 Liangliang Xu1,3
Liangliang Xu1,3 Peng Xiang1,3
Peng Xiang1,3 Baowen Zhang1,3
Baowen Zhang1,3 Kun Yang4Fenglin Liu1,3Zesong Wang1,3Yanjun Jin1,3Longjun Deng2Weixiong Gan2
Kun Yang4Fenglin Liu1,3Zesong Wang1,3Yanjun Jin1,3Longjun Deng2Weixiong Gan2 Zhaobin Song1,3*
Zhaobin Song1,3*- 1Key Laboratory of Bio-Resources and Eco-Environment of Ministry of Education, College of Life Sciences, Sichuan University, Chengdu, China
- 2Yalong River Hydropower Development Company, Ltd., Chengdu, China
- 3Observation and Research Station of Sichuan Province of Fish Resources and Environment in Upper Reaches of the Yangtze River, College of Life Sciences, Sichuan University, Chengdu, China
- 4Institute of Ecology, China West Normal University, Nanchong, China
The fish gut microbiome plays an important role in nutrition absorption and energy metabolism. Studying the gut microbes of cold-water fish is important to understand the dietary adaptation strategies in extreme environments. In this study, the gut samples of Schizothorax wangchiachii (SW, herbivorous), Schizothorax kozlovi (SK, omnivorous), and Percocypris pingi (PP, carnivorous) in the upper Yangtze River were collected, and we sequenced 16S rRNA amplicon to study the potential relationship between gut microbes and host species. The results showed that gut microbial composition and diversity were significantly different between the three cold-water fishes. These fishes had different key taxa in their gut microbes, including bacteria involved in the breakdown of food (e.g., Cetobacterium, Aeromonas, and Clostridium sensu stricto 10). The highest alpha diversity indices (e.g., Chao 1 index) were identified in the herbivore (SW), followed by the carnivore (PP), and the lowest in the omnivore (SK). Non-metric multidimensional scaling (NMDS) results revealed that the gut microbial community of these species was different between host species. The neutral community model (NCM) showed that the microbial community structure of SW was shaped by stochastic processes, and the highest species dispersal was found in SW, followed by PP, and the lowest in SK. The results of niche breadth agreed with these findings. Our results demonstrated that host species influenced the gut microbiome composition, diversity, and microbial community assembly processes of the three cold-water fishes. These findings implied that the variation of gut microbiome composition and function plays a key role in digesting and absorbing nutrients from different foods in cold-water fish.
Introduction
Fish gut microbiota is influenced by trophic level (Liu et al., 2016), season (Neuman et al., 2016; Dulski et al., 2020), host habitat (Dehler et al., 2017; Dulski et al., 2020), and intestinal section (Parata et al., 2020). For example, herbivorous, omnivorous, and carnivorous are the main trophic levels of fish and exhibit differences in gut microbiome composition (Liu et al., 2016; Egerton et al., 2018). Cellulose-degrading bacteria (e.g., Aeromonas, Cetobacterium, Leuconostoc, and Bacillus) are dominant in herbivorous fishes (Megalobrama amblycephala and Ctenopharyngodon idellus), while protease-producing bacteria (e.g., Cetobacterium and Halomonas) are dominant in carnivorous fishes (Siniperca chuatsi and Culter alburnus) (Liu et al., 2016). Similarly, this result was found in other fishes (Egerton et al., 2018). The digestive tract of fish is divided into the foregut, midgut, and hindgut. Differences in the composition and diversity of the gut microbiome have been identified between the intestinal sections of few fish species (Tao et al., 2013; Parata et al., 2020; Cheutin et al., 2021). Tao et al. (2013) found that different intestinal sections of Miichthys miiuy showed differences in the number of culturable bacterial colonies; the highest number was found in the midgut (27.4%), followed by the foregut (25.2%), and hindgut (22.9%). Furthermore, alpha diversity (Shannon index; 1.5–4.3) of gut microbes in juvenile Acanthurus triostegus was higher in the midgut than in the hindgut, while the opposite trend was found in adults (Parata et al., 2020).
Niche-based and neutral theories are two important and complementary mechanisms for understanding the microbial community structure (Sloan et al., 2006; Bahram et al., 2016). Niche-based theories assume that deterministic processes (i.e., deterministic abiotic factors: pH and temperature; biotic factors: competition and predation) shape the microbial community structure (Gipsi et al., 2015; Wei et al., 2016). On the contrary, neutral theories consider that stochastic processes (e.g., birth, death, immigration, and limited dispersal) shape the microbial community structure (Rosindell et al., 2011; Zhou and Ning, 2017). Numerous studies have reported that stochastic processes play an important role in shaping the microbial community structure of the environment (Chen et al., 2019), and in animals (Zhao et al., 2022). However, it is challenging to quantify the relative importance of stochastic processes due to the diversity and complexity of microorganisms. To untangle the relative importance of stochastic processes in microbial community assembly, the neutral community model (NCM) was proposed by Sloan et al. (2006), and it has been widely used for quantifying the importance of stochastic processes (Sloan et al., 2006; Zhao, 2014; Chen et al., 2019).
Schizothorax wangchiachii, S. kozlovi, and P. pingi belong to Cyprinidae and are important commercial cold-water fish distributed in the upper reaches of the Yangtze River and its tributaries (Yue, 2000). Previous studies have shown that the wild population of the three cold-water fishes has decreased due to anthropogenic activities including overexploitation, hydropower development, and water pollution (Jiang et al., 2007; Feng et al., 2011). Thus, multiple studies of population improvement measures such as artificial propagation (He et al., 2014; Zhao, 2014) and stock enhancement (Deng et al., 2016) have been undertaken to protect and improve the resources of these fishes in the wild environment. However, the fry of the three fish species has been mostly fed with mixed feed during artificial breeding (Liu et al., 2015), and this is different for wild populations. For example, S. wangchiachii is an herbivorous fish and feeds on periphyton (e.g., diatoms) (Huang, 2018), S. kozlovi is an omnivorous fish and mostly feeds on algae (e.g., Spirogyra and Cymbella) and aquatic insects (e.g., Diptera and Libellulidae) (Zhang and Dai, 2011), and P. pingi is a carnivorous fish and feeds on other fishes (e.g., S. wangchiachii) (Chen et al., 2015). Consequently, differences in diet composition may influence the survival and growth of the hatchery-reared individuals of these species after stocking for enhancement. Similarly, previous studies showed that the fish gut microbiome played a major role in adapting to dietary changes (Liu et al., 2016). Therefore, the gut microbiome studies of the three cold-water fishes in the wild are helpful to explore the relationship between gut microbes and diet and would provide essential data for stock enhancement programs of the species.
To investigate the potential relationship between the gut microbiome and host species or intestinal sections, we examined the gut samples of three cold-water fishes (S. wangchiachii, S. kozlovi, and P. pingi) from the lower reaches of the Yalong River, a tributary of the upper Yangtze River (Figure 1 and Supplementary Table 1). High-throughput sequencing of the bacterial 16s rRNA gene was used to obtain microbial data. This study was performed to answer the following questions: (1) What is the influence of host species on the gut microbiome of the three cold-water fishes? (2) What is the survival strategy of adapting to different dietary compositions in the three cold-water fishes?
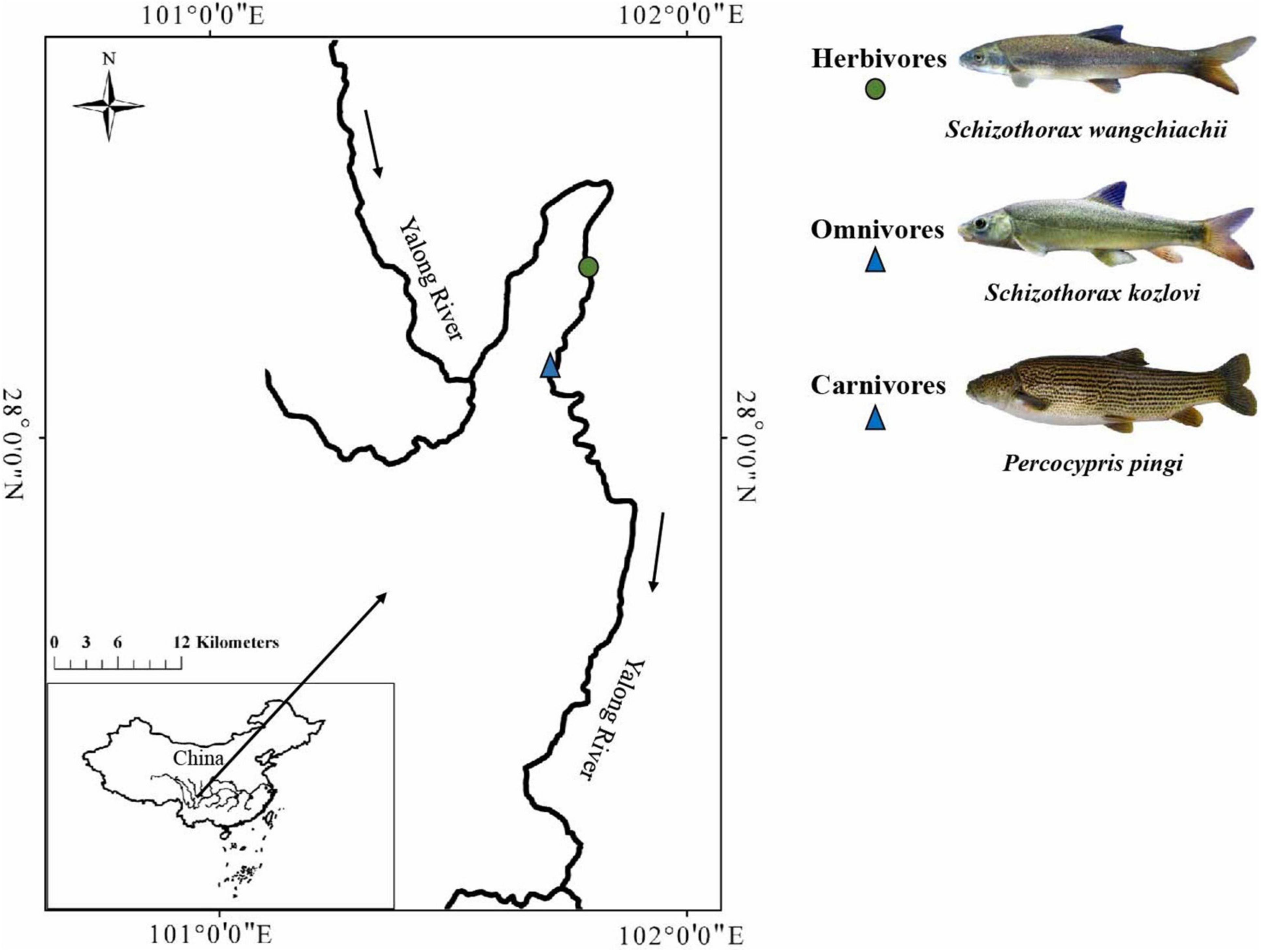
Figure 1. Sampling sites of the three cold-water fishes investigated. The solid circle represents the sampling sites for herbivores, and the solid triangle represents the sampling sites for omnivores and carnivores.
Materials and Methods
Sample Collection
Gut samples were collected from individuals of S. wangchiachii (SW; foregut: 5, midgut: 5, hindgut: 5), S. kozlovi (SK; foregut: 5, midgut: 5, hindgut: 5), and P. pingi (PP; foregut: 5, midgut: 5, hindgut: 5) in the lower reaches of the Yalong River, Sichuan Province, China (Figure 1 and Supplementary Table 1). The average water temperature and altitude of the sampling sites (E: 101°86′15, N: 28°27′41 ∼ E: 101°59′91, N: 28°17′23) in the lower reaches of the Yalong River were 11.2°C and 1,500 m, respectively. In the sampling water area, drift nets were used to capture fish in April 2021. Each fish was euthanized with MS-222 (0.6–1.0 g/L) after being caught, and its foregut, midgut, and hindgut were cut using sterile scissors. These fishes had a digestive tract that consisted of a foregut: enlarged and with thicker walls; midgut: curved and folded; and hindgut: intestinal diameter gradually narrowed. Based on the digestive tract structure of the three fish species, the intestinal contents from the foregut, midgut, and hindgut were collected. To avoid cross-contamination during sampling, the gut contents were collected from each central part of the foregut, midgut, and hindgut. All gut contents were immediately transferred to 2-ml aseptic centrifuge tubes in the field and stored at –20°C in a portable refrigerator. Finally, all samples of gut contents were stored at –80°C in the laboratory for DNA extraction.
DNA Extraction and 16S rRNA Sequencing
Gut contents were thawed in ice and then were used to extract DNA by using the QIAamp DNA Stool Mini Kit (Qiagen, Valencia, CA) according to the manufacturer’s instructions. The highly variable V4-V5 region of the 16S rRNA gene was selected and amplified with bacterial-specific universal primers 515F (5′-GTGCCAGCMGCCGCGG-3′) and 907R (5′-CCGTCAATTCMTTTRAGT-3′) (Caporaso et al., 2012). PCR was performed with conditions as follows: initial denaturation at 95°C for 5 min, 35 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 45 s; and extension at 72°C for 10 min. The products of PCR amplification were sent to Mingke Biotechnology Co., Ltd. (Hangzhou, China) for high-throughput sequencing on the Illumina HiSeq Platform (Hiseq2500 PE250).
Sequencing Data Analysis
The QIIME 1.9 software package (Caporaso et al., 2010) was used to process the raw paired-end sequences (2,273,925 reads from 45 samples). The clean sequences (2,134,418) were generated using the trimming analysis and by removing low-quality reads (the base with a mass value below 20 at the tail of read), splicing (the minimum length overlap is 10 bp and the maximum mismatch ratio is 0.2), and quality control (remove chimeras) (Edgar, 2010). After we removed the low-quality reads, the length (bp) distribution of the 45 samples was between 301 and 400 bp (99.96%). Finally, the operational taxonomic units (OTUs) were clustered, with > 97% sequence identity, and each OTU was classified by the annotation against the Silva 132 database (Release132)1 (Christian et al., 2013). Sequencing data have been uploaded to the NCBI (accession number PRJNA789150).
Bioinformatics Analysis
Gut microbial composition (i.e., phylum and genus levels) among different species were compared by using the Mann–Whitney U-test and the Kruskal–Wallis H-test in Stamp 2.1.3 (Parks et al., 2014). The bar plot was generated by R 2.0 (Kolde, 2015). The linear discriminant analysis (LDA) effect size (LEfSe) method was used to test the differences in the composition of gut microbes of the three fish species (Segata et al., 2011). During the co-occurrence analysis, the relative abundance of bacteria genera was input into Cytoscape 3.4 (Shannon et al., 2003). The plugin CoNet (Faust and Raes, 2016) was used to generate the network plots using these parameters (Spearman index, ρ = 0.5), and the top 60 genera are shown in the plot. The Venn diagram was generated by R software to test the differences in the number of shared and exclusive OTUs among the three fish species.
Alpha diversity was calculated by the Chao 1 index, the Shannon index, a phylogenetic index, and observed OTU numbers. The Mann–Whitney U-test and the one−way analysis of variance (ANOVA) were used to analyze the significant differences between intestinal sections (foregut, midgut, and hindgut) of the three cold-water fishes in Stamp 2.1.3 (Parks et al., 2014). The box plot was used to visualize the results. To analyze the differences in the microbial communities between gut samples, we conducted a PERMANOVA (number of permutations: 999) based on three distances (Bray–Curtis distance, unweighted UniFrac distance, and weighted UniFrac distance). The adonis function in the vegan package (Dixon, 2003) was used to perform a PERMANOVA on the three distances to obtain the R2-value (effect size), and the percentage of variation between species. Non-metric multidimensional scaling (NMDS) was used to visualize the beta-diversity results (Anderson, 2010).
The bacterial function of the three cold-water fishes was predicted by PICRUSt (Phylogenetic Investigation of Communities by Reconstruction of Unobserved States) (Langille et al., 2013). The functional profiles of microbial communities were generated by blasting the 16s RNA gene to the Kyoto Encyclopedia of Genes and Genomes (KEGG) database. In the PICRUSt analysis, the significant KEGG pathways (level 2) among species were analyzed via the one−way analysis of variance (ANOVA) in Stamp 2.1.3 (Parks et al., 2014). Furthermore, Bonferroni was used to correct the p-value (Benjamini and Yekutieli, 2001). The heat map was generated by the heatplus package in R 2.0 (Kolde, 2015) and used to visualize the functional metabolic profiles.
To determine the potential importance of neutral processes in community assembly, the NCM was used to predict the relationship between OTU detection frequency and their relative abundance across the wider metacommunity (Sloan et al., 2006). In the NCM, Nm is an estimate of dispersal between communities. The parameter Nm determines the correlation between occurrence frequency and regional relative abundance, N describes the metacommunity size, and m is the immigration rate. The parameter R2 indicates the overall fit of the neutral model (Sloan et al., 2006). All statistics were analyzed within 95% confidence intervals (CIs) and calculated by bootstrapping with 1,000 replicates. All computations were performed in R 2.0 software.2
The niche width approach (Levins, 1968) was used to quantify habitat specialization across the three cold-water fishes in R 2.0 software (see text footnote 2). Greater values of niche breadth indicated that the taxa are widely present and evenly distributed on the large scale, whereas lower values indicated that the taxa occupied fewer habitats and had a disordered distribution. The Kruskal–Wallis H-test and the one−way analysis of variance (ANOVA) were used to calculate the significant differences between the niche breadth of the three fish species in Stamp 2.1.3 (Parks et al., 2014).
Results
Overview of Gut Samples Data
After data quality processing, we obtained 1,928,859 qualified sequences from 45 samples, an average of 47,232 ± 9,610 sequences per sample. Rarefaction curves showed that the overall quality of sequencing was excellent, and further analysis could be undertaken (Supplementary Figure 1). The gut microbiota of the three cold-water fishes (SW, SK, and PP) were composed of 3,634 OTUs based on 97% sequence similarity. The OTUs were annotated against the Silva 132 database, and 40 phyla, 414 families, and 974 genera were identified (Supplementary Tables 2–4).
Composition of the Bacterial Community of the Three Cold-Water Fishes
Significant differences in the gut microbial composition between the three fish species (SW, SK, and PP) were identified (Figures 2, 3 and Table 1). Overall, at the phylum level, the dominant phyla were Proteobacteria (SW: 41.50%; SK: 33.40%; PP: 49.14%) and Fusobacteria (SW: 8.66%; SK: 41.16%; PP: 22.53%) (Figure 2, Supplementary Figure 2, and Supplementary Table 2). We identified significant differences in the gut microbe composition of the three fish species between intestinal sections (foregut, midgut, and hindgut) (Mann–Whitney U-test; both, p < 0.05) (Figure 2B and Table 1). The relative abundance of Cyanobacteria and Planctomycetes showed significant differences and downward trends between the herbivore (SW), omnivore (SK), and carnivore (PP) (Mann–Whitney U-test; both, p < 0.05) (Cyanobacteria: SW: 24.39%, SK: 1.61%, PP: 0.17%; Planctomycetes: SW: 13.36%, SK: 1.12%, PP: 0.29%) (Figure 2, Table 1, Supplementary Figure 3, and Supplementary Table 2). In SW vs. SK and SW vs. PP, significant differences in gut microbiome composition were found, while no significant differences were observed in SK vs. PP (Table 1).
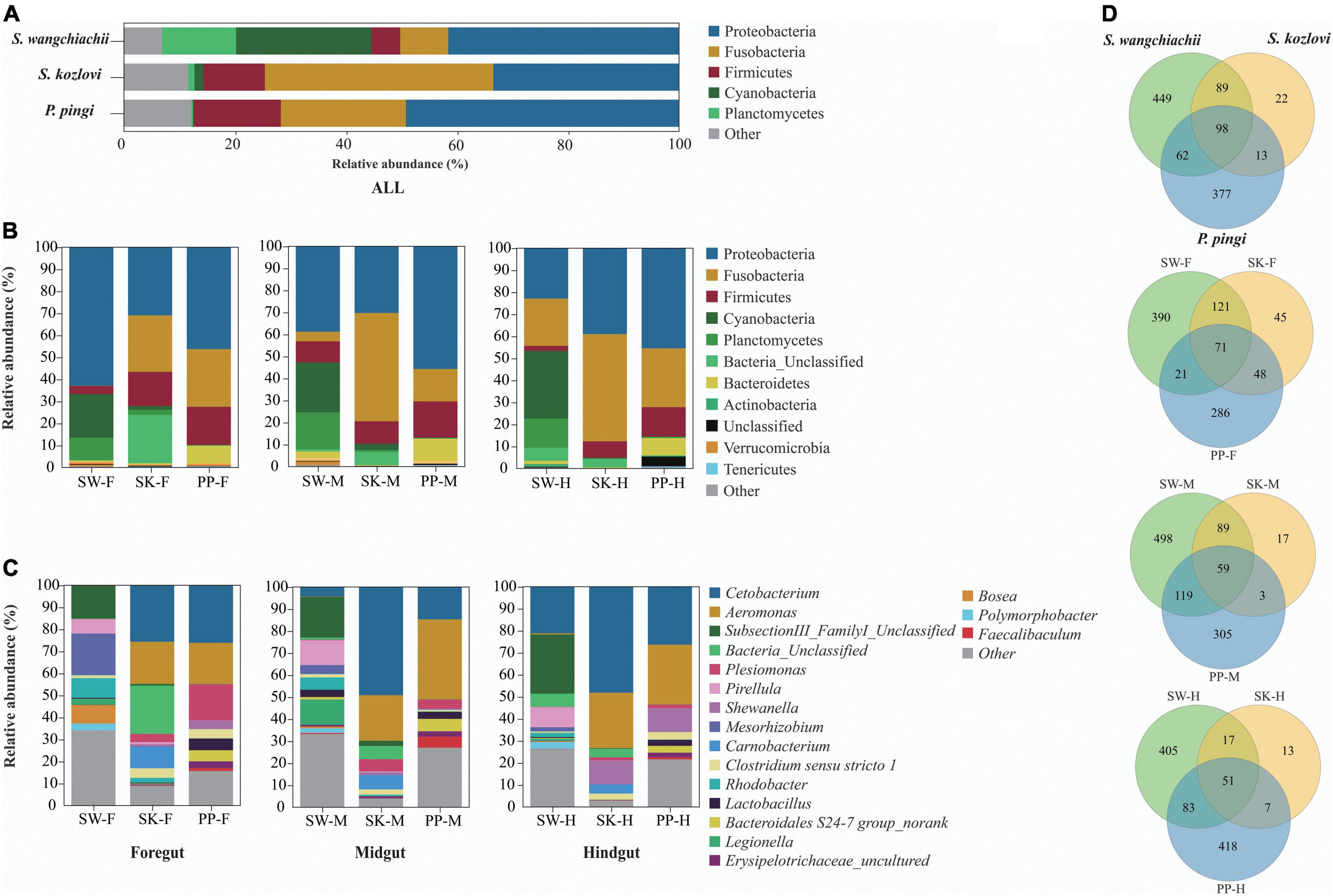
Figure 2. Gut microbiome compositions of the three cold-water fishes. (A–C) The bar plot represents the gut microbiome composition of S. wangchiachii, S. kozlovi, and P. pingi among different intestinal sections at phylum and genus levels. (D) Venn diagrams based on the OTU level to analyze the differences of gut microbes of SW, SK, and PP. SW, S. wangchiachii; SK, S. kozlovi; PP, P. pingi; F, foregut; M, midgut; H, hindgut.
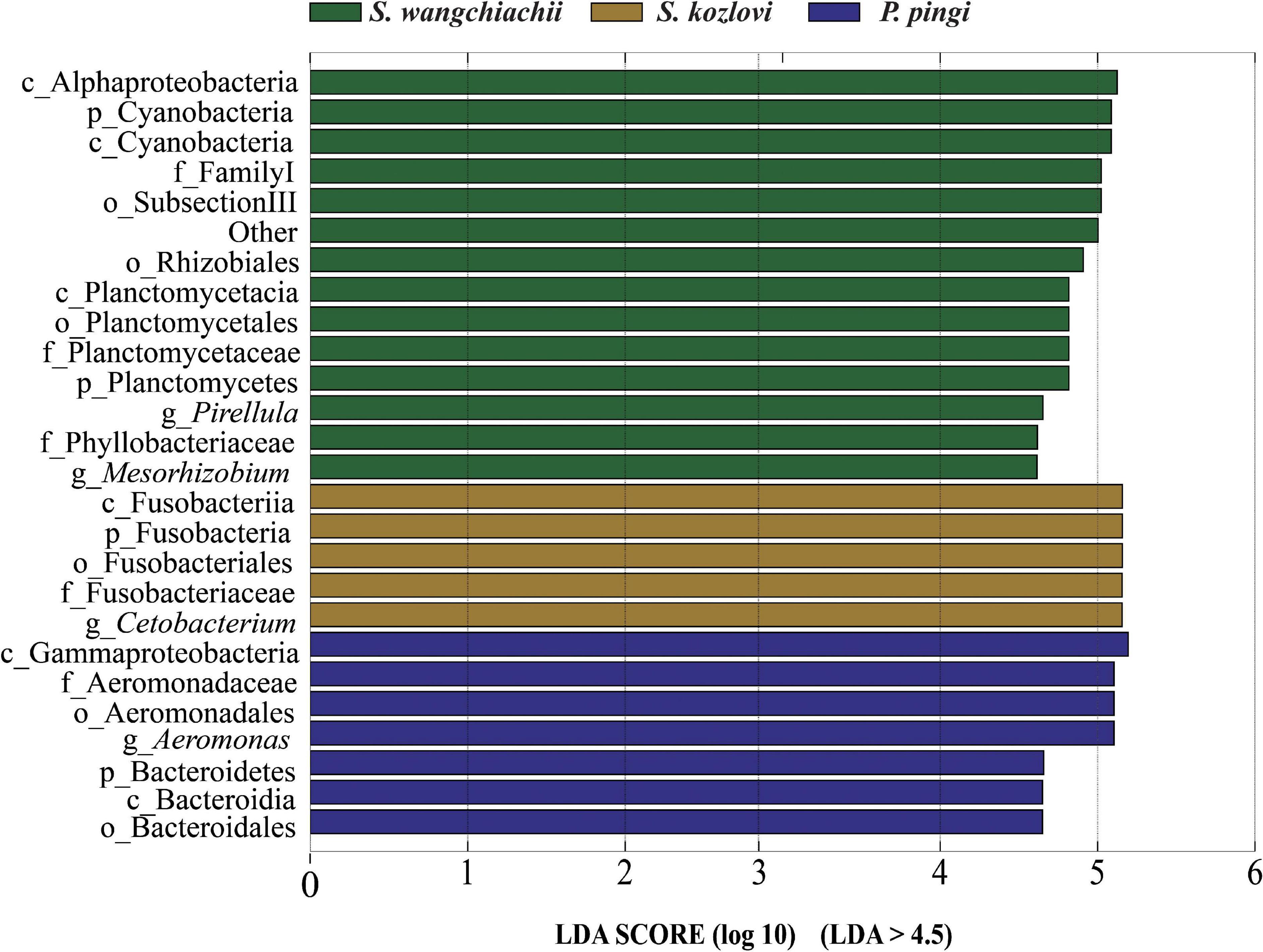
Figure 3. Variation in the composition of the three cold-water fishes. Linear discriminant analysis effect size (LEfSe) analysis of gut microbiota composition of the three fish species (LDA > 4.5).
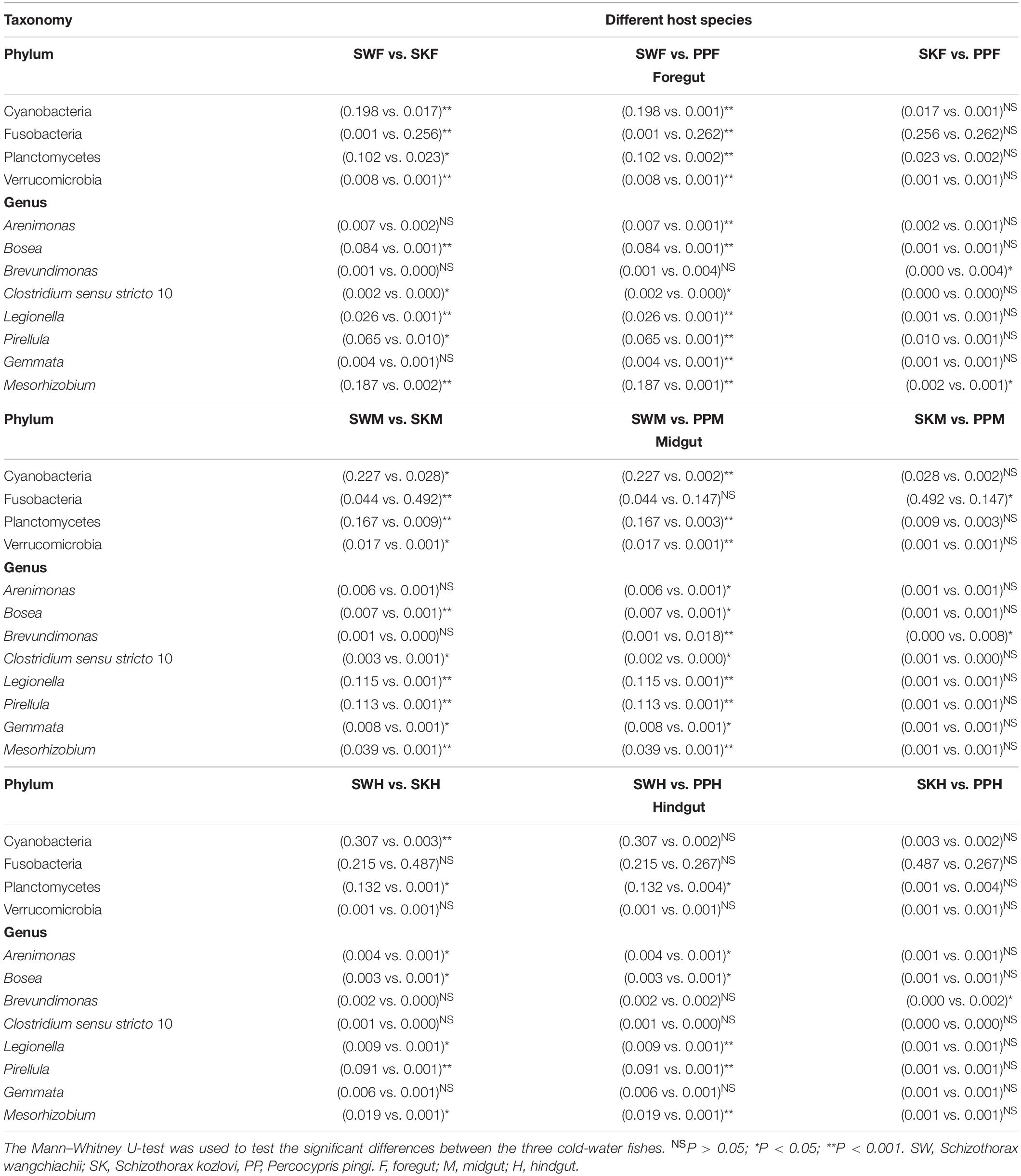
Table 1. Comparison of the relative abundance of gut samples among different host species.
At the genus level, overall, Cetobacterium (SW: 8.53%; SK: 40.95%; PP: 22.38%) and Aeromonas (SW: 0.24%; SK: 21.69%; PP: 27.35%) dominated the bacterial composition (Figure 2C and Supplementary Table 4). The relative abundance of Brevundimonas showed an upward trend from the omnivore (SK), herbivore (SW), to carnivore (PP) (SK: 0%, SW: 0.04%, PP: 2.6%) (Table 1). Significant differences in the relative abundance of Bosea, Clostridium sensu stricto 10, and Pirellula were identified between the three species (Mann–Whitney U-test; both, p < 0.05) (Figure 3 and Table 1). For example, Clostridium sensu stricto 10 was significantly different between SW-F and SK-F (Mann–Whitney U-test; p < 0.01) (Table 1). At the OTU level, the highest number of shared OTUs was identified between SW and SK (number: 89), followed by SW and PP (number: 62), and the lowest between PP and SK (number: 13) (Figure 2D). SW had the highest number of unique OTUs (449), followed by PP (377), and the lowest number of unique OTUs in SK (22) (Figure 2D). Among different intestinal sections (foregut, midgut, and hindgut), the highest number of unique OTUs (498) was detected in SW-M, while the lowest number of unique OTUs (13) was recorded in SK-H (Figure 2D).
Co-occurrence Analysis of the Three Cold-Water Fishes
The co-occurrence analysis showed that the network of gut microbes between the three fish species illustrated distinct co-occurrence patterns (Figure 4). The node in the network diagram mainly belonged to eight phyla: Proteobacteria, Fusobacteria, Firmicutes, Cyanobacteria, Planctomycetes, Bacteria, Bacteroidetes, and Verrucomicrobia (Figure 4). In this study, the bacteria in the center of the co-occurrence were treated as the key taxa (Eiler et al., 2012). SubsectionIII_FamilyI_Unclassified (Cyanobacteria) and Allobaculum (Firmicutes) were the key taxa in SW, and it tended to be positively correlated with Pirellula, Planctomyces, Cyanobacteria_Unclassified, and Gloeocapsa (Figure 4A). The relative abundance of SubsectionIII_FamilyI_ Unclassified showed increased trends from foregut (15%) to midgut (18%) to hindgut (27%) (Supplementary Table 4). The same key taxa were found in SK and PP (Figures 4B,C). Cetobacterium (Fusobacteria) and Streptococcus (Firmicutes) were the key taxa in SK and PP (Figures 4B,C). The higher relative abundance of Cetobacterium (the average relative abundance; SK: 41%; PP: 22%) was three and twofold more than Streptococcus, respectively (the average relative abundance; SK: 0.09%; PP: 0.10%) (Figures 4B,C and Supplementary Table 4).
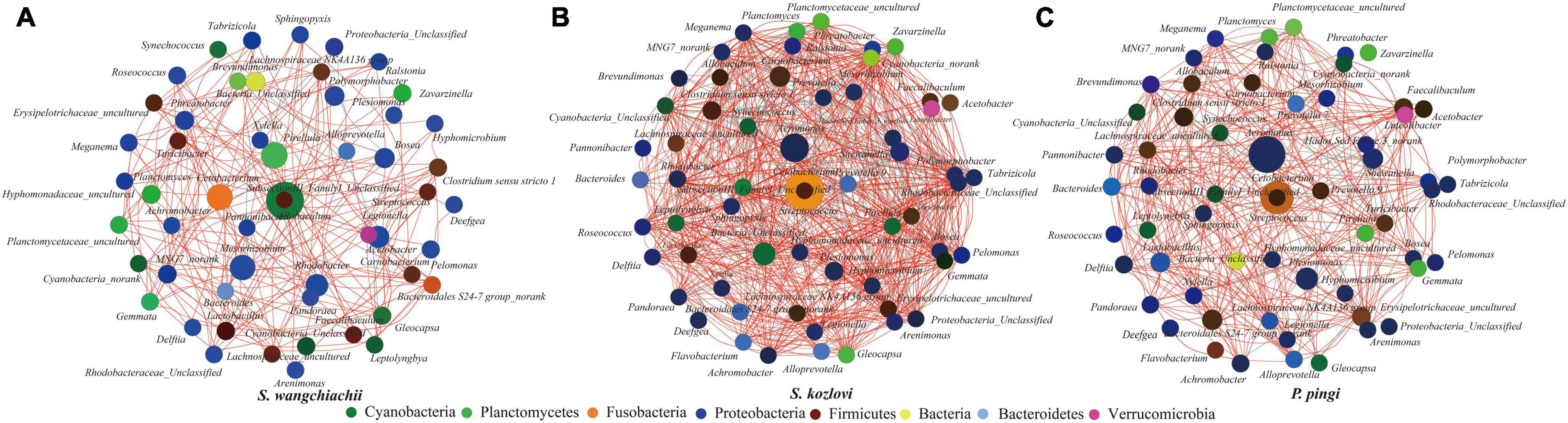
Figure 4. Co-occurrence analysis of the gut microbes (genus level) of the three cold-water fishes (with Spearman index; ρ = 0.5) (A–C). The top 60 genera were used to generate the co-occurrence networks. The node represents the bacteria genus, the size of the node represents the relative abundance, and the color indicates their taxonomic assignment. The red line between the nodes represents a positive correlation, and the blue line indicates a negative correlation.
Alpha and Beta Diversity of Gut Microbes of the Three Cold-Water Fishes
Overall, the Chao 1 and phylogeny indices significantly increased from the omnivorous (SK) to carnivorous (PP) to herbivorous (SW) fish (Kruskal–Wallis H-test, p < 0.05) (Figures 5A,B and Supplementary Table 5). For example, in the midgut (M), the highest Chao 1 index and phylogenetic index were found in SW (mean ± Sd; Chao 1 index: 1,395 ± 211; phylogenetic index: 77 ± 12), followed by PP (Chao 1 index: 466 ± 237; phylogenetic index: 44 ± 18), and the lowest in SK (Chao 1 index: 343 ± 254; phylogenetic index: 23 ± 12) (Figure 5A and Supplementary Table 5). The two alpha diversity indices showed significant differences between the gut microbes of the three fish species (SW, SK, PP) (Kruskal–Wallis H-test, p < 0.05) (Figures 5A,B). At the species level, in SW, the highest Chao 1 index was identified in M (midgut) (mean ± Sd; Chao 1 index: 1,395 ± 211), followed by F (foregut) (Chao 1 index: 1,182 ± 339) and H (hindgut) (Chao 1 index: 995 ± 485) (Supplementary Table 5). In SK, F (Chao 1 index: 457 ± 346) had the highest alpha diversity, followed by M (Chao 1 index: 343 ± 254), and the lowest in H (Chao 1 index: 224 ± 237) (Supplementary Table 5). In PP, the highest Chao 1 index was identified in H (Chao 1 index: 459 ± 365), followed by M (Chao 1 index: 446 ± 237) and F (Chao 1 index: 411 ± 160) (Supplementary Table 5).
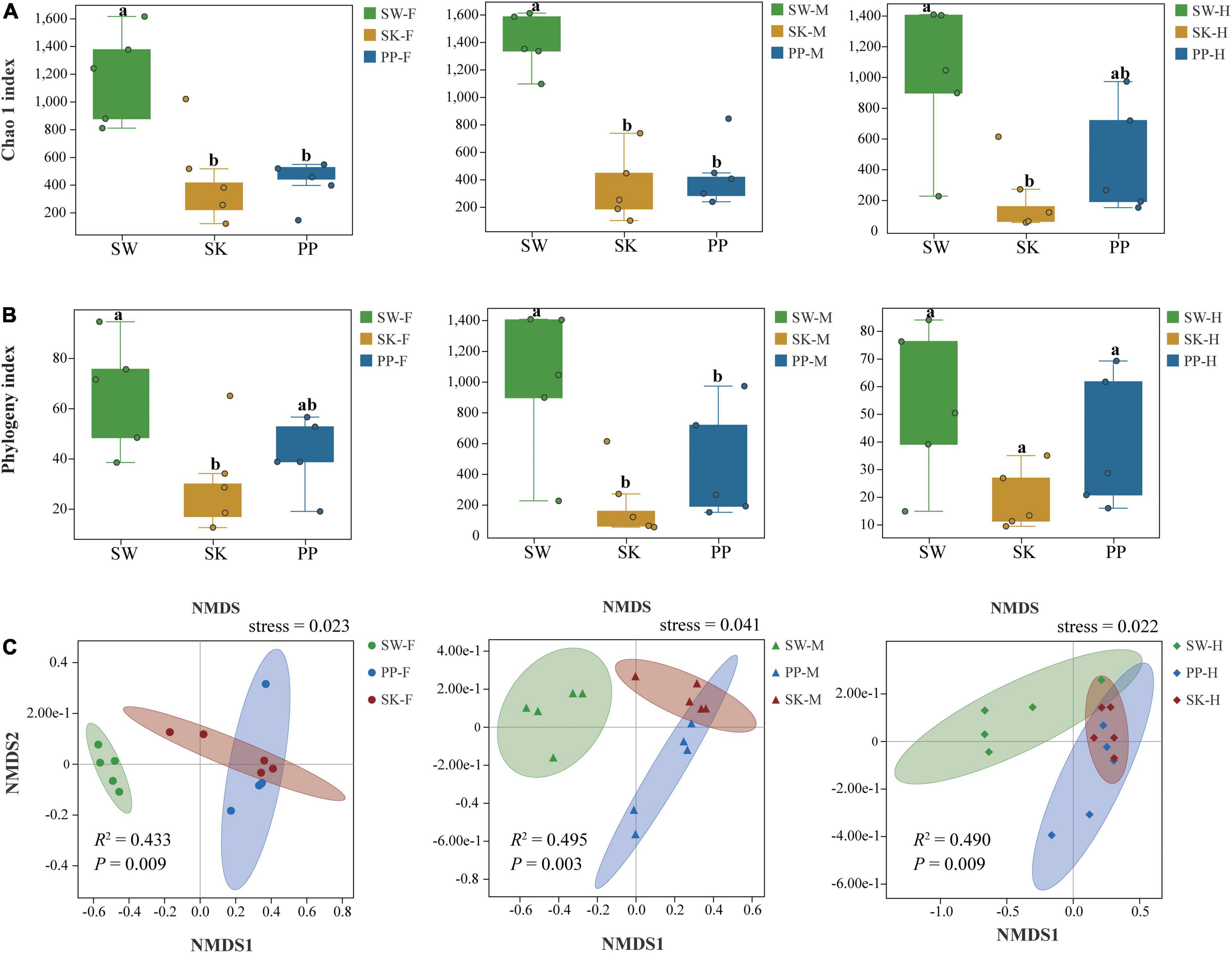
Figure 5. Alpha and beta diversity of gut microbes of the three cold-water fishes. (A) The Chao 1 index was calculated for the foregut, midgut, and hindgut of SW, SK, and PP. (B) The phylogeny index was calculated for the foregut, midgut, and hindgut of SW, SK, and PP. In the box plot, the top line represents the upper quartile (75th percent), the bottom line represents the lower quartile (25th percent), and the line between the top and bottom lines represents the median. The black points represented the outliers. (C) Non-metric multidimensional scaling (NMDS) analysis based on Bray–Curtis distances to explore the dissimilarity in the gut microbes of the three cold-water fishes among different intestinal sections. From left to right: foregut, midgut, and hindgut.
The gut microbial composition of the three cold-water fishes showed significant differences between different species (Figure 5C and Supplementary Figure 4 and Table 2) (PERMANOVA; both, p < 0.05). Among F, M, and H, the NMDS plots showed that the gut microbial communities of SW, SK, and PP were different and could be easily distinguished (Figure 5C). The PERMANOVA test on the three distances had the same result (PERMANOVA; both, p < 0.05) (Table 2). Furthermore, we found that the microbial community composition of SK was more similar to PP than to SW (Figure 5C).
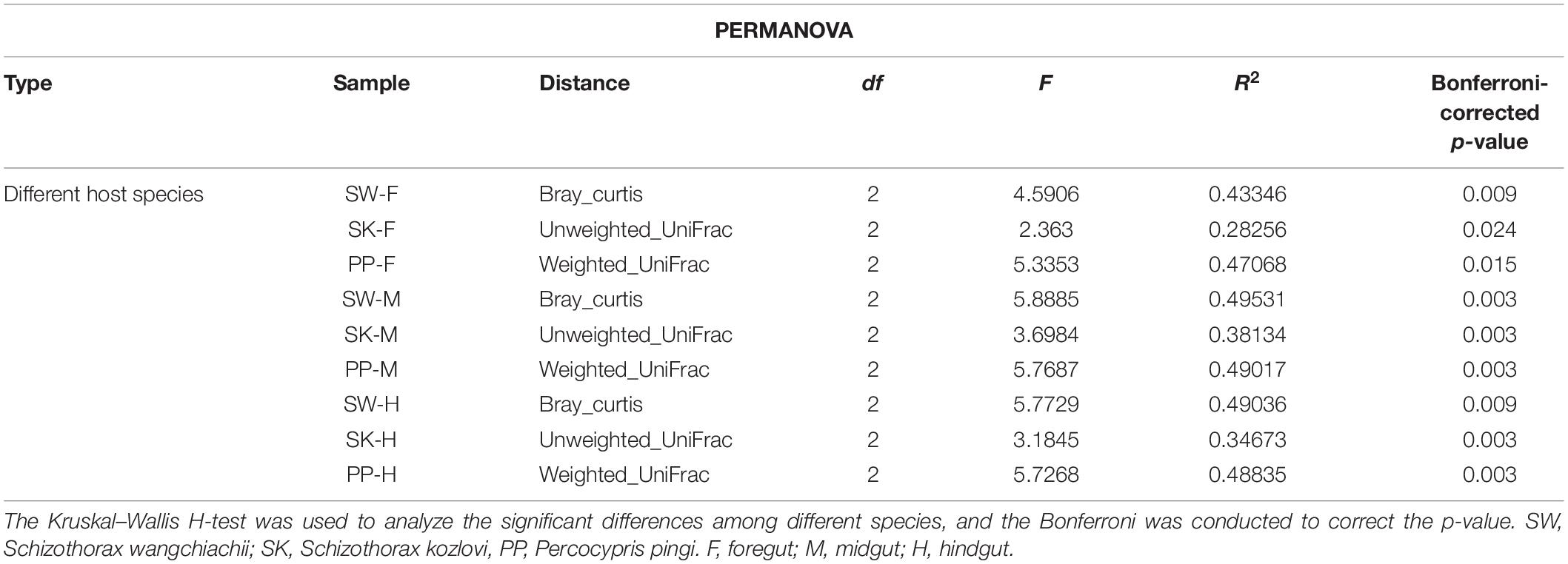
Table 2. Results of the PERMANOVA for the gut samples of the three cold-water fishes.
Potential Functional Groups
Overall, the KEGG pathway analysis (level 2) showed significant differences in the abundance of some diet-related functional categories between the herbivore (SW), omnivore (SK), and carnivore (PP) (one-way ANOVA, both p < 0.05) (Figure 6 and Supplementary Figure 5), for example, metabolism of cofactors and vitamins, amino acid metabolism, membrane transport, and lipid metabolism (Figure 6 and Supplementary Figure 5). Overall, a higher abundance of functional categories was found in SW than in SK and PP (Figures 6B–D). Furthermore, significant differences in the gut microbiota of the three fish species between intestinal sections (foregut, midgut, and hindgut) represented differences in KEGG pathways (level 2) (one-way ANOVA, both p < 0.05) (Figure 6). A higher abundance of metabolism-related functional categories was identified in the midgut (e.g., SW-M and PP-M) than in the foregut and hindgut (Figure 6).
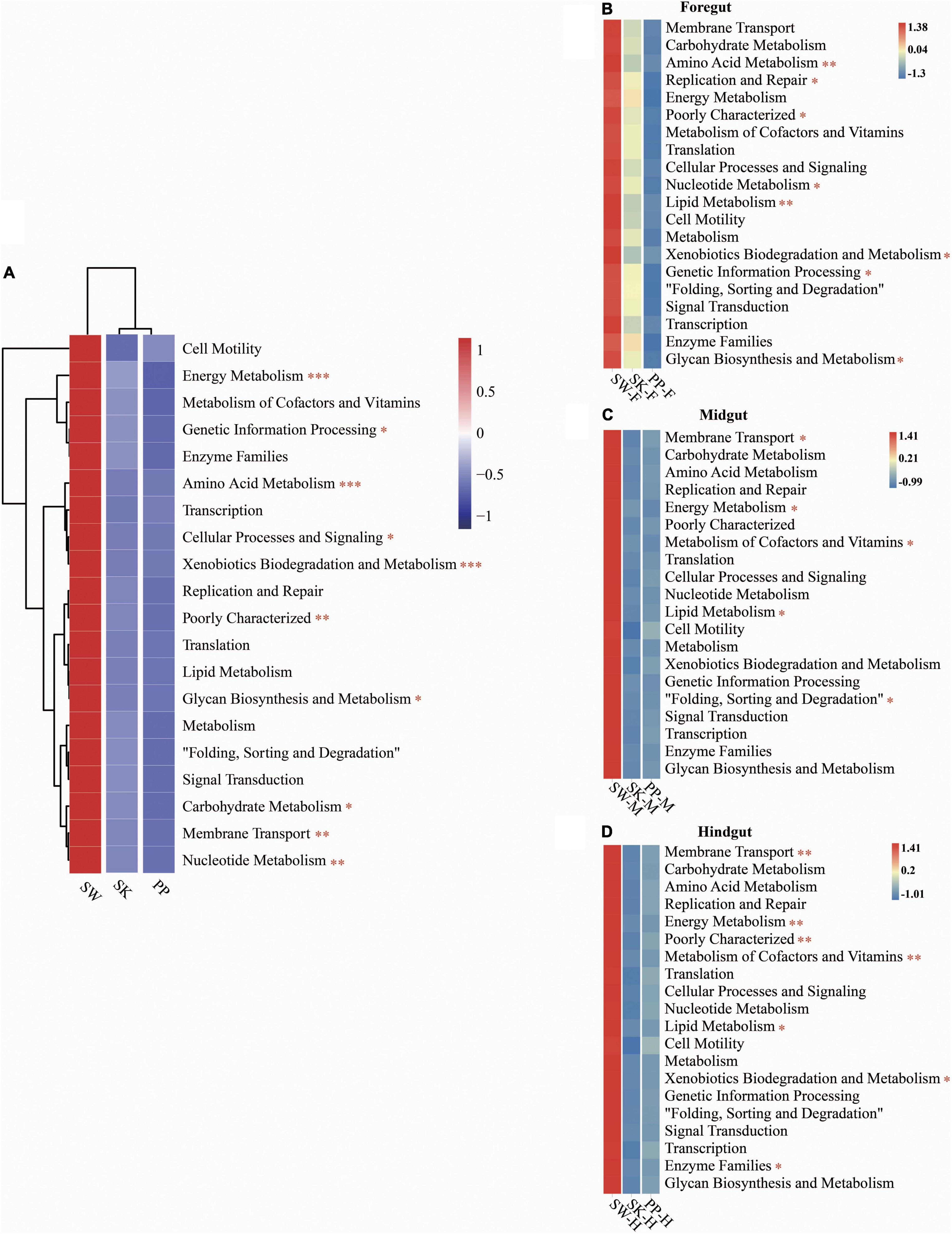
Figure 6. Metabolic functional profiles of the gut microbiome of the three cold-water fishes (KEGG level 2). (A–D) Metabolic functional profiles of the gut microbiome of all species, S. wangchiachii, S. kozlovi, and P. pingi among different intestinal sections. F, foregut; M, midgut; H, hindgut. The one-way analysis of variance (ANOVA) was used to identify significant differences between different samples. *p < 0.05; **p < 0.01; ***p < 0.001.
Microbial Community Assembly Processes of the Three Cold-Water Fishes
The NCM successfully estimated the relationship between the occurrence frequency of OTUs and their relative abundance variations (Figures 7A–C), with 72, 12, and 56% of explained community variance for SW (S. wangchiachii), SK (S. kozlovi), and PP (P. pingi), respectively. Furthermore, the value of R2 was higher in SW (R2 = 0.719) than in PP (R2 = 0.587) (Figures 7A–C). These results indicated that stochastic processes play a key role in shaping the microbial community assembly in SW. The highest Nm-value was found in SW (Nm = 11,837), followed by SK (Nm = 1,676), and the lowest in PP (Nm = 880) (Figures 7A–C). This finding implied that species dispersal of gut microbes was higher in SW than in SK, or PP.
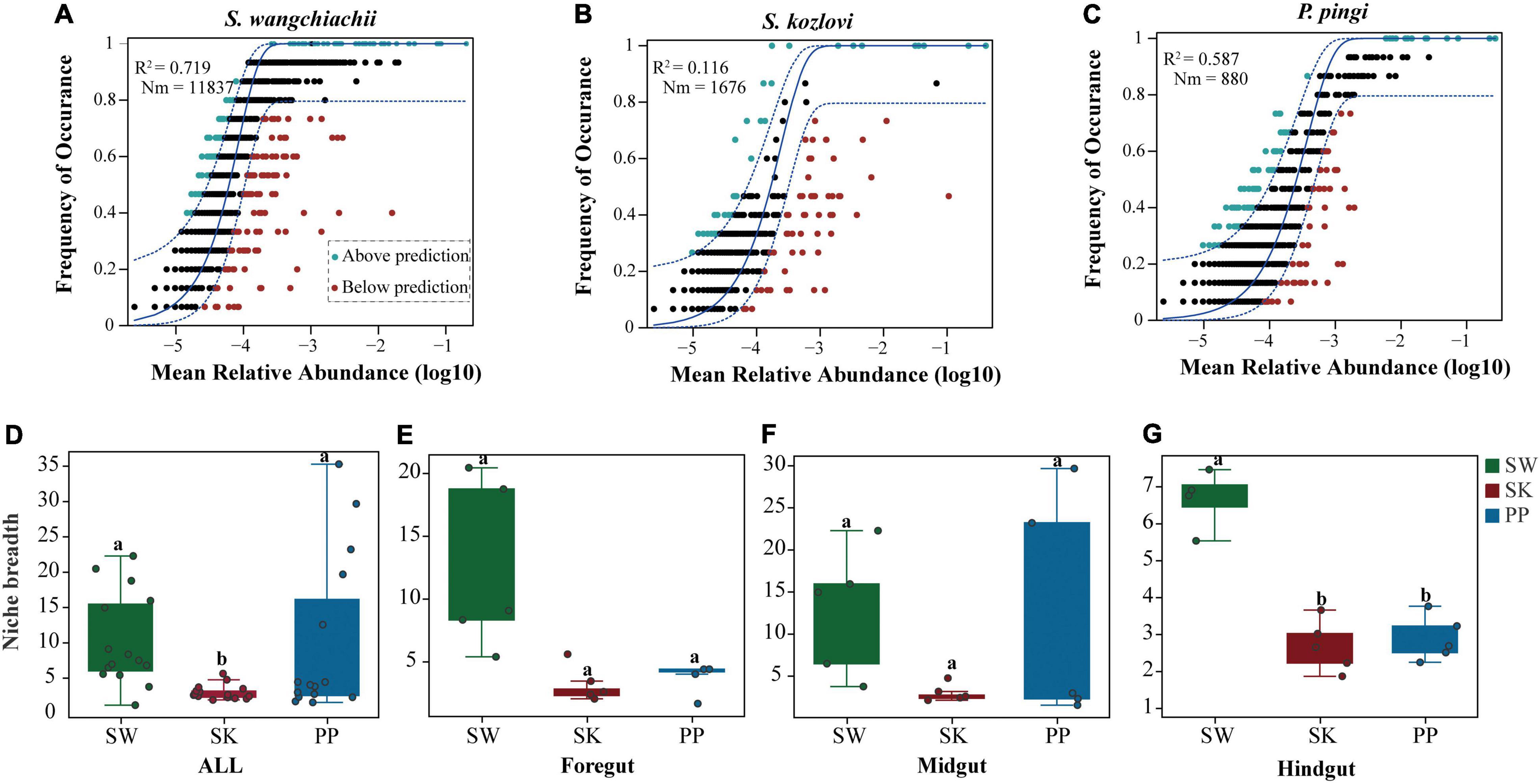
Figure 7. Community assembly process measurements using the niche width and neutral community model (NCM) of the three cold-water fishes. (A–C) Predicted occurrence frequencies of S. wangchiachii, S. kozlovi, and P. pingi gut microbes, respectively. The solid blue line represents the best fit to the NCM, and the dashed blue line indicates 95% confidence intervals around the model prediction. The green point indicates OTUs that occur more frequently than predicted by the NCM, and the red point represents OTUs that occur less frequently than predicted by the model. R2 represents the fit to this model, and Nm represents the metacommunity size times immigration. (D–G) The relative abundance of the niche width of the three cold-water fishes. SW, S. wangchiachii; SK, S. kozlovi; PP, P. ping.
The niche breadth was used to estimate the community-level habitat of the three cold-water fishes (Figures 7D–G). Overall, the highest value of niche breadth was identified in SW (mean ± Sd: 10 ± 7), followed by PP (9 ± 11), and the lowest in SK (3 ± 1) (Figures 7D–G). Moreover, a significant difference in the niche breadth value was found between the three species (Kruskal–Wallis H-test, p < 0.05) (Figures 7D–G).
Discussion
Composition Differences of Gut Microbes of the Three Cold-Water Fishes
Fish gut microbiota plays an important role in nutrition digestion and absorption (Wu et al., 2015; Liu et al., 2016; Li et al., 2017). Previous studies have reported that host feeding on more protein may result in a greater Firmicutes-to-Bacteroidetes ratio of the gut microbiome (De Filippo et al., 2010; Gong et al., 2021). At the phylum level, we found that the Firmicutes-to-Bacteroidetes ratio exhibited an upward trend from SW to SK and PP (Supplementary Table 2). The ratio of protein in food also showed an upward trend from SW to SK to PP (Supplementary Table 1). Therefore, these results implied that the higher Firmicutes-to-Bacteroidetes ratio may help SK and PP to obtain more energy from a protein-rich diet (De Filippo et al., 2010; Gong et al., 2021). Furthermore, the relative abundance of Cyanobacteria in gut microbes significantly decreased from SW to SK and PP (Supplementary Table 2). A previous study showed that Cyanobacteria is an important food source for fish (Currin et al., 2011). Thus, the difference in the relative abundance of Cyanobacteria in the fish gut microbiome is related to the variation in food composition. This result was consistent with our observations. The ratio of algae in food composition showed a downward trend from SW (herbivore) to SK (omnivore) and then to PP (carnivore) (Zhang and Dai, 2011; Chen, 2013; Huang, 2018). Variation in gut microbial composition between species may contribute to digesting food and absorbing nutrients from different dietary compositions (Parata et al., 2020).
Differences in the composition of gut microbes were identified between the three fish species at the genus level (Figure 3 and Table 1). SubsectionIII_FamilyI_Unclassified (Cyanobacteria) was the core bacteria in SW (herbivore) (Figure 4A). Cyanobacteria is one of the main food sources for SW (Huang, 2018). Hence, the core genera SubsectionIII_FamilyI_Unclassified may play a key role in helping SW to digest and absorb nutrients from Cyanobacteria. Furthermore, some cellulose-degrading bacteria were found in the gut microbes of SW (Supplementary Table 4). These findings were in accordance with those of previous studies (Li et al., 2015, 2017; Liu et al., 2016). Clostridium sensu stricto 10 (C. sensu stricto 10), and Clostridium sensu stricto 13 (C. sensu stricto 13) found in herbivores (e.g., grass carp) had the ability to digest xylan, hemicellulose, and cellulose (Uffen, 1997; Uz and Ogram, 2006). Therefore, these bacteria may play an important role in nutrient absorption from periphyton (e.g., diatoms and Cyanobacteria) in SW (Supplementary Table 1). Several protease-producing bacteria (e.g., Cetobacterium, Aeromonas, and C. sensu stricto 1) were observed in SK (omnivore) and PP (carnivore) (Supplementary Table 4). Moreover, Cetobacterium and Aeromonas were the core bacteria in the gut microbes of SK and PP, respectively (Figures 4B,C). A previous study showed that Cetobacterium was the dominant genera in carnivorous fish, with the ability to ferment carbohydrates and peptides (Finegold et al., 2003). Aeromonas plays a vital role in nutrient absorption in fish (Namba et al., 2007; Nayak, 2010). The C. sensu stricto 1 found in carnivorous and omnivorous fish had the ability to break down proteins (Schwab et al., 2011). Therefore, these protease-producing bacteria (e.g., Cetobacterium, Aeromonas, and C. sensu stricto 1) may help SK and PP to utilize nutrition and harvest energy from protein-rich foods (e.g., aquatic insects and fish) (Supplementary Table 1).
Variation of Gut Microbiota Diversity Within the Three Cold-Water Fish Species
Host species can influence the alpha and beta diversity of fish gut microbiomes (Liu et al., 2016; Li et al., 2017). These findings agreed with our results, where we found significant differences in alpha and beta diversities of gut microbes between species (Figure 5). Furthermore, the highest alpha diversity indices were found in SW, followed by PP, and the lowest in SK (Figures 5A,B and Supplementary Table 5). Li et al. (2017) also found higher alpha diversity in the herbivore (M. amblycephala grass carp) than in the omnivore (C. carpio crucian carp) (Li et al., 2017). The higher alpha diversity in SW is likely due to its herbivorous diet, where it feeds on periphyton (e.g., diatoms and Cyanobacteria) (Huang, 2018). The SW needs to swim and forage extensively to acquire more food to meet its growth and reproduction requirements. The gut microbes of SW may acquire more microbes from the aquatic environment than other species and increase the alpha diversity.
Differences in diversity (alpha and beta diversity) of other fish gut microbiomes have been found between intestinal sections (Ye et al., 2014; Parata et al., 2020; Cheutin et al., 2021). The midgut and hindgut microbiomes in surgeonfish (Acanthurus triostegus) represented significant differences in the Shannon index and multi-dimensional scaling (MDS) (Parata et al., 2020). In this study, the midgut exhibited the highest diversity indices, followed by the foregut and hindgut Supplementary Table 5). Our results were inconsistent with those of previous studies (Ye et al., 2014; Yang et al., 2019). The results in our study may be due to a combination of factors. The habitat of the three cold-water fishes surveyed was in lower water temperatures and higher altitudes than the warm-water fishes in Yang et al. (2019). Furthermore, the alpha diversity of fish gut microbiomes between intestinal sections may be affected by variables such as pH (in intestinal lumen), food composition, and oxygen concentration (Ye et al., 2014; Yang et al., 2019). Thus, the cold, high-altitude habitat of these fishes may explain the differences found in other studies.
Compartmentalization of the Functional Differences Between the Three Cold-Water Fishes
The gut microbial function is of great significance in understanding the mechanism of the host adapting to different diets (Gong et al., 2021). The KEGG pathways (level 2) were enriched in diet-related functional categories, such as carbohydrate metabolism, amino acid metabolism, and lipid metabolism that showed differences between species (Figure 6 and Supplementary Figure 5). This result was in agreement with a previous study by Liu et al. (2016) that found that the abundance of carbohydrate metabolism and lipid metabolism showed differences in herbivorous, omnivorous, and carnivorous fishes.
The food compositions were different between the herbivore (SW), omnivore (SK), and carnivore (PP) (Chen, 2013; Zhao, 2014; Huang, 2018; Supplementary Table 1). The periphytic algae were the dominant food of SW and are rich in fatty acids (De Castro Araújoand Garcia, 2005; Huang, 2018). The highest abundance of lipid metabolism was found in SW, followed by PP, and the lowest in SK (Figure 6 and Supplementary Figure 5). Therefore, differences in the relative abundance of lipid metabolism of gut microbes that were identified between species were to adapt to different fatty acid contents of food. The highest abundance was identified in the KEGG pathway of membrane transport in SW. A previous study showed that this pathway may help hosts increase the efficiency of nutrient absorption and adapt to low-temperature environments (Xia et al., 2021). SW (herbivore) is distributed in the upper reaches of the Yangtze River and its tributaries (low water temperature) and feeds on periphytic algae (e.g., diatoms and Cyanobacteria) (Yue, 2000; Huang, 2018). Therefore, the pathway of membrane transport enriched in the gut microbes of SW may play a key role in absorbing nutrients from periphyton and adapting to a low-water temperature environment. In the fish gut microbiome, different intestinal sections (foregut, midgut, and hindgut) play different roles in digesting and absorbing nutrients (Li et al., 2018). It is known that the midgut is the major site of digestion and absorption of nutrients with a higher abundance of diet-related gut microbiome functional categories than other intestinal sections (Chew et al., 2018). In the present study, the abundance of the metabolism pathway (e.g., carbohydrate metabolism, amino acid metabolism, and lipid metabolism) was higher in the midgut than in the foregut and hindgut in the three cold-water fish (Figure 6 and Supplementary Figure 5), which was in accordance with the results of a previous study (Chew et al., 2018). Thus, the gut microbiome of the midgut enriched in diet-related functional categories may play an important role in digesting and absorbing nutrients from the different diets in the three cold-water fishes.
Variation of Gut Microbiota Assembly of the Three Cold-Water Fishes
Studying gut microbiota assembly of animals is crucial to understanding the contribution of ecological processes to the structure of microbial communities in microbial ecology (Sloan et al., 2006; Yan et al., 2016). The NCM and niche breadth are valid approaches for exploring gut microbiota assembly and have been successfully applied to many studies (Yan et al., 2016; Chen et al., 2019). In this study, from SK to PP and then to SW, the contribution of stochastic processes showed an increased trend in shaping the bacterial community structure (Figures 7A–C). This result was consistent with that of Yan et al. (2016) which was focused on the bacterial community assembly processes of herbivorous (Ctenopharyngodon idellus) and carnivorous (Siniperca chuatsi, Silurus meridionalis) species in China. Their results showed that the bacterial community structure of these fish species was mainly driven by stochastic processes (i.e., drift process) (the relative abundance of drift process: Ctenopharyngo_ don idellus: 68%; Siniperca chuatsi: 66%; Silurus meridionalis: 67%) (Yan et al., 2016).
Moreover, our findings showed that the highest species dispersal was found in SW, followed by PP, and the lowest in SK (Figures 7A–C). Similarly, the results of niche breadth agreed with these findings (Figures 7D–G). This phenomenon may be explained by the different feeding behaviors of the three fish species. SW is herbivorous and feeds on low-nutrient algae (e.g., diatoms) (Huang, 2018) and forages extensively across its habitat seeking its food. However, PP and SK feed mainly on fish and aquatic insects, respectively (Zhang and Dai, 2011; Chen, 2013). Thus, compared to PP and SK, SW requires a larger feeding area to obtain food for growth and reproduction. Overall, these findings illustrated that host species influenced the microbial community assembly processes in the three cold-water fishes.
Conclusion
The present study demonstrated that host species influenced the gut microbial composition, diversity, function, and assembly processes of the three cold-water fishes. The gut microbiome of these species had different key bacteria and showed significant differences in several bacteria that were involved in nutrient absorption and diet-related functional categories. The highest alpha diversity indices were identified in the herbivore (SW), followed by the carnivore (PP), and then the omnivore (SK). The gut microbial community showed significant differences between different species. The highest species dispersal was found in SW, followed by PP, and the lowest in SK. Overall, the results implied that increasing the relative abundance of food digesting bacteria and changing the abundance of diet-related pathways may be very important for the three cold-water fishes to digest and uptake nutrients from different foods.
Data Availability Statement
The data presented in this study are deposited in the NCBI repository, accession number PRJNA789150.
Ethics Statement
The animal study was reviewed and approved by the Sichuan Provincial Department of Agriculture and Rural Affairs; Approval Letter of Sichuan Agriculture (2021).
Author Contributions
ZS conceived the project. LX performed the experiments. LX, PX, BZ, KY, FL, ZW, YJ, LD, and WG collected the sample. LX analyzed the data. LX and ZS wrote the manuscript. All authors gave final approval for the publication.
Funding
This study was supported by grants from the Yalong River Hydropower Development Company Ltd. (grant no. YLDC-ZBA-2018116) and the National Natural Science Foundation of China (grant no. 31900373).
Conflict of Interest
LD and WG were employed by the company Yalong River Hydropower Development Company, Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank the Fish Reproduction Station of Jinping-Guandi, China, for providing the place for dissecting experiments. We thank Megan Price for her help with language editing.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.906299/full#supplementary-material
Footnotes
References
Anderson, M. J. (2010). A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 26, 32–46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
Bahram, M., Kohout, P., Anslan, S., Harend, H., Abarenkov, K., and Tedersoo, L. (2016). Stochastic distribution of small soil eukaryotes resulting from high dispersal and drift in a local environment. ISME. J. 10, 885–896. doi: 10.1038/ismej.2015.164
Benjamini, Y., and Yekutieli, D. (2001). The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 29, 1165–1188. doi: 10.1214/aos/1013699998
Caporaso, G., Kuczynsk, I. J., Stombaugh, J., Bittinge, R. K., Bushman, F., Costello, E., et al. (2010). QIIME allows integration and analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Huntley, J., Fierer, N., et al. (2012). Ultra-high-throughput microbial community analysis on the illumina hiseq and miseq platforms. ISME. J. 6, 1621–1624.
Chen, W., Ren, K., Isabwe, A., Chen, H., Liu, M., and Yang, J. (2019). Stochastic processes shape microeukaryotic community assembly in a subtropical river across wet and dry seasons. Microbiome 7:138. doi: 10.1186/s40168-019-0749-8
Chen, Y. (2013). The Genetic Characterization and Population Genetic Diversity of Schizothorax kozlovi Nikolsky. SIchuan: Sichuan Agricultural University. [in Chinese].
Chen, Y., Xu, D., and Shi, W. (2015). Effects of environmental factors on behavior preference of aquatic animal. Chin. Agric. Sci. Bull. 31, 18–24. [in Chinese],
Cheutin, M. C., Villéger, S., Hicks, C. C., Robinson, J. P. W., Graham, N. A. J., Marconnet, C., et al. (2021). Microbial shift in the enteric bacteriome of coral reef fish following climate-driven regime shifts. Microorganisms 9, 1711–1730. doi: 10.3390/microorganisms9081711
Chew, Y. M., Lye, S., Salleh, M., and Yahya, A. (2018). 16S rRNA metagenomic analysis of the symbiotic community structures of bacteria in foregut, midgut, and hindgut of the wood-feeding termite Bulbitermes sp. Symbiosis 76, 187–197. doi: 10.1007/s13199-018-0544-5
Christian, Q., Elmar, P., Pelin, Y., Jan, G., Timmy, S., Pablo, Y., et al. (2013). The silva ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, 590–596. doi: 10.1093/nar/gks1219
Currin, C. A., Levin, L. A., Talley, T. S., Michener, R., and Talley, D. (2011). The role of cyanobacteria in Southern California salt marsh food webs. Mar. Ecol. 32, 346–363. doi: 10.1111/j.1439-0485.2011.00476.x
De Castro Araújo, S., and Garcia, V.M.T. (2005). Growth and biochemical composition of the diatom Chaetoceros cf. wighamii brightwell under different temperature, salinity and carbon dioxide levels. I. protein, carbohydrates and lipids. Aquaculture 246, 405–412. doi: 10.1016/j.aquaculture.2005.02.051
De Filippo, C., Cavalieri, D., Di Paola, M., Ramazzotti, M., Poullet, J. B., Massart, S., et al. (2010). Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. U.S.A 107, 14691–14696. doi: 10.1073/pnas.1005963107
Dehler, C. E., Secombes, C. J., and Martin, S. A. M. (2017). Environmental and physiological factors shape the gut microbiota of Atlantic salmon parr (Salmo salar L.). Aquaculture 467, 149–157. doi: 10.1016/j.aquaculture.2016.07.017
Deng, L. G., Wang, H., and Gan, W. X. (2016). Management of earlier stage operation of fish proliferation and releasing station for Jinping-Guandi Hydropower Station. Water Res. Hydropower Eng. 47, 279–287. [in Chinese] doi: 10.1577/M07-224.1
Dulski, T., Kozłowski, K., and Ciesielski, S. (2020). Habitat and seasonality shape the structure of tench (Tinca tinca L.) gut microbiome. Sci. Rep. 10, 4460–4471. doi: 10.1038/s41598-020-61351-1
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics. 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Egerton, S., Culloty, S., Whooley, J., Stanton, C., and Ross, R. P. (2018). The gut microbiota of marine fish. Front. microbiol. 9:873. doi: 10.3389/fmicb.2018.00873
Eiler, A., Heinrich, F., and Bertilsson, S. (2012). Coherent dynamics and association networks among lake bacterioplankton taxa. ISME. J. 6, 330–342. doi: 10.1038/ismej.2011.113
Faust, K., and Raes, J. (2016). CoNet app: inference of biological association networks using cytoscape. F1000Res. 5:1519. doi: 10.12688/f1000research.9050.2
Feng, H., Xia, Z., Nan, Z., and Lu, Z. (2011). Does hydrologic regime affect fish diversity? A case study of the Yangtze Basin (China). Environ Biol Fishes. 92, 569–584. doi: 10.1007/s10641-011-9880-5
Finegold, S. M., Vaisanen, M.-L., Molitoris, D. R., Tomzynski, T. J., Song, Y., Liu, C., et al. (2003). Cetobacterium somerae sp. nov. from human feces and emended description of the genus Cetobacterium. Syst. Appl. Microbiol. 26, 177–181. doi: 10.1078/072320203322346010
Gipsi, L. M., Faust, K., Henry, N., Decelle, J., Colin, S., Carcillo, F., et al. (2015). Determinants of community structure in the global plankton interactome. Science. 348, 874–875. doi: 10.1126/science.1262073
Gong, L., Liu, B., Wu, H., Feng, J., and Jiang, T. (2021). Seasonal dietary shifts alter the gut microbiota of avivorous bats: implication for adaptation to energy harvest and nutritional utilization. Msphere 6:e0046721. doi: 10.1128/mSphere.00467-21
He, Y., Wu, X., Zhu, Y., Li, H., Li, X., and Yang, D. (2014). Effect of rearing temperature on growth and thermal tolerance of Schizothorax (Racoma) kozlovi larvae and juveniles. J. Therm. Biol. 46, 24–30. doi: 10.1016/j.jtherbio.2014.09.009
Huang, J. (2018). The Study of Ontogeny and Feeding Ecology on Larvae of Schizothorax Wangchiachii. Jingzhou: School of Animal Science Yangtze University. [in Chinese].
Jiang, H., Xie, S. G., Zhao, W. Q., and Chang, J. B. (2007). Changes of fish assemblages after construction of Ertan Reservoir in Yalongjiang River. Acta Hydrobiol. Sinica 4, 532–539. [in Chinese], doi: 10.1016/j.pmrj.2012.09.624
Kolde, R. (2015). pheatmap: Pretty Heatmaps. R Package Version 2.1. Available online at: CRAN.R-project.org/package=pheatmap
Langille, M., Zaneveld, J., Caporaso, J. G., Mcdonald, D., Knights, D., Reyes, J. A., et al. (2013). Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 31, 814–821. doi: 10.1038/nbt.2676
Levins, R. (1968). Evolution in Changing Environments: Some Theoretical Explorations. (MPB-2). Princeton, NJ: Princeton University Press.
Li, T., Long, M., Gatesoupe, F.-J., Zhang, Q., Li, A., and Gong, X. (2015). Comparative analysis of the intestinal bacterial communities in different species of carp by pyrosequencing. Microb. Ecol. 69, 25–36. doi: 10.1007/s00248-014-0480-8
Li, T., Long, M., Li, H., Gatesoupe, F. J., Zhang, X., Zhang, Q., et al. (2017). Multi-omics analysis reveals a correlation between the host phylogeny, gut microbiota and metabolite profiles in cyprinid fishes. Front. microbiol. 8:454. doi: 10.3389/fmicb.2017.00454
Li, X., Yu, Y., Li, C., and Yan, Q. (2018). Comparative study on the gut microbiotas of four economically important Asian carp species. Sci. China. Life. Sci. 61, 696–705. doi: 10.1007/s11427-016-9296-5
Liu, H., Guo, X., Gooneratne, R., Lai, R., Zeng, C., Zhan, F., et al. (2016). The gut microbiome and degradation enzyme activity of wild freshwater fishes influenced by their trophic levels. Sci. Rep. 6, 24340–24352. doi: 10.1038/srep24340
Liu, Y., Zhu, T. B., Wu, X. B., Yan, W. B., and Yang, D. G. (2015). Observation of embryonic and early larval development of Schizothorax wangchiachii. Fish. Sci. 32, 2238–2244. doi: 10.16378/j.cnki.1003-1111.2015.11.003
Namba, A., Mano, N., and Hirose, H. (2007). Phylogenetic analysis of intestinal bacteria and their adhesive capability in relation to the intestinal mucus of carp. J. Appl. Microbiol. 102, 1307–1317. doi: 10.1111/j.1365-2672.2006.03204.x
Nayak, S. K. (2010). Role of gastrointestinal microbiota in fish. Aquacult. Res. 41, 1553–1573. doi: 10.1111/j.1365-2109.2010.02546.x
Neuman, C., Hatje, E., Zarkasi, K. Z., Smullen, R., Bowman, J. P., and Katouli, M. (2016). The effect of diet and environmental temperature on the faecal microbiota of farmed Tasmanian Atlantic Salmon (Salmo salar L.). Aquacult. Res. 47, 660–672.
Parata, L., Nielsen, S., Xing, X., Thomas, T., Egan, S., and Vergés, A. (2020). Age, gut location and diet impact the gut microbiome of a tropical herbivorous surgeonfish. FEMS. Microbiol. Ecol. 96:fiz179. doi: 10.1093/femsec/fiz179
Parks, D. H., Tyson, G. W., Hugenholtz, P., and Beiko, R. G. (2014). STAMP: statistical analysis of taxonomic and functional profiles. Bioinformatics. 30, 3123–3124. doi: 10.1093/bioinformatics/btu494
Rosindell, J., Hubbell, S. P., and Etienne, R. S. (2011). The unified neutral theory of biodiversity and biogeography at age ten. Trends Ecol. Evol. 26, 340–348. doi: 10.1016/j.tree.2011.03.024
Schwab, C., Cristescu, B., Northrup, J. M., Stenhouse, G. B., and Gänzle, M. (2011). Diet and environment shape fecal bacterial microbiota composition and enteric pathogen load of grizzly bears. PLoS. One. 6:e27905. doi: 10.1371/journal.pone.0027905
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W. S., et al. (2011). Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60–R78. doi: 10.1186/gb-2011-12-6-r60
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Sloan, W. T., Lunn, M., Woodcock, S., Head, I. M., Nee, S., and Curtis, T. P. (2006). Quantifying the roles of immigration and chance in shaping prokaryote community structure. Environ. Microbiol. 8, 732–740. doi: 10.1111/j.1462-2920.2005.00956.x
Tao, S., Wang, J., Liu, X., He, F., and Yu, K. (2013). Preliminary research on bacterial community structure and diversity in digestive tract of Miichthys miiuy. South China Fish. Sci. 9, 8–15. [in Chinese], doi: 10.3969/j.issn.2095-0780.2013.04.002
Uffen, R. L. (1997). Xylan degradation: a glimpse at microbial diversity. J. Microbiol. Biotechn. 19, 1–6. doi: 10.1038/sj.jim.2900417
Uz, I., and Ogram, A. V. (2006). Cellulolytic and fermentative guilds in eutrophic soils of the Florida Everglades. FEMS Microbiol. Ecol. 57, 396–408. %J FEMS Microbiology Ecology doi: 10.1111/j.1574-6941.2006.00120.x
Wei, G., Li, M., Li, F., Li, H., and Gao, Z. (2016). Distinct distribution patterns of prokaryotes between sediment and water in the Yellow River estuary. Appl. Microbiol. Biotechnol. 100, 9683–9697. doi: 10.1007/s00253-016-7802-3
Wu, S., Ren, Y., Peng, C., Hao, Y., Xiong, F., Wang, G., et al. (2015). Metatranscriptomic discovery of plant biomass-degrading capacity from grass carp intestinal microbiomes. FEMS Microbiol. Ecol. 91:fiv107. doi: 10.1093/femsec/fiv107
Xia, T., Yao, Y., Wang, C., Dong, M., Wu, Y., Li, D., et al. (2021). Seasonal dynamics of gut microbiota in a cohort of wild Tibetan macaques (Macaca thibetana) in western China. Glob. Ecol. Conserv. 25:e01409. doi: 10.1016/j.gecco.2020.e01409
Yan, Q., Li, J., Yu, Y., Wang, J., He, Z., Van Nostrand, J. D., et al. (2016). Environmental filtering decreases with fish development for the assembly of gut microbiota. Environ. Microbiol. 18, 4739–4754. doi: 10.1111/1462-2920.13365
Yang, G., Jian, S. Q., Cao, H., Wen, C., Hu, B., Peng, M., et al. (2019). Changes in microbiota along the intestine of grass carp (Ctenopharyngodon idella): community, interspecific interactions, and functions. Aquaculture 498, 151–161. doi: 10.1016/j.aquaculture.2018.08.062
Ye, L., Amberg, J., Chapman, D., Gaikowski, M., and Liu, W.-T. (2014). Fish gut microbiota analysis differentiates physiology and behavior of invasive Asian carp and indigenous American fish. ISME J. 8, 541–551. doi: 10.1038/ismej.2013.181
Yue, P. (2000). Fauna Sinica. Osteichthys. Cypriniformes III. HONG KONG: China Scientific Book Services. [in Chinese].
Zhang, X., and Dai, Y. (2011). Feeding habitsand resources protection of Schizothorax kozlovi. J. Hydroecol. 32, 1–5. [in Chinese],
Zhao, G. (2014). Embryonic and postembryonic development of Percocypris pingi (Tchang). Agric. Biotechnol. 3, 51–56. [in Chinese],
Zhao, Z., Jiang, J., Zheng, J., Pan, Y., Dong, Y., Chen, Z., et al. (2022). Exploiting the gut microbiota to predict the origins and quality traits of cultured sea cucumbers. Environ. Microbiol. doi: 10.1111/1462-2920.15972 [Epub ahead of print].
Keywords: gut microbes, host species, core bacteria, cold-water fish, stochastic processes, dietary adaptation
Citation: Xu L, Xiang P, Zhang B, Yang K, Liu F, Wang Z, Jin Y, Deng L, Gan W and Song Z (2022) Host Species Influence the Gut Microbiota of Endemic Cold-Water Fish in Upper Yangtze River. Front. Microbiol. 13:906299. doi: 10.3389/fmicb.2022.906299
Received: 28 March 2022; Accepted: 22 June 2022;
Published: 18 July 2022.
Edited by:
Ram Krishan Negi, University of Delhi, IndiaReviewed by:
Himani Khurana, University of Delhi, IndiaCarlos Puch Hau, Center for Research and Advanced Studies—Mérida Unit, Mexico
Copyright © 2022 Xu, Xiang, Zhang, Yang, Liu, Wang, Jin, Deng, Gan and Song. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhaobin Song, zbsong@scu.edu.cn