 Laise de Moraes1,2
Laise de Moraes1,2 Luciane Amorim Santos1,2,3
Luciane Amorim Santos1,2,3 Liã Bárbara Arruda4
Liã Bárbara Arruda4 Maria da Purificação Pereira da Silva5Márcio de Oliveira Silva5José Adriano Góes Silva1,2,5André Ramos5Marcos Bastos dos Santos2
Maria da Purificação Pereira da Silva5Márcio de Oliveira Silva5José Adriano Góes Silva1,2,5André Ramos5Marcos Bastos dos Santos2 Felipe Guimarães Torres2Cibele Orge2Antonio Marcos dos Santos Teixeira2Thiago Santos Vieira2Laura Ramírez6
Felipe Guimarães Torres2Cibele Orge2Antonio Marcos dos Santos Teixeira2Thiago Santos Vieira2Laura Ramírez6 Manuel Soto6
Manuel Soto6 Maria Fernanda Rios Grassi2,3
Maria Fernanda Rios Grassi2,3 Isadora Cristina de Siqueira2Dorcas Lamounier Costa7
Isadora Cristina de Siqueira2Dorcas Lamounier Costa7 Carlos Henrique Nery Costa7
Carlos Henrique Nery Costa7 Bruno de Bezerril Andrade2,3
Bruno de Bezerril Andrade2,3 Kevan Akrami1,2
Kevan Akrami1,2 Camila Indiani de Oliveira1,2,3
Camila Indiani de Oliveira1,2,3 Viviane Sampaio Boaventura1,2,8
Viviane Sampaio Boaventura1,2,8 Manoel Barral-Netto1,2†
Manoel Barral-Netto1,2† Aldina Barral1,2†
Aldina Barral1,2† Anne-Mieke Vandamme9,10†
Anne-Mieke Vandamme9,10† Johan Van Weyenbergh9†
Johan Van Weyenbergh9† Ricardo Khouri1,2,9*
Ricardo Khouri1,2,9*- 1Programa de Pós-graduação em Ciências da Saúde, Faculdade de Medicina da Bahia, Universidade Federal da Bahia, Salvador, Brazil
- 2Instituto Gonçalo Moniz, Fundação Oswaldo Cruz, Salvador, Brazil
- 3Escola Bahiana de Medicina e Saúde Pública, Salvador, Brazil
- 4Centre for Clinical Microbiology, Division of Infection & Immunity, University College London, London, United Kingdom
- 5Centro Estadual Especializado em Diagnóstico, Assistência e Pesquisa, Secretaria de Saúde do Estado da Bahia, Salvador, Brazil
- 6Departamento de Biología Molecular, Facultad de Ciencias, Centro de Biología Molecular Severo Ochoa, Consejo Superior de Investigaciones Científicas, Universidad Autónoma de Madrid, Madrid, Spain
- 7Laboratório de Leishmanioses, Instituto de Doenças Tropicais Natan Portella, Universidade Federal do Piauí, Teresina, Brazil
- 8Hospital Santa Izabel, Salvador, Brazil
- 9Department of Microbiology, Immunology and Transplantation, Rega Institute for Medical Research, Clinical and Epidemiological Virology, Leuven, Belgium
- 10Center for Global Health and Tropical Medicine, Instituto de Higiene e Medicina Tropical, Universidade Nova de Lisboa, Lisbon, Portugal
Visceral leishmaniasis is an opportunistic disease in HIV-1 infected individuals, unrecognized as a determining factor for AIDS diagnosis. The growing geographical overlap of HIV-1 and Leishmania infections is an emerging challenge worldwide, as co-infection increases morbidity and mortality for both infections. Here, we determined the prevalence of people living with HIV (PWH) with a previous or ongoing infection by Leishmania infantum and investigated the virological and immunological factors associated with co-infection. We adopted a two-stage cross-sectional cohort (CSC) design (CSC-I, n = 5,346 and CSC-II, n = 317) of treatment-naïve HIV-1-infected individuals in Bahia, Brazil. In CSC-I, samples collected between 1998 and 2013 were used for serological screening for leishmaniasis by an in-house Enzyme-Linked Immunosorbent Assay (ELISA) with SLA (Soluble Leishmania infantum Antigen), resulting in a prevalence of previous or ongoing infection of 16.27%. Next, 317 PWH were prospectively recruited from July 2014 to December 2015 with the collection of sociodemographic and clinical data. Serological validation by two different immunoassays confirmed a prevalence of 15.46 and 8.20% by anti-SLA, and anti-HSP70 serology, respectively, whereas 4.73% were double-positive (DP). Stratification of these 317 individuals in DP and double-negative (DN) revealed a significant reduction of CD4+ counts and CD4+/CD8+ ratios and a tendency of increased viral load in the DP group, as compared to DN. No statistical differences in HIV-1 subtype distribution were observed between the two groups. However, we found a significant increase of CXCL10 (p = 0.0076) and a tendency of increased CXCL9 (p = 0.061) in individuals with DP serology, demonstrating intensified immune activation in this group. These findings were corroborated at the transcriptome level in independent Leishmania- and HIV-1-infected cohorts (Swiss HIV Cohort and Piaui Northeast Brazil Cohort), indicating that CXCL10 transcripts are shared by the IFN-dominated immune activation gene signatures of both pathogens and positively correlated to viral load in untreated PWH. This study demonstrated a high prevalence of PWH with L. infantum seropositivity in Bahia, Brazil, linked to IFN-mediated immune activation and a significant decrease in CD4+ levels. Our results highlight the urgent need to increase awareness and define public health strategies for the management and prevention of HIV-1 and L. infantum co-infection.
1. Introduction
HIV is the etiological agent of Acquired Immunodeficiency Syndrome (AIDS), a slowly progressive disease characterized by chronic immune activation resulting from the loss of cell-mediated immune function (Costin, 2007; Perkins et al., 2015). Host genetic factors (e.g., HLA genotypes) and host immune response (e.g., release of proinflammatory cytokines and chemokines), as well as the presence of co-infections, can directly influence the rate of disease progression (Claiborne et al., 2015; Kouri et al., 2015; Perkins et al., 2015).
Neglected Tropical Diseases (NTDs) are endemic among the most impoverished populations in Africa, Asia and Latin America. Among these NTDs, visceral leishmaniasis (VL), caused by infection with Leishmania infantum, is associated with increased morbidity and mortality in people with HIV-1 (PWH) but has not been recognized as a diagnostic criterion for AIDS (van Griensven et al., 2014). The World Health Organization (WHO) identified the geographical overlap of both infections as an emerging challenge in successfully controlling the HIV-1 infection in countries considered endemic for leishmaniasis, such as Brazil (World Health Organization, 2015).
The few studies evaluating the prevalence of HIV-1 and L. infantum coinfection on a national level in Brazil (Sousa-Gomes et al., 2011; Coura-Vital et al., 2014) have highlighted regions endemic for leishmaniasis: Distrito Federal (Carranza-Tamayo et al., 2009), Ceará (Távora et al., 2015), Maranhão (Carvalho et al., 2013), Mato Grosso (Luz et al., 2018), Mato Grosso do Sul (Botelho and Natal, 2009), Minas Gerais (Souza et al., 2012; Cota et al., 2014), Pernambuco (Sousa et al., 2018), Piauí (Soares et al., 2008), Rio Grande do Norte (Lima et al., 2017), Sergipe (Santos et al., 2019), and Tocantins (Albuquerque et al., 2014). Bahia is also considered an endemic area for leishmaniasis, with an incidence rate of 0.8/100,000 inhabitants for VL reported in 2022 (Secretaria de Saúde do Estado da Bahia, 2022b). In addition, the increase of geographical overlap between HIV-1 and Leishmania infection in Bahia (Secretaria de Saúde do Estado da Bahia, 2022a, 2022b) has raised public health concerns. However, no epidemiological studies have attempted to investigate the prevalence of co-infection to date. Therefore, we investigated the seroprevalence of L. infantum in PWH in the state of Bahia, Brazil and attempted to identify virological and immunological factors associated with co-infection, supported by publicly available transcriptomic data.
2. Methods
2.1. Study population and design
A two-stage cross-sectional cohort (CSC) study (CSC-I, 1998–2013 and CSC-II, 2014–2015) was designed to determine the seroprevalence of L. infantum in PWH and to evaluate the association with demographic, clinical, virological, and immunological parameters (Figure 1). From 1998 to 2015, individuals with a new confirmed diagnosis of HIV-1 at the Specialized Center for Diagnosis, Care and Research (CEDAP), a state government public health reference service located in the city of Salvador, Bahia-Brazil, were included in this study. Their remaining blood sample was utilized for the outlined assays herein.
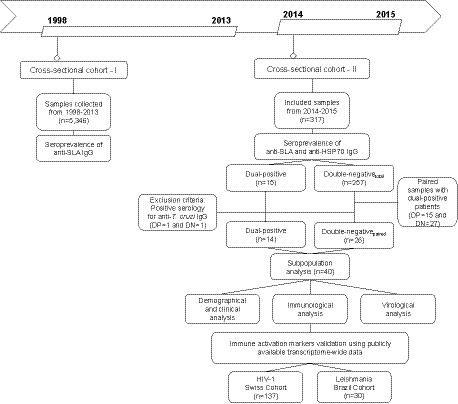
Figure 1. Flow chart of study design detailing the two-stage cross-sectional and prospective cohort study among people living with HIV in Bahia, Brazil.
First, serological reactivity to L. infantum was assessed in plasma or serum samples from the retrospective cross-sectional cohort (n = 5,346) collected between 1998 and 2013. In the second stage, serological reactivity to L. infantum was assessed in a prospective cross-sectional cohort of 317 PWH (sampled between July 2014 and December 2015), with demographic and clinical data (sex, age, HIV-1 viral load, CD4+, CD8+, CD4+/CD8+ ratio and CD45+ T cell counts) prospectively obtained from the clinical records. Additionally, we performed a subpopulation analysis on a subset of PWH identified as either double-positive or double-negative using two independent anti-Leishmania immunoassays. Based on these stringently defined categories, we compared virological and immunological factors influenced by L. infantum seropositivity. The study’s recruitment did not interfere with standard medical care, and patients were treated according to the general Brazilian/WHO guidelines. This study was conducted following the Declaration of Helsinki and was approved by the Institutional Review Board of the Gonçalo Moniz Institute (IGM-FIOCRUZ) (protocol number 1.764.505).
2.2. Detection of anti-Leishmania and anti-Trypanosoma antibodies
All samples underwent a previously described and validated in-house enzyme-linked immunosorbent assay (ELISA) for the detection of IgG against Soluble Leishmania infantum Antigen (SLA), which we previously demonstrated as a sensitive biomarker (90% sensitivity and 95% specificity) of current or past infection with the parasite (Charlab et al., 2000). SLA preparation and ELISA procedures were performed as previously described (Souza et al., 2013). For the prospective cohort of 317 samples from PWH with digitalized clinical records, results of the SLA immunoassay were compared with a second, previously described ELISA employing recombinant L. infantum heat shock protein 70 (HSP70), for which we once found a sensitivity of 74% and a specificity of 73% (Soto et al., 2015). To avoid false-positive results for L. infantum due to cross-reactivity among Trypanosomatidae parasites, an ELISA for anti-Trypanosoma cruzi IgG (Euroimmun) was employed in a subset of 42 samples with dual-positive (DP) (n = 15) and double-negative (n = 27) serology for anti-SLA and anti-HSP70.
2.3. Control samples and ELISA cutoff value
Positive (n = 6) Leishmania-infected patient samples and negative (n = 47) control samples previously identified by the intradermal Leishmanin (also known as Montenegro) Delayed-type Hypersensitivity (DTH) skin test (obtained from Fundação Ezequiel Dias, Belo Horizonte, Brazil), with clinical and laboratorial confirmation, were used to establish anti-SLA and anti-HSP70 ELISA cutoff values. For each assay, specific cutoff values were determined using the mean OD values obtained from the included negative samples plus three times the observed standard deviation, with each assay’s index values corresponding to the OD/cutoff ratio (Supplementary Figure S1).
2.4. Quantification of immunological markers
A Cytometric Bead Array (CBA) Human Inflammatory Cytokines Kit (BD Biosciences) was employed to quantify plasma concentrations of IL-1β, IL-6, IL-10, IL-12p70, and TNF, while CCL2/MPC-1, CCL5/RANTES, CXCL8/IL-8, CXCL9/MIG, and CXCL10/IP-10 quantification was performed using a CBA Human Chemokine Kit (BD Biosciences), each following the manufacturer’s instructions. Data acquisition and analysis were performed using a FACSArray Bioanalyzer (BD Biosciences) and FlowJo v10.0.5 (Tree Star) software, respectively.
2.5. HIV-1 sequencing analyses
Viral RNA isolation was performed using a QIAamp Viral RNA Mini Kit (QIAGEN) following the manufacturer’s instructions. The protease/reverse transcriptase (PR/RT) region was amplified and sequenced as previously described (Barreto et al., 2006). Outer polymerase chain reaction (PCR) was performed using a SuperScript III One-Step RT-PCR System with Platinum Taq DNA Polymerase (Thermo Fisher Scientific) and the following primers: K1 (CAGAGCCAACAGCCCCACC) and K2 (TTTCCCCACTAACTTCTGTATGTCATTGACA) (Kozal et al., 1996). Inner PCR was performed using Platinum Taq DNA Polymerase High Fidelity (Thermo Fisher Scientific) with the following primers: DP16 (CCTCAAATCACTCTTTGGCAAC) and RT4 (AGTTCATAACCCATCCAAAG) (Pieniazek et al., 1991). The generated inner PCR products were sequenced using a BigDye Terminator v.3.1 Cycle Sequencing Kit (Applied Biosystems) with capillary electrophoresis run on an ABI 3500xL Genetic Analyzer (Applied Biosystems) employing the following primers: F1 (GTTGACTCAGATTGGTTGCAC), F2 (GTATGTCATTGACAGTCCAGC) (Frenkel et al., 1995), DP10 (CAACTCCCTCTCAGAAGCAGGAGCCG), DP11 (CCATTCCTGGCTTTAATTTTACTGGTA) (Janini et al., 1996), RT4 (AGTTCATAACCCATCCAAAG), GABO1 (CTCARGACTTYTGGGAAGTTC), and GABO2 (GCATCHCCCACATCYAGTACTG) (Barreto et al., 2006).
Sequence visualization, editing and assembly were performed using Geneious v10.0.8 (Dotmatics) software. HIV-1 subtyping was determined using the REGA HIV-1 Subtyping Tool v.3.4.11 and the jpHMM-HIV approach (Schultz et al., 2009). Phylogenetic analysis was conducted using HIV-1 subtype references retrieved from the Los Alamos HIV sequence database2 (Supplementary Table S1). Phylogenetic inference was performed using PhyML v3.0 (Guindon et al., 2010), applying the BIONJ method and using the General Time Reversible (GTR) nucleotide substitution model with 1,000 bootstrap replicates.
2.6. Transcriptome and systems biology analysis
Transcriptome analysis of publicly available data was performed as recently described (Vivarini et al., 2017; Delgobo et al., 2019; Subramanian et al., 2019). Overlap ratio was calculated as: k/K = # Genes in Overlap (k)/# Genes in Gene Set (K) using Molecular Signatures Database (MSigDb). HIV-1 (Swiss HIV Cohort, n = 137) (Rotger et al., 2010) and Leishmania-infected individuals (Piaui Northeast Brazil Cohort, n = 30) (Gardinassi et al., 2016) were analyzed using GEO2R from Gene Expression Omnibus (GEO).
2.7. Statistical analysis
Since data were not normally distributed, as identified by the Shapiro–Wilk and D’Agostino-Pearson tests (GraphPad Prism v7.0), results were analyzed using non-parametric tests: Mann–Whitney test and Spearman’s correlation, as well as Fisher’s exact test, with p < 0.05 considered statistically significant.
3. Results
3.1. Detection of IgG antibodies against Leishmania infantum antigens (anti-SLA and anti-HSP70) in people living with HIV-1
To determine the seroprevalence of L. infantum among PWHs, 5,346 plasma or serum samples from treatment-näive HIV-1-infected individuals diagnosed between 1998 and 2013 were tested for the presence of IgG antibodies against soluble L. infantum antigen (anti-SLA) (Figure 1).
Positivity for anti-SLA serology was detected in 870/5,346 samples, resulting in a prevalence rate of 16.27% (Figure 2). We included additional positive control samples from patients with a clinically confirmed diagnosis of visceral leishmaniasis (VL) and laboratory-confirmed L. infantum infection in the absence (n = 16) or presence of HIV-1 co-infection (n = 10) (Zacarias et al., 2017). The detection rate of anti-SLA antibodies of the positive control samples was 81.25% for HIV-1−VL+ patients and 100% for HIV-1+VL+ patients (Figure 2). Besides, we found humoral immune response against SLA in VL patients quantitatively similar regardless of the presence of HIV-1 co-infection (Mann Whitney p = 0.71, HIV-negative vs. HIV-positive). Thus, we could confirm the high specificity of the L. infantum anti-SLA IgG serology protocol employed herein.
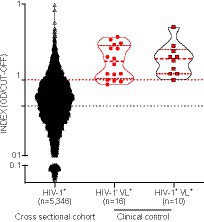
Figure 2. Detection of anti-L. infantum IgG antibodies in plasma/serum samples collected from PWH between 1998 and 2013. Violin plot with a dot-plot overlay of OD index values representative of L. infantum anti-SLA serology in the retrospective PWH cohort (n = 5,346) and a clinical- and laboratory-confirmed control group (HIV-1-VL+, n = 16; and HIV-1-VL+, n = 10). Filled black circles (●): PWH cross-sectional cohort samples; open triangles (Δ): clinical HIV-1 negative controls with VL; filled red squares with black borders (▪): clinical controls with HIV-1 positive controls and VL.
Subsequently, to validate our findings, we enrolled an independent prospective CSC between 2014 and 2015, composed of 317 treatment-näive HIV-1-infected individuals from which complete demographic and clinical data were obtained from digitalized clinical records. Moreover, we combined anti-SLA serology with a second specific assay to detect IgG antibodies against L. infantum HSP70 recombinant protein (anti-HSP70) (Figure 1). The overall prevalence of anti-SLA positive serology in the prospective cohort was 15.46% (49/317), similar to the cross-sectional cohort results. The prevalence of anti-HSP70 positive serology in the prospective cohort was 8.20% (26/317) (Figure 3). Positivity for both anti-SLA and anti-HSP70 (i.e., double-positivity – DP) was detected in 4.73% (15/317) of the cohort; anti-SLA positive serology alone was detected in 10.73% (34/317); anti-HSP70 serology alone in only 3.47% (11/317) and the union of both anti-SLA and anti HSP70 positive serology was detected in 18.93% (Figure 3). Important, no statistical differences (Kruskal–Wallis test) were observed in viral load (p = 0.95), CD4+ (p = 0.93), CD8+ (p = 0.59), CD4+/CD8+ (p = 0.92), and CD45+ (p = 0.89) T cells values comparing DP, SLA, HSP70, and HSP70+SLA groups (Supplementary Table S6).
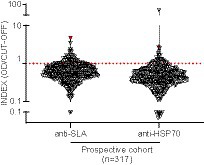
Figure 3. Detection of anti-L. infantum IgG antibodies in plasma samples collected from PWH between 2014 and 2015. Violin plot with a dot-plot overlay of OD index values representative of L. infantum anti-SLA and anti-HSP70 serology in the PWH prospective cohort (n = 317). The red symbols represent the DP individual diagnosed with visceral leishmaniasis >20 years ago.
Based on these findings, DP (n = 15) was adopted as the most stringent criterium for L. infantum infection. Assessment of standard diagnosis of previous or ongoing visceral leishmaniasis (VL) documented in the clinical records showed that one of the 15 DP subjects had VL reported >20 years ago (anti-SLA index = 4.71; anti-HSP70 index = 2.67).
After two decades of clinical diagnosis, this pronounced humoral response most likely reflects chronic antigen exposure, supporting the concept that Leishmania infection is lifelong in most patients as observed in murine models of disease (Bogdan et al., 2000; Seyed et al., 2018; Paduch et al., 2019). These results, taken together, demonstrated that a significant number of PWH in the state of Bahia are unaware of their exposure to and/or infection by L. infantum and are at risk of developing new or relapsing visceral leishmaniasis.
3.2. Demographic and clinical markers associated with dual-positive L. infantum serology in PWH
The CSC-II (n = 317) consisted of predominantly young males (72.23%) with a median age of 32 years old at the time of HIV diagnosis, median viral load of 40,370 copies/mL (IQR = 7,032-141,359), CD4+ T cell count of 369 cells/μL (IQR = 163–609), CD8+ T cell count of 1,025 cells/μL (IQR = 715–1,441), and CD45+ T cell count of 2,144 cells/μL (IQR = 1,507–2,763) (Table 1).
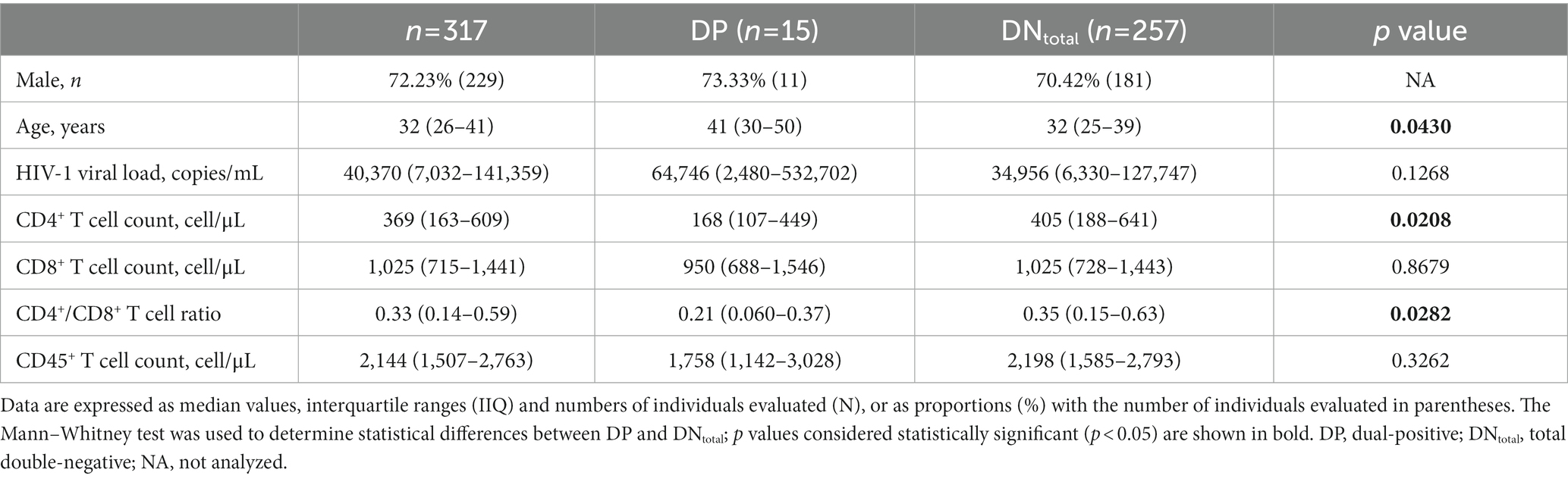
Table 1. Epidemiological, virological, and immunological characteristics of PWH in Bahia (Brazil) between 2014 and 2,105, stratified according to DP and DNtotal.
Comparative analyses between the DP (n = 15) and total double-negative (DNtotal) (n = 257) groups indicated that individuals with antibodies against both SLA and HSP70 L. infantum were older (p = 0.0430), presented lower baseline CD4+ T cell counts (p = 0.0208) and lower CD4+/CD8+ T cell ratios (p = 0.0282), as well as a tendency towards higher HIV-1 viral load levels (p = 0.1268) suggestive of an advanced stage of HIV-1 infection than individuals without L. infantum antibodies (Table 1). Indeed, a large proportion of DP individuals (10/15, 66.67%) had <200 CD4+ T cells/μL at the time of HIV-1 diagnosis, which is considered AIDS-defining.
3.3. Subpopulation analysis reveals virological and immunological markers associated with dual-positive L. infantum serology in PWH
To ensure specificity for L. infantum in the DP serological samples and to exclude the possibility of cross-reactivity due to polyclonal activation and hypergammaglobulinemia, anti-Trypanosoma cruzi IgG serology was performed. Samples demonstrating positivity for anti-T. cruzi IgG (n = 2) were excluded from subsequent analyses (Supplementary Figure S2). Next, we performed a subpopulation analysis by matching DN individuals to the DP group by gender, age, viral load and CD4+ T cell count. This strategy resulted in the establishment of a third group (DNpaired) (Figure 1), thereby mitigating the possibility of bias arising from polyclonal activation, hypergammaglobulinemia, high viral load or low CD4+ T cell levels. Controlling for these parameters limited the potential for variations in the HIV-1 transcriptomic profile and the state of immune activation. No statistical differences were observed between the DP and DNpaired groups regarding sociodemographic and clinical parameters, which underscores the pairing strategy’s equity (Table 2).
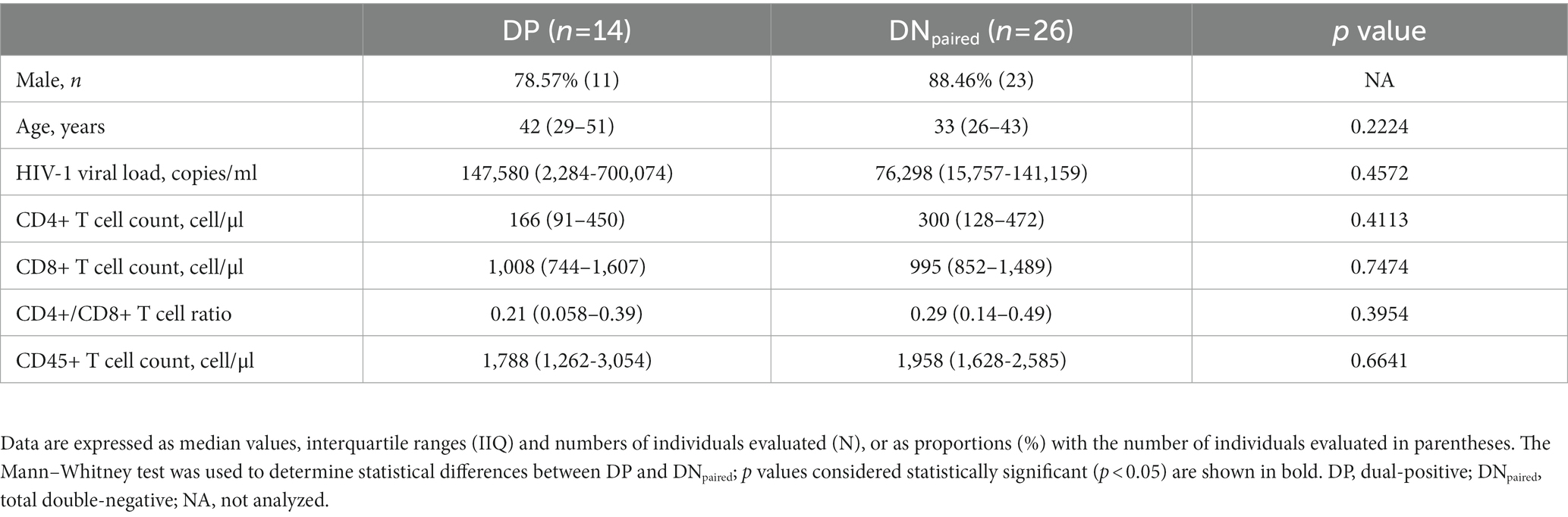
Table 2. Epidemiological, virological, and immunological characteristics of PWH in Bahia (Brazil) between 2014 and 2,105, stratified according to DP and DNpaired.
3.4. Subpopulation analysis reveals virological and immunological markers associated with dual-positive L. infantum serology in PWH
To investigate possible associations between transmission clusters of a more-aggressive HIV subtype and worsen HIV clinical status, HIV-1 subtyping was performed in the DP and DNpaired subset to infer. After excluding samples with a viral load <1,000 copies/mL, both groups were pooled together (n = 19) and submitted to phylogenetic analysis using the jpHMM-HIV model (Supplementary Figure S3). Concerning subtype distribution, subtype B (63.16%, 12/19) was the most prevalent, followed by subtypes C (15.79%, 3/19), recombinant BF (15.79%, 3/19), and D (5.26%, 1/19). No statistical differences in subtype were observed between the DP and DNpaired groups (p = 0.67) (Supplementary Table S2). No transmission cluster was identified. HIV-1 subtyping showed no impact on L. infantum serology titers, viral load, T cell count, or any other host immunological marker evaluated, indicating that HIV-1 subtypes B, C, D, F1, and BF exhibit similar virological and immunological outcomes in HIV/L. infantum co-infection.
Next, we quantified circulating cytokines and chemokines that might reflect an increase in systemic immune activation and have been previously associated with infection to L. infantum (Caldas et al., 2005; Peruhype-Magalhães et al., 2006; Frade et al., 2011; Carneiro et al., 2016). The systemic levels of cytokines tested (IL-1β, IL-6, IL-10, IL-12p70, and TNF) were below the detection limits in all samples (DP and DNpaired groups), concordant with our previous findings in a large Cuban cohort of PWH (Kouri et al., 2015). Conversely, most chemokines (except IL-8) were readily detected in most PWH, demonstrating a significant increase in CXCL10/IP-10 (p = 0.0076) and a tendency towards higher CXCL9/MIG levels (p = 0.061) (Figure 4) in the DP group compared to DNpaired, suggesting increased immune activation in the group with DP serology for L. infantum.
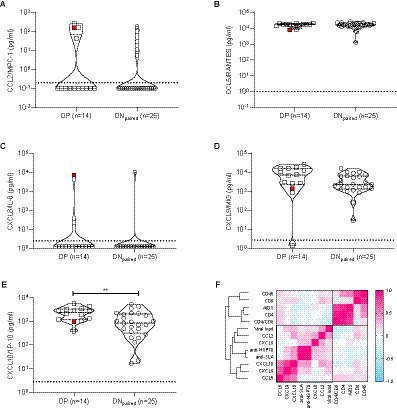
Figure 4. Chemokine profiles obtained from plasma/serum samples of DP and DNpaired groups. (A)–(E) Violin plots with dot-plot overlays illustrating chemokine plasma levels (pg/mL). Statistical comparisons made by the Mann–Whitney test in plasma levels (pg/mL) of CCL2/MPC-1 (p = 0.43), CCL5/RANTES (p = 0.94), CXCL8/IL-8 (p = 0.54), CXCL9/MIG (p = 0.061), and CXCL10/IP-10 (p = 0.0076) chemokines. Open squares (): subgroup of double-positive PWH (positive for both anti-SLA and anti-HSP70 L. infantum antigens); open circle (○): subgroup of paired double-negative PWH (serologically negative for anti-SLA and anti-HSP70 L. infantum antigens). The red symbol represents the DP individual diagnosed with visceral leishmaniasis >20 years ago. (F) Heat map shows the correlation (pink: positive correlation; blue: negative correlation; white: no correlation) of epidemiological, virological and immunological characteristics of PWH.
This finding, together with the decreased CD4+ T cell counts found in our previous analysis (Table 2), corroborates the hypothesis that therapy-naïve HIV-1-infected subjects previously or currently infected with L. infantum exhibit a more intensified state of immune activation than HIV-1 positive, L. infantum seronegative subjects. During clinical follow-up, the introduction of antiretroviral therapy restored CD4+ T cell counts (median values of CD4+ T cell counts after 4–5 years of follow-up: DP = 620 cells/μL and DNpaired = 870 cells/μL) and reduced viral load levels to undetectable limits similarly in each group (Figure 5), with no statistically significant differences between groups (AUC comparison, Mann–Whitney test: VL: p = 0.6773; CD4+: p = 0.5239; CD8+: p = 0.6986; Ratio CD4+/CD8+: p = 0.9044; CD45+: p = 0.7540).
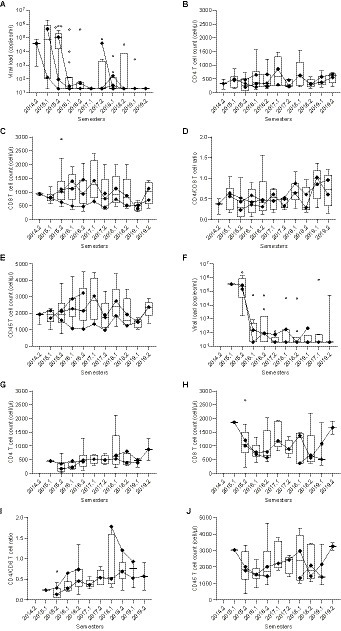
Figure 5. Long-term follow-up of CD4+, CD8+, CD4/CD8 ratio, CD45+, and viral load levels after therapy introduction in DP and DNpaired groups. (A)–(J) Box plots with line plot overlays connecting medians of CD4+, CD8+, CD4+/CD8+ ratio, CD45+ and viral load levels during follow-up. (A) and (F) Follow-up of viral load (copies/mL) in DNpaired (A) and DP (F) groups. (B) and (G) Follow-up of CD4+ T cell count (cells/μL) in DNpaired (B) and DP (G) groups. (C) and (H) Follow-up of CD8+ T cell count (cells/μL) in DNpaired (C) and DP (H) groups. (D) and (I) Follow-up of CD4+/CD8+ (ratio) in DNpaired (D) and DP (I) groups. (E) and (J) Follow-up of CD45+ T cell count (cells/μL) in DNpaired (E) and DP (J) groups.
3.5. CXCL10 is shared between the HIV and visceral leishmaniasis gene signatures, while CXCL10, CXCL9, and CCL2 positively correlate with HIV viral load in untreated PWH
Given the small number of immune activation markers and modest cohort size for the paired DP and DN groups, we aimed to validate and extend our findings by cross-examining independent cohorts of HIV-1 (Swiss HIV Cohort, n = 137) or Leishmania-infected individuals with and without active disease (Piaui Northeast Brazil, n = 30) with publicly available transcriptome-wide data (Rotger et al., 2010; Gardinassi et al., 2016). As shown in Figures 6A,B, a significant overlap (41 genes, enrichment p < 0.0001) was found between the HIV and visceral leishmaniasis gene signatures when comparing the top 500 transcripts significantly upregulated during active disease in both untreated HIV patients and untreated visceral leishmaniasis patients, as compared to their paired post-treatment groups. A systems biology analysis of the 41 genes shared by HIV and leishmaniasis active disease signatures revealed several significantly enriched biological processes, encompassing immune activation broadly and, more specifically, type I IFN signaling and antiviral response. The overlapping genes included CXCL10, which we found to be overexpressed in the DP group of our prospective cohort, as well as STAT1, the major transcription factor activated by IFN signaling. As detailed in Figure 6A and Supplementary Tables S3–S5, enriched gene sets from the Molecular Signatures Database (MSigDb) representing ‘immune activation’ included graft-versus-host disease, antigen recognition and blister cytotoxicity, while enriched gene sets representing ‘type I IFN/antiviral response’ comprised those upregulated in the liver by HBV infection, by IFN-alpha treatment of fibroblasts in vitro and by IFN-beta treatment of multiple sclerosis patients in vivo. Of interest, IFNG transcripts significantly decrease in monoinfected visceral leishmaniasis individuals, confirming our previous results obtained at the protein level for systemic IFN-γ in two independent VL cohorts (Caldas et al., 2005; Peruhype-Magalhães et al., 2006).

Figure 6. CXCL10 is shared between the HIV and visceral leishmaniasis disease signatures while CXCL10, CXCL9, and CCL2 transcripts positively correlate with HIV viral load in untreated PWH. Venn diagrams show overlap between (A) active disease gene signatures, i.e., genes upregulated in untreated PWH (Swiss cohorts) and visceral leishmaniasis patients (as compared to post-treatment) and (C) genes upregulated in untreated PWH and asymptomatic Leishmania-infected individuals (who did not develop disease). Systems biology analysis shows a significant enrichment of gene sets comprising immune activation and type I IFN signaling (right-hand panels). (B) Selective up-regulation of CXCL10 and STAT1 (as a positive control for IFN signaling) transcripts in both untreated visceral leishmaniasis patients (upper panel) and L. infantum-exposed individuals and (D) Selective up-regulation (upper panel) and positive correlation to viral load (lower panel) of CXCL10, CXCL9, and CCL2 and STAT1 (as a positive control for IFN signaling) transcripts in untreated PWH (Swiss cohort, n = 137).
To define a specific ‘Leishmania infection’ gene signature, we used the top 500 significantly upregulated transcripts from Leishmania-infected individuals who did not develop disease, for which Leishmania infection was documented by a positive skin DTH reaction, as compared to matched DTH-negative individuals from the same endemic area (Gardinassi et al., 2016). Surprisingly, the overlap between the ‘untreated HIV’ and ‘Leishmania-seropositive (DP)’ gene signatures was even more pronounced than the overlap between the ‘untreated HIV’ and ‘untreated visceral leishmaniasis’ gene signatures, both by quantitative (65 shared genes, Figure 6B) and qualitative measures (median gene set overlap 9.4% vs. 3.2%, Mann–Whitney p < 0.0001, Supplementary Table S3), indicating that even asymptomatic Leishmania infection has a profound effect on the systemic immune response. Again, CXCL10 and STAT1 were shared among the two disease signatures (Figure 6B), and enriched gene sets included type I IFN signaling (genes upregulated by IFN-beta treatment of multiple sclerosis patients in vivo and by IFN-alpha treatment of fibroblasts in vitro) and immune activation (inflammatory response to LPS, TNF signaling).
Validation of our DP cohort results revealed that CXCL10, but not CCL2, CCL5 or CXCL8 transcripts, were significantly upregulated by Leishmania infantum infection with or without clinically definite visceral leishmaniasis (Figure 6C upper and lower panel). In the Swiss HIV Cohort, we found that CXCL10, CCL2 and, to a lesser extent, CXCL9 transcripts were significantly upregulated during untreated HIV-1 infection. In contrast, no significant effect was observed for CXCL8 in either asymptomatic or symptomatic Leishmania infantum infection, supporting our negative findings for CXCL8 genetic variation in visceral leishmaniasis patients from the same endemic region (Frade et al., 2011). However, CXCL8 transcripts were significantly downregulated during untreated HIV-1 infection (Figure 6D upper panel). Finally, as shown in Figure 6D (lower panel), CCL2, CXCL9, and CXCL10 transcripts were significantly correlated (even after stringent correction for genome-wide testing) with set point viral load in untreated PWH from the Swiss HIV Cohort (n = 137), confirming the results from our prospective cross-sectional cohort (Figure 4F).
4. Discussion
The geographic overlap of HIV-1 and L. infantum infections raises the possibility of co-infection, which several groups have shown to increase morbidity and mortality (Coura-Vital et al., 2014; Lima et al., 2017, 2018; Viana et al., 2017; Diniz et al., 2018; Fontoura et al., 2018; Luz et al., 2018; Rezaei et al., 2018; Sousa et al., 2018; Botana et al., 2019; Diro et al., 2019; Horrillo et al., 2019; Maia-Elkhoury et al., 2019; Prestes-Carneiro et al., 2019; Santos et al., 2019; Silva de Lima et al., 2019; van Griensven et al., 2019). Several groups have studied the extent of this overlap in several Brazilian regions. However, no studies have attempted to determine the seroprevalence of L. infantum among PWH in the state of Bahia, despite the significant prevalence of both infections (Secretaria de Saúde do Estado da Bahia, 2022a, 2022b). We used a two-stage cross-sectional cohort approach (CSC-I, n = 5,346 and CSC-II, n = 317) and identified positive serology for L. infantum ranging from 4.73% (DP for anti-SLA and anti-HSP70) to 18.93% in the CSC-II (union of all positive samples for anti-SLA and anti-HSP70), with an intermediate seroprevalence for only anti-SLA of 16.27% in the CSC-I and 15.46% in the CSC-II.
The observed frequencies of Leishmania seroprevalence in PWH using any of the serology strategies in both the cross-sectional cohorts support previous findings falling within the range of what has already been described in other Brazilian regions – Ceará: 5.4–6.6% (Távora et al., 2015); Distrito Federal: 16.0% (Carranza-Tamayo et al., 2009); Maranhão: 4.2% (Carvalho et al., 2013); Mato Grosso: 9.9% (Luz et al., 2018); Mato Grosso do Sul: 5% (Botelho and Natal, 2009); Pernambuco: 5.6% (Sousa et al., 2018); Piauí: 8.2% (Soares et al., 2008); Rio Grande do Norte: 4.9% (Lima et al., 2017); Sergipe: 4.5% (Santos et al., 2019); and Tocantins: 2.1% (Albuquerque et al., 2014). Moreover, demographic data indicated that all DP patients were born in the macroregions surrounding the cities of Salvador and Feira de Santana, which are responsible for 33.8% (1,074/3,172) of notified VL cases in Bahia between 2007 and 2015 (Secretaria de Saúde do Estado da Bahia, 2018).
The observed discrepancy between the results obtained with the two different immunoassays might be explained by the rigor employed to determine cutoffs for each serological test and the imbalance in protein fractions between these two antigen-based diagnostic methods. SLA is a heterogeneous mix of antigens and has a lower concentration of HSP70 than parasite structural proteins (actin, tubulin, etc.). Even though it will pick up a broader set of samples, some samples with low antibody levels will fail. On the other hand, the recombinant HSP70 antigen is homogenous and highly concentrated and has a better detection limit with better sensitivity than other recombinant proteins (rH2A, rH2B, rH3, rH4, and rKMP11) and high specificity when evaluated against sera from patients with Chagas’ disease, Tuberculosis, Leprosy or Systemic Lupus Erythematosus (Souza et al., 2013). We considered only those samples demonstrating dual positivity to increase confidence regarding past or present L. infantum infection. This strategy was underscored by the identification of one individual with a previous history of VL in the DP group and no cases with an earlier history of VL in the DNpaired group. Furthermore, we confirmed that no statistical differences regarding epidemiological, virological, and immunological characteristics were observed comparing DP, SLA, HSP70 and HSP70+SLA groups.
Unfortunately, only serum or plasma samples were available for HIV-1 RNA extraction, thus precluding DNA analysis to detect L. infantum by qPCR, which we acknowledge as one of the limitations of our study. However, Fukutani et al. (2014) demonstrated a concordance of 68% between anti-SLA serology and PCR, and found a 5.40% rate of ongoing asymptomatic L. infantum infection among blood donors in Salvador, Bahia. Of note, this concordance between PCR and serology could be compromised by the reduced serology sensitivity (66%) to identify true positive ongoing Leishmania-infected cases among PWH. In addition, we also highlight as a limitation the absence of Leishmanin skin test in our study (DTH) to access cellular immune response present during Leishmania infection, which is more sensitive and specific to determine ongoing asymptomatic infection than serological assays (Botana et al., 2019). Nevertheless, positive IgG serology for L. infantum indicates immunological memory of at least a previous Leishmania infection before HIV diagnosis. Strikingly, we identified a DP individual with a pronounced humoral response two decades after a VL clinical diagnosis. This ongoing humoral response most likely reflects chronic antigen exposure, emphasizing the concept that Leishmania infection is lifelong in most patients, as well as murine models of disease (Bogdan et al., 2000; Seyed et al., 2018; Paduch et al., 2019). A wealth of studies worldwide (Oryan and Akbari, 2016; Grande et al., 2017; Zacarias et al., 2017; Bruhn et al., 2018; Guedes et al., 2018; Luz et al., 2018) and a recent systematic review (Fontoura et al., 2018) have demonstrated a high frequency of relapse of VL in PWH, as well as increased mortality. Relapse of VL has been reported after anti-leishmaniasis treatment, even years after the patient was considered clinically cured, especially in PWH and/or immunosuppressive conditions (Horrillo et al., 2019; Silva de Lima et al., 2019).
Prior large studies, including from our group, have found striking differences in the prevalence of VL within different regions in Brazil, as well as Southern Europe. The highest seroprevalence was observed in Mato Grosso, 41.40% of blood donors, in contrast to other regions in Brazil [Salvador: 5.40% of 700 (Fukutani et al., 2014); Paraná: 11.40% of 176 (Braga et al., 2015); Fortaleza: 17.10% of 431 (Monteiro et al., 2016)], and in Southern Europe [France: 13.40% of 565 (le Fichoux et al., 1999); Spain: 3.10% of 1,437 (França et al., 2018)].
Studies conducted in populations of newly diagnosed HIV-1-infected individuals from regions endemic for leishmaniasis in Brazil reported a prevalence of HIV-1/L. infantum co-infection ranging from 4.20 to 20.20%. In these studies, L. infantum infection was determined by parasitological, serological and/or molecular testing (Soares et al., 2008; Carranza-Tamayo et al., 2009; Orsini et al., 2012; Carvalho et al., 2013). However, in co-infection studies focused on VL patients, which were diagnosed by parasitological, molecular or immunofluorescence methods, the prevalence of HIV-1/L. infantum coinfection ranged from 36.60 to 55.60% (Souza et al., 2012; Cota et al., 2014; Druzian et al., 2015).
The present study also investigated possible differences in the prevalence of HIV-1 variants associated with L. infantum infection in PWH. With these subtyping results, we would like to investigate if a possible worsen disease progression in PWH with positive serology for L. infantum cases was related to cluster of transmission of a more-aggressive HIV subtype (e. g., HIV Subtype D) (Khouri and Vandamme, 2015). However, no differences were found concerning HIV-1 isolates between matched samples of patients infected (DP) or not (DNpaired) to L. infantum. Within these isolates, subtype B (63.16%) was found to be the most prevalent, followed by C (15.79%), recombinant BF (15.79%) and D (5.26%). Recent studies in Bahia that evaluated HIV-1 variant distribution have described the wide distribution of subtype B, followed by a smaller proportion of recombinant BF units (Araujo et al., 2010; Monteiro-Cunha et al., 2011; Santos et al., 2011). Of note, the higher prevalence of subtype C (15.79%) found herein stands in contrast to other studies also conducted in this region, which reported a very low prevalence ranging from 1.70 to 2.50% (Chavez et al., 2007; Santos et al., 2009; Monteiro-Cunha et al., 2011).
Moreover, our confirmation of the circulation of subtype D in the state is following work by Monteiro et al. (2009), who previously reported the presence of subtype D in individuals from Feira de Santana (0.60% of polF/envD subtype), a city located ~100 km from Salvador. It is worth noting that the subtyping analysis performed herein involved three different methodologies (REGA HIV-1 Subtyping Tool, phylogeny and jpHMM-HIV probabilistic modeling), which lends support to the presently reported distribution. Furthermore, our subtype identification was performed using HIV-1 pol sequences, inferring subtype frequencies can vary when complete genome sequencing is performed.
Prior research into host immune responses has demonstrated that L. infantum infection induces a higher level of immune activation in co-infected PWH, which could lead to a more rapid progression to AIDS (Ipp et al., 2014; Claiborne et al., 2015; Hoffmann et al., 2016; Utay and Hunt, 2016). Our study found a roughly five-fold increase in circulating CXCL10/IP-10 and CXCL9/MIG chemokines in PWH exposed to L. infantum, supporting a link between positive serology for L. infantum and immune activation, which we validated at the transcript level for CXCL10 in independent HIV-1 and Leishmania-infected cohorts. These chemokines are strongly induced by both type I (IFN-α/β) and type II IFN (IFN-γ) (Kedzierska and Crowe, 2001; Mehla, 2013; Kenway-Lynch et al., 2014; Utay and Douek, 2016). It has been previously reported that Th1 subpopulations of CD4+ CD95+ T cells in HIV-1 infected individuals are highly susceptible to HIV-1 envelope gp120-induced apoptosis (Accornero et al., 1997; Cummins and Badley, 2010), which may corroborate the deleterious role of intensified immune activation (Costa et al., 2012; Sun et al., 2015; Rodrigues et al., 2016). In a submitted manuscript (Khouri et al.), we demonstrated an increase in both systemic soluble Fas as well as membrane-bound Fas/CD95+ expression in CD8+ cells in HIV/VL co-infection, as compared to HIV-1 or VL mono-infected groups, in a large cohort of individuals recruited during an outbreak of VL in Piaui, another state in the same northeast region of Brazil. Since we previously demonstrated STAT1-mediated IFN signaling as a preferential inducer of Fas/CD95+ at both the protein and transcript level (Farre et al., 2008; Khouri et al., 2018), this study as well as Khouri et al. (submitted) underscore the pivotal role of IFN-driven immune activation in HIV-1 and Leishmania infantum co-infection and its deleterious effect on both AIDS progression and visceral leishmaniasis.
The present study aimed to describe the prevalence and consequences of L. infantum infection in a population of antiretroviral treatment-naïve PWH at diagnosis. We found a high seroprevalence of L. infantum in treatment-naïve PWH in Bahia (Brazil) associated with a more intensified state of immune activation as compared to HIV-1 mono-infected subjects, possibly exacerbating progression to AIDS, which was observed in 66.67% in co-infected individuals at diagnosis. This immune activation and disease progression in PWH is reversed by immediate ART implementation (Buzón et al., 2010). Brazil adopted therapy as prevention (TasP) for HIV in 2013, which through our National Health System (SUS) offers and treats every HIV diagnosed patient in the country, independent of the CD4+ T cell level. However, in Brazil, Visceral Leishmaniasis cases are investigated only when severe VL-like symptoms are already present, precluding ongoing asymptomatic or mild visceral leishmaniasis diagnosis. Thus, Leishmania-infected and ART-naïve PWH with low CD4+ T cells (<200 cell/μL) and high viral load are at high risk of developing severe visceral leishmaniasis (Viana et al., 2017). These findings underscore the urgent need to increase awareness and define public health strategies for the management and prevention of HIV-1 and L. infantum co-infection.
Data availability statement
The data presented in this study have been deposited in the NCBI GenBank under accession numbers MW596907, MW596966, MW597006, MW597017, MW597019, MW597022, MW597023, MW597030, MW597036, MW597044, MW597056, MW597059–MW597061, MW597063, MW597068, MW597073–MW597075.
Ethics statement
The studies involving human participants were reviewed and approved by Institutional Review Board of the Gonçalo Moniz Institute (IGM-FIOCRUZ) (protocol number 1.764.505). The patients/participants provided their written informed consent to participate in this study.
Author contributions
RK and JW contributed to conception, design of the study and funding acquisition. LM, MP, FT, MBS, AT, and TV contributed to data curation. LM, LS, LA, CO, and LR performed formal analysis. MOS, AR, IS, JS, DC, CC, and MG were responsible for the investigation. RK and JW developed the methodology. AB, A-MV, JV, MS, MOS, AR, DC, and CC provided resources. JW, RK, and LM contributed to visualization. LM and RK wrote the first draft of the manuscript. RK, A-MV, JV, MB-N, AB, LS, LA, VB, MS, BA, KA, and CO wrote sections of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001, RK; Fundação de Amparo à Pesquisa do Estado da Bahia (FAPESB, grant APP0032/2016, RK); Brazilian National Council for Scientific and Technological Development (CNPq, grant 65083/2015-8, LS); FAPESB/CNPq (008/2014 PRONEX grant 8111/2014, AB); Fonds voor Wetenschappelijk Onderzoek Vlaanderen (grant G0D6817N, A-MV and JW). None of the funding organizations had any role in the study design, data collection, data interpretation or writing of this report.
Acknowledgments
The authors would like to thank Andris K. Walter for his assistance with English language revision and manuscript copyediting, as well as the RPT01B Genomic Platform at the Gonçalo Moniz Institute (IGM-FIOCRUZ) for the use of their facilities.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1221682/full#supplementary-material
Footnotes
1. ^https://www.genomedetective.com/app/typingtool/hiv
2. ^https://www.hiv.lanl.gov/content/sequence/NEWALIGN/align.html
References
Accornero, P., Radrizzani, M., Delia, D., Gerosa, F., Kurrle, R., and Colombo, M. P. (1997). Differential susceptibility to HIV-GP120-sensitized apoptosis in CD4+ T-cell clones with different T-helper phenotypes: role of CD95/CD95L interactions. Blood 89, 558–569. doi: 10.1182/blood.V89.2.558
Albuquerque, L. C. P., Mendonça, I. R., Cardoso, P. N., Baldaçara, L. R., Borges, M. R. M. M., et al. (2014). HIV/AIDS-related visceral leishmaniasis: a clinical and epidemiological description of visceral leishmaniasis in northern Brazil. Rev. Soc. Bras. Med. Trop. 47, 38–46. doi: 10.1590/0037-8682-0180-2013
Araujo, A. F., Brites, C., Monteiro-Cunha, J., Santos, L. A., Galvao-Castro, B., and Alcantara, L. C. J. (2010). Lower prevalence of human immunodeficiency virus type 1 Brazilian subtype B found in northeastern Brazil with slower progression to AIDS. AIDS Res. Hum. Retrovir. 26, 1249–1254. doi: 10.1089/aid.2010.0068
Barreto, C. C., Nishyia, A., Araújo, L. V., Ferreira, J. E., Busch, M. P., and Sabino, E. C. (2006). Trends in antiretroviral drug resistance and clade distributions among HIV-1-infected blood donors in Sao Paulo. Brazil. J. Acquir. Immune Defic. Syndr. 41, 338–341. doi: 10.1097/01.qai.0000199097.88344.50
Bogdan, C., Donhauser, N., Döring, R., Röllinghoff, M., Diefenbach, A., and Rittig, M. G. (2000). Fibroblasts as host cells in latent leishmaniosis. J. Exp. Med. 191, 2121–2130. doi: 10.1084/jem.191.12.2121
Botana, L., Ibarra-Meneses, A. V., Sánchez, C., Castro, A., San Martin, J. V., Molina, L., et al. (2019). Asymptomatic immune responders to Leishmania among HIV positive patients. PLoS Negl. Trop. Dis. 13:e0007461. doi: 10.1371/journal.pntd.0007461
Botelho, A. C. A., and Natal, D. (2009). Primeira descrição epidemiológica da leishmaniose visceral em Campo Grande, Estado de Mato Grosso do Sul. Rev. Soc. Bras. Med. Trop. 42, 503–508. doi: 10.1590/S0037-86822009000500006
Braga, L. S., Navasconi, T. R., Leatte, E. P., Skraba, C. M., Silveira, T. G. V., and Ribas-Silva, R. C. (2015). Presence of anti-Leishmania (Viannia) braziliensis antibodies in blood donors in the west-central region of the state of Paraná. Brazil. Rev. Soc. Bras. Med. Trop. 48, 622–625. doi: 10.1590/0037-8682-0043-2015
Bruhn, F. R. P., Morais, M. H. F., Bruhn, N. C. P., Cardoso, D. L., Ferreira, F., and Rocha, C. M. B. M. (2018). Human visceral leishmaniasis: factors associated with deaths in Belo Horizonte, Minas Gerais state, Brazil from 2006 to 2013. Epidemiol. Infect. 146, 565–570. doi: 10.1017/S0950268818000109
Buzón, J., Massanella, M., Llibre, J. M., Esteve, A., Dahl, V., Puertas, M. C., et al. (2010). HIV-1 replication and immune dynamics are affected by raltegravir intensification of HAART-suppressed subjects. Nat. Med. 16, 460–465. doi: 10.1038/nm.2111
Caldas, A., Favali, C., Aquino, D., Vinhas, V., van Weyenbergh, J., Brodskyn, C., et al. (2005). Balance of IL-10 and interferon-γ plasma levels in human visceral leishmaniasis: implications in the pathogenesis. BMC Infect. Dis. 5:113. doi: 10.1186/1471-2334-5-113
Carneiro, M. W., Fukutani, K. F., Andrade, B. B., Curvelo, R. P., Cristal, J. R., Carvalho, A. M., et al. (2016). Gene expression profile of high IFN-γ producers stimulated with Leishmania braziliensis identifies genes associated with cutaneous Leishmaniasis. PLoS Negl. Trop. Dis. 10:e0005116. doi: 10.1371/journal.pntd.0005116
Carranza-Tamayo, C. O., Santana, T., Assis, M.De, Teresa, A., Neri, B., Cupolillo, E., et al. (2009). Prevalence of Leishmania infection in adult HIV/AIDS patients treated in a tertiary-level care center in Brasilia, Federal District, Brazil. Trans. R. Soc. Trop. Med. Hyg. 103, 743–748. doi: 10.1016/j.trstmh.2009.01.014
Carvalho, F. L., Aires, D. L. S., Segunda, Z. F., Azevedo, C., Corrêa, R. D. G. C. F., Aquino, D. M. C., et al. (2013). Perfil epidemiológico dos indivíduos HIV positivo e coinfecção HIV-Leishmania em um serviço de referência em São Luís, MA. Brasil. Cien. Saude Colet. 18, 1305–1312. doi: 10.1590/S1413-81232013000500015
Charlab, R., Barral-Netto, M., Valenzuela, J. G., Barral, A., Caldas, A., Honda, E., et al. (2000). Human immune response to sand fly salivary gland antigens: a useful epidemiological marker? Am. J. Trop. Med. Hyg. 62, 740–745. doi: 10.4269/ajtmh.2000.62.740
Chavez, H. H., Tran, T.-A., Dembele, B., Nasreddine, N., Lambotte, O., Gubler, B., et al. (2007). Lack of evidence for prolonged double-long terminal repeat episomal HIV DNA stability in vivo. JAIDS J. Acquir. Immune Defic. Syndr. 45, 247–249. doi: 10.1097/QAI.0b013e3180415dc2
Claiborne, D. T., Prince, J. L., Scully, E., Macharia, G., Micci, L., Lawson, B., et al. (2015). Replicative fitness of transmitted HIV-1 drives acute immune activation, proviral load in memory CD4 + T cells, and disease progression. Proc. Natl. Acad. Sci. 112, E1480–E1489. doi: 10.1073/pnas.1421607112
Costa, A. S. A., Costa, G. C., Aquino, D. M. C., Mendonça, V. R. R., Barral, A., Barral-Netto, M., et al. (2012). Cytokines and visceral leishmaniasis: a comparison of plasma cytokine profiles between the clinical forms of visceral leishmaniasis. Mem. Inst. Oswaldo Cruz 107, 735–739. doi: 10.1590/S0074-02762012000600005
Cota, G. F., de Sousa, M. R., de Mendonça, A. L. P., Patrocinio, A., Assunção, L. S., de Faria, S. R., et al. (2014). Leishmania-HIV co-infection: clinical presentation and outcomes in an urban area in Brazil. PLoS Negl. Trop. Dis. 8, e2816–e2818. doi: 10.1371/journal.pntd.0002816
Coura-Vital, W., Araújo, V. E. M., Reis, I. A., Amancio, F. F., Reis, A. B., and Carneiro, M. (2014). Prognostic factors and scoring system for death from visceral Leishmaniasis: an historical cohort study in Brazil. PLoS Negl. Trop. Dis. 8, e3374–e3312. doi: 10.1371/journal.pntd.0003374
Cummins, N. W., and Badley, A. D. (2010). Mechanisms of HIV-associated lymphocyte apoptosis: 2010. Cell Death Dis. 1:e99. doi: 10.1038/cddis.2010.77
Delgobo, M., Mendes, D. A. G. B., Kozlova, E., Rocha, E. L., Rodrigues-Luiz, G. F., Mascarin, L., et al. (2019). An evolutionary recent IFN/IL-6/CEBP axis is linked to monocyte expansion and tuberculosis severity in humans. elife 8, 1–32. doi: 10.7554/eLife.47013
Diniz, L. F. B., Souza, C. D. F., and Carmo, R. F. (2018). Epidemiology of human visceral leishmaniasis in the urban centers of the lower-middle São Francisco Valley, Brazilian semiarid region. Rev. Soc. Bras. Med. Trop. 51, 461–466. doi: 10.1590/0037-8682-0074-2018
Diro, E., Edwards, T., Ritmeijer, K., Fikre, H., Abongomera, C., Kibret, A., et al. (2019). Long term outcomes and prognostics of visceral leishmaniasis in hiv infected patients with use of pentamidine as secondary prophylaxis based on CD4 level: a prospective cohort study in Ethiopia. PLoS Negl. Trop. Dis. 13, e0007132–e0007117. doi: 10.1371/journal.pntd.0007132
Druzian, A. F., Souza, A. S., Campos, D. N., Croda, J., Higa, M. G., Dorval, M. E. C., et al. (2015). Risk factors for death from visceral leishmaniasis in an urban area of Brazil. PLoS Negl. Trop. Dis. 9, e0003982–e0003911. doi: 10.1371/journal.pntd.0003982
Farre, L., Bittencourt, A. L., Silva-Santos, G., Almeida, A., Silva, A. C., Decanine, D., et al. (2008). Fas-670 promoter polymorphism is associated to susceptibility, clinical presentation, and survival in adult T cell leukemia. J. Leukoc. Biol. 83, 220–222. doi: 10.1189/jlb.0407198
Fontoura, I. G., Barbosa, D. S., de Andrade Paes, A. M., Santos, F. S., Neto, M. S., Fontoura, V. M., et al. (2018). Epidemiological, clinical and laboratory aspects of human visceral leishmaniasis (HVL) associated with human immunodeficiency virus (HIV) coinfection: a systematic review – CORRIGENDUM. Parasitology 145:1819. doi: 10.1017/S0031182018001166
Frade, A. F., Oliveira, L. C., Costa, D. L., Costa, C. H. N., Aquino, D., Van Weyenbergh, J., et al. (2011). TGFB1 and IL8 gene polymorphisms and susceptibility to visceral leishmaniasis. Infect. Genet. Evol. 11, 912–916. doi: 10.1016/j.meegid.2011.02.014
França, A., Pompilio, M. A., Pontes, E. R. J. C., de Oliveira, M. P., Pereira, L. O. R., Lima, R. B., et al. (2018). Leishmania infection in blood donors: a new challenge in leishmaniasis transmission? PLoS One 13:e0198199. doi: 10.1371/journal.pone.0198199
Frenkel, L. M., Wagner, L. E., Atwood, S. M., Cummins, T. J., and Dewhurst, S. (1995). Specific, sensitive, and rapid assay for human immunodeficiency virus type 1 pol mutations associated with resistance to zidovudine and didanosine. J. Clin. Microbiol. 33, 342–347. doi: 10.1128/jcm.33.2.342-347.1995
Fukutani, K. F., Figueiredo, V., Celes, F. S., Cristal, J. R., Barral, A., Barral-Netto, M., et al. (2014). Serological survey of Leishmaniainfection in blood donors in Salvador. Northeastern Brazil. BMC Infect. Dis. 14:422. doi: 10.1186/1471-2334-14-422
Gardinassi, L. G., Garcia, G. R., Costa, C. H. N., Costa Silva, V., and de Miranda Santos, I. K. F. (2016). Blood transcriptional profiling reveals immunological signatures of distinct states of infection of humans with Leishmania infantum. PLoS Negl. Trop. Dis. 10:e0005123. doi: 10.1371/journal.pntd.0005123
Grande, E., Zucchetto, A., Suligoi, B., Grippo, F., Pappagallo, M., Virdone, S., et al. (2017). Multiple cause-of-death data among people with AIDS in Italy: a nationwide cross-sectional study. Popul. Health Metrics 15:19. doi: 10.1186/s12963-017-0135-3
Guedes, D. L., Medeiros, Z., Dionísio da Silva, E., Martins de Vasconcelos, A. V., Santana da Silva, M., Lopes da Silva, M. A., et al. (2018). Visceral Leishmaniasis in hospitalized HIV-infected patients in Pernambuco. Brazil. Am. J. Trop. Med. Hyg. 99, 1541–1546. doi: 10.4269/ajtmh.17-0787
Guindon, S., Dufayard, J.-F., Lefort, V., Anisimova, M., Hordijk, W., and Gascuel, O. (2010). New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst. Biol. 59, 307–321. doi: 10.1093/sysbio/syq010
Hoffmann, M., Pantazis, N., Martin, G. E., Hickling, S., Hurst, J., Meyerowitz, J., et al. (2016). Exhaustion of activated CD8 T cells predicts disease progression in primary HIV-1 infection. PLoS Pathog. 12:e1005661. doi: 10.1371/journal.ppat.1005661
Horrillo, L., Castro, A., Matía, B., Molina, L., García-Martínez, J., Jaqueti, J., et al. (2019). Clinical aspects of visceral leishmaniasis caused by L. infantum in adults. Ten years of experience of the largest outbreak in Europe: what have we learned? Parasit. Vectors 12:359. doi: 10.1186/s13071-019-3628-z
Ipp, H., Zemlin, A. E., Erasmus, R. T., and Glashoff, R. H. (2014). Role of inflammation in HIV-1 disease progression and prognosis. Crit. Rev. Clin. Lab. Sci. 51, 98–111. doi: 10.3109/10408363.2013.865702
Janini, L. M., Pieniazek, D., Peralta, J. M., Schechter, M., Tanuri, A., Vicente, A. C. P., et al. (1996). Identification of single and dual infections with distinct subtypes of human immunodeficiency virus type 1 by using restriction fragment length polymorphism analysis. Virus Genes 13, 69–81. doi: 10.1007/BF00576981
Kedzierska, K., and Crowe, S. M. (2001). Cytokines and HIV-1: interactions and clinical implications. Antivir. Chem. Chemother. 12, 133–150. doi: 10.1177/095632020101200301
Kenway-Lynch, C. S., Das, A., Lackner, A. A., and Pahar, B. (2014). Cytokine/chemokine responses in activated CD4 + and CD8 + T cells isolated from peripheral blood, bone marrow, and axillary lymph nodes during acute simian immunodeficiency virus infection. J. Virol. 88, 9442–9457. doi: 10.1128/JVI.00774-14
Khouri, R., Silva-Santos, G., Dierckx, T., Menezes, S. M., Decanine, D., Theys, K., et al. (2018). A genetic IFN/STAT1/FAS axis determines CD4 T stem cell memory levels and apoptosis in healthy controls and adult T-cell leukemia patients. Onco. Targets. Ther. 7:e1426423. doi: 10.1080/2162402X.2018.1426423
Khouri, R., and Vandamme, A. M. (2015). Virus genetic variability involvement in transmissibility of HIV-1 immune activation and disease progression. Future Virol. 10, 1259–1262. doi: 10.2217/fvl.15.96
Kouri, V., Khouri, R., Alemán, Y., Abrahantes, Y., Vercauteren, J., Pineda-Peña, A.-C., et al. (2015). CRF19_cpx is an evolutionary fit HIV-1 variant strongly associated with rapid progression to AIDS in Cuba. EBioMedicine 2, 244–254. doi: 10.1016/j.ebiom.2015.01.015
Kozal, M. J., Shah, N., Shen, N., Yang, R., Fucini, R., Merigan, T. C., et al. (1996). Extensive polymorphisms observed in HIV–1 clade B protease gene using high–density oligonucleotide arrays. Nat. Med. 2, 753–759. doi: 10.1038/nm0796-753
le Fichoux, Y., Quaranta, J.-F., Aufeuvre, J.-P., Lelievre, A., Marty, P., Suffia, I., et al. (1999). Occurrence of Leishmania infantum Parasitemia in asymptomatic blood donors living in an area of Endemicity in southern France. J. Clin. Microbiol. 37, 1953–1957. doi: 10.1128/JCM.37.6.1953-1957.1999
Lima, Á. L. M., de Lima, I. D., Coutinho, J. F. V., de Sousa, Ú. P. S. T., Rodrigues, M. A. G., Wilson, M. E., et al. (2017). Changing epidemiology of visceral leishmaniasis in northeastern Brazil: a 25-year follow-up of an urban outbreak. Trans. R. Soc. Trop. Med. Hyg. 111, 440–447. doi: 10.1093/trstmh/trx080
Lima, I. D., Lima, A. L. M., Mendes-Aguiar, C., Coutinho, J. F. V., Wilson, M. E., Pearson, R. D., et al. (2018). Changing demographics of visceral leishmaniasis in Northeast Brazil: lessons for the future. PLoS Negl. Trop. Dis. 12:e0006164. doi: 10.1371/journal.pntd.0006164
Luz, J. G. G., Naves, D. B., Meira, G. A., Dias, J. V. L., and Fontes, C. J. F. (2018). Visceral leishmaniasis in a Brazilian endemic area: an overview of occurrence, HIV coinfection and lethality. Rev. Inst. Med. Trop. Sao Paulo 60, 1–9. doi: 10.1590/s1678-9946201860012
Maia-Elkhoury, A. N. S., Sierra Romero, G. A., Valadas, O. B., Sousa-Gomes, M., Lauletta Lindoso, J. A., Cupolillo, E., et al. (2019). Premature deaths by visceral leishmaniasis in Brazil investigated through a cohort study: a challenging opportunity? PLoS Negl. Trop. Dis. 13:e0007841. doi: 10.1371/journal.pntd.0007841
Mehla, R. (2013). Chemokine deregulation in HIV infection: role of interferon gamma induced Th1-chemokine signaling. J. Clin. Cell. Immunol. 4:1779. doi: 10.4172/2155-9899.S7-004
Monteiro, J. P., Alcantara, L. C. J., de Oliveira, T., Oliveira, A. M., Melo, M. A. G., Brites, C., et al. (2009). Genetic variability of human immunodeficiency virus-1 in Bahia state, northeast, Brazil: high diversity of HIV genotypes. J. Med. Virol. 81, 391–399. doi: 10.1002/jmv.21414
Monteiro, D. C. S., Sousa, A. Q., Lima, D. M., Fontes, R. M., Praciano, C. C., Frutuoso, M. S., et al. (2016). Leishmania infantum infection in blood donors, northeastern Brazil. Emerg. Infect. Dis. 22, 739–740. doi: 10.3201/eid2204.150065
Monteiro-Cunha, J. P., Araujo, A. F., Santos, E., Galvao-Castro, B., and Alcantara, L. C. J. (2011). Lack of high-level resistance mutations in HIV type 1 BF recombinant strains circulating in Northeast Brazil. AIDS Res. Hum. Retrovir. 27, 623–631. doi: 10.1089/AID.2010.0126
Orsini, M., Canela, J. R., Disch, J., Maciel, F., Greco, D., Toledo, A., et al. (2012). High frequency of asymptomatic Leishmania spp. infection among HIV-infected patients living in endemic areas for visceral leishmaniasis in Brazil. Trans. R. Soc. Trop. Med. Hyg. 106, 283–288. doi: 10.1016/j.trstmh.2012.01.008
Oryan, A., and Akbari, M. (2016). Worldwide risk factors in leishmaniasis. Asian Pac J Trop Med 9, 925–932. doi: 10.1016/j.apjtm.2016.06.021
Paduch, K., Debus, A., Rai, B., Schleicher, U., and Bogdan, C. (2019). Resolution of cutaneous Leishmaniasis and persistence of Leishmania major in the absence of arginase 1. J. Immunol. 202, 1453–1464. doi: 10.4049/jimmunol.1801249
Perkins, M. R., Bartha, I., Timmer, J. K., Liebner, J. C., Wollinsky, D., Günthard, H. F., et al. (2015). The interplay between host genetic variation, viral replication, and microbial translocation in untreated HIV-infected individuals. J. Infect. Dis. 212, 578–584. doi: 10.1093/infdis/jiv089
Peruhype-Magalhães, V., Martins-Filho, O. A., Prata, A., Silva, L. D. A., Rabello, A., Teixeira-Carvalho, A., et al. (2006). Mixed inflammatory/regulatory cytokine profile marked by simultaneous raise of interferon-γ and interleukin-10 and low frequency of tumour necrosis factor-α+ monocytes are hallmarks of active human visceral Leishmaniasis due to Leishmania chagasi infectio. Clin. Exp. Immunol. 146, 124–132. doi: 10.1111/j.1365-2249.2006.03171.x
Pieniazek, D., Peralta, J. M., Ferreira, J. A., Krebs, J. W., Owen, S. M., Sion, F. S., et al. (1991). Identification of mixed HIV-1/HIV-2 infections in Brazil by polymerase chain reaction. AIDS 5, 1293–1300. doi: 10.1097/00002030-199111000-00002
Prestes-Carneiro, L. E., Spir, P. R. N., Fontanesi, M., Pereira Garcia, K. G., Flores, E. F., et al. (2019). Unusual manifestations of visceral leishmaniasis in children: a case series and its spatial dispersion in the western region of São Paulo state. Brazil. BMC Infect. Dis. 19:70. doi: 10.1186/s12879-018-3652-1
Rezaei, Z., Sarkari, B., Dehghani, M., Layegh Gigloo, A., and Afrashteh, M. (2018). High frequency of subclinical Leishmania infection among HIV-infected patients living in the endemic areas of visceral leishmaniasis in Fars province, southern Iran. Parasitol. Res. 117, 2591–2595. doi: 10.1007/s00436-018-5949-9
Rodrigues, V., Cordeiro-da-Silva, A., Laforge, M., Silvestre, R., and Estaquier, J. (2016). Regulation of immunity during visceral Leishmania infection. Parasit. Vectors 9:118. doi: 10.1186/s13071-016-1412-x
Rotger, M., Dang, K. K., Fellay, J., Heinzen, E. L., Feng, S., Descombes, P., et al. (2010). Genome-wide mRNA expression correlates of viral control in CD4+ T-cells from HIV-1-infected individuals. PLoS Pathog. 6:e1000781. doi: 10.1371/journal.ppat.1000781
Santos, E. D. S., Araújo, A. F., Galvão-Castro, B., and Alcantara, L. C. J. (2009). Diversidade genética do vírus da imunodeficiência humana tipo 1 (HIV-1) em mulheres infectadas de uma cidade do nordeste do Brasil. Rev. Bras. Ginecol. e Obs. 31, 609–614. doi: 10.1590/S0100-72032009001200006
Santos, G. O., Jesus, N. P. S., Cerqueira-Braz, J. V., Santos, V. S., and Lemos, L. M. D. (2019). Prevalence of HIV and associated factors among visceral leishmaniasis cases in an endemic area of Northeast Brazil. Rev. Soc. Bras. Med. Trop. 52, e20180257–e20180254. doi: 10.1590/0037-8682-0257-2018
Santos, L. A., Monteiro-Cunha, J. P., Araújo, A. F., Brites, C., Galvão-Castro, B., and Alcantara, L. C. J. (2011). Detection of distinct human immunodeficiency virus type 1 circulating recombinant forms in Northeast Brazil. J. Med. Virol. 83, 2066–2072. doi: 10.1002/jmv
Schultz, A.-K., Zhang, M., Bulla, I., Leitner, T., Korber, B., Morgenstern, B., et al. (2009). jpHMM: improving the reliability of recombination prediction in HIV-1. Nucleic Acids Res. 37, W647–W651. doi: 10.1093/nar/gkp371
Secretaria de Saúde do Estado da Bahia (2018). Diretoria de Vigilância Epidemiológica. Informe Epidemiológico de Leishmaniose Visceral (LV) - Bahia [Epidemiological Report of Visceral Leishmaniasis (LV) - Bahia], Bahia: Secretaria de Saúde do Estado da Bahia. Available at: https://www.saude.ba.gov.br/wp-content/uploads/2017/11/2018-Boletim-de-Leishmaniose-Vivsceral-n.-01.pdf
Secretaria de Saúde do Estado da Bahia (2022a). Diretoria de Vigilância Epidemiológica. Boletim Epidemiológico HIV/Aids [Epidemiological Report HIV/Aids], Bahia: Secretaria de Saúde do Estado da Bahia. Available at: https://www.saude.ba.gov.br/wp-content/uploads/2017/11/boletim-HIV.Aids-2022-final-1.pdf
Secretaria de Saúde do Estado da Bahia (2022b). Diretoria de Vigilância Epidemiológica. Informe Epidemiológico da Leishmaniose Visceral no Estado da Bahia [Epidemiological Report of Visceral Leishmaniasis in the State of Bahia], Bahia: Secretaria de Saúde do Estado da Bahia. Available at: https://www.saude.ba.gov.br/wp-content/uploads/2017/11/boletimLeishVisceral_No01_2021.pdf
Seyed, N., Peters, N. C., and Rafati, S. (2018). Translating observations from Leishmanization into non-living vaccines: the potential of dendritic cell-based vaccination strategies against Leishmania. Front. Immunol. 9, 1–10. doi: 10.3389/fimmu.2018.01227
Silva de Lima, U. R., Vanolli, L., Moraes, E. C., Ithamar, J. S., and Azevedo, C. (2019). Visceral leishmaniasis in Northeast Brazil: what is the impact of HIV on this protozoan infection? PLoS One 14:e0225875. doi: 10.1371/journal.pone.0225875
Soares, V. Y. R., Lúcio Filho, C. E. P., De Carvalho, L. I. M., Silva, A. M. M. D. M., and Eulálio, K. D. (2008). Clinical and epidemiological analysis of patients with HIV/AIDS admitted to a reference hospital in the northeast region of Brazil. Rev. Inst. Med. Trop. Sao Paulo 50, 327–332. doi: 10.1590/S0036-46652008000600003
Soto, M., Corvo, L., Garde, E., Ramírez, L., Iniesta, V., Bonay, P., et al. (2015). Coadministration of the three antigenic Leishmania infantum poly (a) binding proteins as a DNA vaccine induces protection against Leishmania major infection in BALB/c mice. PLoS Negl. Trop. Dis. 9:e0003751. doi: 10.1371/journal.pntd.0003751
Sousa, J. M., Dos, S., Ramalho, W. M., and Melo, M. A. (2018). Demographic and clinical characterization of human visceral leishmaniasis in the state of Pernambuco, Brazil between 2006 and 2015. Rev. Soc. Bras. Med. Trop. 51, 622–630. doi: 10.1590/0037-8682-0047-2018
Sousa-Gomes, M. L., Maia-Elkhoury, A. N. S., and Pelissari, D. M., Sena, J. M. de, and Cechinel, M. P. (2011). Coinfecção Leishmania-HIV no Brasil: aspectos epidemiológicos, clínicos e laboratoriais. Epidemiol. Serv. Saúde 20, 519–526. doi: 10.5123/S1679-49742011000400011
Souza, G. F., Biscione, F., Greco, D. B., and Rabello, A. (2012). Slow clinical improvement after treatment initiation in Leishmania/HIV coinfected patients. Rev. Soc. Bras. Med. Trop. 45, 147–150. doi: 10.1590/S0037-86822012000200001
Souza, A. P., Soto, M., Costa, J. M. L., Boaventura, V. S., de Oliveira, C. I., Cristal, J. R., et al. (2013). Towards a more precise serological diagnosis of human Tegumentary Leishmaniasis using Leishmania recombinant proteins. PLoS One 8:e66110. doi: 10.1371/journal.pone.0066110
Subramanian, K., Dierckx, T., Khouri, R., Menezes, S. M., Kagdi, H., Taylor, G. P., et al. (2019). Decreased RORC expression and downstream signaling in HTLV-1-associated adult T-cell lymphoma/leukemia uncovers an antiproliferative IL17 link: a potential target for immunotherapy? Int. J. Cancer 144, 1664–1675. doi: 10.1002/ijc.31922
Sun, H., Kim, D., Li, X., Kiselinova, M., Ouyang, Z., Vandekerckhove, L., et al. (2015). Th1/17 polarization of CD4 T cells supports HIV-1 persistence during antiretroviral therapy. J. Virol. 89, 11284–11293. doi: 10.1128/JVI.01595-15
Távora, L. G. F., Nogueira, M. B., and Gomes, S. T. (2015). Visceral Leishmaniasis/HIV co-infection in Northeast Brazil: evaluation of outcome. Brazilian J. Infect. Dis. 19, 651–656. doi: 10.1016/j.bjid.2015.07.004
Utay, N. S., and Douek, D. C. (2016). Interferons and HIV infection: the good, the bad, and the ugly. Pathog. Immun. 1, 107–116. doi: 10.20411/pai.v1i1.125
Utay, N. S., and Hunt, P. W. (2016). Role of immune activation in progression to AIDS. Curr. Opin. HIV AIDS 11, 131–137. doi: 10.1097/COH.0000000000000242
van Griensven, J., Ritmeijer, K., Lynen, L., and Diro, E. (2014). Visceral Leishmaniasis as an AIDS defining condition: towards consistency across WHO guidelines. PLoS Negl. Trop. Dis. 8, e2916–e2918. doi: 10.1371/journal.pntd.0002916
van Griensven, J., van Henten, S., Mengesha, B., Kassa, M., Adem, E., Endris Seid, M., et al. (2019). Longitudinal evaluation of asymptomatic Leishmania infection in HIV-infected individuals in north-West Ethiopia: a pilot study. PLoS Negl. Trop. Dis. 13:e0007765. doi: 10.1371/journal.pntd.0007765
Viana, G. M. C., Silva, M. A. C. N., Garcia, J. V. S., Guimarães, H. D., Arcos Júnior, G. F., Santos, A. V. A., et al. (2017). Epidemiological profile of patients co-infected with visceral leishmaniasis and HIV/AIDS in northeast. Brazil. Rev. Soc. Bras. Med. Trop. 50, 613–620. doi: 10.1590/0037-8682-0494-2017
Vivarini, C., Calegari-Silva, T. C., Saliba, A. M., Boaventura, V. S., França-Costa, J., Khouri, R., et al. (2017). Systems approach reveals nuclear factor erythroid 2-related factor 2/protein kinase R crosstalk in human cutaneous Leishmaniasis. Front. Immunol. 8, 1–18. doi: 10.3389/fimmu.2017.01127
World Health Organization (2015). Investing to overcome the global impact of neglected tropical diseases: Third WHO report on neglected tropical diseases Available at: https://scholar.google.com/scholar?as_vis=1&q=Investing+to+overcome+the+global+impact+of+neglected+tropical+diseases:+Third+WHO+report+on+neglected+tropical+diseases
Keywords: HIV-1, Leishmania infantum, visceral leishmaniasis, immune activation, hostpathogen interaction
Citation: de Moraes L, Santos LA, Arruda LB, Silva MPP, Silva MO, Silva JAG, Ramos A, Santos MB, Torres FG, Orge C, Teixeira AMS, Vieira TS, Ramírez L, Soto M, Grassi F, Siqueira IC, Costa DL, Costa CHN, Andrade BB, Akrami K, de Oliveira CI, Boaventura VS, Barral-Netto M, Barral A, Vandamme A-M, Van Weyenbergh J and Khouri R (2023) High seroprevalence of Leishmania infantum is linked to immune activation in people with HIV: a two-stage cross-sectional study in Bahia, Brazil. Front. Microbiol. 14:1221682. doi: 10.3389/fmicb.2023.1221682
Edited by:
John S. Spencer, Colorado State University, United StatesReviewed by:
Hira Nakhasi, Center for Biologics Evaluation and Research (FDA), United StatesVicente Larraga, Spanish National Research Council (CSIC), Spain
Copyright © 2023 de Moraes, Santos, Arruda, Silva, Silva, Silva, Ramos, Santos, Torres, Orge, Teixeira, Vieira, Ramírez, Soto, Grassi, Siqueira, Costa, Costa, Andrade, Akrami, de Oliveira, Boaventura, Barral-Netto, Barral, Vandamme, Van Weyenbergh and Khouri. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ricardo Khouri, ricardo.khouri@fiocruz.br
†These authors have contributed equally to this work and share first authorship