 Sara Cuesta-Sancho1†
Sara Cuesta-Sancho1† Denisse Márquez-Ruiz1†
Denisse Márquez-Ruiz1† Francisco Illanes-Álvarez1
Francisco Illanes-Álvarez1 Irene Campaña-Gómez1
Irene Campaña-Gómez1 Andrés Martín-Aspas1María Teresa Trujillo-Soto2Alberto Romero3
Andrés Martín-Aspas1María Teresa Trujillo-Soto2Alberto Romero3 Fátima Galán2Manuel Rodríguez-Iglesias2
Fátima Galán2Manuel Rodríguez-Iglesias2 Mercedes Márquez-Coello1‡
Mercedes Márquez-Coello1‡ José-Antonio Girón-González1*‡
José-Antonio Girón-González1*‡- 1Unidad de Enfermedades Infecciosas, Servicio de Medicina Interna, Facultad de Medicina, Hospital Universitario Puerta del Mar, Instituto de Investigación e Innovación en Ciencias Biomédicas de Cádiz (INiBICA), Universidad de Cádiz, Cádiz, Spain
- 2Servicio de Microbiología, Facultad de Medicina, Hospital Universitario Puerta del Mar, Instituto de Investigación e Innovación en Ciencias Biomédicas de Cádiz (INiBICA), Universidad de Cádiz, Cádiz, Spain
- 3Unidad de Enfermedades Infecciosas, Facultad de Medicina, Hospital Universitario Puerto Real, Instituto de Investigación e Innovación en Ciencias Biomédicas de Cádiz (INiBICA), Universidad de Cádiz, Cádiz, Spain
Objective: To evaluate the serum expression of microRNAs (miRNAs) with ability to modulate the human immunodeficiency (HIV) replication or inflammatory status in people living with HIV (PLWH).
Methods: Forty healthy controls and two groups of PLWH were evaluated: (a) Group 1 (n = 30), patients with detectable viral load at inclusion, analyzed before receiving antiretroviral therapy (ART) and 12 months after initiating it; (b) Group 2 (n = 55), PLWH with prolonged undetectable viral load. Intestinal barrier disruption (I-FABP) and bacterial translocation (16S rDNA) markers, inflammatory markers such as interleukin (IL)-6 and sCD163, immune activation and expression of specific miRNAs were evaluated.
Results: Serum concentrations of I-FABP, 16S rDNA, IL-6, sCD163 and activated T lymphocytes were increased in PLWH. Serum miR-34a was overexpressed at inclusion and remained elevated after ART. The expression of the remaining miRNAs that modulate HIV infectivity (miR-7, mir-29a, miR-150, and miR-223) was similar in PLWH and controls. Related to miRNAs implicated in inflammation (miR-21, miR-155, and miR-210), significant overexpression were observed in miR-21 and miR-210 levels in untreated PLWH, but levels were restored in those patients treated for a long period.
Conclusion: A sustained overexpression of miR-34a was detected even after prolonged HIV controlled replication. miR-21 and miR-210 can be considered new markers of inflammation with high sensitivity to its modifications.
Introduction
After acute infection by the human immunodeficiency virus (HIV), a depletion of CD4+ T cells in gut-associated lymphoid tissues occurs (Brenchley et al., 2006). The administration of antiretroviral therapy (ART) only partially repairs gut mucosal injury (Jiang et al., 2009). Due to the CD4+ T cell loss and intestinal barrier damage, microbial translocation is detected in HIV-infected patients (Brenchley et al., 2006; Raffatellu et al., 2008; Sandler and Douek, 2012; Hunt et al., 2014). Inflammatory and immune activation are consequences, among others, of the intestinal microbial translocation (Sandler and Douek, 2012; Hunt et al., 2014). Immune activation increases the number of CD4+ T lymphocytes that can be target of HIV (Sandler and Douek, 2012). In addition, persistent inflammation and immune activation have been associated with certain complications in people living with HIV (PLWH), such as metabolic syndrome, type 2 diabetes mellitus or cardiovascular disease (Triant and Grinspoon, 2007).
The inflammatory and immune activation can be modulated, among others, by certain microRNAs (miRNAs) present in PLWH (Sadri Nahand et al., 2020). miRNAs are single-stranded RNAs, of approximately 22 nucleotides, transcribed from DNA genes, but not translated into proteins, whose function is to regulate the expression of other genes. Certain miRNAs can promote (miR-34a) or inhibit (miR-7, miR-29a, miR-150 and miR-223) the replication of HIV in peripheral blood mononuclear cells, promoting the underexpression or overexpression of genes that regulate its cell cycle (Huang et al., 2007; Triboulet et al., 2007; Ahluwalia et al., 2008; Chiang et al., 2012; Fabbri et al., 2012; Duskova et al., 2013; Farberov et al., 2015; Kapoor et al., 2015; Liu et al., 2015; Monteleone et al., 2015; Murray et al., 2015; Hu et al., 2017; Sheng et al., 2017; Bashti et al., 2018). In addition, it has been demonstrated that some miRNAs modulate the increased activation of innate or adaptive immunity (Fabbri et al., 2012; Witwer et al., 2012; Murray et al., 2015; Ruelas et al., 2015; Ballegaard et al., 2017; Jin et al., 2017; Sheng et al., 2017), either by promoting a change in the macrophage phenotype toward an anti-inflammatory state (miR-21; Sheng et al., 2017), decreasing the secretion of proinflammatory cytokines (miR-210; Ballegaard et al., 2017), or by increasing the proportion of regulatory T lymphocytes (miR-155; Witwer et al., 2012). It must be remembered that the immune activation increases the number of CD4+ T lymphocytes that can be targeted by HIV (Sandler and Douek, 2012).
The study of these miRNAs in HIV infection has been carried out mostly in a cross-sectional manner. We hypothesize that changes in the concentration of the miRNAs will correlate with the HIV viral load or with the concentrations of proinflammatory molecules or immune activation before and after ART.
Our objective is to perform a quantitative evaluation of the expression of miRNAs with ability to modulate the HIV replication or inflammatory status in PLWH in different clinical-therapeutic situations: (a) before ART, (b) after 12 months of treatment, when HIV replication is controlled, and (c) in a sample of HIV-infected individuals that had undergone a prolonged time of therapy, to evaluate the long-term consequences of viral suppression on the inflammatory and immune modifications as well as on miRNAs levels. Specifically, the miRNAs studied were those related to infectivity of HIV (miR-34a-activator-, miR-7, miR-29a, miR-150, miR-223-inhibitors; Huang et al., 2007; Ahluwalia et al., 2008; Chiang et al., 2012; Farberov et al., 2015; Kapoor et al., 2015; Monteleone et al., 2015; Hu et al., 2017) or involved in the activation of inflammatory and immune systems (miR-21, miR-155, and miR-210; Fabbri et al., 2012; Witwer et al., 2012; Murray et al., 2015; Ballegaard et al., 2017).
Patients and methods
Study design
A prospective, observational study of consecutive cases of HIV infection recruited from a cohort of patients followed up in HIV outpatient clinics at the Puerta del Mar and Puerto Real University Hospitals (Cádiz, Spain) was conducted.
Eighty-five PLWH were divided in two groups: (1) Group 1 included 30 untreated patients with detectable viral load. They were evaluated at the beginning of the study and started ART. They were analyzed just before starting ART and after 12 months with ART. (2) Group 2 included 55 patients with chronic HIV infection and under ART treatment [undetectable HIV loads for a median of 180 months (range, 38–357 months)]. Forty age-and gender-matched healthy controls were recruited.
The exclusion criteria were: (1) acute HIV infection; (2) presence of other opportunistic infections (including viral hepatitis, Pneumocystis jirovecii, toxoplasmosis, tuberculosis, cytomegalovirus infections, etc.) or neoplasms. Our screening procedure followed the guidelines established by the Spanish Group for AIDS Study (GeSIDA-Grupo de Estudio de la SEIMC, 2022); (3) active drug use (cocaine, heroin, amphetamines) or significant alcohol ingestion (greater than 50 g/day); (4) treatments that could have modified the determination of inflammation-related molecules or cells (pentoxifylline, anti-inflammatory or immunosuppressive drugs); (5) red blood cell or plasma transfusion in the month before inclusion; (6) non-acceptance of follow-up.
Definitions
The duration of the HIV infection was established based on the first positive anti-HIV test.
When the HIV load was less than 50 copies/mL (Abbott RealTime HIV-1, Abbott Park, IL, United States), HIV replication was considered to be controlled.
Increased serum concentration of intestinal fatty acid-binding protein (I-FABP) was indicative of gut barrier disruption (Hunt et al., 2014). The bacterial translocation was detected by serum 16S ribosomal DNA (16S rDNA) levels (Abad-Fernández et al., 2013).
Markers of inflammatory activation were serum concentrations of interleukin (IL)-6 and soluble CD163 (sCD163). The activated CD4 and CD8 T lymphocytes were those that simultaneously expressed DR and CD38 on the membrane. Exhausted lymphocytes were those that expressed CD57 and did not express CD28. Finally, those subpopulations that expressed the cell death marker PD1 were analyzed.
Study schedule
The study protocol included: (1) clinical history, nadir CD4+ T cell count and HIV load, record of previous ART and time with undetectable HIV load in the cases in which it applies; (2) CD4+ T cell count and HIV load at inclusion in the study, (3) peripheral blood sampling at inclusion for analysis of miRNAs, bacterial translocation and inflammatory and immune parameters.
Comparison of results of cell markers and molecules between healthy controls and Group 1 patients (at inclusion) was performed.
In patients with detectable HIV loads at baseline (Group 1), ART was initiated according to the Spanish Group for AIDS Study guidelines (GeSIDA-Grupo de Estudio de la SEIMC, 2022). These patients were followed for 12 months. Bacterial translocation markers, inflammatory and immune parameters and miRNAs were analyzed just before initiation of ART and after 12 months of ART. An index was calculated to quantify the percentage of increase or decrease of molecules after 12 months of ART:
100 × (Molecule concentration after 12 months of ART – Molecule concentration at baseline)/Molecule concentration at baseline.
Current ART was continued in those patients with undetectable HIV loads at inclusion (Group 2). Study parameters were determined only at inclusion.
Comparison of results of cell markers and molecules between Group 1 patients after 12 months of treatment (when undetectable HIV viral load had been obtained) and Group 2 individuals (with a longer period of undetectability) was performed.
Laboratory methods
Serum was obtained by centrifugation at 1500 g during 15 min at room temperature of blood samples in pyrogen-free heparinized tubes (Biofreeze, Costar, United States), and it was subsequently frozen down at-80°C until its use.
16S rDNA was detected after an initial extraction of DNA (QIAamp DNA Mini Kit; QIAgen, Hilden, Germany) and quantification by spectrophotometry (BioRad, Hercules, CA, United States), as previously described (Márquez-Coello et al., 2021).
Quantikine Human Immunoassays (R&D, Minneapolis, MN, United States) were used to quantify serum concentrations of I-FABP, IL-6 and sCD163, following the indications of the manufacturer.
Fresh blood samples from pyrogen-free heparinized tubes with EDTA (Biofreeze, Costar, United States) were used for flow cytometry. Activated CD4+ and CD8+ T lymphocytes (CD4+ and CD8+ DR+CD38+), senescent status (CD4+ and CD8+ CD57+CD28-) and predisposed to death (CD4+ and CD8+ PD1+) were characterized, as previously described (Márquez-Coello et al., 2021). The stained cells were acquired and analyzed in a BD FACSCanto™ II Cell cytometer, using BD FACSDiva™ Software (BD Biosciences, San Jose, CA, United States). In each case, dead cells were excluded and 300,000 cells were acquired. Blank tubes with no antibodies were used to confirm the specificity of the staining and discriminate the sample from the background.
RNA from serum was isolated using the miRNeasy Serum/Plasma Advanced Kit (QIAGEN, Hilden, Germany), following the instructions of the manufacturer. It was subsequently retrotranscripted with the miRCURY LNA RT Kit (QIAGEN, Hilden, Germany), according to the manufacturer. They were amplified by qPCR, using SYBR® Green master mix and specific LNA primers for miRNAs of interest in the CFX 96 Real-Time System (BioRad, Hercules, CA, United States) thermocycler. Primers for the spike-ins UniSp2 and UniSp5 were used as controls for the RNA extraction. Primers for the spike-in UniSp6 were used as controls for the retrotranscription. Hsa-miR-103a-3p was used as a positive control and data were normalized to this control (Ruiz-de-León et al., 2019). Expression levels of the individual miRNAs were determined by 2−ΔΔCt (Livak and Schmittgen, 2001). All amplifications were done in triplicate. Results were shown as relative units (RU).
Statistical analysis
Data were expressed as absolute numbers (percentage) or as median values [25–75 interquartile range (IQR)]. Categorical variables were compared using the chi-square test or Fisher’s exact test. The Mann–Whitney U test or ANOVA was used to compare quantitative variables from two independent groups. For comparison of three or more independent groups, Kruskal-Wallis test was used. Friedman’s or Wilcoxon’s rank tests were used to perform paired analysis of variables. The association between quantitative variables was analyzed using the Spearman correlation test. A two-tailed p value of <0.05 was considered to be significant. SPSS 22.0 statistical software package (SPSS Inc., Chicago, IL, United States) was used to perform statistical analysis.
Ethical aspects
This study was performed according to the Helsinki Declaration. The Comité Coordinador de Ética de la Investigación Biomédica de Andalucía (Spain) approved the project (PI-0128-2018-2, Consejería de Salud de la Junta de Andalucía, Spain). Each participant gave written informed consent.
Results
The demographic, immune and virological characteristics of PLWH (Groups 1 and 2) and controls are shown in Table 1.
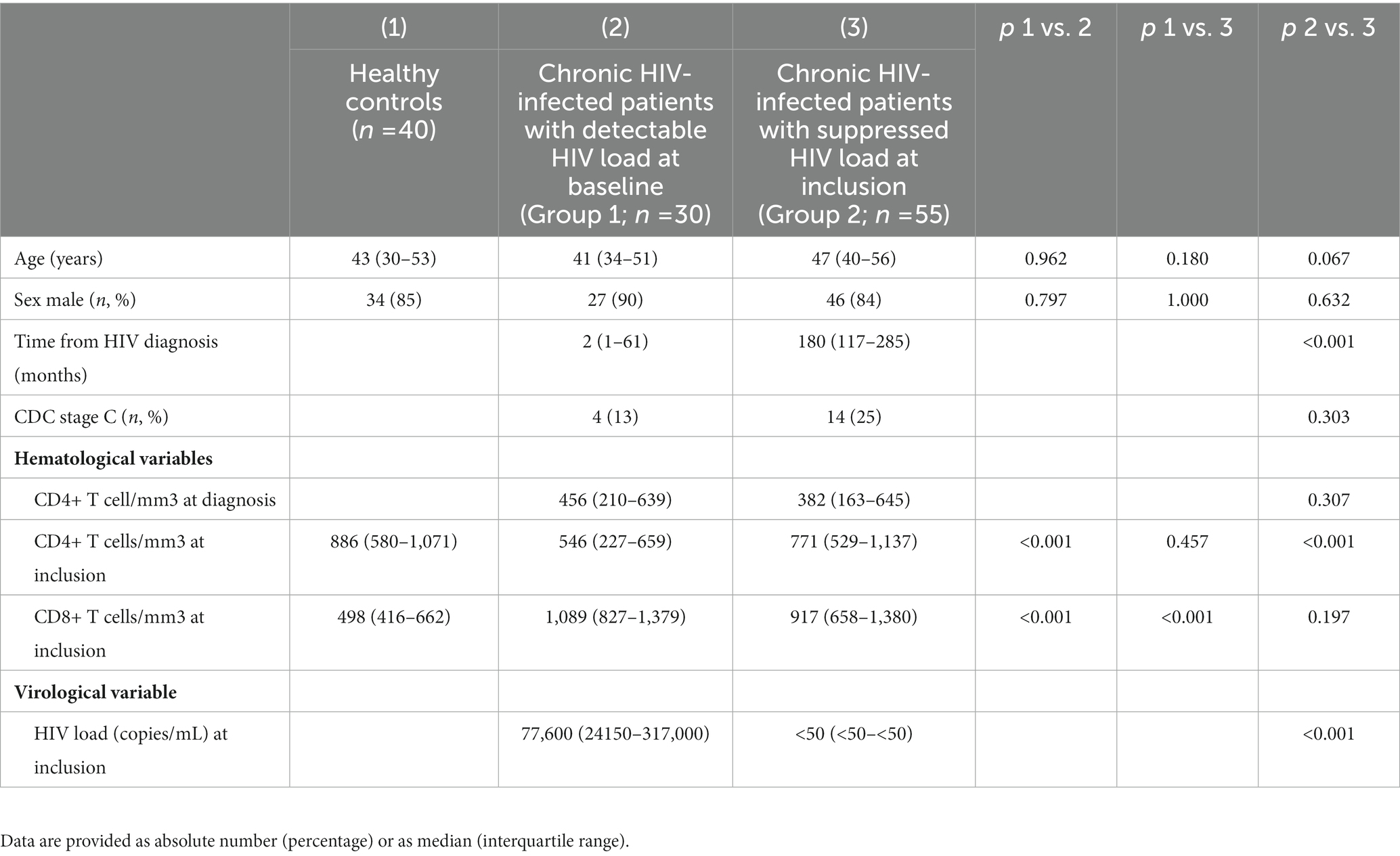
Table 1. Hematological, immune, and virological characteristics of healthy controls and HIV-infected patients.
Comparative analysis of untreated people living with HIV and healthy controls
Serum concentrations of I-FABP, 16S rDNA, IL-6, and sCD163 were significantly higher in untreated PLWH with respect to healthy controls. The activation of CD4+ and CD8+ T cells (as measured by membrane expression of CD38 and HLA-DR antigens) and the percentage of those expressing death receptor (PD1) were significantly increased in PLWH with respect to healthy controls (Table 2).
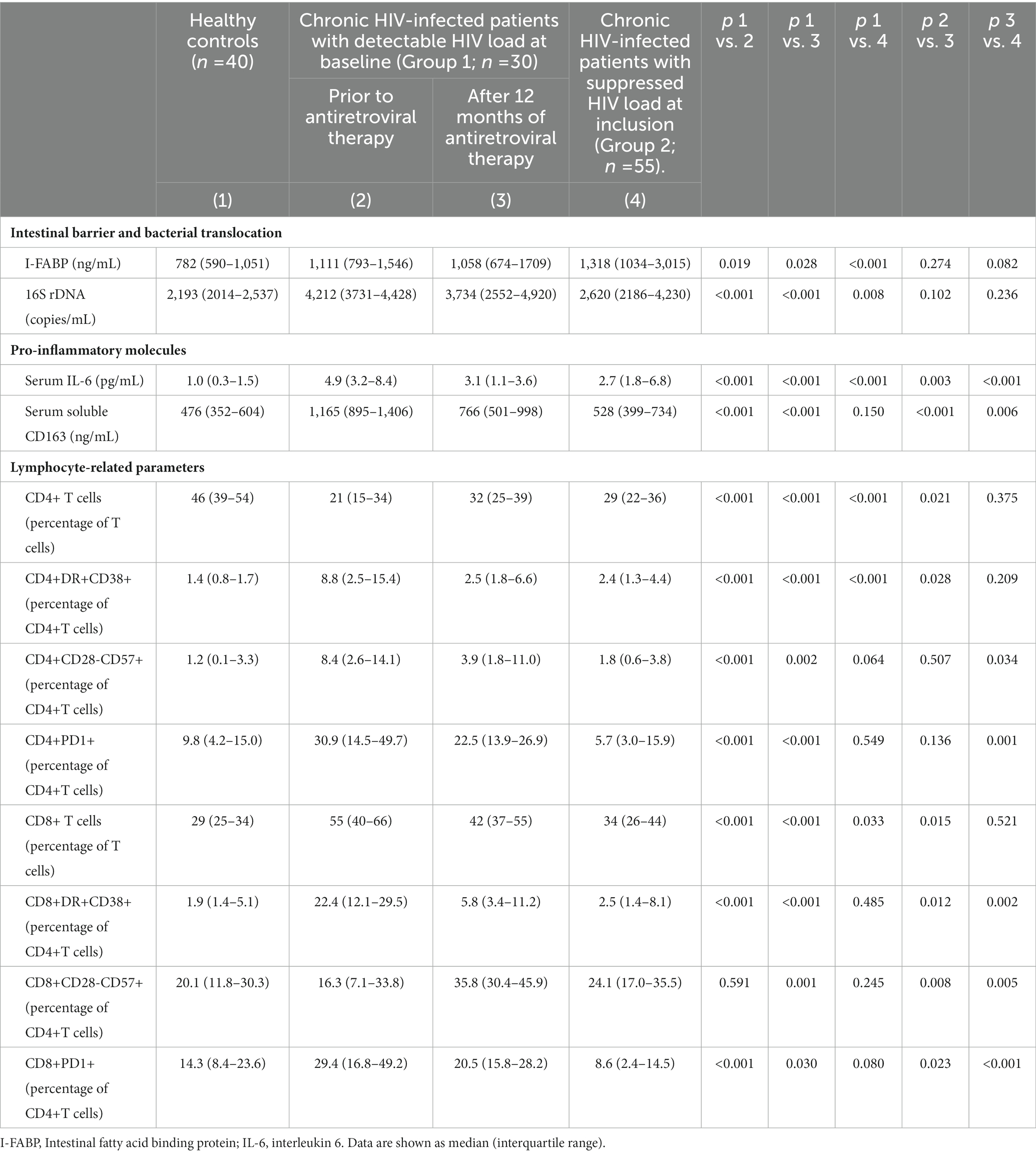
Table 2. Gut barrier status, bacterial translocation and immune characteristics of healthy controls and patients with HIV infection, untreated at baseline, analyzed at inclusion and after 12 months of antiretroviral therapy, and of chronically treated HIV patients with undetectable HIV load at inclusion.
Expression of miR-34a, a miRNA related with facilitation of HIV infection, was significantly higher in untreated PLWH compared with controls. On the contrary, levels of those related with inhibition of HIV replication (miR-7, miR-29a, miR-150, and miR-223) were similar in PLWH and controls (Table 3). There was no significant correlation between the values of these miRNAs and the viral load of the PLWH (data not shown).
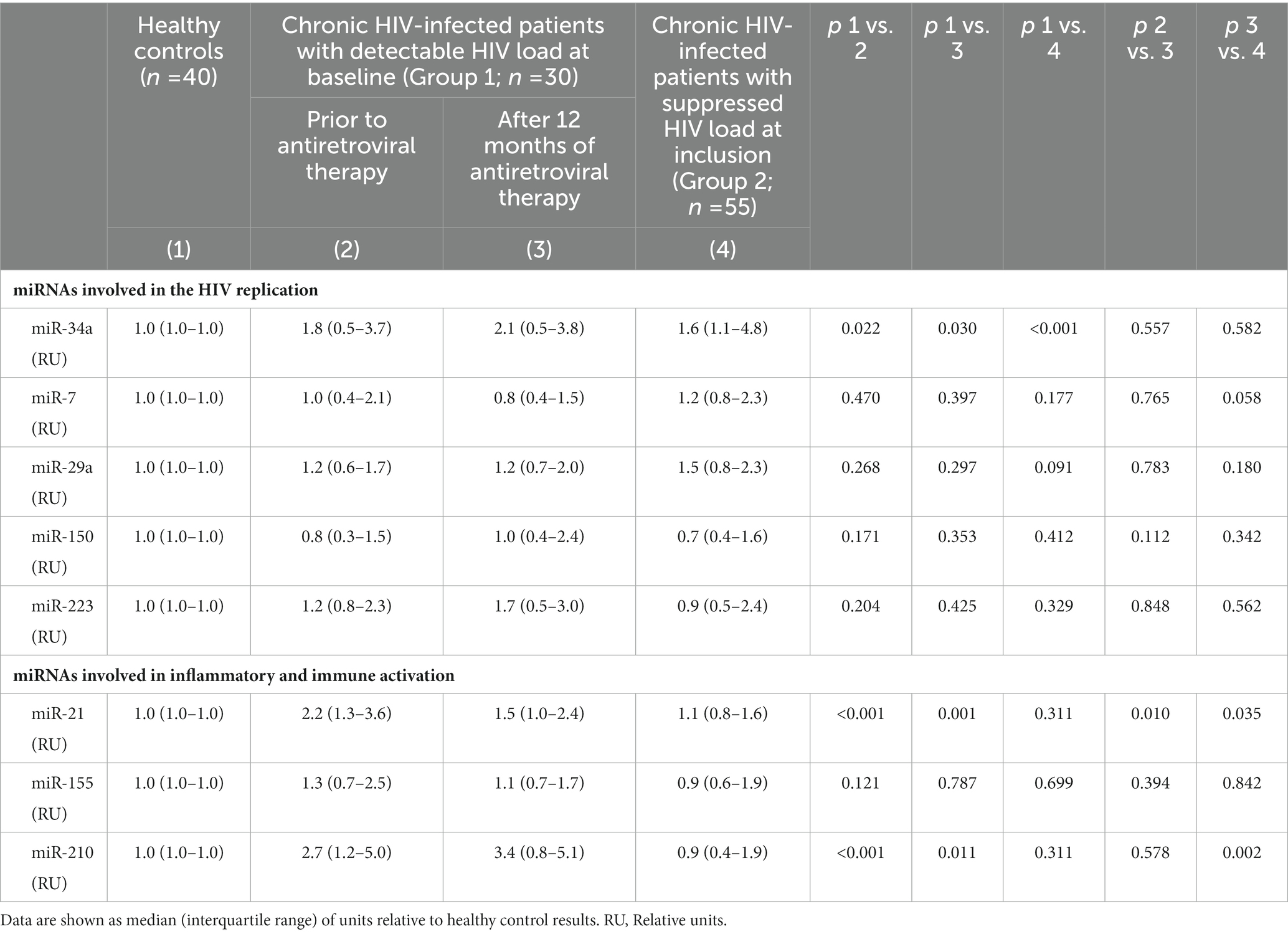
Table 3. Quantification of the miRNAs involved in the inhibition or facilitation of HIV infection, and of modulation of inflammation and immune activation, in healthy controls and patients with HIV infection, untreated at baseline, analyzed at inclusion and after 12 months of antiretroviral therapy, and of chronically treated HIV patients with undetectable HIV load at inclusion.
miR-21 and miR-210 levels, implicated in the inflammatory activation, were significantly higher in PLWH than in healthy controls (Table 3). In PLWH, no significant correlation was detected between the values of miRNAs involved in inflammatory or immune regulation and CD4 values (nadir or at the time of inclusion) or viral load. Likewise, there was no significant correlation between the values of these miRNAs and the concentrations of IL-6, sCD163 or the count of activated T lymphocytes (CD4+DR+CD38+ and CD8+DR+CD38+; Figures 1–3).
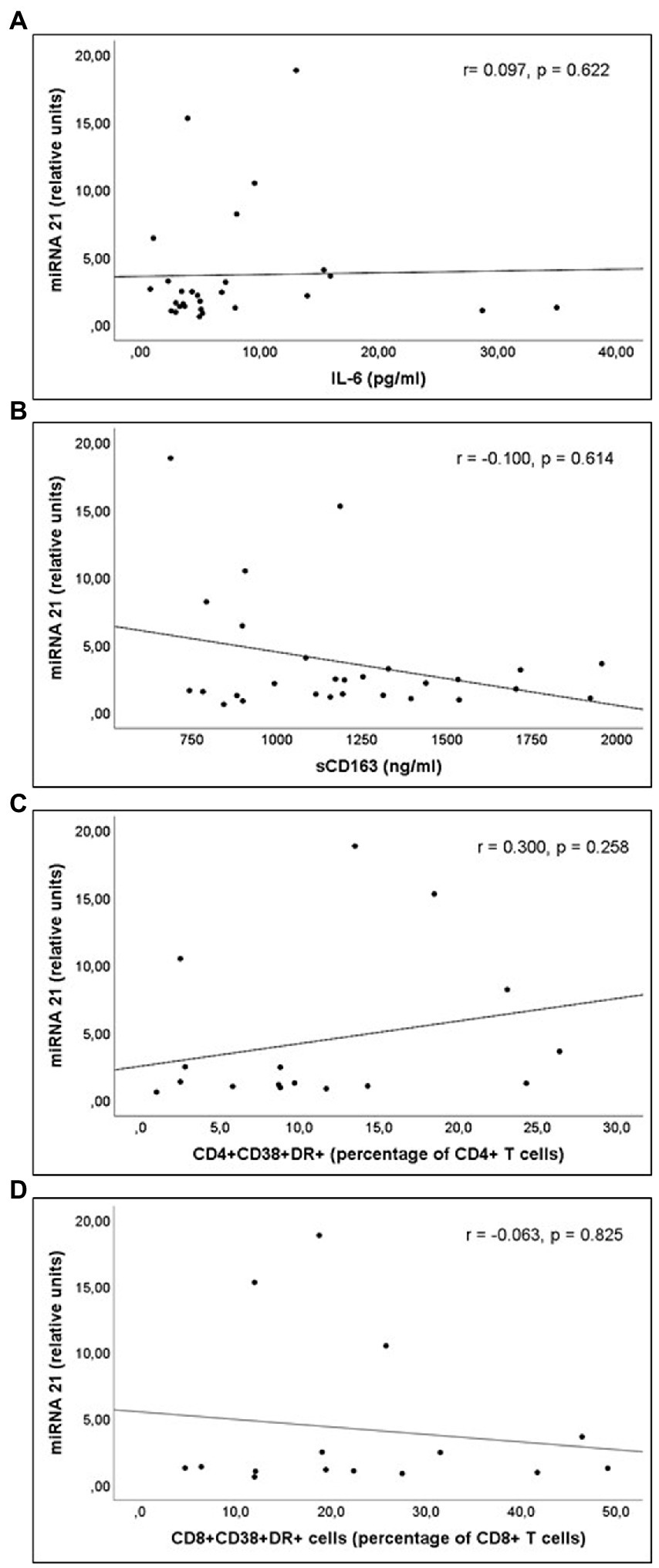
Figure 1. Correlations of miR-21 levels and (A) interleukin 6 (IL-6) levels; (B) soluble CD163 (sCD163) levels; (C) CD4+CD38+DR+ percentage of CD4+ T cell lymphocytes; and (D) CD8+CD38+DR+ percentage of CD8+ T cell lymphocytes count, in untreated people living with HIV.
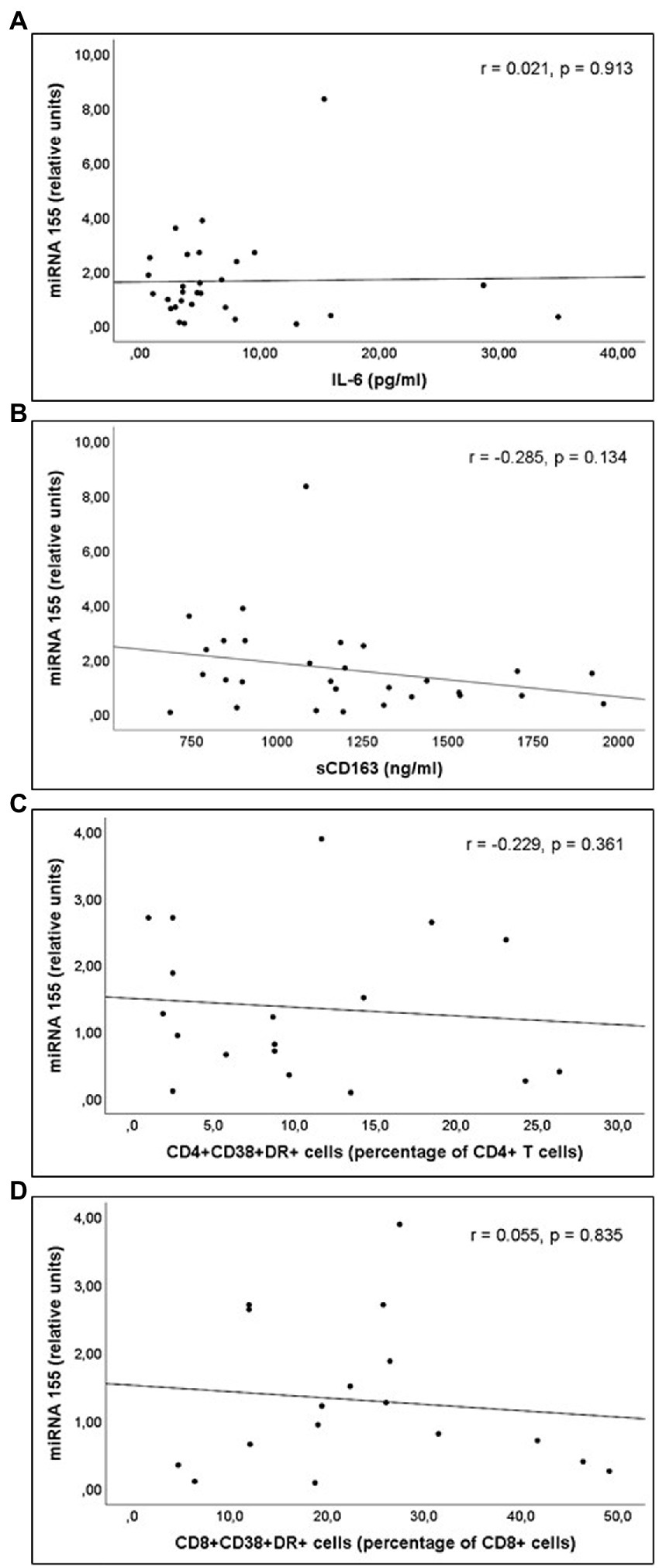
Figure 2. Correlations of miR-155 levels and (A) interleukin 6 (IL-6) levels; (B) soluble CD163 (sCD163) levels; (C) CD4+CD38+DR+ percentage of CD4+ T cell lymphocytes; and (D) CD8+CD38+DR+ percentage of CD8+ T cell lymphocytes count, in untreated people living with HIV.
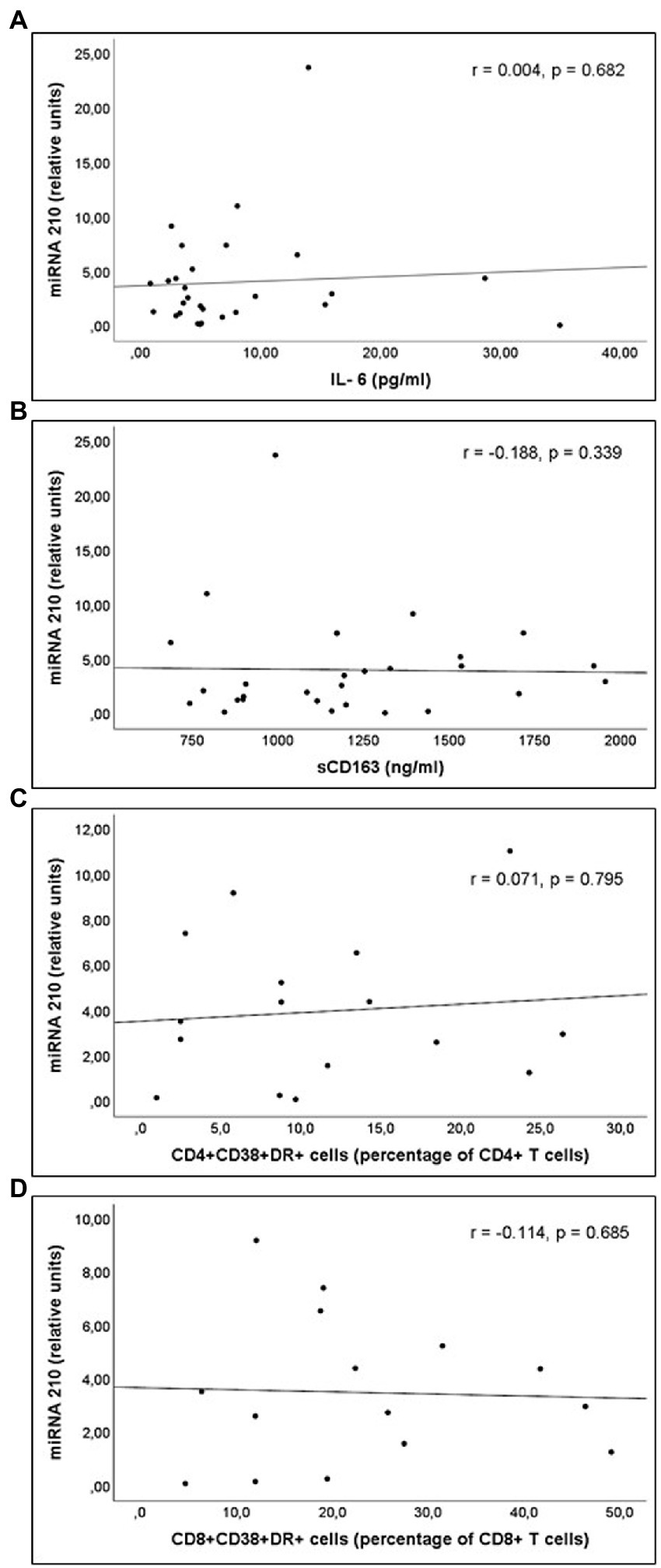
Figure 3. Correlations of miR-210 levels and (A) interleukin 6 (IL-6) levels; (B) soluble CD163 (sCD163) levels; (C) CD4+CD38+DR+ percentage of CD4+ T cell lymphocytes; and (D) CD8+CD38+DR+ percentage of CD8+ T cell lymphocytes count, in untreated people living with HIV.
Evolution of gut barrier alteration, bacterial translocation, immune markers, and modulating miRNAs in group 1 patients after antiretroviral therapy
Group 1 patients started ART and achieved undetectable HIV loads at 6 months. They were treated either with tenofovir alafenamide and emtricitabine (24 cases) or abacavir and lamivudine (6 cases) associated with integrase inhibitors (24 cases), darunavir/cobicistat (5 cases) or rilpivirine (1 case; Table 2).
During the follow-up, I-FABP and 16S rDNA concentrations remained elevated, without significant differences when compared with baseline values. IL-6 and sCD163 serum concentrations decreased significantly during the follow-up, although they remained elevated relative to healthy controls. CD4+ and CD8+ activated cells (CD38+DR+), as well as CD8+CD28-CD57+ and CD8+PD1+, significantly decreased after 12 months. The proportions of CD4+ expressing PD1 or CD28-CD57+ remained similar during the follow-up (Table 2).
The analysis of miRNAs demonstrated that only miR-21 significantly decreases during the follow-up (Table 3). A significant correlation between the decrease of miR-21 and that of CD163 was observed (Figure 4).
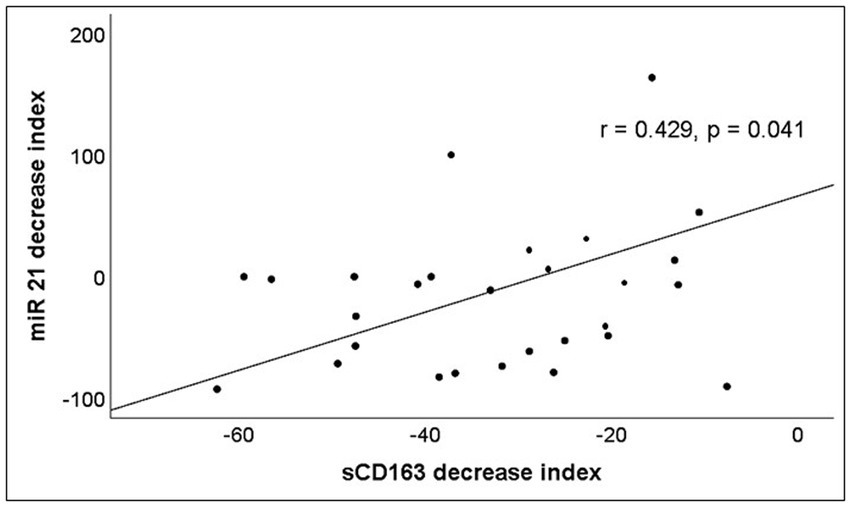
Figure 4. Correlation between the percentage of increase or decrease of serum levels of miR-21 and sCD163 after 12 months of ART. The decrease index was calculated according to the following formula: 100 x (Molecule concentration after 12 months of ART – Molecule concentration at baseline)/Molecule concentration at baseline.
Comparison of characteristics of HIV-infected patients (group 1) after antiretroviral therapy during 12 months and patients with chronically undetectable HIV load (group 2)
Characteristics observed after 12 months of ART in the 30 HIV-infected individuals that had been untreated at baseline (Group 1) were compared with those of 55 chronically infected patients with undetectable HIV at inclusion as a consequence of previous ART (Group 2) [undetectable HIV load for a median of 180 (range, 38–357) months] (Table 1). Group 2 individuals were being treated either with tenofovir (disoproxil or alafenamide) and emtricitabine (47 patients) or abacavir and lamivudine (8 patients) combined with integrase inhibitors (27 patients), darunavir/cobicistat (15 patients) or rilpivirine (13 patients).
Both I-FABP and 16S rDNA were similar in Group 2 patients when compared to Group 1 individuals treated for only 12 months. A significantly lower serum concentration of IL-6 and sCD163 was observed in Group 2 patients compared to Group 1 subjects treated with ART. Serum I-FABP, 16S rDNA and IL-6 levels in Group 2 patients were significantly higher than those of healthy controls (Table 2).
Group 2 patients showed an increase in the percentage of activated CD4+ T cells with respect to healthy controls. CD4+CD28-CD57+, CD4+PD1+, CD8+CD38+DR+, CD8+CD28-CD57+, and CD8+PD1+ proportions were similar in Group 2 patients and in healthy controls; these values were significantly lower in PLWH with undetectable HIV at inclusion (Group 2) compared with those treated for 12 months (Group 1).
There was no significant difference in the levels of miRNAs implicated in HIV replication between PLWH with undetectable HIV at inclusion (Group 2) compared with those treated for 12 months (Group 1). miR-34a levels were significantly higher in Group 2 patients compared with healthy controls. Analysis of miRNAs implicated in the regulation of inflammatory response demonstrated that significant differences were found in miR-21 and miR-210 expression: their levels were significantly higher in PLWH treated during only 12 months (Group 1) compared with those of Group 2. miR-21 and mir-210 concentrations were similar in healthy controls and Group 2 PLWH.
Analysis of the correlations between the parameters indicative of inflammatory or immune activation and the miRNAs involved in these responses was performed in PLWH with prolonged undetectable viral load (Group 2). A statistically significant correlation was observed between serum miR-21 expression and concentrations of sCD163, as well as between miR-21 expression and CD4+ T lymphocytes/mm3 values at the time of inclusion (Figure 5). Likewise, a significant correlation was observed between serum miR-21 levels and percentages of CD4+PD1+ and CD8+PD1+ cells (Figure 6).
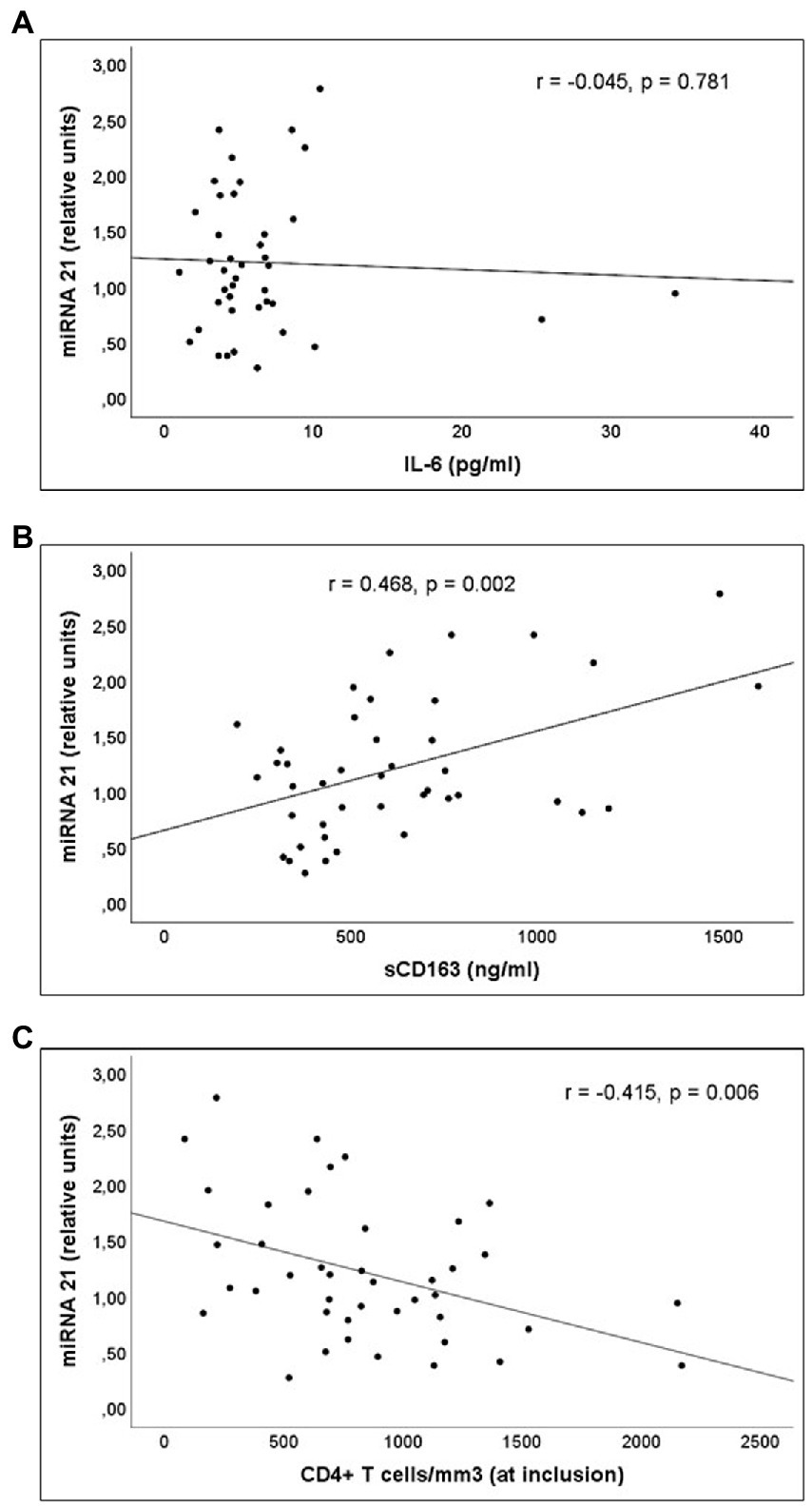
Figure 5. Correlations of miR-21 levels and serum concentration of (A) interleukin (IL)-6, (B) sCD163 and (C) CD4+ T cells/mm3 count at inclusion, in Group 2 patients (PLWH with prolonged undetectable viral load).
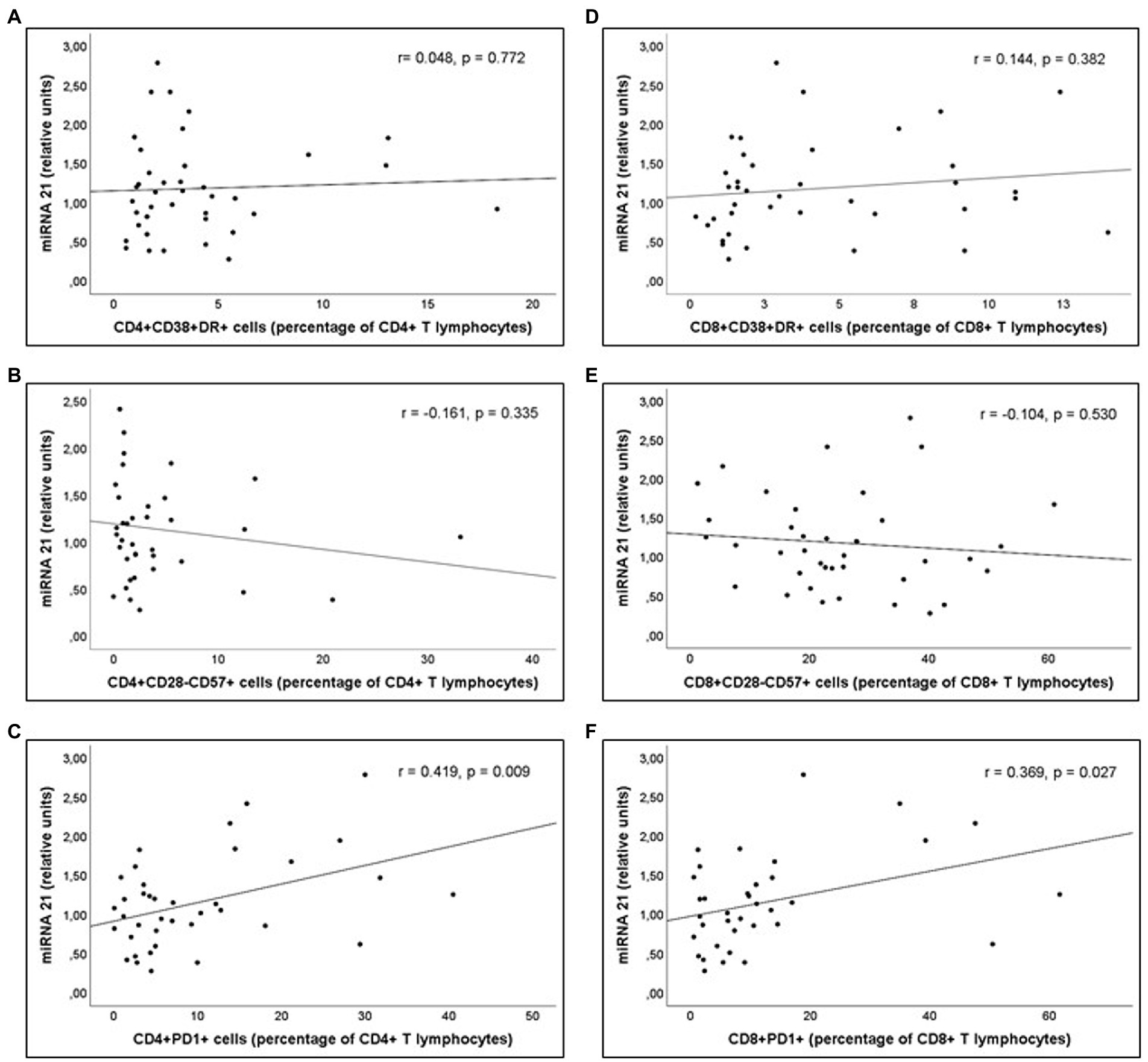
Figure 6. Correlations of miR-21 levels and (A) Percentage of CD4+DR+CD38+ (CD4+ T activated lymphocytes), (B) Percentage of CD4+CD28-CD57+ (“exhausted” CD4+ T lymphocytes), (C) Percentage of CD4+PD1+ (“predisposed to death” CD4+ T lymphocytes), (D) Percentage of CD8+DR+CD38+ (CD8+ T activated lymphocytes), (E) Percentage of CD8+CD28-CD57+ (“exhausted” CD8+ T lymphocytes), and (F) Percentage of CD8+PD1+ (“predisposed to death” CD8+ T lymphocytes), in Group 2 patients (PLWH with prolonged undetectable viral load).
In PLWH with prolonged undetectable viral load (Group 2), the influence of ART on inflammatory or immune activation parameters together with the miRNAs expression was performed. No significant differences were observed in these parameters depending on the different ART treatments [tenofovir plus emtricitabine or abacavir plus lamivudine (Supplementary Table 1)]. Similarly, no statistically significant differences were found in these values depending on whether the third drug was a non-nucleoside reverse transcriptase inhibitor, an integrase inhibitor, or a viral protease inhibitor (Supplementary Table 2).
Discussion
The present study has shown the existence of a permanent alteration of intestinal permeability, bacterial translocation and inflammatory and immune activation in PLWH. The analysis of the miRNAs reveals the existence of high values of some of those involved in facilitating HIV infection and in modulating inflammatory activation.
As it has previously been communicated (Brenchley et al., 2006; Jiang et al., 2009), intestinal barrier alteration (assessed by increased concentration of I-FABP) and microbial translocation (assessed by the elevated levels of 16S rDNA in serum), as well as inflammatory (measured by IL-6 or sCD163 levels) and T lymphocytes activation (CD4+DR+CD38+ and CD8+DR+CD38+ percentages) were detected in untreated PLWH. The proportion of T lymphocytes exhausted or expressing death receptors were also significantly elevated in these individuals. After controlling viral replication by ART, and even when bacterial translocation is not significantly modified, there was a decrease in inflammatory and immune activation, although it persisted at values higher than those detected in healthy controls.
This study has analyzed the role of miRNAs that could modify the infective capacity of HIV or modulate the inflammatory and immune activation of these individuals. The selected miRNAs have been those that have shown the ability to influence any of these phases, according to existent literature (Huang et al., 2007; Ahluwalia et al., 2008; Chiang et al., 2012; Fabbri et al., 2012; Witwer et al., 2012; Kapoor et al., 2015; Monteleone et al., 2015; Murray et al., 2015; Ballegaard et al., 2017).
miR-34a expression, but not that of miR-7, miR-29a, miR-150 or miR-223, was increased in untreated PLWH compared with controls. A positive feedback between miR-34a and HIV has been demonstrated: HIV induces the expression of miR-34a in T lymphocytes and miR-34a favors the replication of HIV in these cells, inhibiting either the phosphatase 1 nuclear-targeting subunit (Kapoor et al., 2015), the histone deacetylase sirtuin 1 (Hu et al., 2017) or the human potassium channel protein TASK-1 (Farberov et al., 2015), that regulate negatively HIV transcription. The miR-34a is significantly upregulated in mononuclear cells of PLWH with high viral load (Duskova et al., 2013). Our data support such a state of upregulation by providing evidence of increased peripheral blood miR-34a levels in PLWH. Additionally, we have been able to verify that the overexpression of this miRNA persists even after controlling viral replication by ART for very long periods. It is possible to speculate that, even when these patients had an undetectable HIV viral load in peripheral blood, the virus could be active in some other location (mainly lymph nodes or gut-associated lymphoid tissue) and the stimulus for the synthesis of miR-34a persisted. Furthermore, this suggests the existence of a favorable state for virus replication in case the ART blockade disappears. Our data open a new line toward the possibility of drug treatments directed against miR-34a in conjunction with ARTs. On the other hand, miR-34a has been implicated in endothelial cell senescence and vascular aging in PLWH (Zhan et al., 2016). The persistence of an overexpression of miR-34a, even after long periods of viral undetectability, could justify the demonstrated higher prevalence of cardiovascular disease in PLWH (Gibellini et al., 2013).
Several miRNAs (miR-7, miR-29a, miR-150, and miR-223) have enriched expression in resting primary CD4+T lymphocytes and have direct target sites on HIV-1 mRNA, suggesting that they could influence resistance against HIV (Huang et al., 2007). miR-7 may be induced in response to infections of several mammalian viruses, although the mechanisms through which it exerts its effect are not clarified (Zhang et al., 2015). miR-29 downregulates HIV-1 Nef transcripts and Nef protein expression, as well as Tat-dependent transactivation of viral gene expression (Chiang et al., 2012). miR-150 could influence HIV replication by targeting a cellular cofactor (CCNT1) via c-Myb (Huang et al., 2007). miR-223 attenuates viral gene expression by directly targeting HIV-1 mRNAs in dormant CD4+ T cells where HIV replication is silenced (Yuan et al., 2021). Upregulation of these miRNAs leads to an inhibition of HIV replication (Leo et al., 2021).
The overexpression of these miRNAs has been demonstrated in different cell types (Wang et al., 2009; Munshi et al., 2014; Narla et al., 2018; Biswas et al., 2019), although the results in this regard are controversial (Wang et al., 2009; Munshi et al., 2014; Liu et al., 2015; Bazié et al., 2022). An overexpression of miR-7 (Biswas et al., 2019), miR-29 (Narla et al., 2018), miR-150 (Munshi et al., 2014) and miR-223 (Munshi et al., 2014; Narla et al., 2018) has been observed in the PLWH plasma by some authors, while others have detected values similar to those of healthy controls (Bazié et al., 2022) or, when analyzed in cell populations, decreased (Wang et al., 2009; Munshi et al., 2014; Liu et al., 2015). It is possible that these discrepancies, as well as the differences of some of them with our results, are due to the characteristics of the patients included, specifically derived from their viral load or CD4+ T lymphocyte counts, or to the inclusion of treated and untreated patients, without differentiation between them. Our data show that expression of miR-7, −29a, −150, and −223 are similar in viremic patients and healthy controls, and that these serum levels do not change after a short or long period of therapy. These data argue against an important modulation of the viral infectivity by these miRNAs in patients with HIV infection.
miRNAs inhibit HIV-1 expression by either directly interfering with viral mRNAs or by modulating host innate immunity. IL-6 stimulates the expression of CD163 molecules (Fabriek et al., 2005) and thus sCD163 concentrations can be considered, at least in part, a consequence of IL-6-promoted proinflammatory activation. In our study, IL-6, sCD163 and activated T lymphocytes were selected as markers of inflammatory or immune activation.
miR-155 controls differentiation of CD4+ T cells into the T helper type 1 (Th1), Th2, and Th17 subsets of T helper cells and it affects the development of T regulatory (Treg) cells (Seddiki et al., 2014). miR-155 levels are increased after in vitro macrophage stimulation of Toll-like receptors 3 and 4 (Witwer et al., 2012). High levels of miR-155 have been found previously in total PBMCs, CD4 T cells and CD8 T cells in PLWH (Jin et al., 2017; Zhang et al., 2021). In our series, miR-155 level was similar in controls and in PLWH, treated or untreated. It is possible that the difference with other authors is due to the fact that in our study the levels observed were those of serum and not those of PBMCs. In any case, the absence of correlation with activation parameters and the absence of significant change in serum levels after ART is significant.
The expression of miR-210 is induced by LPS in macrophages. This miRNA works as a negative regulator for LPS-induced expression of proinflammatory cytokines – it targets NF-κB1 to decrease NF-κB activation (Qi et al., 2012). It has been demonstrated that miR-210 was upregulated in PBMCs from treated HIV-infected individuals (Ballegaard et al., 2017). In our case, miR-210 was increased in untreated or treated PLWH for 12 months compared with healthy controls. However, and in the same way as other inflammatory parameters, its concentration was similar to healthy controls in those treated for very long periods.
miR-21 is increased in response to oxidative stress or LPS in macrophages and regulates macrophage functions (Krichevsky and Gabriely, 2009; Thulasingam et al., 2011). miR-21 inhibits inflammation by targeting JAK2/STAT3 pathway (Sheng et al., 2017). Interestingly, it modulates the cytokine output of macrophages, from a proinflammatory to anti-inflammatory profile, suggesting that miR-21 is involved in macrophage polarization (Barnett et al., 2016). Besides its anti-inflammatory role, it has also been hypothesized to be involved in the permeability of the intestinal barrier, since it regulates the intestinal epithelial tight junction permeability through PTEN/PI3K/Akt signaling pathway (Zhang et al., 2015). Increased levels of miR-21 have been observed in treated HIV-infected patients (Chettimada et al., 2020). Our work has shown that miR-21 expression levels are increased in untreated subjects, as do the serum concentrations of IL-6 and sCD163. A short period of treatment (12 months) of previously untreated patients (Group 1) induces a decrease in both IL-6 and sCD163 as well as miR-21, although they remain elevated compared to healthy controls. The decreases (measured as a percentage from baseline values) of sCD163 and miR-21 were positively and significantly correlated. Patients with long-term treatment (Group 2) present values of both sCD163 and miR-21 similar to those of healthy individuals. Consequently, we can affirm that miR-21 is a new marker of inflammatory activation, with high sensitivity to the changes experienced in it.
In the same way as sCD163 and miR-21, there was a significant decrease in the percentages of CD4+PD1+ and CD8+PD1+ (T lymphocytes predisposed to death) in PLWH with a long treatment time (Group 2) compared to those treated with ART for only 12 months (Group 1). In fact, patients of Group 2 show values similar to those of the healthy controls. The relationship of miR-21 with the expression of PD1 receptors in T lymphocytes has not been previously described.
Limitations
Although each reported anti-HIV miRNA has been validated by one or more research groups, there are no miRNAs universally identified as anti-HIV miRNAs. These studies cannot be compared to each other easily because of the differences in patients’ detection methods and candidate miRNAs studied, as well as in the types of samples.
Conclusion
In conclusion, the present study has analyzed PLWH in different clinical situations (untreated, after a short period of treatment-12 months-or after a prolonged period of treatment-180 months) and has demonstrated the usefulness of three miRNAs in the follow-up of these individuals: (a) miR-34a, as a marker that possibly implies the ability to reactivate HIV after treatment withdrawal, and (b) miR-21 and miR-210, that would probably act as regulatory factors of inflammatory activation.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving human participants were reviewed and approved by Comité Coordinador de Ética de la Investigación Biomédica de Andalucía (Spain). The patients/participants provided their written informed consent to participate in this study.
Author contributions
SC-S, DM-R, MM-C, and J-AG-G conceived and designed the experiments. SC-S, DM-R, FI-Á, IC-G, AM-A, MT-S, AR, FG, and MR-I performed the experiments. SC-S, DM-R, FI-Á, IC-G, MM-C, and J-AG-G analyzed the data. SC-S, DM-R, FI-Á, IC-G, MT-S, FG, and MR-I contributed reagents, materials, and analysis tools. SC-S, DM-R, FI-Á, IC-G, MM-C, and J-AG-G wrote the draft. All authors contributed to conception of the study, and critical revision of the manuscript, and saw and approved the final version.
Funding
This work was supported by the Consejería de Salud de la Junta de Andalucía, Spain, Iniciativa territorial integrada 2014-2020 para la provincia de Cádiz [No PI-0076-2017]; by the Consejería de Salud de la Junta de Andalucía, Spain, Proyectos de investigación en Salud [No PI-0128-2018]; and by Instituto de Salud Carlos III, Acción Estratégica en Salud 2019 [No PI19/01361], Spain. Co-financed by European Regional Development Fund (FEDER).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1136718/full#supplementary-material
References
Abad-Fernández, M., Vallejo, A., Hernández-Novoa, B., Díaz, L., Gutiérrez, C., Madrid, N., et al. (2013). Correlation between different methods to measure microbial translocation and its association with immune activation in long-term suppressed HIV-1-infected individuals. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 64, 149–153. doi: 10.1097/QAI.0b013e31829a2f12
Ahluwalia, J. K., Khan, S. Z., Soni, K., Rawat, P., Gupta, A., Hariharan, M., et al. (2008). Human cellular microRNA hsa-miR-29a interferes with viral nef protein expression and HIV-1 replication. Retrovirology 5:117. doi: 10.1186/1742-4690-5-117
Ballegaard, V., Ralfkiaer, U., Pedersen, K. K., Hove, M., Koplev, S., Brændstrup, P., et al. (2017). MicroRNA-210, MicroRNA-331, and MicroRNA-7 are differentially regulated in treated HIV-1-infected individuals and are associated with markers of systemic inflammation. J. Acquir. Immune Defic. Syndr. 74, e104–e113. doi: 10.1097/QAI.0000000000001191
Barnett, R. E., Conklin, D. J., Ryan, L., Keskey, R. C., Ramjee, V., Sepulveda, E. A., et al. (2016). Anti-inflammatory effects of miR-21 in the macrophage response to peritonitis. J. Leukoc. Biol. 99, 361–371. doi: 10.1189/jlb.4A1014-489R
Bashti, O., Noruzinia, M., Garshasbi, M., and Abtahi, M. (2018). miR-31 and miR-145 as potential non-invasive regulatory biomarkers in patients with endometriosis. Cell J. 20, 84–89. doi: 10.22074/cellj.2018.4915
Bazié, W. W., Boucher, J., Goyer, B., Traoré, I. T., Kania, D., Somé, D. Y., et al. (2022). Plasma vesicular miR-155 as a biomarker of immune activation in antiretroviral treated people living with HIV. Front. Immunol. 13:916599. doi: 10.3389/fimmu.2022.916599
Biswas, S., Haleyurgirisetty, M., Lee, S., Hewlett, I., and Devadas, K. (2019). Development and validation of plasma miRNA biomarker signature panel for the detection of early HIV-1 infection. EBioMedicine 43, 307–316. doi: 10.1016/j.ebiom.2019.04.023
Brenchley, J. M., Price, D. A., Schacker, T. W., Asher, T. E., Silvestri, G., Rao, S., et al. (2006). Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 12, 1365–1371. doi: 10.1038/nm1511
Chettimada, S., Lorenz, D. R., Misra, V., Wolinsky, S. M., and Gabuzda, D. (2020). Small RNA sequencing of extracellular vesicles identifies circulating miRNAs related to inflammation and oxidative stress in HIV patients. BMC Immunol. 21:57. doi: 10.1186/s12865-020-00386-5
Chiang, K., Sung, T. L., and Rice, A. P. (2012). Regulation of cyclin T1 and HIV-1 replication by microRNAs in resting CD4+ T lymphocytes. J. Virol. 86, 3244–3252. doi: 10.1128/JVI.05065-11
Duskova, K., Nagilla, P., Le, H. S., Iyer, P., Thalamuthu, A., Martinson, J., et al. (2013). MicroRNA regulation and its effects on cellular transcriptome in human immunodeficiency virus-1 (HIV-1) infected individuals with distinct viral load and CD4 cell counts. BMC Infect. Dis. 13:250. doi: 10.1186/1471-2334-13-250
Fabbri, M., Paone, A., Calore, F., Galli, R., Gaudio, E., Santhanam, R., et al. (2012). MicroRNAs bind to toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. U. S. A. 109, E2110–E2116. doi: 10.1073/pnas.1209414109
Fabriek, B. O., Dijkstra, C. D., and van den Berg, T. K. (2005). The macrophage scavenger receptor CD163. Immunobiology 210, 153–160. doi: 10.1016/j.imbio.2005.05.010
Farberov, L., Herzig, E., Modai, S., Isakov, O., Hizi, A., and Shomron, N. (2015). MicroRNA-mediated regulation of p 21 and TASK1 cellular restriction factors enhances HIV-1 infection. J. Cell Sci. 128, 1607–1616. doi: 10.1242/jcs.167817
GeSIDA-Grupo de Estudio de la SEIMC. (2022) Documento de consenso de GeSIDA/Plan Nacional sobre el Sida respecto al tratamiento antirretroviral en adultos infectados por el virus de la inmunodeficiencia humana (Actualización enero 2022) (Accessed January 1, 2023).
Gibellini, D., Borderi, M., Clò, A., Morini, S., Miserocchi, A., Bon, I., et al. (2013). HIV-related mechanisms in atherosclerosis and cardiovascular diseases. J. Cardiovasc. Med. (Hagerstown) 14, 780–790. doi: 10.2459/JCM.0b013e3283619331
Hu, G., Liao, K., Yang, L., Pendyala, G., Kook, Y., Fox, H. S., et al. (2017). Tat-mediated induction of miRs-34a & -138 promotes astrocytic activation via downregulation of SIRT1: implications for aging in HAND. J. Neuroimmune Pharmacol. 12, 420–432. doi: 10.1007/s11481-017-9730-0
Huang, J., Wang, F., Argyris, E., Chen, K., Liang, Z., Tian, H., et al. (2007). Cellular micro RNAs contribute to HIV-1 latency in resting primary CD4+ T lymphocytes. Nat. Med. 13, 1241–1247. doi: 10.1038/nm1639
Hunt, P. W., Sinclair, E., Rodriguez, B., Shive, C., Clagett, B., Funderburg, N., et al. (2014). Gut epithelial barrier dysfunction and innate immune activation predict mortality in treated HIV infection. J. Infect. Dis. 210, 1228–1238. doi: 10.1093/infdis/jiu238
Jiang, W., Lederman, M. M., Hunt, P., Sieg, S. F., Haley, K., Rodriguez, B., et al. (2009). Plasma levels of bacterial DNA correlate with immune activation and the magnitude of immune restoration in persons with antiretroviral-treated HIV infection. J. Infect. Dis. 199, 1177–1185. doi: 10.1086/597476
Jin, C., Cheng, L., Höxtermann, S., Xie, T., Lu, X., Wu, H., et al. (2017). MicroRNA-155 is a biomarker of T-cell activation and immune dysfunction in HIV-1-infected patients. HIV Med. 18, 354–362. doi: 10.1111/hiv.12470
Kapoor, R., Arora, S., Ponia, S. S., Kumar, B., Maddika, S., and Banerjea, A. C. (2015). The miRNA miR-34a enhances HIV-1 replication by targeting PNUTS/PPP1R10, which negatively regulates HIV-1 transcriptional complex formation. Biochem. J. 470, 293–302. doi: 10.1042/BJ20150700
Krichevsky, A. M., and Gabriely, G. (2009). miR-21: a small multi-faceted RNA. J. Cell. Mol. Med. 13, 39–53. doi: 10.1111/j.1582-4934.2008.00556.x
Leo, C. G., Mincarone, P., Tumolo, M. R., Panico, A., Guido, M., Zizza, A., et al. (2021). MiRNA expression profiling in HIV pathogenesis, disease progression and response to treatment: a systematic review. Epigenomics 13, 1653–1671. doi: 10.2217/epi-2021-0237
Liu, M. Q., Zhao, M., Kong, W. H., Peng, J. S., Wang, F., Qiu, H. Y., et al. (2015). Antiretroviral therapy fails to restore levels of HIV-1 restriction miRNAs in PBMCs of HIV-1-infected MSM. Medicine 94:e2116. doi: 10.1097/MD.0000000000002116
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Márquez-Coello, M., Ruiz-Sánchez, C., Martín-Aspas, A., Fernández Gutiérrez del Álamo, C., Illanes-Álvarez, F., Cuesta-Sancho, S., et al. (2021). Neutrophil expression of T and B immunomodulatory molecules in HIV infection. Front. Immunol. 12:670966. doi: 10.3389/fimmu.2021.670966
Monteleone, K., Selvaggi, C., Cacciotti, G., Falasca, F., Mezzaroma, I., D’Ettorre, G., et al. (2015). MicroRNA-29 family expression and its relation to antiviral immune response and viro-immunological markers in HIV-1-infected patients. BMC Infect. Dis. 15:51. doi: 10.1186/s12879-015-0768-4
Munshi, S. U., Panda, H., Holla, P., Rewari, B. B., and Jameel, S. (2014). MicroRNA-150 is a potential biomarker of HIV/AIDS disease progression and therapy. PLoS One 9:e95920. doi: 10.1371/journal.pone.0095920
Murray, D. D., Suzuki, K., Law, M., Trebicka, J., Neuhaus, J., Wentworth, D., et al. (2015). Circulating microRNAs in sera correlate with soluble biomarkers of immune activation but do not predict mortality in ART treated individuals with HIV-1 infection: a case control study. PLoS One 10:e0139981. doi: 10.1371/journal.pone.0139981
Narla, V., Bhakta, N., Freedman, J. E., Tanriverdi, K., Maka, K., Deeks, S. G., et al. (2018). Unique circulating MicroRNA profiles in HIV infection. J. Acquir. Immune Defic. Syndr. 79, 644–650. doi: 10.1097/QAI.0000000000001851
Qi, J., Qiao, Y., Wang, P., Li, S., Zhao, W., and Gao, C. (2012). microRNA-210 negatively regulates LPS-induced production of proinflammatory cytokines by targeting NF-κB1 in murine macrophages. FEBS Lett. 586, 1201–1207. doi: 10.1016/j.febslet.2012.03.011
Raffatellu, M., Santos, R. L., Verhoeven, D. E., George, M. D., Wilson, R. P., Winter, S. E., et al. (2008). Simian immunodeficiency virus-induced mucosal interleukin-17 deficiency promotes salmonella dissemination from the gut. Nat. Med. 14, 421–428. doi: 10.1038/nm1743
Ruelas, D. S., Chan, J. K., Oh, E., Heidersbach, A. J., Hebbeler, A. M., Chavez, L., et al. (2015). MicroRNA-155 reinforces HIV latency. J. Biol. Chem. 290, 13736–13748. doi: 10.1074/jbc.M115.641837
Ruiz-de-León, M. J., Jiménez-Sousa, M. A., Moreno, S., García, M., Gutiérrez-Rivas, M., León, A., et al. (2019). Lower expression of plasma-derived exosome miR-21 levels in HIV-1 elite controllers with decreasing CD4 T cell count. J. Microbiol. Immunol. Infect. 52, 667–671. doi: 10.1016/j.jmii.2018.07.007
Sadri Nahand, J., Bokharaei-Salim, F., Karimzadeh, M., Moghoofei, M., Karampoor, S., Mirzaei, H. R., et al. (2020). Micro RNAs and exosomes: key players in HIV pathogenesis. HIV Med. 21, 246–278. doi: 10.1111/hiv.12822
Sandler, N. G., and Douek, D. C. (2012). Microbial translocation in HIV infection: causes, consequences and treatment opportunities. Nat. Rev. Microbiol. 10, 655–666. doi: 10.1038/nrmicro2848
Seddiki, N., Brezar, V., Ruffin, N., Lévy, Y., and Swaminathan, S. (2014). Role of miR-155 in the regulation of lymphocyte immune function and disease. Immunology 142, 32–38. doi: 10.1111/imm.12227
Sheng, B., Zhao, L., Zang, X., Zhen, J., and Chen, W. (2017). miR-375 ameliorates sepsis by downregulating miR-21 level via inhibiting JAK2-STAT3 signaling. Biomed. Pharmacother. 86, 254–261. doi: 10.1016/j.biopha.2016.11.147
Thulasingam, S., Massilamany, C., Gangaplara, A., Dai, H., Yarbaeva, S., Subramaniam, S., et al. (2011). miR-27b*, an oxidative stress-responsive microRNA modulates nuclear factor-kB pathway in RAW 264.7 cells. Mol. Cell. Biochem. 352, 181–188. doi: 10.1007/s11010-011-0752-2
Triant, V. A., and Grinspoon, S. K. (2007). Vascular dysfunction and cardiovascular complications. Curr. Opin. HIV AIDS 2, 299–304. doi: 10.1097/COH.0b013e3281e7e831
Triboulet, R., Mari, B., Lin, Y. L., Chable-Bessia, C., Bennasser, Y., Lebrigand, K., et al. (2007). Suppression of microRNA-silencing pathway by HIV-1 during virus replication. Science 315, 1579–1582. doi: 10.1126/science.1136319
Wang, X., Ye, L., Hou, W., Zhou, Y., Wang, Y. J., Metzger, D. S., et al. (2009). Cellular microRNA expression correlates with susceptibility of monocytes/macrophages to HIV-1 infection. Blood 113, 671–674. doi: 10.1182/blood-2008-09-175000
Witwer, K. W., Watson, A. K., Blankson, J. N., and Clements, J. E. (2012). Relationships of PBMC microRNA expression, plasma viral load, and CD4+ T-cell count in HIV-1-infected elite suppressors and viremic patients. Retrovirology 9:5. doi: 10.1186/1742-4690-9-5
Yuan, S., Wu, Q., Wang, Z., Che, Y., Zheng, S., Chen, Y., et al. (2021). miR-223: an immune regulator in infectious disorders. Front. Immunol. 12:781815. doi: 10.3389/fimmu.2021.781815
Zhan, J., Qin, S., Lu, L., Hu, X., Zhou, J., Sun, Y., et al. (2016). miR-34a is a common link in both HIV-and antiretroviral therapy-induced vascular aging. Aging 8, 3298–3310. doi: 10.18632/aging.101118
Zhang, X., Liu, D., Zhang, S., Wei, X., Song, J., Zhang, Y., et al. (2015). Host-virus interaction: the antiviral defense function of small interfering RNAs can be enhanced by host microRNA-7 in vitro. Sci. Rep. 5:9722. doi: 10.1038/srep09722
Zhang, L., Shen, J., Cheng, J., and Fan, X. (2015). MicroRNA-21 regulates intestinal epithelial tight junction permeability. Cell Biochem. Funct. 33, 235–240. doi: 10.1002/cbf.3109
Keywords: HIV, people living with HIV, miRNAs, IL-6, sCD163, I-FABP, bacterial translocation
Citation: Cuesta-Sancho S, Márquez-Ruiz D, Illanes-Álvarez F, Campaña-Gómez I, Martín-Aspas A, Trujillo-Soto MT, Romero A, Galán F, Rodríguez-Iglesias M, Márquez-Coello M and Girón-González J-A (2023) Expression profile of microRNAs related with viral infectivity, inflammatory response, and immune activation in people living with HIV. Front. Microbiol. 14:1136718. doi: 10.3389/fmicb.2023.1136718
Edited by:
Svetlana Khaiboullina, University of Nevada, Reno, United StatesReviewed by:
Simona Anticoli, National Institute of Health (ISS), ItalyVeron Ramsuran, University of KwaZulu-Natal, South Africa
Copyright © 2023 Cuesta-Sancho, Márquez-Ruiz, Illanes-Álvarez, Campaña-Gómez, Martín-Aspas, Trujillo-Soto, Romero, Galán, Rodríguez-Iglesias, Márquez-Coello and Girón-González. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José-Antonio Girón-González, joseantonio.giron@uca.es
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work and share last authorship