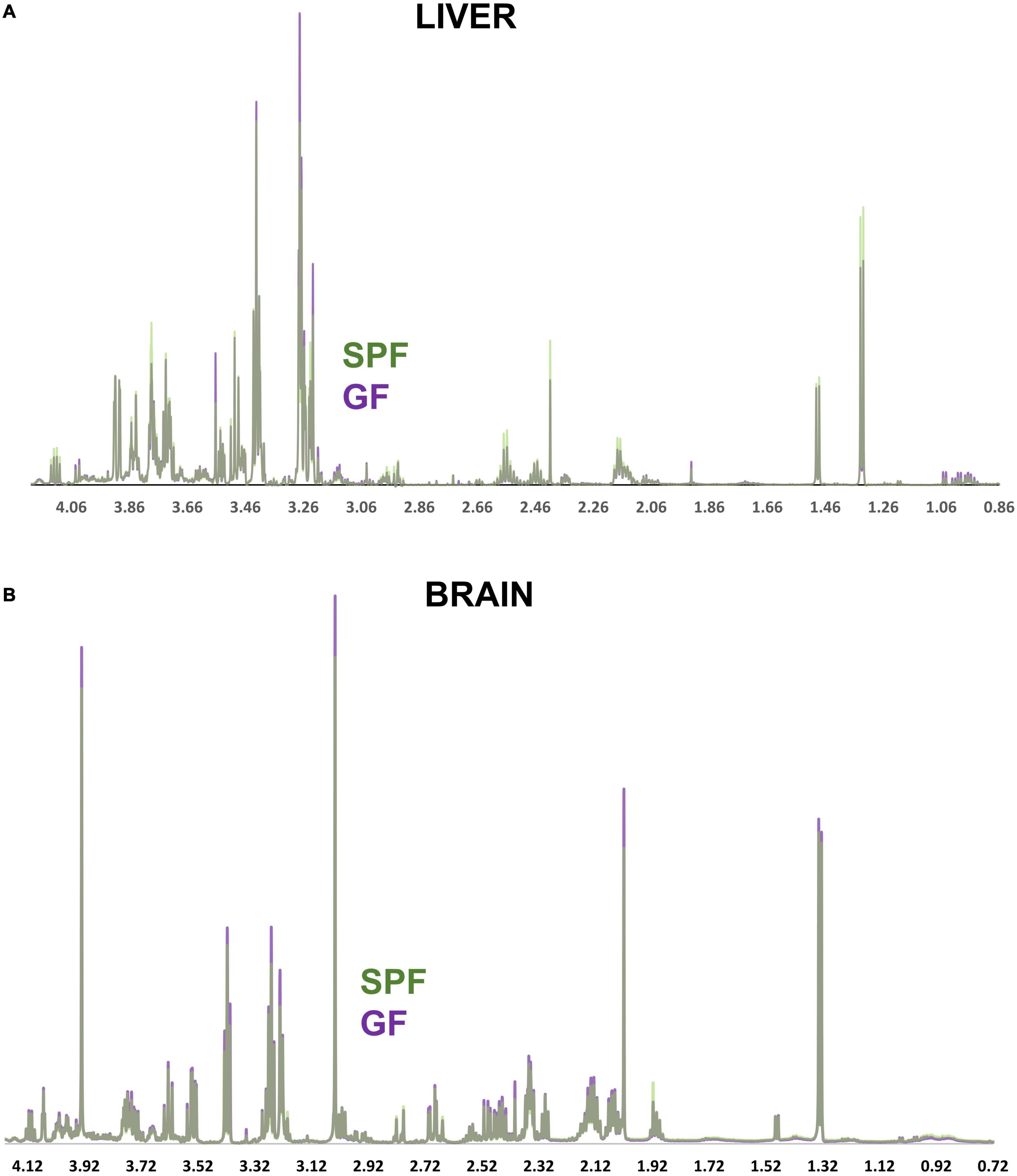
Figure 3. NMR spectra comparing the GF and SPF liver and brain metabolome. (A) Median spectral intensity of the GF (purple, n = 6) and SPF (green, n = 6) liver metabolome. (B) Median spectral intensity of the GF (purple, n = 6) and SPF (green, n = 6) brain metabolome. Several metabolites were altered by absence of bacterial flora in both liver and brain GF tissue (Figures 4, 5). In the brain, taurine, myoinositol, creatine, glutamate, glutamine, choline, and NAA were significantly increased in GF mice relative to SPF animals (Figure 4). Conversely, acetate and various lipoprotein resonances were reduced in GF animals (Figure 4). Inspection of the corresponding loadings plot indicates that cerebral lactate and taurine levels may have contributed to the variation observed in the GF mice brain samples (Figure 2B).

Table 2. Summary of metabolic changes in the brain and liver of germ free mice.
Several metabolites were altered by absence of bacterial flora in both liver and brain GF tissue (Figures 4, 5). In the brain, taurine, myoinositol, creatine, glutamate, glutamine, choline, and NAA were significantly increased in GF mice relative to SPF animals (Figure 4). Conversely, acetate and various lipoprotein resonances were reduced in GF animals (Figure 4). Inspection of the corresponding loadings plot indicates that cerebral lactate and taurine levels may have contributed to the variation observed in the GF mice brain samples (Figures 2A, B).
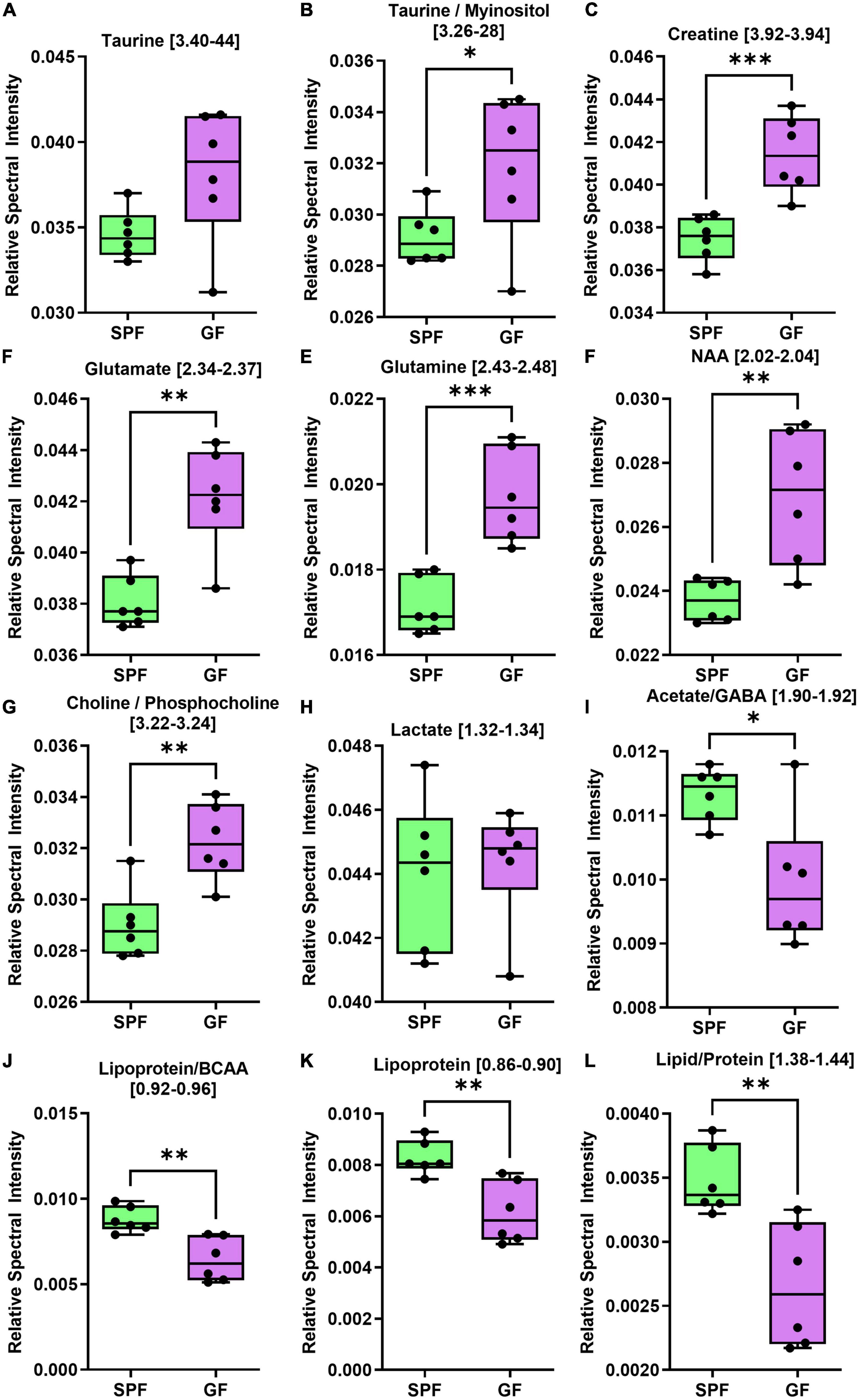
Figure 4. The brain (cerebrum) metabolome is altered in GF (n = 6) compared to SPF (n = 6) mice. Boxplots represent the interquartile range (IQR), and whiskers display minimum/maximum values. (A) Taurine was increased in GF (p = 0.0649). (B) Taurine/myoinositol (p = 0.0481*), (C) creatine (p = 0.0010***), (D) glutamate (p = 0.0013**), (E) glutamine (p = 0.0006*), (F) NAA (p = 0.0046**), and (G) choline/phosphocholine (p = 0.0028**) were all significantly increased in GF. (H) There were no significant variations in lactate (p = 0.7975). (I) Acetate/GABA (p = 0.0120*), (J) lipoprotein/BCAA (p = 0.0031**), (K) lipoprotein (p = 0.0033**), and (L) lipid/protein (p = 0.0043**) were all significantly lower in GF. In the liver, several amino acids were increased in GF mice relative to SPF controls (Figure 5), whereas certain microbial metabolites (succinate, TMAO) were significantly reduced in relative abundance.
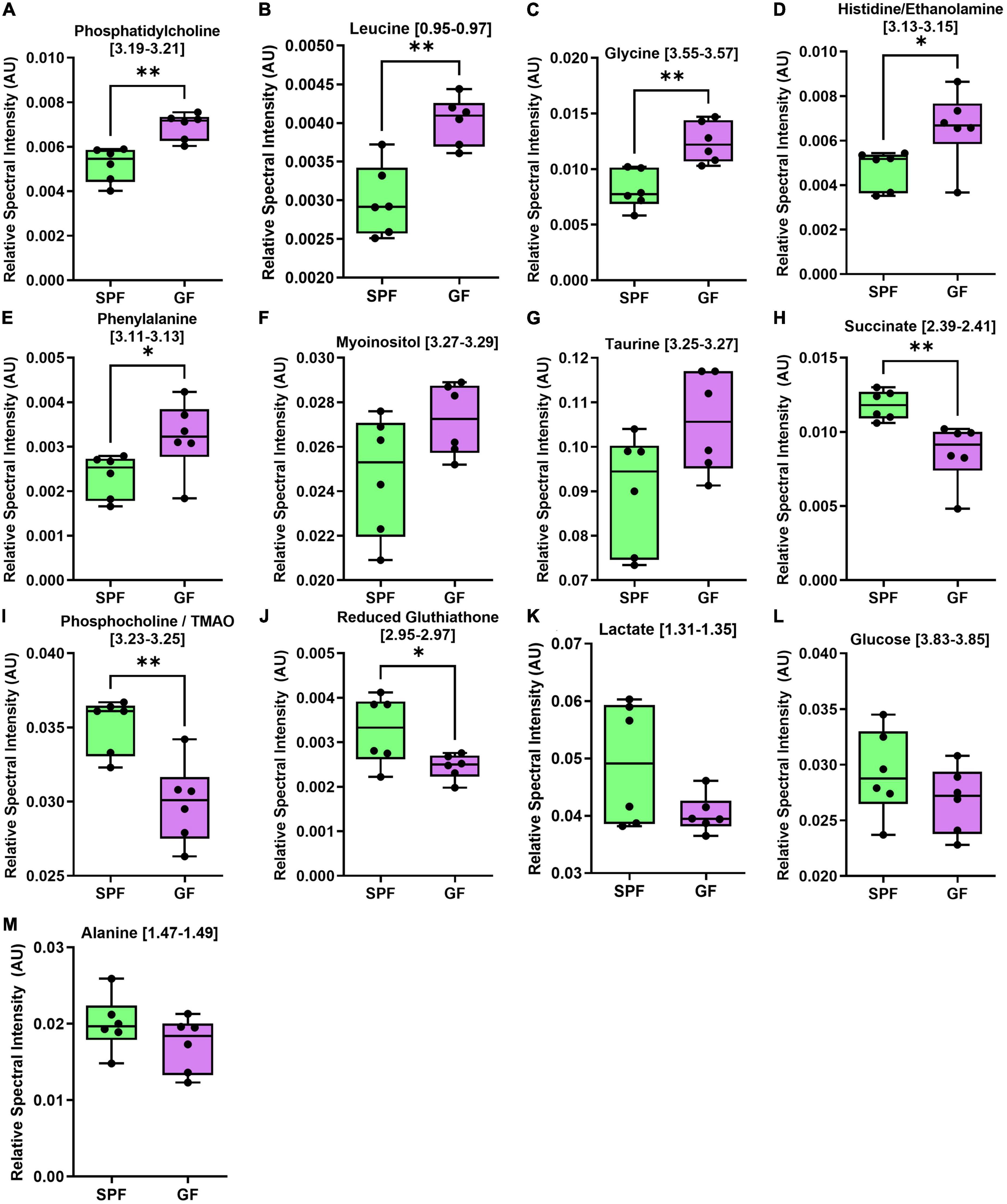
Figure 5. The liver metabolome is altered in GF (n = 7) compared to SPF (n = 5) mice. Boxplots represent the interquartile range (IQR), and whiskers display minimum/maximum values. (A) Phosphatidylcholine (p = 0.0015**), (B) leucine (p = 0.0010**), (C) glycine (p = 0.0019**), (D) histidine/ethanolamine (p = 0.0327*), and (E) phenylalanine (p = 0.0451*) were all significantly increased in GF. (F) Myoinositol (p = 0.0532) and (G) taurine (p = 0.0812) were increased in GF. (H) Succinate (p = 0.0057**), (I) phosphocholine/TMAO (p = 0.0030**), and (J) reduced glutathione (p = 0.0499*) were significantly decreased in GF. (K) Lactate (p = 0.1012), (L) glucose (p = 0.2489), and (M) alanine (p = 0.2156) were not significantly different between groups.
In the liver, several amino acids were increased in GF mice relative to SPF controls (Figure 5), whereas certain microbial metabolites (succinate, TMAO) were significantly reduced in relative abundance.
Germ-free mice have significant changes in gene expression in brain and liver tissue
In the frontal cortex of GF and SPF mice, we profiled the mRNA expression of 22 genes (Table 1) relating to neuroplasticity, immunity, and serotonin and glutamate receptors (Figure 6 and Supplementary Figure 1). In the liver, 6 genes were investigated, including IL-1B, SAA2, GFAP, IL-6, TNF, and TLR4 (Figure 7 and Table 1).
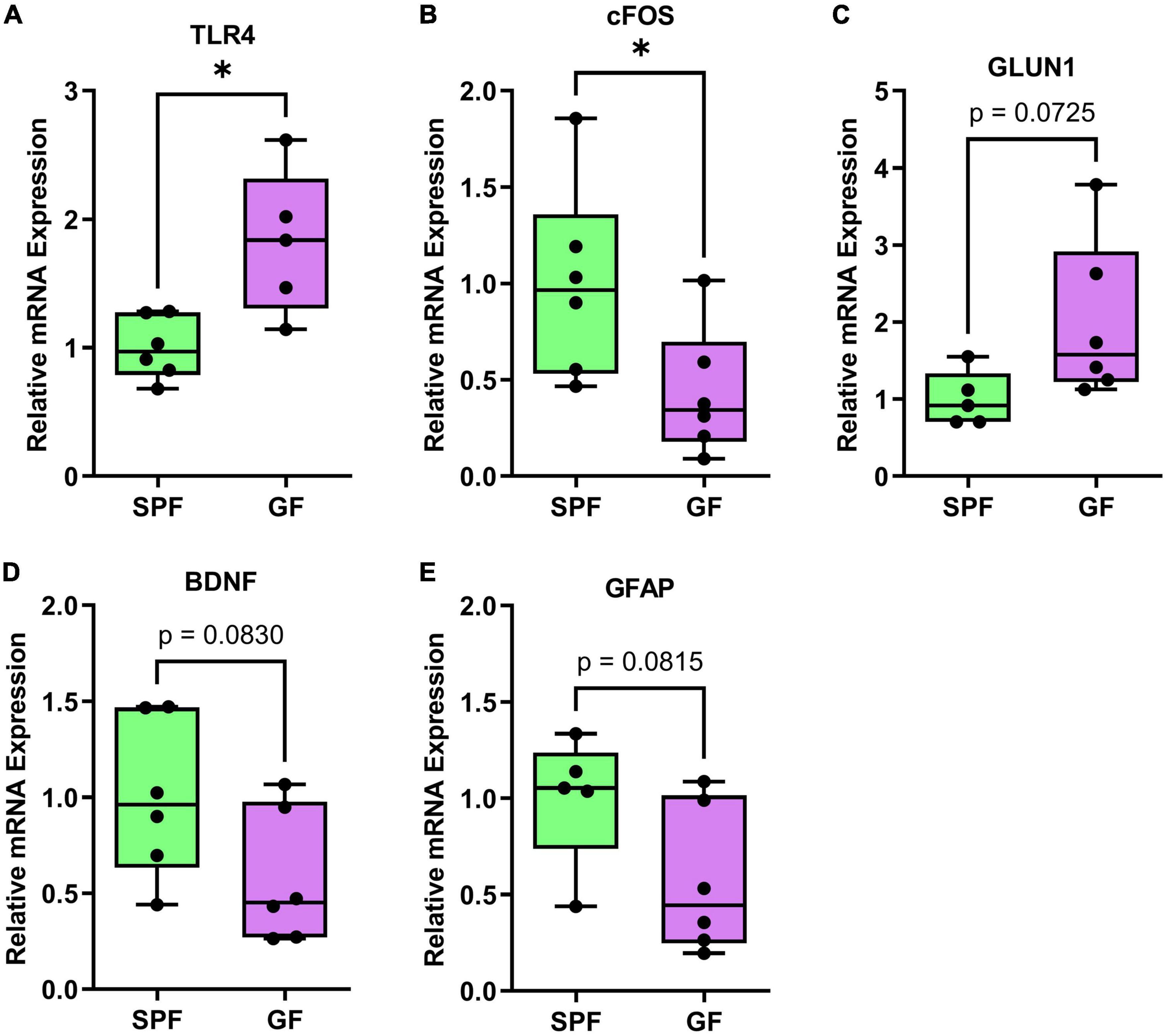
Figure 6. Significant or trending genes from brain qPCR analysis in GF (n = 6) and SPF (n = 6) mice. *p < 0.05, trending p < 0.1. Boxplots represent the interquartile range (IQR) and whisker display minimum/maximum values. (A) TLR4 had significantly higher expression in GF (p = 0.0101*). (B) cFOS had significantly lower expression (p = 0.0435*). (C) GLUN1 (p = 0.0725) had higher expression, (D) BDNF (p = 0.0830) and (E) GFAP (p = 0.0815) had lower expression.
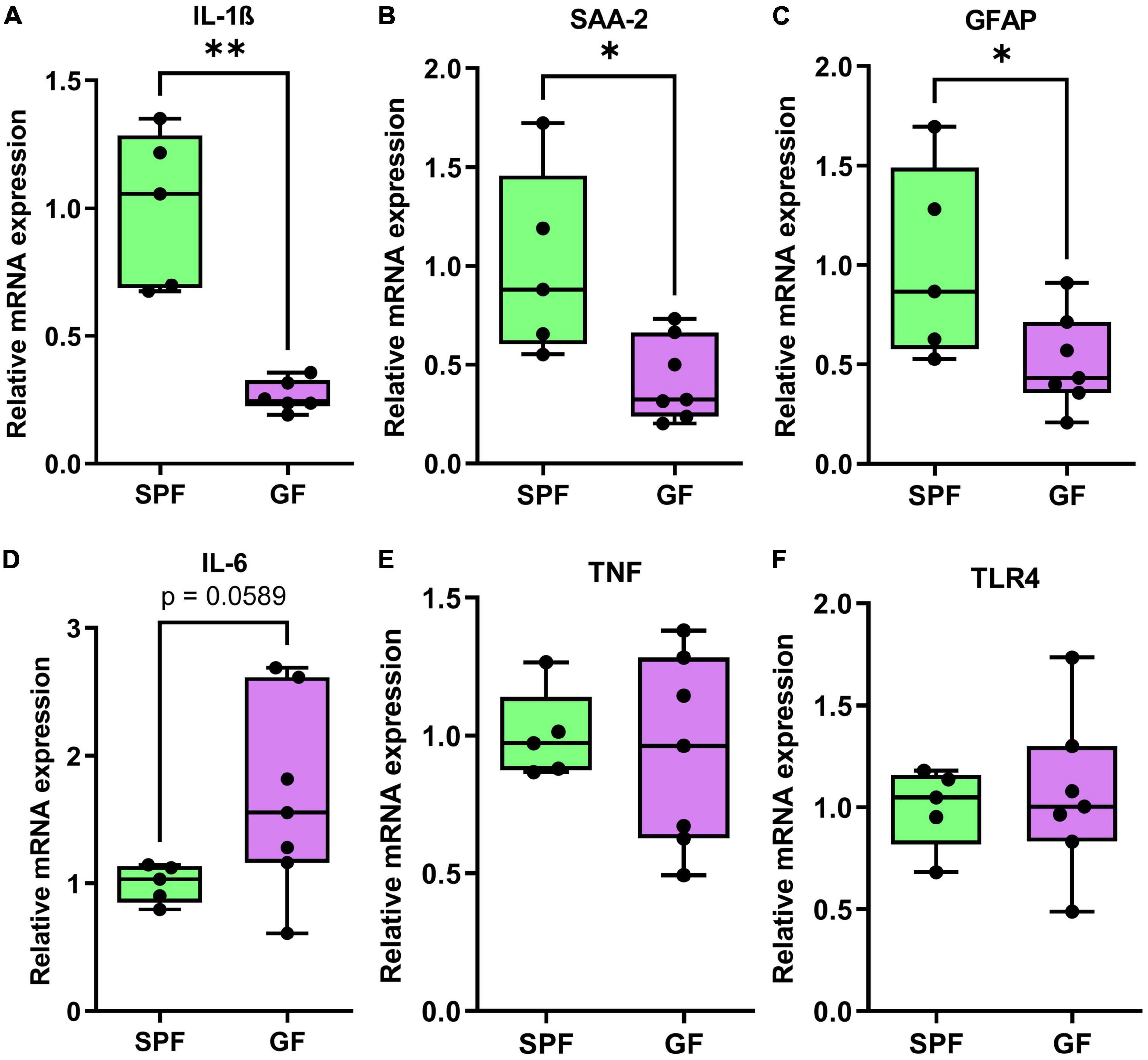
Figure 7. qPCR genes analysis of liver tissue in GF (n = 7) and SPF (n = 5) mice. Boxplots represent the interquartile range (IQR) and whiskers display minimum/maximum values. (A) Interleukin-1ß (p = 0.0050**), (B) serum amyloid A (p = 0.0162*), and (C) glial fibrillary acidic protein (p = 0.0424*) were all significantly lower in GF. (D) Interleukin-6 (p = 0.0589) was trending higher in GF animals relative to SPF animals. (E) TNF (p = 0.7184) and (F) toll-like receptor-4 (p = 0.7961) were not significantly different between GF and SPF animals.
In the prefrontal cortex, TLR4 was increased in GF mice relative to SPF controls (p = 0.01), whereas cFOS was decreased (p = 0.044). BDNF (p = 0.083) and GLUN1 (p = 0.073) showed a numeric (non-significant) reduction and increase in GF mice, respectively.
In the liver, mRNA expression of immunity-related factors IL-1B (p = 0.0050), SAA2 (p = 0.016), and GFAP (p = 0.042) were reduced in GF mice relative to SPF controls (Figure 6). IL-6 (p = 0.0589) was trending higher in GF animals, but there was no difference in mRNA levels of TNF or TLR4 in the liver (Figure 7).
Correlations between metabolites and gene expression
Correlations between gene expression and key brain and liver metabolites were analyzed to infer potential connections between metabolic changes and neuroplasticity. A heatmap was used to visualize correlations (Figure 8); those that remained significant following FDR correction are shown in Supplementary Figure 2. Most notably, cFOS expression correlated significantly with several brain metabolites that were also significantly altered by the GF phenotype, whereas only liver succinate was significantly correlated with PFC cFOS expression (Figure 8 and Supplementary Figure 3).
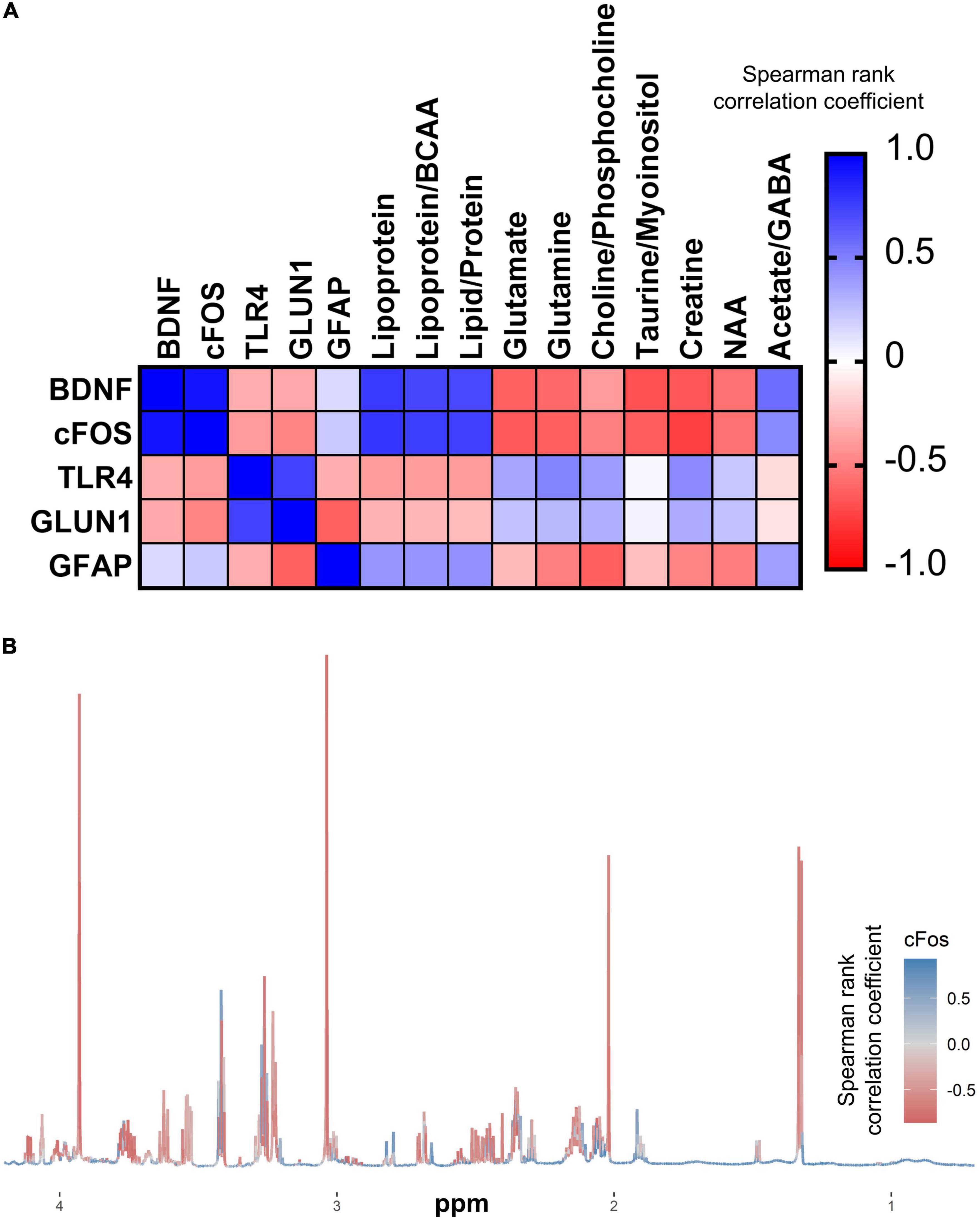
Figure 8. cFOS expression is intimately linked to cerebral metabolism. (A) Correlation matrix of qPCR gene expression in the PFC with cerebral metabolites that were significantly altered in GF (n = 6) vs. SPF (n = 6) mice. (B) Median NMR spectrum (n = 12) of the brain metabolome, colored by correlation with PFC cFOS expression.
Discussion
In this study, we aimed to define key intermediary factors (metabolites) involved in gut microbiota to brain communication, with implications for the role of the MGB axis in anxiety and other mood disorders (Table 2). We inferred altered brain function from significant changes to brain gene expression between age and strain -matched GF and SPF mice. We report significant gene expression and metabolomic changes in GF compared to SPF juvenile mice in liver and brain tissue. Of particular note, we identified a significant reduction in cFOS expression in the prefrontal cortex of GF mice, which may relate to the robust reduced-anxiety phenotype demonstrated by GF animals (Neufeld et al., 2011; De Palma et al., 2015). Furthermore, our results suggest a direct relationship between the microbiota regulation of host metabolism and brain plasticity, as cFOS expression correlated with a number of brain metabolites that were altered between GF and SPF animals. These findings shed light on the complex mechanisms underlying the MGB axis and provide insights into potential avenues for the development of novel treatments for stress-related psychiatric disorders such as anxiety.
Liver metabolism
Microbiota contribute to normal host metabolism. In the GF liver, we identified significantly higher levels of several amino acids usually metabolized by commensals, including leucine, glycine, and phenylalanine. Similarly, lower levels of metabolic by-products were observed, such as succinate and trimethylamine N-oxide (TMAO). These findings are consistent with several reports comparing liver (Mardinoglu et al., 2015) and blood (Lai et al., 2021) metabolomic differences between GF and SPF mice, and likely represent an altered gut amino acid profile (due to the lack of microbial metabolism in GF mice) entering hepatic tissue via the portal vein.
Liver and brain immunity
Reduced immune markers in the liver (IL-1ß, GFAP, and SAA-2), suggests immunological homeostasis may be disrupted (Furusawa et al., 2013). GFAP is expressed on stellate cells in the liver, which are normally thought to be quiescent but may be activated in response to liver injury or inflammation (Winau et al., 2007). It is unsurprising that hepatic SAA-2 follows a similar expression pattern, as SAA-2 is known to be a primary mediator of the downstream inflammatory response in activated hepatic stellate cells (Siegmund et al., 2016). It is important to note that SAA-2 may be upregulated several-hundred-fold after an inflammatory challenge (Yates et al., 2022). Thus, a modest 50% reduction in expression may be interpreted as a reduction in basal expression levels, possibly due to a lack of pathogen-associated molecular patterns (PAMPs) entering the portal circulation from the gut.
Although we were initially surprised to find no significant difference in liver TLR4 expression between GF and SPF mice, this finding has been reported in peripheral tissues by other laboratories (Amaral et al., 2008). It is possible that TLR4 is constitutively expressed in the liver, and therefore, the discrepancy between GF and SPF mice lies in the level of TLR4 activation and downstream inflammatory signaling. Our results showed reduced expression of IL-1B, SAA-2, and GFAP in GF mice relative to SPF mice, suggesting that the absence of circulating microbial antigens reduces basal inflammatory tone in the periphery.
Our study revealed a novel finding of increased TLR4 expression in the brains of GF mice. We propose that the absence of exposure of brain endothelial cells to circulating PAMPs, such as endotoxin, may have led to an adaptive increase in TLR4 expression to maintain homeostasis. Alternatively, previous research has shown that microbiota-derived acetate contributes to microglia homeostasis in the rodent brain (Erny et al., 2021). Since we identified a similar reduction in brain acetate levels in GF mice, it is possible that aberrant TLR4 expression could be due to disrupted microglia function, which has not been previously reported.
TLR4 has been proposed to play a significant role in modulating synaptic plasticity during neurodevelopment (Okun et al., 2011), with the exact role of neuronal TLR4 expression yet to be fully understood. Previous studies have suggested an interaction between TLR4 expression and NMDAR activity (Kashima and Grueter, 2017). Notably, TLR4-deficient animals exhibit deficits in drug reward learning, indicating that microbiota-induced alterations in TLR4 expression during key neurodevelopmental periods may influence anhedonia through changes in synaptic physiology, specifically NMDAR levels.
Alterations in synaptic plasticity
We found reduced cortical expression of the synaptogenesis plasticity protein, brain-derived neurotrophic factor (BDNF) in GF mice, corroborated by numerous studies examining expression in the cortex, but also other brain regions, including the hippocampus and amygdala (Diaz Heijtz et al., 2011; Neufeld et al., 2011; Clarke et al., 2013). Chronic stress negatively impacts BDNF expression, via glucocorticoid mechanisms (Chen et al., 2017) and so altered development of the hypothalamic pituitary adrenal (HPA) axis in GF mice may explain aberrant BDNF expression, though further studies assessing behavioral and physiological responses to stress are required (Sudo et al., 2004). Depression is also associated with a decrease in BDNF, demonstrated by post-mortem analysis in the PFC of depressed individuals (Karege et al., 2005) and in animal models of depression (Li et al., 2016). Indeed, a prevailing theory of depression pathophysiology is a reduced capacity to neurochemically adapt to stress through changes in plasticity (Liu et al., 2017). Disruption of early life commensal-dependent HPA axis development may contribute to depression through BDNF suppression. Disruptions to normal microbiota engraftment in early life due to caesarean birth (Dominguez-Bello et al., 2010) or antibiotic treatment (Fouhy et al., 2012) may therefore have long-term consequences for brain function and increase the risk of stress-related psychiatric conditions.
The microbiome affects the levels of short-chain fatty acids
Despite considerable debate about the exact microbiota composition alterations in MDD, it is consistently found that bacteria responsible for producing SCFAs (namely, butyrate and acetate), such as Faecalibacterium, Lachnospiraceae, and Lactobacillus are reduced in chronically stressed and depressive individuals (Jiang et al., 2015; Aizawa et al., 2019). Here, we identified reduced brain acetate in GF mice relative to SPF controls. Acetate in particular appears to be crucial for the maintenance of microglial complexity suggesting that continuous microbial input and diversity is continuously required for normal function (Erny et al., 2021). The involvement of microglia in synaptic pruning and developmental neural circuit shaping may contribute to the regulation of stress- and depression-related circuitry in adulthood, further exemplifying how developmental dysbiosis may promote future stress-related depression (Schafer et al., 2012).
Short-chain fatty acids contribution to healthy brain function illuminates a putative therapeutic target. Huang et al. (2021) found acetate supplementation promoted hippocampal plasticity and induced antidepressant-like effects in a chronic social defeat stress (CSDS) mouse model. While this study has exciting potential, it is limited in that it only looked at the positive control, fluoxetine, in terms of behavior not gene expression; it may have been useful to assess if antidepressants produced similar gene expression changes to acetate, possibly indicating similar underlying mechanisms. Nonetheless, since acetate (in the form of glyceryl triacetate) has already been found safe for human use (Madhavarao et al., 2009), it paves the way for clinical human trials to explore its therapeutic potential in stress-related psychiatric disorders.
Exploring correlations between gene expression and metabolomic profiles
cFOS expression in the prefrontal cortex was significantly correlated with the abundance of several metabolites in the cerebrum associated with the GF phenotype. cFOS protein dime rises with members of the Jun family to form activator protein-1 (AP-1), a transcription factor with widespread activating effects on gene expression, such as on neurotransmitter-producing enzymes, glutamate decarboxylase (producing GABA) and tyrosine hydroxylase (producing dopamine) (Najlerahim et al., 1991; Sabban et al., 1995). Alterations in cFOS activity could disrupt the brain’s excitatory-inhibitory balance or dopaminergic modulation, potentially with consequences on anxiety-like behaviors. Given the role of cFOS role as a master regulator, our data putatively suggests an avenue in which microbiota, via modulating the metabolomic landscape, may regulate host brain plasticity.
It is currently unknown which metabolites, and by what mechanism, may regulate cFOS expression, but our findings raise some suggestions. Previous data reveal taurine administration to have a suppressive effect on peripheral cFOS expression in mesenchymal cells (Terashima et al., 2003). Although not yet demonstrated in the brain, there may be overlap in peripheral and central taurine actions which would explain our negative correlation between taurine and cFOS. The observed significant increase in extracellular glutamate in GF mice could result in calcium-mediated neuronal excitotoxic death, reducing the amount of cortical neuronal activity, reflected by reduced cFOS, given its role as a marker of neural activity. A microglial culture study found glutamate receptor activation induced cFOS expression initially, but high concentrations dramatically decreased cFOS expression, supporting our hypothesis (Eun et al., 2004). Preliminary findings also implicate acetate: CSDS-induced depression results in suppressed cFOS expression which can be mostly restored by acetate supplementation (Huang et al., 2021). Given the high level of correlational significance with cFOS and altered metabolites, it warrants further study to determine if a causal link between microbially-affected metabolite levels and cFOS expression exists. This may suggest another potential mechanism of microbial involvement in mood and potentially dysbiosis’ contribution to stress-related psychiatric disorders.
Future directions
Several further research avenues can be explored from this finding. Firstly, a panel of behavioral assays, addressing anxiety-like behavior, depressive-like behavior, anhedonia, and memory, could be employed to determine how altered cFOS expression and brain metabolite levels affect emotional and cognitive behaviors. While several studies have identified reduced anxiety-like behavior in male and female GF mice in adulthood (Luczynski et al., 2016), it would also be important to determine whether this phenotype exists in adolescent mice. Additionally, gene expression in other brain regions (amygdala and hippocampus) could be mapped to the cortex metabolome and other distinct brain regions (brainstem and cerebellum) to more comprehensively investigate gene-metabolite associations. This approach, however, would necessitate mass spectrometry techniques or pooled sampling to overcome the issue of limited tissue mass per brain region.
Electrophysiology experiments could also be employed to understand how changes in brain gene expression and metabolite levels correspond to synaptic changes between brain regions, which may be of particular interest given the observed significant/trending reduction in PFC cFOS/BDNF, respectively, and significant/trending increase in TLR4/GLUN1, respectively. Specifically, neuronal activity could be investigated in depression-related PFCbrainstem projections (Warden et al., 2012) or midbrainamygdala projections (Chaudhury et al., 2013; Koo et al., 2019; Morel et al., 2022).
Such experiments could be followed-up by the reintroduction of individual commensal strains into GF mice, observing metabolomic and neuronal gene expression changes, and the impact on depressive and anxiety-like behaviors. Mapping neurochemical and behavioral changes to individual commensal strains will progress the MGB axis field toward the development of novel therapeutics, and further elucidate the microbiota’s role in brain metabolism.
Limitations
Whilst this study provided significant evidence for the impact of the gut microbiome on the host, there are important limitations. Firstly, it may be useful to confirm whether changes in mRNA expression levels reflect changes in protein abundance, particularly of FOS protein. Nevertheless, it has been suggested that cFOS transcription, not translation, serves as a readout for neuronal activity in the PFC, and by extension, neuronal plasticity (Sheng et al., 1990). Indeed, this is supported by the significant correlation between cFOS mRNA (upstream of FOS protein translation) and cerebral metabolite levels (downstream of FOS protein translation).
Secondly, only female GF and SPF mice were used to assess the gene expression and metabolomic correlations in this study. While the use of female mice is justified by the increased prevalence of disorders of gut-brain interaction, such as irritable bowel syndrome in females (Canavan et al., 2014), we recommend that further research explores whether sex differences may exist in the GF phenotype on PFC gene expression and metabolite levels. In this study, we performed qPCR on prefrontal cortex tissue and metabolomics on the contralateral cerebrum. Therefore, the correlation analysis compares different brain regions, which may limit the conclusions drawn from these results. Conversely, comprehensive metabolomic studies of the mouse brain across the lifespan have shown that in “adolescence” (3 weeks), the cerebrum exhibits a high degree of correlation between distinct brain regions (Ding et al., 2021). Whilst differences in the aforementioned study were noted between the cerebrum as a whole, the brainstem, and cerebellum, and between cerebral regions in adult and aged mice, the high degree of convergence within the adolescent cerebrum is most pertinent to our current study. Importantly, the paper utilized 3-week-old, female, C57Bl/6 mice, identical to our own study. Separately, we have demonstrated similar findings, whereby cerebrum metabolite levels and PFC mRNA correlations are stronger in 3-week-old mice compared to 16-week-old mice (Radford-Smith et al., 2022). Therefore, it is unlikely that performing metabolomics on just the contralateral PFC region would show different results to the metabolomics and correlation analyses performed here.
Conclusion
This study demonstrated that microbiota clearly exert a significant impact on host physiology. Until recently, their effect on brain function was largely underestimated. This study observed gene expression and metabolomic changes, identifying novel correlations between cFOS expression and several metabolites. Our pilot study provides preliminary evidence that the host metabolome profile, modified by the microbial composition, may influence brain gene expression, at least partially through regulation of the widespread transcriptional regulator, cFOS and putatively through HPA axis regulation. The microbiome’s impact on metabolite levels and gene expression, may be crucially implicated in depression, through neuroinflammatory, and neuroplasticity mechanisms. It is therefore not surprising that dysbiosis, even in early life, is strongly implicated in stress-related psychiatric disorders. The gut microbiome and associated metabolites, such as SCFAs, may be an important therapeutic target for novel or augmenting treatment strategies for those afflicted with depression.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation. The liver and brain metabolomics data analyzed in this study is available online at http://dx.doi.org/10.5287/ora-zv6mnkajn.
Ethics statement
This animal study was reviewed and approved by the UK Animal Welfare and Ethical Review Body. All animal procedures were carried out in accordance with UK Home Office Animals (Scientific Procedures) Act (1986) and associated Home Office guidelines under license P996B4A4E.
Author contributions
TP and DR-S performed the research and conducted the analysis. TP, DR-S, and DA drafted the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was funded by an MRC Confidence in Concepts grant. The APC was funded by the UKRI block grant—Open Access Fund, University of Oxford.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnmol.2023.1155620/full#supplementary-material
References
Aizawa, E., Tsuji, H., Asahara, T., Takahashi, T., Teraishi, T., Yoshida, S., et al. (2019). Bifidobacterium and Lactobacillus counts in the gut microbiota of patients with bipolar disorder and healthy controls. Front. Psychiatry 9:730. doi: 10.3389/fpsyt.2018.00730
Amaral, F. A., Sachs, D., Costa, V. V., Fagundes, C. T., Cisalpino, D., Cunha, T. M., et al. (2008). Commensal microbiota is fundamental for the development of inflammatory pain. Proc. Natl. Acad. Sci. 105, 2193–2197. doi: 10.1073/pnas.0711891105
Barreau, F., Ferrier, L., Fioramonti, J., and Bueno, L. (2004). Neonatal maternal deprivation triggers long term alterations in colonic epithelial barrier and mucosal immunity in rats. Gut 53, 501–506. doi: 10.1136/gut.2003.024174
Borghi, S. M., Fattori, V., Pinho-Ribeiro, F. A., Domiciano, T. P., Miranda-Sapla, M. M., Zaninelli, T. H., et al. (2019). Contribution of spinal cord glial cells to L. amazonensis experimental infection-induced pain in BALB/c mice. J Neuroinflammation 16:113. doi: 10.1186/s12974-019-1496-2
Borst, K., Dumas, A. A., and Prinz, M. (2021). Microglia: Immune and non-immune functions. Immunity 54, 2194–2208. doi: 10.1016/j.immuni.2021.09.014
Bravo, J. A., Forsythe, P., Chew, M. V., Escaravage, E., Savignac, H. M., Dinan, T. G., et al. (2011). Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. U.S.A. 108, 16050–16055. doi: 10.1073/pnas.1102999108
Brescia, P., and Rescigno, M. (2021). The gut vascular barrier: A new player in the gut–liver–brain axis. Trends Mol. Med. 27, 844–855. doi: 10.1016/j.molmed.2021.06.007
Canavan, C., West, J., and Card, T. (2014). The epidemiology of irritable bowel syndrome. Clin. Epidemiol. 6, 71–80. doi: 10.2147/CLEP.S40245
Cebra, J. J. (1999). Influences of microbiota on intestinal immune system development. Am. J. Clin. Nutr. 69, 1046S–1051S. doi: 10.1093/ajcn/69.5.1046s
Chaudhury, D., Walsh, J. J., Friedman, A. K., Juarez, B., Ku, S. M., Koo, J. W., et al. (2013). Rapid regulation of depression-related behaviours by control of midbrain dopamine neurons. Nature 493, 532–536. doi: 10.1038/nature11713
Chen, H., Lombès, M., and Le Menuet, D. (2017). Glucocorticoid receptor represses brain-derived neurotrophic factor expression in neuron-like cells. Mol. Brain 10:12. doi: 10.1186/s13041-017-0295-x
Clarke, G., Grenham, S., Scully, P., Fitzgerald, P., Moloney, R. D., Shanahan, F., et al. (2013). The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 18, 666–673. doi: 10.1038/mp.2012.77
Cryan, J. F., O’Riordan, K. J., Cowan, C. S. M., Sandhu, K. V., Bastiaanssen, T. F. S., Boehme, M., et al. (2019). The microbiota-gut-brain axis. Physiol. Rev. 99, 1877–2013. doi: 10.1152/physrev.00018.2018
De Palma, G., Blennerhassett, P., Lu, J., Deng, Y., Park, A. J., Green, W., et al. (2015). Microbiota and host determinants of behavioural phenotype in maternally separated mice. Nat. Commun. 6:7735. doi: 10.1038/ncomms8735
Diaz Heijtz, R., Wang, S., Anuar, F., Qian, Y., Björkholm, B., Samuelsson, A., et al. (2011). Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. U.S A. 108, 3047–3052. doi: 10.1073/pnas.1010529108
Ding, J., Ji, J., Rabow, Z., Shen, T., Folz, J., Brydges, C. R., et al. (2021). A metabolome atlas of the aging mouse brain. Nat. Commun. 12:6021. doi: 10.1038/s41467-021-26310-y
Dominguez-Bello, M. G., Costello, E. K., Contreras, M., Magris, M., Hidalgo, G., Fierer, N., et al. (2010). Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. 107, 11971–11975. doi: 10.1073/pnas.1002601107
Erny, D., Dokalis, N., Mezö, C., Castoldi, A., Mossad, O., Staszewski, O., et al. (2021). Microbiota-derived acetate enables the metabolic fitness of the brain innate immune system during health and disease. Cell Metab. 33, 2260–2276.e7. doi: 10.1016/j.cmet.2021.10.010
Erny, D., Hrabì de Angelis, A. L., Jaitin, D., Wieghofer, P., Staszewski, O., David, E., et al. (2015). Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 18, 965–977. doi: 10.1038/nn.4030
Eun, S.-Y., Hong, Y. H., Kim, E. H., Jeon, H., Suh, Y. H., Lee, J. E., et al. (2004). Glutamate receptor-mediated regulation of c-fos expression in cultured microglia. Biochem. Biophys. Res. Commun. 325, 320–327. doi: 10.1016/j.bbrc.2004.10.035
Fouhy, F., Guinane, C. M., Hussey, S., Wall, R., Ryan, C. A., Dempsey, E. M., et al. (2012). High-throughput sequencing reveals the incomplete, short-term recovery of infant gut microbiota following parenteral antibiotic treatment with ampicillin and gentamicin. Antimicrob. Agents Chemother. 56, 5811–5820. doi: 10.1128/AAC.00789-12
Furusawa, Y., Obata, Y., Fukuda, S., Endo, T. A., Nakato, G., Takahashi, D., et al. (2013). Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504, 446–450. doi: 10.1038/nature12721
Hebert, J. C., Radford-Smith, D. E., Probert, F., Ilott, N., Chan, K. W., Anthony, D. C., et al. (2021). Mom’s diet matters: Maternal prebiotic intake in mice reduces anxiety and alters brain gene expression and the fecal microbiome in offspring. Brain Behav. Immun. 91, 230–244. doi: 10.1016/j.bbi.2020.09.034
Huang, W., Hu, W., Cai, L., Zeng, G., Fang, W., Dai, X., et al. (2021). Acetate supplementation produces antidepressant-like effect via enhanced histone acetylation. J. Affect. Disord. 281, 51–60. doi: 10.1016/j.jad.2020.11.121
Iñiguez, S. D., Flores-Ramirez, F. J., Themann, A., and Lira, O. (2021). Adolescent fluoxetine exposure induces persistent gene expression changes in the hippocampus of adult male C57BL/6 mice. Mol. Neurobiol. 58, 1683–1694. doi: 10.1007/s12035-020-02221-9
Jiang, H., Ling, Z., Zhang, Y., Mao, H., Ma, Z., Yin, Y., et al. (2015). Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 48, 186–194. doi: 10.1016/j.bbi.2015.03.016
Karege, F., Vaudan, G., Schwald, M., Perroud, N., and La Harpe, R. (2005). Neurotrophin levels in postmortem brains of suicide victims and the effects of antemortem diagnosis and psychotropic drugs. Brain Res. Mol. Brain Res. 136, 29–37. doi: 10.1016/j.molbrainres.2004.12.020
Kashima, D. T., and Grueter, B. A. (2017). Toll-like receptor 4 deficiency alters nucleus accumbens synaptic physiology and drug reward behavior. Proc. Natl. Acad. Sci. 114, 8865–8870. doi: 10.1073/pnas.1705974114
Koo, J. W., Chaudhury, D., Han, M.-H., and Nestler, E. J. (2019). Role of mesolimbic brain-derived neurotrophic factor in depression. Biol. Psychiatry 86, 738–748. doi: 10.1016/j.biopsych.2019.05.020
Kronsten, V. T., Tranah, T. H., Pariante, C., and Shawcross, D. L. (2022). Gut-derived systemic inflammation as a driver of depression in chronic liver disease. J. Hepatol. 76, 665–680. doi: 10.1016/j.jhep.2021.11.008
Lai, Y., Liu, C.-W., Yang, Y., Hsiao, Y.-C., Ru, H., and Lu, K. (2021). High-coverage metabolomics uncovers microbiota-driven biochemical landscape of interorgan transport and gut-brain communication in mice. Nat. Commun. 12:6000. doi: 10.1038/s41467-021-26209-8
Li, M., Du, W., Shao, F., and Wang, W. (2016). Cognitive dysfunction and epigenetic alterations of the BDNF gene are induced by social isolation during early adolescence. Behav. Brain Res. 313, 177–183. doi: 10.1016/j.bbr.2016.07.025
Liu, W., Ge, T., Leng, Y., Pan, Z., Fan, J., Yang, W., et al. (2017). The role of neural plasticity in depression: from hippocampus to prefrontal cortex. Neural Plast. 2017, 6871089. doi: 10.1155/2017/6871089
Luczynski, P., McVey Neufeld, K.-A., Oriach, C. S., Clarke, G., Dinan, T. G., and Cryan, J. F. (2016). Growing up in a bubble: using germ-free animals to assess the influence of the gut microbiota on brain and behavior. Int. J. Neuropsychopharmacol. 19:yw020. doi: 10.1093/ijnp/pyw020
Madhavarao, C. N., Arun, P., Anikster, Y., Mog, S. R., Staretz-Chacham, O., Moffett, J. R., et al. (2009). Glyceryl triacetate for Canavan disease: a low-dose trial in infants and evaluation of a higher dose for toxicity in the tremor rat model. J. Inherit. Metab. Dis. 32:640. doi: 10.1007/s10545-009-1155-3
Mardinoglu, A., Shoaie, S., Bergentall, M., Ghaffari, P., Zhang, C., Larsson, E., et al. (2015). The gut microbiota modulates host amino acid and glutathione metabolism in mice. Mol. Syst. Biol. 11:834. doi: 10.15252/msb.20156487
Morel, C., Montgomery, S. E., Li, L., Durand-de Cuttoli, R., Teichman, E. M., Juarez, B., et al. (2022). Midbrain projection to the basolateral amygdala encodes anxiety-like but not depression-like behaviors. Nat. Commun. 13:1532. doi: 10.1038/s41467-022-29155-1
Najlerahim, A., Showell, D. G., and Pearson, R. C. (1991). Transient increase in glutamic acid decarboxylase mRNA in the cerebral cortex following focal cortical lesion in the rat. Exp. Brain Res. 87, 113–118. doi: 10.1007/BF00228512
Neufeld, K. M., Kang, N., Bienenstock, J., and Foster, J. A. (2011). Reduced anxiety-like behavior and central neurochemical change in germ-free mice. Neurogastroenterol. Motil. 23, 255–264. doi: 10.1111/j.1365-2982.2010.01620.x
Nicolas, S., McGovern, A. J., Hueston, C. M., O’Mahony, S. M., Cryan, J. F., O’Leary, O. F., et al. (2022). Prior maternal separation stress alters the dendritic complexity of new hippocampal neurons and neuroinflammation in response to an inflammatory stressor in juvenile female rats. Brain Behav. Immun. 99, 327–338. doi: 10.1016/j.bbi.2021.10.016
Okun, E., Griffioen, K. J., and Mattson, M. P. (2011). Toll-like receptor signaling in neural plasticity and disease. Trends Neurosci. 34, 269–281. doi: 10.1016/j.tins.2011.02.005
Qin, J., Li, R., Raes, J., Arumugam, M., Burgdorf, K. S., Manichanh, C., et al. (2010). A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464, 59–65. doi: 10.1038/nature08821
Radford-Smith, D. E., Probert, F., Burnet, P. W. J., and Anthony, D. C. (2022). Modifying the maternal microbiota alters the gut-brain metabolome and prevents emotional dysfunction in the adult offspring of obese dams. Proc. Natl. Acad. Sci. U.S.A. 119, e2108581119. doi: 10.1073/pnas.2108581119
Sabban, E. L., Hiremagalur, B., Nankova, B., and Kvetnanský, R. (1995). Molecular biology of stress-elicited induction of catecholamine biosynthetic enzymes. Ann. N. Y. Acad. Sci. 771, 327–338. doi: 10.1111/j.1749-6632.1995.tb44692.x
Schafer, D. P., Lehrman, E. K., Kautzman, A. G., Koyama, R., Mardinly, A. R., Yamasaki, R., et al. (2012). Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron 74, 691–705. doi: 10.1016/j.neuron.2012.03.026
Schéle, E., Grahnemo, L., Anesten, F., Hallén, A., Bäckhed, F., and Jansson, J.-O. (2013). The gut microbiota reduces leptin sensitivity and the expression of the obesity-suppressing neuropeptides proglucagon (Gcg) and brain-derived neurotrophic factor (Bdnf) in the central nervous system. Endocrinology 154, 3643–3651. doi: 10.1210/en.2012-2151
Semova, I., Carten, J. D., Stombaugh, J., Mackey, L. C., Knight, R., Farber, S. A., et al. (2012). Microbiota regulate intestinal absorption and metabolism of fatty acids in the zebrafish. Cell Host Microbe 12, 277–288. doi: 10.1016/j.chom.2012.08.003
Sheng, M., McFadden, G., and Greenberg, M. E. (1990). Membrane depolarization and calcium induce c-fos transcription via phosphorylation of transcription factor CREB. Neuron 4, 571–582. doi: 10.1016/0896-6273(90)90115-V
Siegmund, S. V., Schlosser, M., Schildberg, F. A., Seki, E., De Minicis, S., Uchinami, H., et al. (2016). Serum amyloid a induces inflammation, proliferation and cell death in activated hepatic stellate cells. PLoS One 11:e0150893. doi: 10.1371/journal.pone.0150893
Sobko, T., Huang, L., Midtvedt, T., Norin, E., Gustafsson, L. E., Norman, M., et al. (2006). Generation of NO by probiotic bacteria in the gastrointestinal tract. Free Radic. Biol. Med. 41, 985–991. doi: 10.1016/j.freeradbiomed.2006.06.020
Strekalova, T., Evans, M., Costa-Nunes, J., Bachurin, S., Yeritsyan, N., Couch, Y., et al. (2015). Tlr4 upregulation in the brain accompanies depression- and anxiety-like behaviors induced by a high-cholesterol diet. Brain Behav. Immun. 48, 42–47. doi: 10.1016/j.bbi.2015.02.015
Sudo, N., Chida, Y., Aiba, Y., Sonoda, J., Oyama, N., Yu, X.-N., et al. (2004). Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J. Physiol. 558, 263–275. doi: 10.1113/jphysiol.2004.063388
Terashima, M., Mitani, T., Hosokawa, Y., Nariai, Y., Imada, K., Kageyama, E., et al. (2003). Suppressive effect of taurine on platelet-derived growth factor (PDGF) BB-induced c-fos and c-jun mRNA expressions through extracellular signal-regulated kinase (ERK) in mesenchymal cell lines. J. Nutr. Sci. Vitaminol. 49, 187–194. doi: 10.3177/jnsv.49.187
Tripathi, A., Debelius, J., Brenner, D. A., Karin, M., Loomba, R., Schnabl, B., et al. (2018). The gut–liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 15, 397–411. doi: 10.1038/s41575-018-0011-z
van der Hee, B., and Wells, J. M. (2021). Microbial regulation of host physiology by short-chain fatty acids. Trends Microbiol. 29, 700–712. doi: 10.1016/j.tim.2021.02.001
Velagapudi, V. R., Hezaveh, R., Reigstad, C. S., Gopalacharyulu, P., Yetukuri, L., Islam, S., et al. (2010). The gut microbiota modulates host energy and lipid metabolism in mice. J. Lipid Res. 51, 1101–1112. doi: 10.1194/jlr.M002774
Vuong, H. E., Pronovost, G. N., Williams, D. W., Coley, E. J. L., Siegler, E. L., Qiu, A., et al. (2020). The maternal microbiome modulates fetal neurodevelopment in mice. Nature 586, 281–286. doi: 10.1038/s41586-020-2745-3
Warden, M. R., Selimbeyoglu, A., Mirzabekov, J. J., Lo, M., Thompson, K. R., Kim, S.-Y., et al. (2012). A prefrontal cortex–brainstem neuronal projection that controls response to behavioural challenge. Nature 492, 428–432. doi: 10.1038/nature11617
Waters, N. J., Holmes, E., Williams, A., Waterfield, C. J., Farrant, R. D., and Nicholson, J. K. (2001). NMR and pattern recognition studies on the time-related metabolic effects of alpha-naphthylisothiocyanate on liver, urine, and plasma in the rat: an integrative metabonomic approach. Chem. Res. Toxicol. 14, 1401–1412. doi: 10.1021/tx010067f
Winau, F., Hegasy, G., Weiskirchen, R., Weber, S., Cassan, C., Sieling, P. A., et al. (2007). Ito cells are liver-resident antigen-presenting cells for activating T cell responses. Immunity 26, 117–129. doi: 10.1016/j.immuni.2006.11.011
Wishart, D. S., Feunang, Y. D., Marcu, A., Guo, A. C., Liang, K., Vázquez-Fresno, R., et al. (2018). HMDB 4.0: the human metabolome database for 2018. Nucleic Acids Res. 46, D608–D617. doi: 10.1093/nar/gkx1089
Yates, A. G., Weglinski, C. M., Ying, Y., Dunstan, I. K., Strekalova, T., and Anthony, D. C. (2022). Nafamostat reduces systemic inflammation in TLR7-mediated virus-like illness. J. Neuroinflammation 19:8. doi: 10.1186/s12974-021-02357-y
Zhao, Z., Ren, Y., Jiang, H., and Huang, Y. (2021). Dexmedetomidine inhibits the PSD95-NMDA receptor interaction to promote functional recovery following traumatic brain injury. Exp. Ther. Med. 21:4. doi: 10.3892/etm.2020.9436
Keywords: microbiota, cFOS, short-chain fatty acids (SCFAs), germ free animal, metabolomics, acetate, glutamate, glutamine
Citation: Pate T, Anthony DC and Radford-Smith DE (2023) cFOS expression in the prefrontal cortex correlates with altered cerebral metabolism in developing germ-free mice. Front. Mol. Neurosci. 16:1155620. doi: 10.3389/fnmol.2023.1155620
Received: 31 January 2023; Accepted: 03 April 2023;
Published: 20 April 2023.
Edited by:
Ismail Zaed, Humanitas University, ItalyReviewed by:
Surjeet Singh, Jackson Laboratory, United StatesJulie-Myrtille Bourgognon, University of Glasgow, United Kingdom
Copyright © 2023 Pate, Anthony and Radford-Smith. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel E. Radford-Smith, daniel.radford-smith@pharm.ox.ac.uk