 Laura García-Descalzo
Laura García-Descalzo Eva García-López2
Eva García-López2- 1Centro de Astrobiología, Department of Planetology and Habitability, CSIC-INTA, Madrid, Spain
- 2Centro de Astrobiología, Department of Molecular Ecology, CSIC-INTA, Madrid, Spain
The old debate of nature (genes) vs. nurture (environmental variables) is once again topical concerning the effect of climate change on environmental microorganisms. Specifically, the Polar Regions are experiencing a drastic increase in temperature caused by the rise in greenhouse gas emissions. This study, in an attempt to mimic the molecular adaptation of polar microorganisms, combines proteomic approaches with a classical microbiological analysis in three bacterial species Shewanella oneidensis, Shewanella frigidimarina, and Psychrobacter frigidicola. Both shewanellas are members of the same genus but they live in different environments. On the other hand, Shewanella frigidimarina and Psychrobacter frigidicola share the same natural environment but belong to a different genus. The comparison of the strategies employed by each bacterial species estimates the contribution of genome vs. environmental variables in the adaptation to temperature. The results show a greater versatility of acclimatization for the genus Shewanella with respect to Psychrobacter. Besides, S. frigidimarina was the best-adapted species to thermal variations in the temperature range 4–30°C and displayed several adaptation mechanisms common with the other two species. Regarding the molecular machinery used by these bacteria to face the consequences of temperature changes, chaperones have a pivoting role. They form complexes with other proteins in the response to the environment, establishing cooperation with transmembrane proteins, elongation factors, and proteins for protection against oxidative damage.
Introduction
Cold is the most frequently spread natural condition that causes stress to live organisms (Rodrigues and Tiedje, 2008). Cold ecosystems are widespread on Earth, given that 80% of the biosphere is permanently frozen (Russell, 1990). About 90% of oceans hold a temperature of ≤5°C (Rodrigues and Tiedje, 2008), and, along with the seas, constitute 70% of the surface of the planet with a medium temperature of 2°C.
The low temperature causes changes in the properties of water even if it does not freeze (Rodrigues and Tiedje, 2008). This restricts its availability for biochemical reactions, limiting the survival of microorganisms that are unable to develop adaptive mechanisms to cope with it. When the temperature decreases, the diffusion of water drops, and its viscosity increases, reducing substrate diffusion rates and disrupting the transport of nutrients in the cells (Rodrigues and Tiedje, 2008). However, the low temperature does also alter other important parameters in cells, such as enzyme activity, membrane fluidity, transcription, translation, cellular division rates, protein folding, and denaturation (Russell, 1990; D’Amico et al., 2006; Rodrigues and Tiedje, 2008; Schumann, 2009; Alcazar et al., 2010).
The study of extremophiles has increased the discovery of new microorganisms and has contributed to understanding how they respond to evolving challenges over time at the molecular level (Irwin and Baird, 2004). Regarding the temperature of growth, microorganisms have been conventionally classified into psychrophiles (cold loving), psychrotrophs (cold tolerant), mesophiles (moderate temperature loving), and thermophiles (heat loving) (Tortora et al., 2010). Psychrophiles are broadly distributed and have developed successful strategies to colonize cold environments and even regions under temperature fluctuations (De Maayer et al., 2014). These strategies include the upward regulation of some genes, enhanced membrane transport, increased biosynthesis of carotenoid pigments, and compatible osmotic solutes (De Maayer et al., 2014; Cray et al., 2015).
Nevertheless, the main actors in these adaptive responses are the proteins, which have a key role in the molecular mechanisms of the cell. They control the balance between nutrients and products, the assembly of macromolecules, the dynamic of nucleic acids, and the correct folding of other proteins (D’Amico et al., 2006). Thus, changes in the composition of the proteome in response to cellular stress conditions are linked with specific adaptive strategies (Maier et al., 2011). Chaperones are a group of essential proteins involved in the quality control of cell machinery (Hartl and Hayer-Hartl, 2002) and in dealing with the consequences of stress (Cooper and Hausman, 2000; Hochachka and Somero, 2002; Beckerman, 2005; Cloutier and Coulombe, 2013). They are constituent proteins responsible for the correct folding and building of other proteins and for eliminating those incorrectly formed during synthesis or as a result of a cell stress situation (Hartl and Hayer-Hartl, 2002; Camberg et al., 2013).
Among the chaperone family, heat-shock proteins (Hsp) play a well-known role in the prevention of aberrant protein aggregates as well as in the facilitation of protein synthesis, translocation, de novo folding, and the assembly of multi-protein complexes (Bromfield and Nixon, 2013). Their amount increases in the cell during stress situations like changes in the temperature of the environment. Not only does heat induce these proteins but the exposure of cells to low temperature can also lead to the synthesis of some Hsps (Hochachka and Somero, 2002; García-Descalzo et al., 2011). The main families of Hsps are highly conserved and classified based on their sequence and their molecular weight (Makhnevych and Houry, 2013). Two large and important families are Hsp70 and Hsp60 (Cooper and Hausman, 2000; Beckerman, 2005). Sometimes, members of these families act together sequentially during protein transport and folding of newly synthesized proteins (Langer et al., 1992; Cooper and Hausman, 2000). Protein DnaK is the main representative of the Hsp70 family in the prokaryotes (Thompson et al., 2012), and it is part of a chaperone system together with other co-chaperones, such as DnaJ, from the Hsp 40 family, and the nucleotide exchange factor GrpE (Ahmad et al., 2011; Perrody et al., 2012; Thompson et al., 2012). In the Hsp60 family, a great number of chaperone complexes are activated by cellular stress, including the GroEL-GroES complex (Ventura et al., 2004), which are responsible for facilitating the folding of other proteins from an incorrect or unfolded conformation to their native form in stressful situations (Cooper and Hausman, 2000).
Extremophiles employ different adaptive approaches to cope with environmental changes (Lin, 2008; Metpally and Reddy, 2009; Paredes et al., 2011). Their comparison is a very valuable tool to identify crucial features of adaptation to cold and fluctuating environments. With this purpose, we designed a comparative proteomic study of three bacteria: a mesophile, a psychrophile, and a psychrotroph. They were subjected to different growth temperature conditions to study the underlying molecular actors that are activated under diverse environmental scenarios. Two of these bacteria belong to the same genus but live in environments with different temperatures, while the other two share the same environment but belong to a different genus. In this way, our findings will clarify to what extent the strategies used by these microorganisms are derived from a similar genome or a similar environment. Detailed information about each strain is summarized in Table 1.
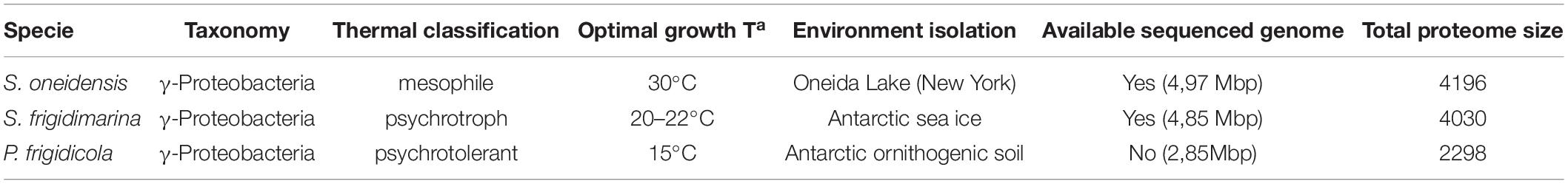
Table 1. Main features of the species used.
Moreover, the knowledge of these molecular mechanisms will have application in different research fields, including biotechnology, medicine, astrobiology, ecology, and climate change investigations (Cavicchioli et al., 2002). The molecules involved in these mechanisms range from enzymes that remain active with high efficiency at very low temperature to be used in the food industry (or in cosmetic or textile detergent production) (Cavicchioli et al., 2002) to biosensors that can report about the state of an ecosystem (Irwin and Baird, 2004), providing valuable information related to climate change or even to biomarkers of habitability in planetary exploration and planetary analog research.
Materials and Methods
Strains and Culture Conditions
Bacteria included in this study were purchased from culture collections: Shewanella oneidensis (MR-1, 700550TM) and Shewanella frigidimarina (ACAM 591, 700753TM) from the Germany Microorganisms and Culture Cell collection (DMSZ) and Psychrobacter frigidicola (ACAM 304, 700361TM) from the American collection (ATCC®). The two first strains belong to the same genus but are distinct regarding their optimal temperature condition and natural environment. S. oneidensis is a mesophile, while S. frigidimarina is a psychrotolerant and P. frigidicola belongs to a different genus and is a psychrophile.
Individual pre-inocula of the three strains were prepared from vials stored in glycerol at −80°C. Aliquots of 100 μl from these vials were incubated at their corresponding optimal growth temperature in tubes with 5 ml of each specific broth (BactoMarine Broth Difco2216 – MB – for S. frigidimarina and P. frigidicola and Luria-Bertani Broth – LB – for S. oneidensis). Once they reached the half exponential phase (corresponding to an optical density (O.D.) at 600 nm about 0.5), 500 μl of each one was inoculated in 250 ml Erlenmeyer flasks with broth at a final volume of 100 ml. Flasks of each species were incubated in triplicate at each experimental temperature of 4, 12, and 30°C for 5 days in aerobic conditions (150 rpm stirring). The O.D. at 600 nm was checked two times per day until it reached the stationary growth phase. Then, pellets of each flask were obtained by centrifugation for 20 min (10,000 × g; 4°C) and washed with phosphate buffer saline (PBS). Pellets were used to extract proteins for subsequent experiments. For the growth curves, statistical analyses were performed using GraphPad Prism version 7.0 (GraphPad Software, La Jolla, CA, United States1). The data were the mean ± SD from three replicates.
Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis and Western-Blot
Proteins from the pellets were extracted to study the potential differences in protein profiles of the three bacteria against different temperature conditions. The preparation of cell extracts and protein determination were performed as described in García-Descalzo et al. (2014).
Protein extracts were separated and analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) for 1 h at 200 V and 400 mA (PowerPac 200, BioRad) in 12% acrylamide-bisacrylamide gels (2.6% crosslinking) using low molecular weight (LMW) (GE Healthcare) marker. Gels were stained with Coomassie Blue or transferred to polyvinyl-difluoride membranes (PVDF) to perform chaperones immunodetection by western blot (WB).
Membranes were firstly incubated overnight in a cold chamber with antibodies anti-DnaK, anti-GroEL, anti-DnaJ, and anti-GroES (Table 2). A second incubation was performed with HRP-linked anti-rabbit or anti-mouse (1:1,000) for 1 h. After several washes with PBS, membranes were revealed by chemiluminescence using ECL Western blotting detection reagents (GE Healthcare).

Table 2. Antibodies for WB and the dilution used.
Two-Dimensional Fluorescence Difference Gel Electrophoresis
The proteins produced in each bacterium at the analyzed temperatures were separated by 2-DE (two-dimensional electrophoresis). Samples (150 μg from each extract) were prepared as previously described (García-Descalzo et al., 2014) with slight modifications. After the first dimension by IEF (isoelectric focusing), which separates proteins based on their isoelectric points, IPG-strips (Immobiline Dry Strips, GE; pH 3-11 NL – non-linear – of 24 cm) were equilibrated in two successive steps with SDS equilibration buffer solutions containing a Tris buffer, 100 mM; urea, 6 M; glycerol, 30% (w/v); and DTT, 0.5% (w/v). Then, they were incubated with iodoacetamide, 4.5% (w/v). The second dimension in SDS-PAGE, to separate proteins by their molecular weight, was carried out on 12% acrylamide (2.6% crosslinking) gels (1.-mm thick). Gels were stained with Coomassie Blue or with MALDI-MS-compatible silver reagent for protein identification (Miller et al., 2006).
To compare the proteins over and under synthesized in each temperature condition, proteins from bacterial cells grown at 4 and 30°C were analyzed by 2-D DIGE. Proteins were extracted from the cultures as indicated before, obtaining four biological replicates of each one. Samples (40 μg) from each experimental condition were made using 7 M urea/2 M thiourea and labeled with DIGE Fluor minimal dyes (GE Healthcare) for their electrophoresis separation and analysis. Two replicates of each experimental condition were labeled with Cy3 and the other two with Cy5. A pool of both temperature conditions was also labeled with Cy2 to be used as an internal standard.
Protein gel images were processed and analyzed using the DeCyder Differential In-gel Analysis (DIA). The spots were co-detected, quantified, normalized, and matched between the four replicate experiments. Protein abundance changes between samples from each strain grouped by the temperatures 4 and 30°C (4/30 ratio) were examined by ANOVA test. Spots present in at least three of four gels per group, with significant ANOVA test (p ≤ 0.05 or p ≤ 0.01), and an averaged 4/30 ratio ≥ ± 2 were considered and selected for further MS analysis in preparative gels. Those spots selected in new gels were excised, digested with trypsin, and identified by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS).
The identification protocol and data search used were based on the method previously described in Cid et al. (2010) and García-Descalzo et al. (2014) with minor variations. Combined peptide mass fingerprint and MS/MS ion search modes were applied against the NCBInr databases (NCBInr 081217 with 7463447 sequences and 2570742364 residues for S. oneidensis; NCBInr 160909 with 9694989 sequences and 3312496757 residues for S. frigidimarina; NCBInr 20131020 with 33055681 sequences and 11532217697 residues and NCBInr 14042010 with 10866589 sequences and 3703552722 residues for P. frigidicola). The MASCOTMT (Matrix Science, London, United Kingdom) database search algorithm was used for protein identification (search parameters: Enzyme-trypsin; fixed modifications carbamidomethyl (C); variable modifications-oxidation (M); mass values-monoisotopic; protein mass unrestricted; peptide mass tolerance-±50 ppm; peptide charge State- 1+; max-missed cleavages-1).
Immunoprecipitation
Cell extracts were immunoprecipitated with monoclonal anti-DnaK and anti-GroEL antibodies (Enzo Life Sciences, United States) to study the DnaK- and the GroEL-associated proteins in the three bacteria studied.
The immunoprecipitation was carried out using Protein G-Sepharose previously washed with PBS. In the first-step, 600 μg of protein extract from each sample was mixed with 3 μl (1 mg/ml) of an antibody (anti-DnaK or anti-GroEL) for 4 h at 4°C with rotation in an immunoprecipitation buffer (IPB) (Tris HCl, 500 mM; pH: 7.5; NaCl, 200 mM; 0.2% Triton X-100; 0.2% NP-40; EDTA, 5 mM; pH: 8, protease inhibitors – Roche – and phenylmethylsulphonyl fluoride, PMSF, 1 mM). The immunoprecipitation was developed in stringent conditions to minimize unspecific complexes. Second, 30 μl of washed Protein-G-Sepharose (50% of slurry) was added, followed by 1 h of incubation in the same previous conditions. Finally, the immunoprecipitates were washed two times in a cold isotonic protease inhibition buffer (IPIB) (Tris HCl, 1 M; pH: 7.5; EDTA, 0.2 M; NaCl, 150 mM; 0.2% SDS) with protease inhibitors (Roche) to remove the unspecific binding. Then, samples were centrifuged for 5 min at 5,000 rpm, discarding the supernatant. The solid phase was subjected to two-dimensional electrophoresis (2 DE), and the subsequent protein identification was carried out by MALDI-TOF MS or kept at −20°C until use.
Results
Cell Growth at 4 and 30°C (and 12°C)
Despite having different optimal temperatures for growth, the three bacteria studied were able to adapt to changing laboratory conditions, mimicking some of the extreme temperatures they could experience in their natural ecosystems. Their growth achieved high O.D. values in the established temperature range, although their lag phase was very different (Figure 1 and Table 3). In the three bacteria, the growth at 4°C demanded a large lag phase; while warmer temperatures depicted a quicker onset, regardless of whether they were mesophile, psychrotolerant, or strict psychrophile. The differences between lag phase duration were statistically significant for the three strains at 4°C as well as for the case of P. frigidicola compared to the other two strains. The exponential phase duration when bacteria grew at 4°C was considerably larger when they grew at 30°C for the three species. Growth rates of each strain at each temperature treatment showed significant differences between S. frigidimarina and S. oneidensis, growing at 4 and 30°C and between S. frigidimarina and P. frigidicola, growing at 12 and 30°C (Table 4). For the two Shewanellas, the highest rate appeared at 30°C, while the rate of P. frigidicola is quite similar in the three temperatures tested.
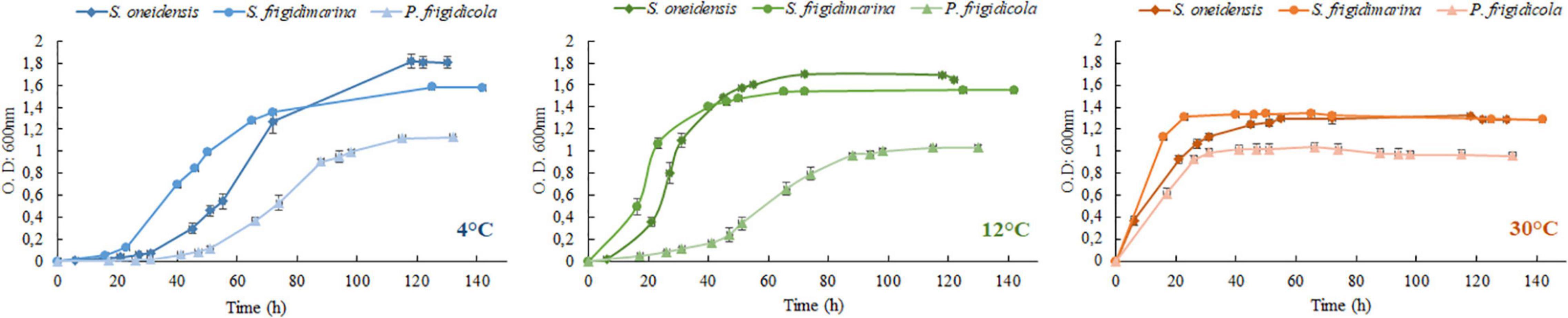
Figure 1. Growth curves of S. oneidensis, S. frigidimarina, and P. frigidicola at 4 (blue), 12 (green), and 30°C (red/orange).
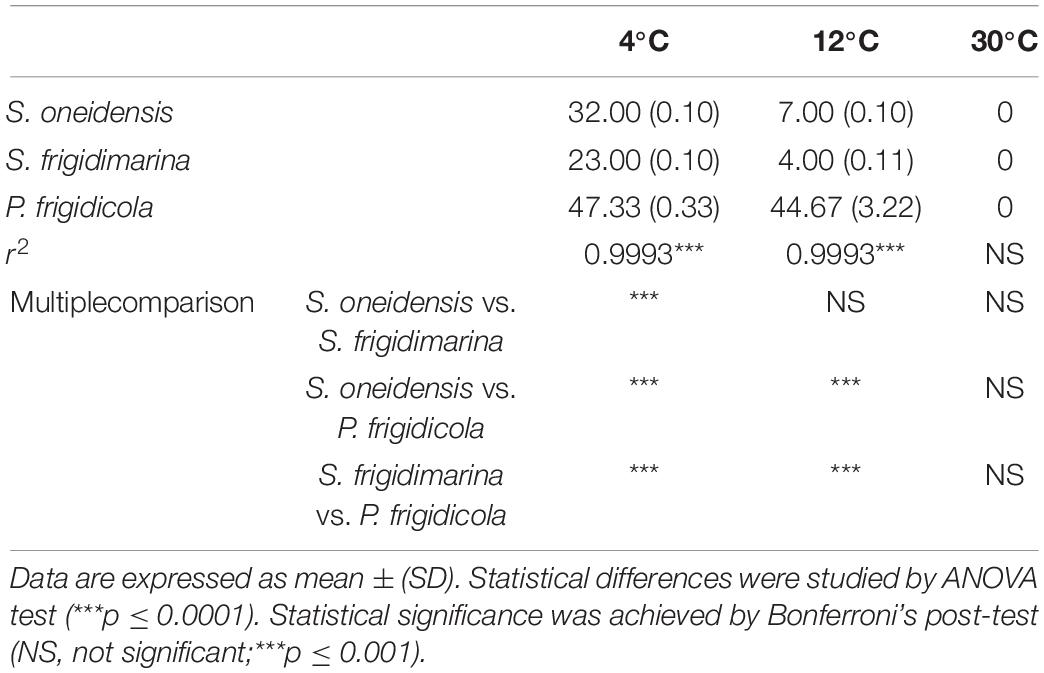
Table 3. Duration of the lag phase in each bacterium at each temperature studied.
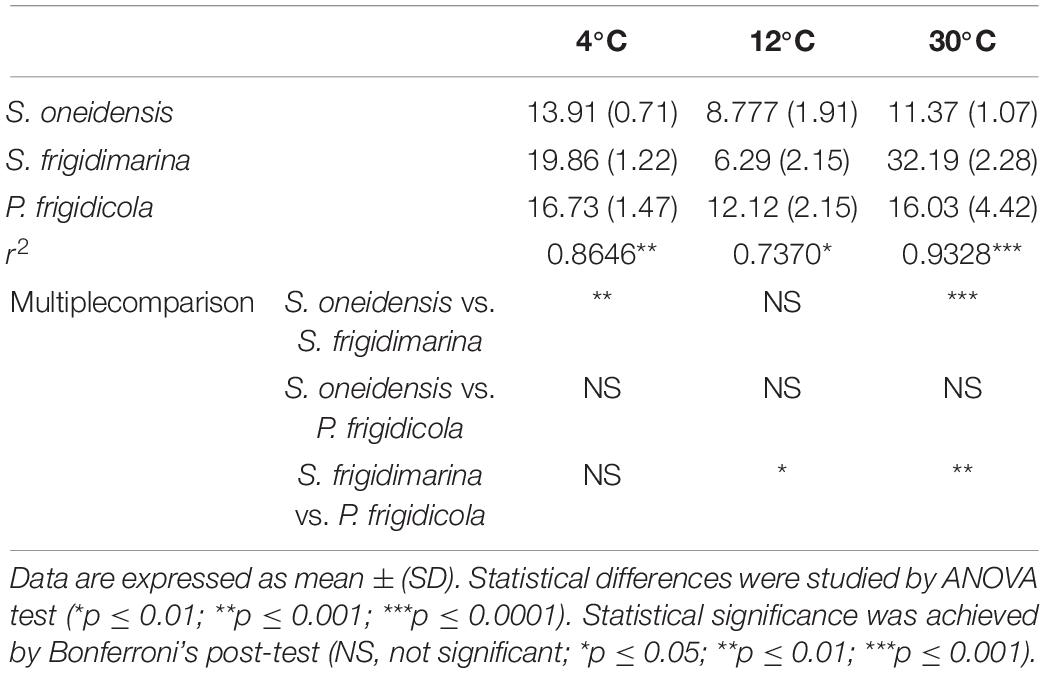
Table 4. Growth rates (h–1) of S. oneidensis, S. frigidimarina, and P. frigidicola at 4, 12, and 30°C.
Synthesis Levels of Chaperones With Temperature Changes Immunodetected by Western Blot
The relative synthesis levels of chaperones GroEL, GroES, DnaK, and DnaJ at 4 and 30°C for each bacterium were studied by WB. Cultures of the mesophile, S. oneidensis, at 30°C showed strong signals of chaperones GroEL/GroES and DnaK (Figure 2A). Cultures at 4°C revealed a weak band corresponding to DnaK, while a strong immunodetection of chaperonine GoES was observed (Figure 2A). These results may suggest that the machinery activated and controlled by these chaperones could be an advantage for the mesophile bacterium, allowing it to thrive in cold conditions and to grow even at temperatures far from its optimum.
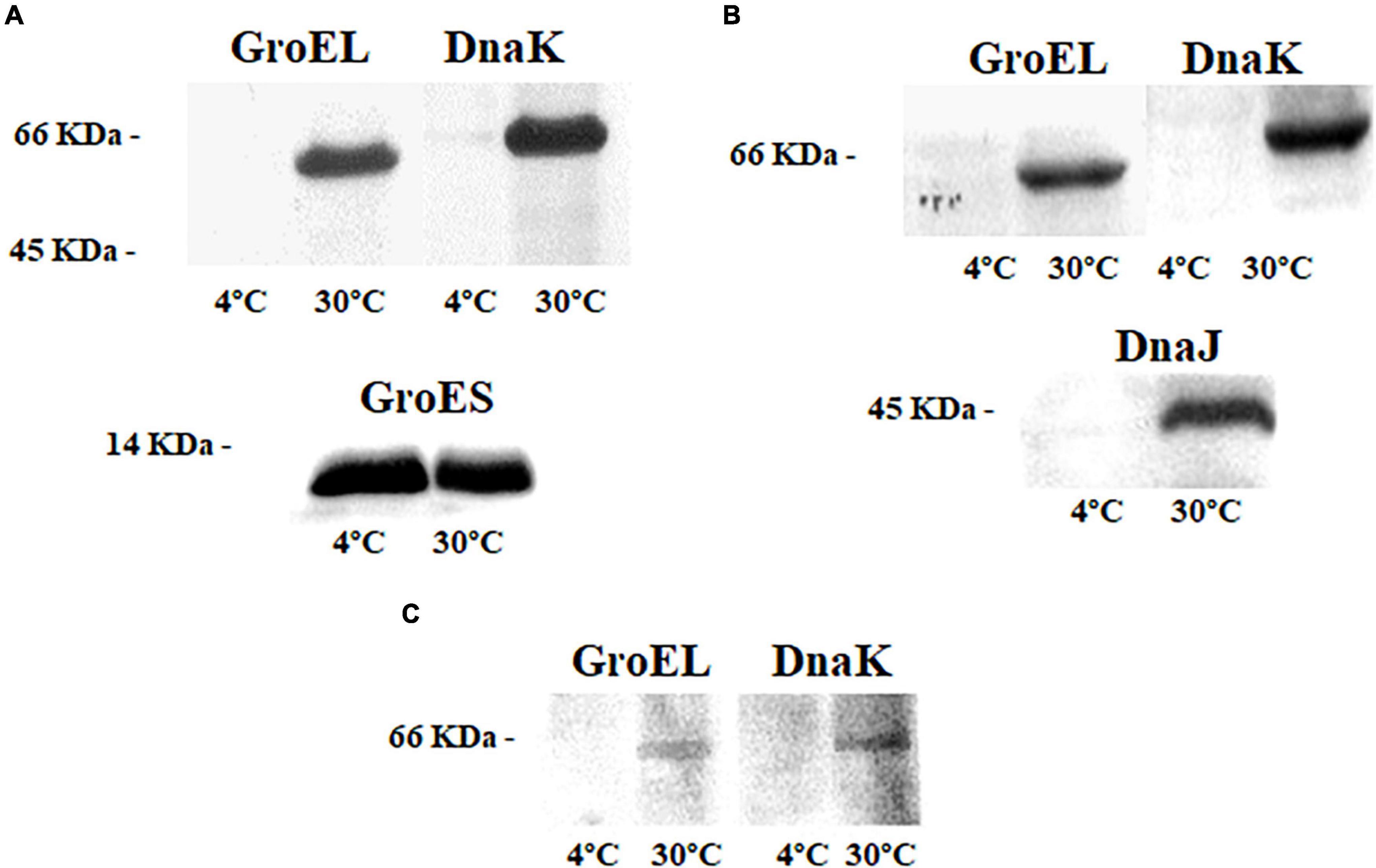
Figure 2. Immunodetection of chaperones in the cell extracts of the three bacteria at 4 and 30°C; (A) GroEL, DnaK, and GroES in Shewanella oneidensis; (B) GroEL, DnaK, and DnaJ in Shewanella frigidimarina; and (C) GroEL and DnaK Psychrobacter frigidicola.
The chaperones GroEL, DnaK, and DnaJ were immunodetected in samples from S. frigidimarina cultured at 30°C (10°C above their optimal growth temperature). Nevertheless, the immunodetection of chaperones in cultures at 4°C was not possible (Figure 2B).
In this case, the chaperone system at 30°C was formed by the chaperone DnaK from the Hsp70 family and its co-chaperonin DnaJ from the Hsp40 family. No levels of the co-chaperonin GroES were detected, despite the strong detection of its partner GroEL.
The immunodetection in samples from cultures of the psychrophile P. frigidicola showed detectable levels of GroEL and DnaK chaperones when cells were grown at 30°C but not at 4°C (Figure 2C). The optimal growth temperature for this microorganism is 15°C. It seems that it needs these chaperones to cope with warm temperatures, while, at cold temperatures, the levels of these proteins remained undetectable by WB.
Identification of Over- and Under-Synthesized Proteins at Warm and Cold Temperatures
Two-dimensional fluorescence difference gel electrophoresis experiments were performed to compare the whole proteomes of each bacterial species cultured at 4 and 30°C. In that way, the molecular machinery used in the adaptation to changes in temperature could be studied.
Two-Dimensional Differential Electrophoresis Analysis of Shewanella oneidensis at 4 and 30°C
2DE-labeled images (Supplementary Figure 2) were matched to analyze the results of 2D-DIGE as explained above. The fluorescence intensity of 395 protein spots was significantly altered by an averaged fold change of ±2 (t-test, p < 0.01). The 51 spots, which showed the greatest change, were selected and excised from the gel (Supplementary Figure 3), digested with trypsin, and identified by MALDI-TOF or MALDI-TOF-TOF MS. Finally, from the 51 spots selected, 44 were identified corresponding to 47 proteins, while 7 spots could not be identified (Table 5).
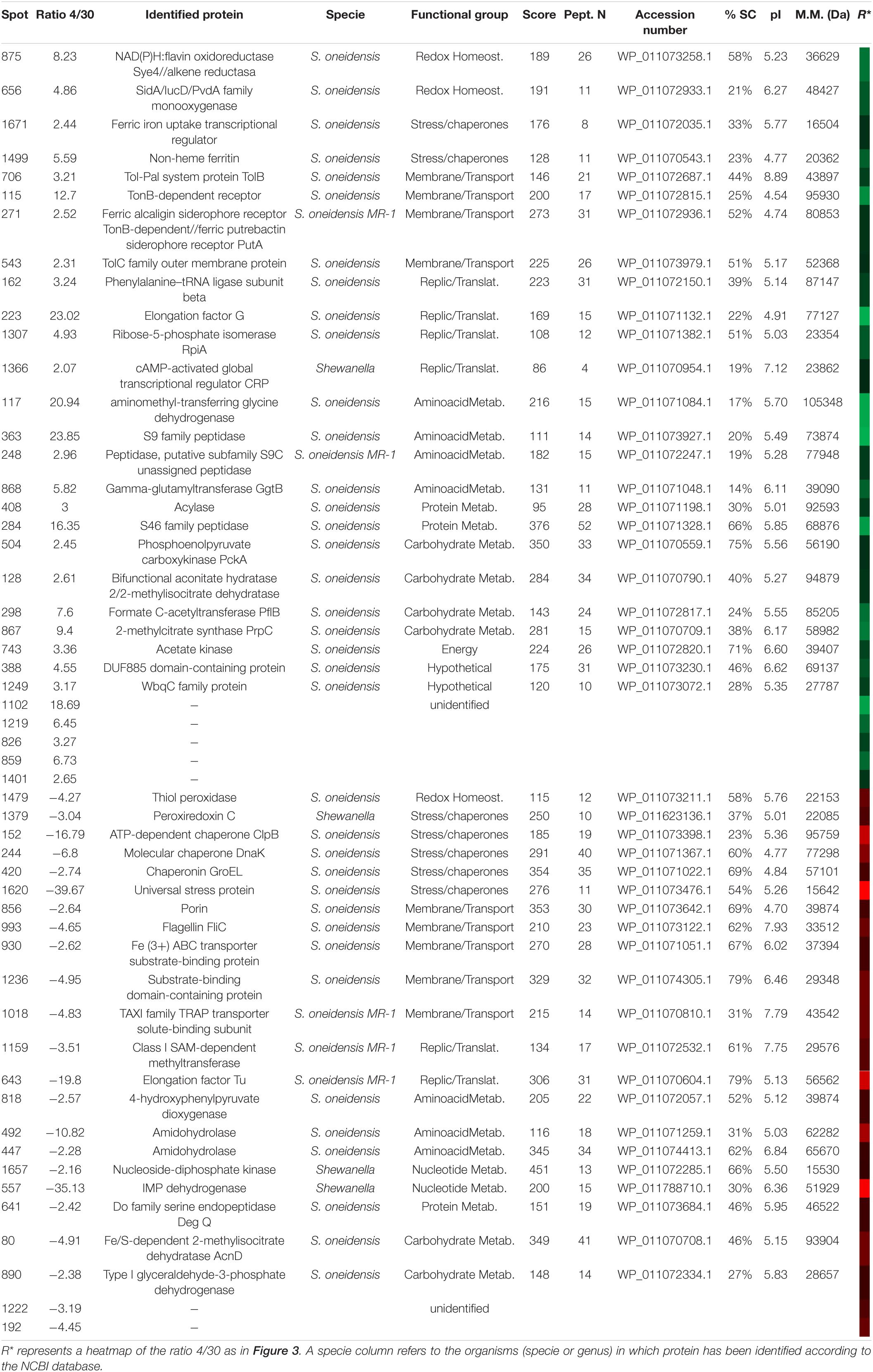
Table 5. Identified proteins in S. oneidensis overexpressed at 4 (green) and 30°C (red).
The identified proteins were classified into functional groups: carbohydrate metabolism, amino acid metabolism, nucleotide metabolism, protein metabolism, energy, redox homeostasis, transport and transmembrane proteins, chaperones and stress response, replication and translation, hypothetical, and an additional group one of the non-identified spots.
The main increased proteins observed at 30°C were especially from the categories related to stress response (GroEL, ClpB protein, DnaK), transport, and membrane proteins [flagellin, iron (III) ABC transporter, substrate-binding domain-containing protein], TAXI family TRAP transporter solute-binding subunit, and outer membrane porine. In cultures, at 4°C, S. oneidensis increased the synthesis of groups involved in replication and translation processes and in the biosynthesis of proteins and aminoacids precursors (Phe-tRNA, elongation factor G (EF-G), RpiA, and the cAMP-CRP). These bacteria also showed an increase in redox enzymes involved in the response to oxidative stress like Flav-Sye4.
Two-Dimensional Differential Electrophoresis Analysis of Shewanella frigidimarina at 4 and 30°C
After matching the fluorescence images (Supplementary Figure 4) and normalizing data from conditions of 4 and 30°C, 128 protein spots showed significant differences based on changes in the abundance ratio of ±1.5-fold with a t-test of p < 0.005. The 43 with the greatest differences were selected and cut off for their identification. About 42 spots were identified (Supplementary Figure 5) corresponding to 31 proteins – some of them with several isoforms – and 5 without satisfactory identification (Table 6).
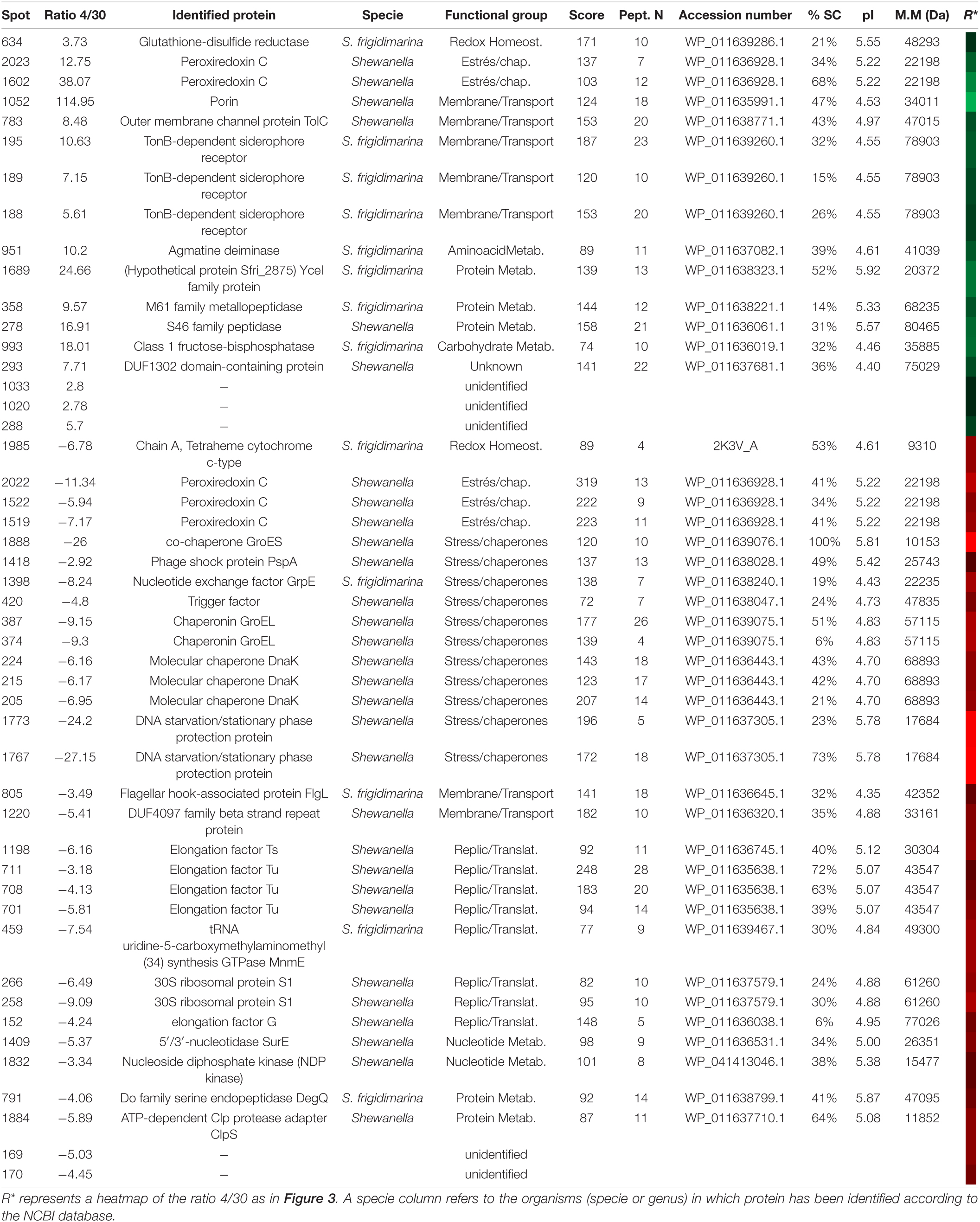
Table 6. Identified proteins in S. frigidimarina overexpressed at 4 (green) and 30°C (red).
Samples at 30°C showed a greater number of proteins identified as over-synthesized. The functional groups more represented were chaperones and stress proteins (GroES, Trigger factor, PspA, Clps adaptor, GrpE, GroEL, and DnaK) as well as the replication and translation proteins (elongation factors and ribosomal proteins). However, these two groups were not over-synthesized in samples at 4°C. At this temperature, the main groups identified were those of transport and membrane proteins. It is noteworthy that, in the 30°C samples, the redox metabolism was represented mainly by the appearance of three isoforms of the peroxiredoxin C protein. Two other isoforms of the same protein also appeared in cultures at 4°C.
Unlike the other two tested bacteria, cultures of S. frigidimarina at both 4°C and 30°C show a high number of isoforms of different proteins [TonB-dependent receptor, peroxiredoxin C, GroEL, DnaK, ferritin, elongation factor Tu (EF-Tu) and RpS1].
Two-Dimensional Differential Electrophoresis Analysis of Psychrobacter frigidicola at 4 and 30°C
The merged fluorescence images were analyzed (Supplementary Figure 6) and normalized. The data from the extracts at 4 and 30°C determined the protein spots of greatest interest. Thus, based on changes in the abundance ratio of ±2.5 times fold between both temperature conditions and with a p < 0.05 as statistically significant, 86 spots were selected. From them, the 39 spots with the greatest differences were cut out (Supplementary Figure 7), resulting in 36 (Table 7) proteins identified and 5 with no satisfactory identification. Due to the recent sequence project of this species, the identification of spots was carried out using databases of the closest phylogenetic species, such as Psychrobacter cryohalolentis, Psychrobacter arcticus, or the genus Psychrobacter in other cases.
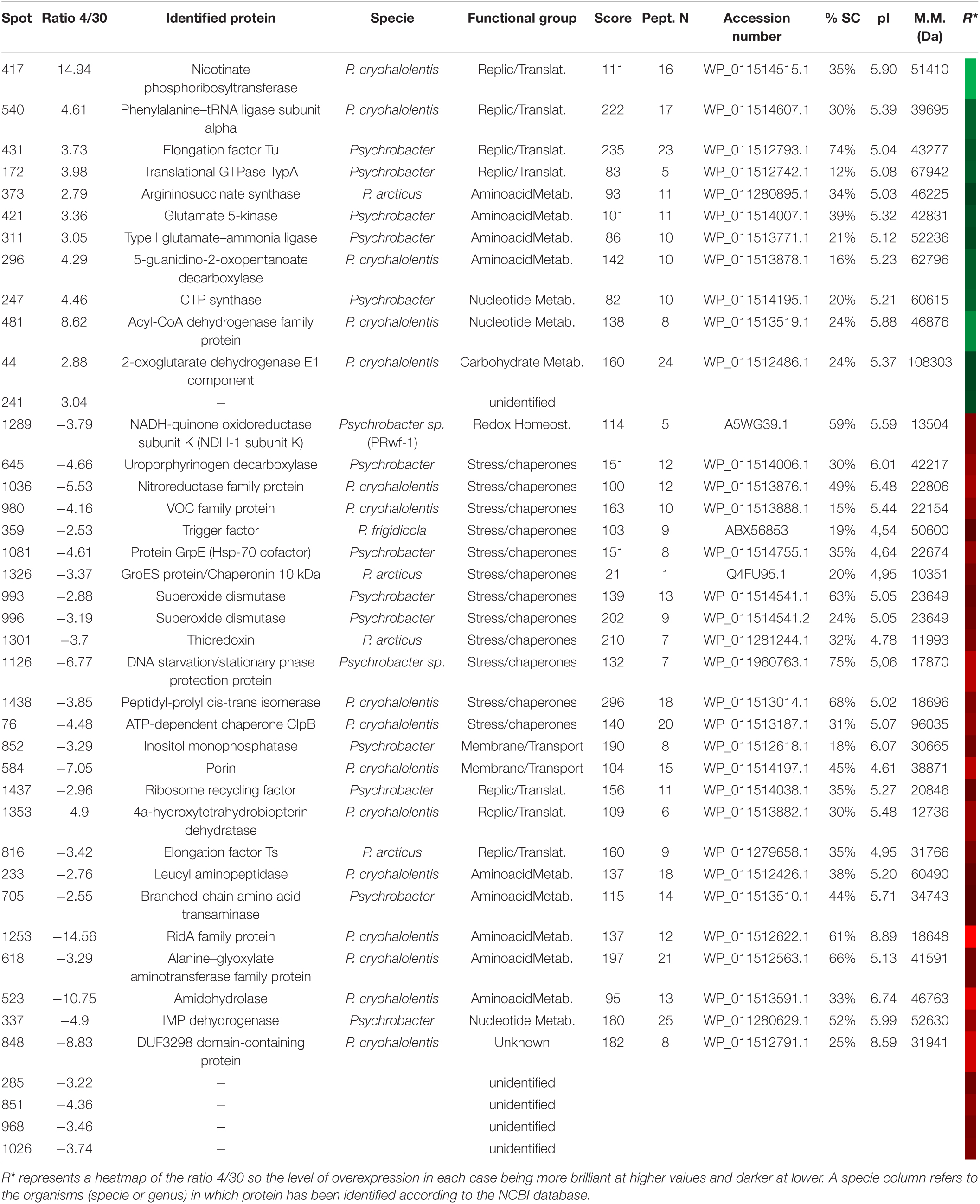
Table 7. Identified proteins in P. frigidicola overexpressed at 4 (green) and 30°C (red).
In this species, we identified more than double the number of proteins over-synthesized at 30°C with regard to 4°C. Similarly to S. frigidimarina, the main group represented at 30°C, that was not found at 4°C, corresponded to chaperones and stress proteins [trigger factor, GrpE, GroES, superoxide dismutase (SOD), thioredoxin, ferritin, peptidyl-prolyl isomerase, and ATPase AAA-2]. Besides, we found, at 30°C, several proteins were involved in redox metabolism that did not appear in the 4°C condition. The main groups over-synthesized in cultures at 4°C were those of transport and transmembrane proteins, like TolC (spot 783), three isoforms of TonB (188, 189, and 195), and porin (1052).
Proteins in Common Revealed by Two-Dimensional Fluorescence Difference Gel Electrophoresis
Those proteins identified in at least two of the three species were grouped by a functional category, comparing their synthesis through changes in the 4/30 ratio and represented in a heatmap (Figure 3).
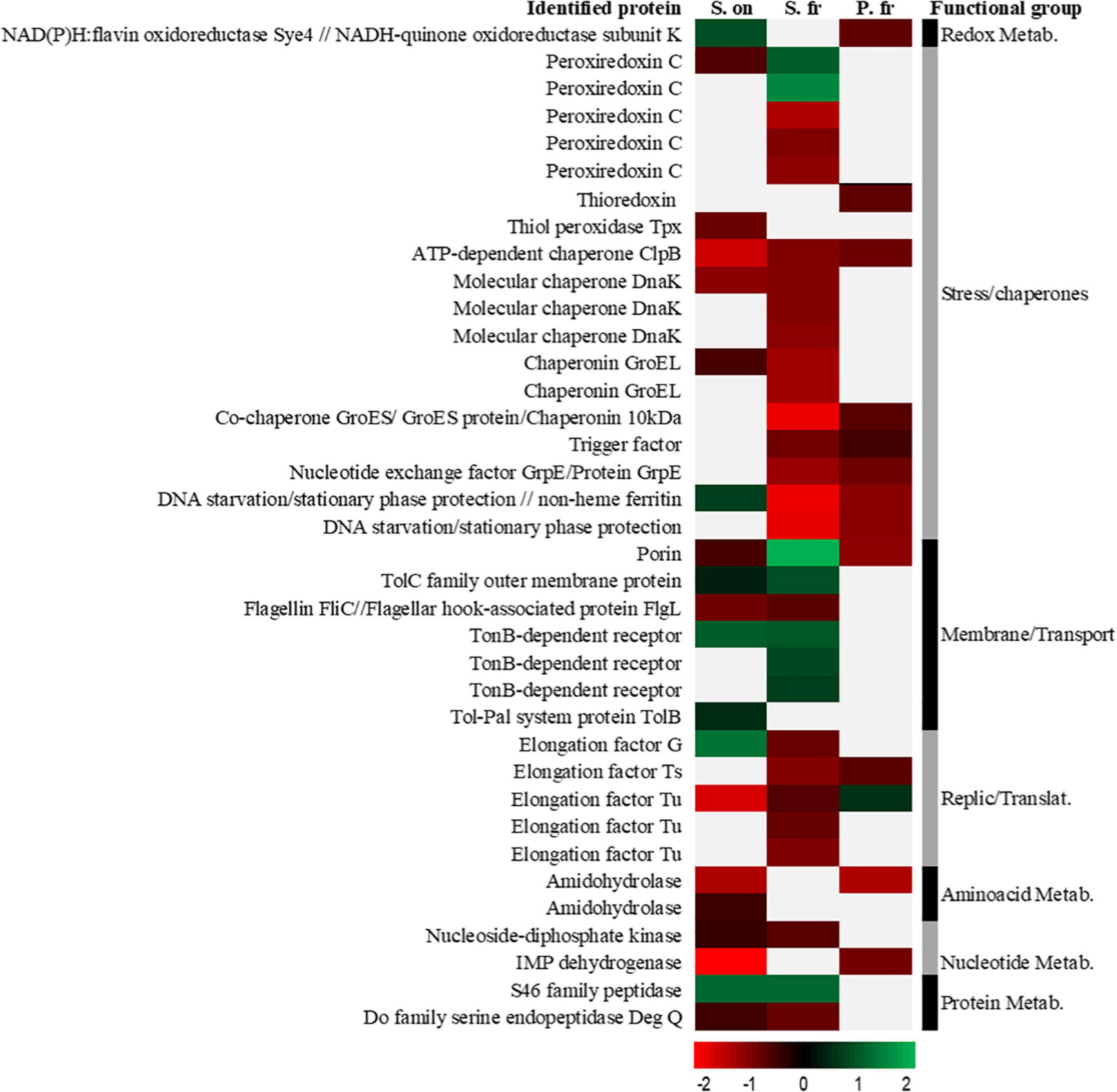
Figure 3. A heat map of ratios from 2D-DIGE experiments of the proteins over-synthesized at 30 (red) and 4°C (green) found in common in at least two of the three studied bacteria. The color and intensity of each protein are calculated as a function of the logarithm based on 10 of the value of its 4/30 ratio obtained in the 2D-DIGE experiment so that values closer to –2 indicate higher ratios in proteins whose synthesis has been increased to 30°C and closer to 2 higher ratios in proteins whose synthesis has been increased to 4°C. The gray/white color indicates the absence of identification of that specific protein in the corresponding species. “S. on” indicates Shewanella oneidensis, “S. fr” indicates Shewanella frigidimarina and “P. fr” indicates Psychrobacter frigidicola.
The protein groups most shared between the three bacterial species were those related to thermal stress, oxidative stress, and transmembrane transport. Mainly, the heatmap showed more coincidences in the production of proteins regarding temperature between S. frigidimarina and the other two species than between the other two species.
Comparison of Molecular Machinery
To compare the proteins in vivo linked to GroEL and DnaK chaperones, immunoprecipitations were carried out with specific anti-GroEL and anti-DnaK antibodies. The immunoprecipitates were analyzed by 2DE, and proteins were identified by mass spectrometry (MS). The extended list of proteins identified that co-immunoprecipitate with DnaK or GroEL in each species and at each temperature are summarized in Supplementary Material (Supplementary Tables 1–4). Figure 4 represents a comparison of the molecular machinery expressed by each bacterial species at each temperature tested that interacts with chaperones GroEL or DnaK.
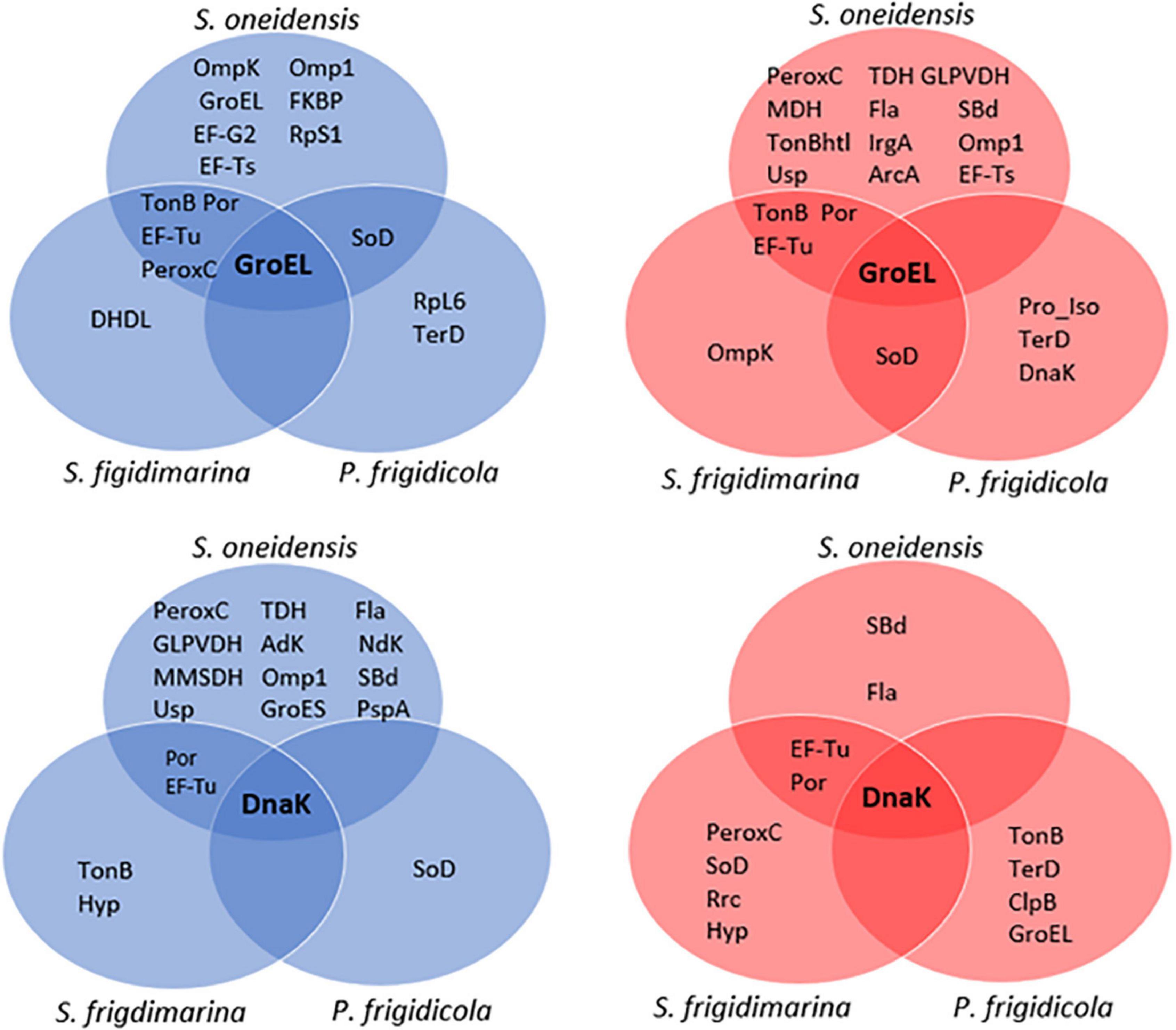
Figure 4. Venn diagrams of identified proteins that coimmunoprecipitate with GroEL (top) and DnaK (down) in the three bacteria at 4 (blue) and 30°C (red).
Regarding the proteomic machinery, P. frigidicola and S. oneidensis used different proteins in these interactions. At warm temperatures, they only had one common interaction (GroEL with SOD). On the other hand, the machinery with more proteins in common was found between the two shewanellas at both temperatures and was related to both DnaK and GroEL. These proteins were elongation factors and transmembrane proteins, and the peroxiredoxin C in the case of immunoprecipitation with GroEL in cultures at 4°C. Specifically, S. oneidensis displayed a higher number of proteins interacting with chaperone GroEL, at 4 and 30°C, and DnaK, at 4°C, than S. frigidimarina and P. frigidicola. In contrast, the immunoprecipitation with DnaK in samples at 30°C showed a slight increase in the number of proteins identified in the species S. frigidimarina and P. frigidicola, compared to S. oneidensis. In fact, P. frigidicola immuno precipitated more proteins at 30°C with antibodies from both chaperones used than at 4°C, while S. frigidimarina showed a similar number of proteins interacting with both chaperone antibodies at 4 and 30°C, with the exception of a slight increase in the case of DnaK at 30°C.
Discussion
The aim of this study was to understand the molecular strategies of three bacteria to cope with temperature changes in their environment. Given that the proteins are the main actors in these strategies and are responsible for the active machinery of the cells (D’Amico et al., 2006), the investigation has mainly focused on them. As mentioned above, this was achieved by comparing the behavior of psychrotolerant bacteria (i) Shewanella frigidimarina, which share a great part of the genome with the mesophile (ii) Shewanella oneidensis and came from the same environment (Antarctica) as the third bacteria (iii) Psychrobacter frigidicola from a different genus.
Growth Dynamics Under Different Temperatures
On the whole, the behavior of the three bacteria studied regarding their growth and biomass multiplication under cold, mild, and warm temperatures appeared relatively similar, with only mild differences in their growth phases (Table 3) and growth rates (Table 4). These differences could inform whether the bacteria are in a normal or stressful situation, taking into account the length of the lag phase. However, this would be just a potential finding taken as sole data (Hamill et al., 2020) that would need to be supported with further analysis by proteomic experiments.
Shewanella oneidensis is considered a mesophile according to its optimal growth temperature (Venkateswaran et al., 1999), but it was originally isolated from Oneida Lake in New York, which is frozen or at temperatures near 0°C for almost half of the year. This could be the reason why it can multiply at 4°C after a long lag phase with a relatively similar growth rate than at 30°C. Besides, this bacterium can behave as a psychrotolerant after a preadaptation period (Hébraud and Potier, 1999) as it has also been described for E. coli (Jones et al., 1987) and other mesophiles isolated from the Arctic and Antarctic permafrost (Steven et al., 2006).
S. frigidimarina and P. frigidicola, whose preinocula were set at 20°C, and S. oneidensis (preinocula at 30°C) showed marked lag phases in their growth at 4 and 12°C. In general, cold-adapted bacteria possess regulating factors that keep the translating machinery active at low temperatures. When their growth pauses due to a shift in temperature, they can activate these regulating mechanisms after an adaptation period and grow again at these new low temperatures (Hébraud and Potier, 1999). For S. frigidimarina, this accommodation period is shorter at 12°C than at 4°C, probably due to the small temperature difference from its optimal growth temperature, although the growth rates are different, being higher at 4°C. This may indicate that the strategies activated during the lag period provide efficient molecular machinery for successful bacterial development. The ACAM 304 strain of the psychrophile P. frigidicola, which has an optimal growth temperature of 15°C, was isolated from ornithogenic soil in Antarctica (Bowman et al., 1996). Its behavior at 4 and 12°C, with relatively long lag phases, could indicate that it has a generalized decrease in affinity at low temperatures (Nedwell and Rutter, 1994). This bacterium is described as a strict psychrophile from an environment where it is competing with psychrotolerants, which are predominant in polar areas (Nedwell and Rutter, 1994) and are better adapted to temperature changes and fluctuations (Baross and Morita, 1978).
In this study, we have observed that the growth of the three species at warm temperatures (30°C), with almost no lag period, implies a fast entry in the stationary phase, especially for the psychro-species. Nevertheless, and contrary to what one might expect, the psychrotolerant and psychrophilic bacteria showed an effective growth at temperatures slightly higher than those of their described limits. The reason could be the well-known effect of the adaptation of laboratory strains. This can expand cellular limits of medium/environmental conditions (Knöppel et al., 2018), susceptibility against chemicals (Herruzo-Cabrera et al., 2004), or even temperature, such as Saccharomyces cerevisiae, which can increase their maximum growth temperature by 3°C (Caspeta and Nielsen, 2015). Furthermore, the growth of the three bacteria at low temperatures reached higher values of O.D., probably due to the improved efficiency in the use of substrates during the first period of incubation, which allows psychrophiles to reach maximum amounts of biomass (Morita, 1975). Microbial growth is the result of a sequence of interrelated chemical reactions (Hébraud and Potier, 1999), and is not only influenced by temperature but also by the availability and quantity of nutrients (Russell, 1990), especially important in the late exponential phase and the entry into the stationary phase.
Changes in Synthesis Levels of Chaperone Systems GroEL/GroES and DnaK/DnaJ With the Temperature
Chaperones GroEL and DnaK have a crucial role in the adaptation to thermal stress through refolding of denatured or misfolded proteins, avoiding the formation of damaging aggregates, or eliminating them by proteolysis (Gragerov et al., 1992; Phadtare, 2004; Robin et al., 2009). On many occasions, they act together in successive steps being the pair DnaK/DnaJ, which first recognizes the substrate protein and stabilizes an intermediate conformation. Subsequently, the pair GroEL/GroES acts by ATP hydrolysis to conform to the native state of the protein (Langer et al., 1992).
The immunodetection in samples at 30°C showed high levels of different chaperones in each strain (GroEL/GroES and DnaK in S. oneidensis, GroEL, DnaK/DnaJ in S. frigidimarina and GroEL and DnaK in P. frigidicola), suggesting that this warm temperature condition represents a stressful situation that promotes the increase of Hsps levels (Lüders et al., 2009; Che et al., 2013). As indicated before, the natural environment of S. oneidensis is a lake that remains frozen or at temperatures near 0°C for 6 months of the year. This bacterium, although described as a mesophile, is able to behave as a psychrotolerant, showing similarities with S. frigidimarina in responses to temperature conditions. It is also known that mesophiles and psychrotolerants sometimes overlap with their thermal properties (Russell, 1990). The signal of GroES immunodetected in S. oneidensis growing at 4°C was relatively strong, which correlates with the fact that this chaperone is greatly produced at low temperatures in some species of the Shewanella genus (Miyake et al., 2007), although, in S. frigidimarina, this chaperone was not detected. S. frigidimarina and P. frigidicola, which are both originally from Antarctica, had undetectable levels of the chaperones tested in cultures at 4°C, suggesting that, even far from their respective optimal growth temperatures (20°C for S. frigidimarina and 15°C for P. frigidicola), they do not require the activity of these Hsps to cope with a cold environment such as at 4°C.
Over-Synthesis of Proteins at 4 and 30°C in the Three Bacteria
Over-Synthesized Proteins at Low Temperature
Cultures of the psycrotrophic S. frigidimarina at 4°C produce mainly a group of proteins related to the low iron availability or its metabolism, membrane transport, and oxidative stress. At low temperatures, the solubility of gases, especially oxygen, increases, and free radicals are more stable. Psychrophiles possess several mechanisms to cope with the oxidative stress of these reactive oxygen species (ROS). These mechanisms include the synthesis of specific reductases to repair oxidized residues, reduction in the number of oxidizable amino acids in the proteins, and the deletion of metabolic pathways that produce ROS (Piette et al., 2010). The increase of proteins like peroxiredoxin C and GDHr, which are involved in the redox homeostasis of the cell by acting as a reducing disulfide (Aslund and Beckwith, 1999), is presumably triggered to restore reduced cell conditions occurring at low temperatures. Membrane proteins identified in these cultures, system TolC-TonB, conform a membranal system to bond, transport, and chelate iron into the cell (Krewulak and Vogel, 2011), thus regulating iron homeostasis. Since cold affects the membrane fluidity (Bajerski et al., 2017; Siliakus et al., 2017), it could also be prompting these proteins for a better regulation of iron homeostasis to be used as a cofactor for the function of other proteins and respiration and to avoid more ROS emergence (Yang et al., 2008).
Similar to S. frigidimarina, when S. oneidensis grew at low temperature, there was an increase in the synthesis of redox enzymes and iron metabolism-related, as well, to an increase in proteins of the transport system, consisting of Ton-B-dependent receptor protein (Spot 115), the Tol-Pal system protein TolB (Spot 706), and TolC (Spot 543). This may be an indicator of an oxidative stress situation at the cold that S. oneidensis and S. frigidimarina face due to the increase of proteins that regulate iron levels and its transport for cell respiration, and accurate homeostasis to avoid oxidative poisoning (Mirus et al., 2009).
Different groups of proteins involved in replication and translation processes and in the biosynthesis of other proteins and aminoacid precursors were also over-accumulated in S. oneidensis at 4°C. Among them, the one with a notably higher ratio (23.02) was EF-G, which plays a key role in the elongation of polypeptides during the synthesis of protein (Savelsbergh et al., 2009; Li et al., 2011). In E. coli, EF-G has been described as susceptible to oxidative stress (Nagano et al., 2012) and plays a chaperone role in protein folding (Caldas et al., 2000). Furthermore, its marked synthesis increase at 4°C in S. oneidensis could be because it behaves like a cold-adapted enzyme, with high molecular flexibility (Ruggiero et al., 2007) and a low activation energy requirement in GTP hydrolysis (Thomas and Cavicchioli, 2000), which is able to keep active at low temperatures.
At 4°C, this bacterium also increased the synthesis of proteins related to intracellular levels of acetyl-CoA. Bacteria often cope with changes in their environment by activating metabolic pathways to produce available nutrients. The phosphorylation of some regulating proteins in response to stress involves mechanisms that include acetyl phosphate, whose gene is induced by a shortage of phosphate in some bacteria (Summers et al., 1999). Temperature and the growth phase are factors that influence intracellular concentrations of acetyl phosphate (Prüss and Wolfe, 1994). Thus, the over-synthesis of enzymes that control intracellular levels of acetyl-CoA, like the acetate kinase, detected in S. oneidensis at 4°C, could respond to the need of the bacterium to generate energy and phosphorylate other proteins to be active in response to a stressful situation.
In this line, there was a remarkable increase in the production of the enzyme acyl-CoA dehydrogenase (ACDH) (Spot 481), also in P. fridigicola at 4°C that could support the activation of the metabolism of this bacterium at cold temperatures. An active metabolism contributes to the generation of energy and electron transfer in respiration with the participation of acetyl-CoA, a product of the reaction catalyzed by ACDH in metabolic pathways such as the tricarboxylic acid cycle. This enzyme belongs to the family of proteins involved in lipid oxidation and in the metabolism of amino acids and acts as an oxidoreductase for the transfer of electrons in the respiratory chain (Kim and Miura, 2004). Possibly, this active metabolism in P. frigidicola cultures at low temperature also requires a high synthesis and presence of NAD co-factors of the main metabolic processes (Gerdes et al., 2006). Interestingly, we did find strong induction of the enzyme nicotine-phosphoribosyl transferase (spot. 417) too, which catalyzes the first reaction in the synthesis of NAD (Dulyaninova et al., 2000).
The active metabolism of this bacterium at 4°C was also depicted by the over-synthesis of proteins involved in the biosynthesis processes of numerous amino acids and nucleotides, as well as in the metabolism of carbohydrates, while there was no induction of chaperons or stress proteins detected by 2D-DIGE. One of the most strongly induced proteins in these samples was the CTP synthase. This enzyme is responsible for the production of CTP from UTP and glutamine, and for the regulation of intracellular levels of CTP (Endrizzi et al., 2004), which is an essential precursor of membrane phospholipids (Chang and Carman, 2008). It also plays a key role in the metabolism of pyrimidines and is very important for the growth rate and the concentration of ribonucleotides and deoxyribonucleotides (Jørgensen et al., 2004).
Over-Synthesized Proteins at High Temperature
When S. frigidimarina grew at warm temperatures (30°C), an increase in the synthesis of several chaperones and stress response proteins was observed. They were mainly proteins that face oxidative, osmotic, and thermal stress situations. Some of them appeared in different isoforms like DnaK (posts 224, 215, 205) and GroEL (Spots 387 and 374) and others as single forms like GrpE (Spot 1398), TF (Spot 420), PspA (Spot 1418), and GroES (Spot 1888). The latter, although undetectable by WB in this study, forms the GroEL-GroES complex that keeps bound in the presence of ATP (Ventura et al., 2004) in the thermal stress response. These chaperones play an important role in the quality control system of post-translational processing. They control the folding and the proper conformation of other proteins to cope with environmental changes such as temperature, cooperating with each other (Liberek et al., 2008; Muga and Moro, 2008; Robin et al., 2009). In response to oxidative stress at warm temperatures, we found in S. frigidimarina two isoforms of the DNA-binding ferritin-like proteins region, DNA-Dps (spots 1773 and 1767), which belongs to Dps family proteins. This family is involved in protecting DNA from damage by chelating iron and by ferroxidase activity (Calhoun and Kwon, 2011). DNA-Dps is also required for starvation response and long-term stationary viability (Jeong et al., 2006) that is why we cannot discard some contribution to its high levels detected to the fact that the samples were collected once cultures reached the stationary phase. Nevertheless, another three isoforms of other protein-related to oxidative stress were also overexpressed in these samples, the peroxiredoxin C (Spots 2022, 1522, 1519). It is part of the large and highly conserved family of the peroxiredoxines, which detoxify peroxides by reducing them (Rhee, 2016). This protein is also known as alkyl hydroperoxide reductase (AhpC) described as a protective enzyme against peroxides in E. coli (Storz and Imlay, 1999) against osmotic stress in Staphylococcus aureus and induced by a temperature increase in Bacillus subtilis, while, in Shewanella putrefaciens, its activity is related to cold acclimation (Leblanc et al., 2003).
This molecular strategy involving proteins against redox, osmotic, and stationary phase stress during warm conditions seems to be common in the two shewanellas studied, being the peroxiredoxin C also identified as over-accumulated in warm cultures of S. oneidensis (Spot 1379). In this bacterium, at 30°C, there were too many other over-synthesized proteins related to oxidative and osmotic stress belonging to the anti-oxidant AhpC-TSA or peroxiredoxin family. That is the case of the Tpx-C (Spot 1479) that protects against oxidative (Storz and Imlay, 1999; Horst et al., 2010; Somprasong et al., 2012), osmotic and cold stress (Leblanc et al., 2003), and that increase in the late stationary phase (Miyake et al., 2007). Thus, the bacteria S. oneidensis, as well as S. frigidimarina, could share a similar mechanism to that of Bacillus subtilis in which this type of enzyme is induced as a response to temperature increase and osmotic stress after the beginning of the stationary phase (Hecker and Völker, 1990; Antelmann et al., 1996).
Regarding other groups of proteins over-synthesized in S. oneidensis at 30°C, we found GroEL (Spot 420), which increases the solubilization of aggregates (Niwa et al., 2012), the ClpB protein (Spot 152), and the DnaK (Spot 244), correlating the immunodetection results observed by WB. Protein GroEL belongs to the Hsp100/Clp AAA + ATPase family and cooperates with the chaperone system DnaK/DnaJ/GrpE in efficient solubilization and reactivation of aberrant protein aggregates formed as a consequence of cellular heat stress (Kedzierska et al., 2003; Mogk et al., 2003; Lee et al., 2004; Doyle et al., 2007). This chaperone restores the activity of other membrane proteins that also appeared over-synthesized in S. oneidensis at 30°C, the porin (spot 856), which forms hydrophilic channels in the membrane and is refolded by GroEL (Goulhen et al., 2004). Similar to S. frigidimarina, several other stress proteins over-accumulated could be indicative of a stress situation for S. oneidensis when grown at 30°C and/or due to be in the stationary phase. This is the case of universal stress protein (Usp) (Spot 1620). It is a small cytoplasmic protein that is greatly induced in many bacteria, such as E. coli, as a response to a wide range of environmental conditions and stressful situations like heat shock, starvation, or in the presence of stressors that prevent cell growth, and agents that damage DNA (Tkaczuk et al., 2013). This protein increases the survival rate of cells during prolonged exposure to these stressful conditions and is phosphorylated when cells enter the stationary growth phase (Sousa and McKay, 2001).
Again, the majority of proteins over-synthesized at 30°C in the case of the psychrophile P. frigidicola belong to the chaperone and stress protein groups. There were several of them involved in redox metabolism, resistance to oxidative stress or in heat stress, and the starvation response as it was observed for the other two bacteria studied. They were oxide and nitro-reductase enzymes that contribute to the defense against oxidative stress by reducing the redox cycle and the toxic nitro-aromatic compounds and quinones (Liochev et al., 1999), as well as degrading ROS and free radicals.
In this regard, two isoforms of the enzyme SOD (Spots 993 and 996), the protein thiorredoxin (Spot 1301) and DNA-Dps (Spot 1126), were induced in P. frigidicola at warm temperatures. The first protein plays a pivotal role in maintaining the redox state of the cell and protects it from oxidative stress (Wang et al., 2013; Lu and Holmgren, 2014). DNA-Dps, identified also in S. frigidimarina, is highly conserved and operates, as indicated before, against several kinds of stressors (Jeong et al., 2006; Karas et al., 2015) in E. coli, including oxidative damage to protect DNA. and it is also required in the normal starvation response and in long-term stationary viability (Almirón et al., 1992; Nair and Finkel, 2004).
Concerning the specific chaperones group, the main proteins found were involved in the heat stress response forming, as was the case for samples of S. frigidimarina, a cooperative role in the folding and maintenance of the proper conformation of other proteins to cope with environmental changes such as temperature.
Comparative Molecular Machinery in Each Bacterium Against Temperature Conditions
Both temperature and oxidative stress are closely related, since temperature influences the solubility of oxygen and the formation of ROS (Piette et al., 2010; Chattopadhyay et al., 2011). Stress proteins were required at warm temperatures, whereas cold temperature seems to promote a similar membrane system in the two shewanellas studied.
The analysis of 2D-DIGE revealed common strategies in several groups of proteins between the three species, depending on growth temperature. The species S. frigidimarina shared over/under-synthesized proteins with S. oneidensis and P. frigidicola, while these two showed fewer proteins in common with one another. The strategy followed by S. frigidimarina, especially at warm temperatures, used shame chaperones and stress proteins with both P. frigidicola (GroES, TF, GrpE, ferritin, stationary phase protection protein) and S. oneidensis (ClpB, GroEL, and DnaK). On the other hand, the mesophile, S. oneidensis, and the psychrophile, P. frigidicola, employed different strategies at 30°C, except for the ClpB, which works together with DnaK to solubilize and reactivate aggregated proteins (Doyle et al., 2007; Doyle et al., 2015). This interaction is specie specific (Miot et al., 2011) and plays an essential role in cell survival under thermal stress conditions (Nagy et al., 2010; Krajewska et al., 2017).
Regarding oxidative stress, members of the superfamily of peroxiredoxins (peroxiredoxin C, thiorredoxin, and thiol peroxidase) were over-synthesized at 30°C in the two species of Shewanella and Psychrobacter. This kind of protein removes peroxides that are toxic to the cell (Storz and Imlay, 1999) and does also protects against osmotic stress and temperature changes in several bacteria (Leblanc et al., 2003). Even though oxidative stress is more typical at low temperatures (Chattopadhyay et al., 2011), it could also occur at 30°C as a response to the by-products of the increased metabolism of these bacteria at moderate to warm temperatures (Piette et al., 2010). Interestingly, the psychrotolerant S. frigidimarina may need to cope with oxidative stress at low temperatures through over-synthetization of other peroxiredoxin C isoforms. S. oneidensis showed a differential behavior in the synthesis of proteins related to the iron homeostasis. While it over-synthesized the non-heme ferritin at 4°C; the other two bacteria increased the levels of the Dps, which, as mentioned before, joins ferritin and protects against oxidative damage (Nair and Finkel, 2004; De Martino et al., 2016).
The metabolism of S. oneidensis and S. frigidimarina at low temperature, below their optimum, uses channels and siderophores that regulate the entrance of ions in the cell. These species at 4°C there induced synthesis of proteins of the outer membrane channels like TolPal System (TonB, TolC, and TolB) and the porin protein (that appears also in P. frigidicola), which interacts with this system for assembling to the membrane (Rigal et al., 1997). The system has an important role in the elimination of toxic compounds from oxidative damage through the membrane (Benz et al., 1993; Fralick, 1996) and its reparation (Zgurskaya et al., 2011) and binds to iron-chelating or siderophore complexes (Mirus et al., 2009). The porin protein, which was present in the three bacteria, could be acting differently in the psychrotolerant than in the other two species. It was over-synthesized at 4°C in S. frigidimarina, probably forming aqueous channels and interacting with the TolPal System (Cascales et al., 2002). However, it could be related to the basal apparatus of the flagellum (Chu et al., 2020; Liao et al., 2021) in S. oneidensis, in which it is over-synthesized at 30°C along with the flagellin protein.
Concerning replication and translation processes, there was an induction of different elongation factors like EF-G, EF-Tu, and EF-Ts, mainly at warm temperatures, except for factor EF-G in the mesophile and EF-Tu in the psychrophile. The EF-Ts is a guanine nucleotide exchanger for EF-Tu by GTPase activity for the separation of the EF-Tu-GDP complex from the ribosome (Palmer et al., 2013). It has a high similarity in different mesophiles such as E. coli, thermophiles such as Thermus thermophilus, and psychrophiles such as Pseudoalteromonas haloplanktis, but it was observed in higher concentration in the latter (Raimo et al., 2004). EF-G has different associations with ribosomes in response to stressful situations (deLivron et al., 2009). This factor is required for growth at low temperatures, as well as in acidic environments in Sinorhizobium meliloti (Kiss et al., 2004) and E. coli (Pfennig and Flower, 2001). It has also been described in other extremophiles such as Psychrobacter cryohalolentis and Thiomicrospiracrunogena, increasing the translation efficiency during the growth at low temperatures (Krishnan and Flower, 2008). Interestingly, EF-Tu that catalyzes the binding of aminoacyl-RNA to the ribosome can also act as a chaperone in the renaturation of other proteins (Masullo et al., 2000). This chaperone role has also been proposed for EF-Ts (Takeshita and Tomita, 2010) in the assembly and maintenance of the Qβ viral RNA polymerase in infected cells.
Regarding the groups of amino acids, nucleic acids, and proteins metabolism, there were no proteins identified that were shared by the three bacteria. Contrary to what has been indicated before for groups of stress and membrane proteins, no coincidences were found for these groups between S. frigidimarina and P. frigidicola, but both bacteria presented similarities with S. oneidensis. This could indicate that the main processes affected by stress and essential for cell survival and adaptation are those involving chaperone activity, detoxifying proteins, and transport through the membrane.
Several protein isoforms of different functional groups (peroxiredoxin C, DnaK, GroEL, EF-Tu, TonB-dependent receptor) present in S. frigidimarina were not observed in S. oneidensis or P. frigidicola. Other psychrotolerant species from Antarctica belonging to the genera Sphingobacterium and Pseudomonas, which can grow in the temperature range of 0–30°C, display differences in the phosphorylation-dephosphorylation state of membrane and cytosolic proteins in response to temperature changes, acting as cell heat sensors (Ray et al., 1994). The presence of isoforms in S. frigidimarina suggested that this psychrotolerant bacterium could have a similar mechanism to recognize environmental conditions and adapt to them, probably by different phosphorylation states or other post-translational modifications (dos Santos et al., 2010; García-Descalzo et al., 2014).
As a further step in understanding the molecular machinery that these three types of bacteria use to adapt to temperature changes, proteins that co-immunoprecipitate alongside the main chaperones DnaK and GroEL were investigated. Analysis of the results revealed protein interactions that could be happening in vivo in the cell as part of an adaptive response to different growth temperatures or as part of their normal metabolism at these temperatures (Figure 4).
Chaperones are essential components of the quality control machinery in cells. They optimize the physiological responses of bacteria by interacting directly with other proteins, taking part in cell signaling, and acting as stress response proteins (Hartl and Hayer-Hartl, 2002). Chaperones GroEL and DnaK are the main representatives of their families (Cooper and Hausman, 2000; Beckerman, 2005; Liberek et al., 2008), and their action has been described in a wide range of bacteria, frequently working together (Fourie and Wilson, 2020; Fatima et al., 2021; Mayer, 2021) and taking part in the regulation of transcription and translation (Parsot et al., 2003).
The molecular machinery involving these chaperones according to temperature was more complex in S. oneidensis, of intermediate complexity in S. frigidimarina, and notably simpler in P. frigidicola. While, in S. oneidensis, the proteins that co-immunoprecipitate with GroEL and DnaK tended to cluster at both warm and cold temperatures; those of P. frigidicola gathered mainly toward warmer temperatures, and S. frigidimarina showed a more equal distribution in the established and combined chaperone-temperature conditions. The groups of proteins that co-immunoprecipitated with the chaperones tested in S. oneidensis and S. frigidimarina shared more elements with each other than they do with proteins co-immunoprecipitated in P. frigidicola. These were proteins particularly involved in membrane transport, elongation processes, and oxidative stress. Characteristically, psychrotrophs are a more diverse group than psychrophiles, and their thermal properties overlap with those of some mesophiles (Russell, 1990). For S. oneidensis, a noticeably higher number of proteins seem to act together with GroEL at both temperatures and with DnaK at 30°C.
In general, the protein interaction network observed was mainly composed of chaperones and co-chaperones (GroES, Usp), membrane proteins involved in the transport of molecules and ions (porins and Ton-B-dependent receptors), and the formation of flagellar structures (flagellin, Omp family); but also proteins related with the defense against oxidative stress (peroxiredoxine, superoxide dismutase) and the elongation factors EF-Tu, EF-Ts, and EF-G. Co-immunoprecipitation revealed that the main processes to cope with temperature changes in these bacteria were those involved in the stress (temperature and oxidative) response. And that they need, especially S. oneidensis, the support of a membrane system that helps to eliminate toxic products resulting in these processes.
It is noteworthy that P. frigidicola, with a simpler proteome and a smaller number of proteins that co-immunoprecipitate with GroEL and Dnak than the two shewanellas, showed the co-appearance of the tellurium ion resistance (TerD) and SOD proteins with GroEL at both temperatures and with DnaK at 30°C for the former and DnaK at 4°C for the latter.
The TerD protein is part of a poorly characterized family involved especially in the response to oxidative stress in several organisms (Chasteen et al., 2009; Anantharaman et al., 2012; Daigle et al., 2015) and is also present in different pathogenic bacteria (Taylor, 1999; Turkovicova et al., 2016; Chen et al., 2019). The presence of this enzyme and SOD suggests that this psychrophilic bacterium is adapted to the frequent situation of oxidative stress at low temperatures due to the stability of ROS, and the metabolic processes that can lead to the generation of ROS at high temperatures (Piette et al., 2011).
Conclusion
In the adaptation of microorganisms to changes in temperature, proteins are the molecular machine that regulates metabolism and cell functions. This study combines proteomic approaches with classical microbiology analysis, like growth dynamics.
We have observed a similar growing behavior between the three species against cold and warm temperatures, being able to achieve akin biomass under the temperatures tested, regardless of whether it is a stress situation or not. Some differences observed in the first stages of growth could give clues about whether the bacteria are in a normal or stressful situation, considering the duration of the lag phase. This observation had to be corroborated by a more detailed analysis, for which proteomic tools are essential.
The analysis of proteomic data showed that, in general, the two bacteria from the genus Shewanella displayed a more versatile adaptive response than the Psychrobacter species tested. These findings could suggest a tendency of an adaptation and evolution from mesophilic to psychrophilic microorganisms, maintaining a similar genome, but this would need to be supported with more genomic studies. Within the two Shewanella species studied, the psychrotolerant S. frigidimarina exhibits temperature-adaptative mechanisms in common with the other two species. This could be due to the genome (common with the mesophile) and the post-translational modifications, which give rise to isoforms (common with the psychrophile). As a result, it is better adapted to thermal variations in the temperature range 4–30°C.
In the molecular machinery to withstand temperature stressful situations, the role of chaperones is essential. In the three species studied, these proteins interact with other proteins forming complexes to provide diverse physiological responses of the cell to different growth temperatures. Equally important for adapting to temperature changes is the establishment of a cooperative system of chaperones with other proteins for the protection against oxidative damage, transmembrane transport, and elongation factors. They, together, are in charge of ensuring the viability and survival of bacteria.
This study is potentially related to pieces of climate change research since microorganisms and their adaptive strategies could be biosensors of the state of environments. An in-depth understanding of how they cope with changes in the temperature conditions could give us information about the molecules overproduced and provide a tool for evaluating the state of the environment. Additionally, it could also contribute to the Astrobiology field. The knowledge about the limits of life and how it adapts to environmental changes is crucial in the study of the habitability of environments with extreme conditions on Earth and in other planetary bodies like Mars or icy moons. The molecules employed to adapt to these extreme conditions could be considered biomarkers of life on Earth and beyond.
Data Availability Statement
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE repository with the dataset identifier PXD030438.
Author Contributions
CC contributed to conceptualization, funding acquisition, editing, and manuscript review. LG-D contributed to experiments performance, analysis, and manuscript writing. EG-L contributed to experiments performance and data acquisition. All authors contributed to the article and approved the submitted version.
Funding
This research has been funded by Grant Nos. PID2019-104205GB-C22 and MDM-2017-0737 Unidad de Excelencia “Maria de Maeztu”-Centro de Astrobiología (INTA-CSIC) by the Spanish Ministry of Science and Innovation/State Agency of Research MCIN/AEI/10.13039/501100011033. EG-L is supported by PTA2016-12325-I grant provided by MCIN/AEI/10.13039/501100011033. LG-D was supported by the Instituto Nacional de Técnica Aeroespacial through the grant FPI “Rafael Calvo Rodés.” This research is part of POLARCSIC activities.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Paula Alcázar for her assistance in reviewing, and Proteomic Unit of the Parque Científico de Madrid and Proteomic Service at Centro Nacional de Investigaciones Cardiovasculares for 2D-DIGE experiments and identification of proteins.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.841359/full#supplementary-material
Abbreviations
SBd, substrate-binding domain-containing protein; TAXI-TRAP, TAXI family TRAP transporter solute-binding subunit; Omp, outer membrane porine; Phe-tRNA, phenylalanile-tRNA ligase subunit beta; RpiA, ribose-5-phosphate isomerase; cAMP-CRP, cAMP-activated global transcriptional regulator CRP protein; Flav-Sye4-NAD(P)H, flavin oxidoreductase Sye4; RpS1, ribosomal protein S1 30S; Pro-Iso, peptidyl-prolyl isomerase; TolC, outer membrane channel protein TolC; TonB, TonB-dependent siderophore receptor; GDHr, glutathione-disulfide reductase protein; PflB, formate C-acetyltransferase; PrpC, protein 2-methylcitrate synthase; Fur, ferric iron uptake transcriptional regulator; TolB, Tol-Pal system protein TolB; DH-E1, 2-oxoglutarate dehydrogenase E1 component; acyl-CoA DH, acyl-CoA dehydrogenase family protein; NpRibT, nicotinate phosphoribosyltransferase; CTP synthase, cytosine triphosphate synthase; PspA, phage shock protein A; ClpS, protease-dependent ATP adaptor protein Clp; Do-DegQ, periplasmic Do family serine endopeptidase DegQ; DNA-Dps, DNA starvation/stationary phase protection protein; Cyto-c-type, Chain A Tetraheme cytochrome c-type; Tpx-C, Thiol peroxidase protein subunit C; Q-nitroreductase -K, K subunit of the NADH quinone nitroreductase family protein.
Footnotes
References
Ahmad, A., Bhattacharya, A., McDonald, R. A., Cordes, M., Ellington, B., Bertelsen, E. B., et al. (2011). Heat shock protein 70 kDa chaperone/DnaJ cochaperone complex employs an unusual dynamic interface. Proc. Natl. Acad. Sci. 108, 18966–18971. doi: 10.1073/pnas.1111220108
Alcazar, A., Garcia-Descalzo, L., and Cid, C. (2010). “Microbial evolution and adaptation in icy worlds,” in Astrobiology: Physical Origin, Biological Evolution and Spatial Distribution, eds S. Hegedus and J. Csonka (New York, NY: Nova Science Publishers), 81–95. doi: 10.3389/fmicb.2017.01407
Almirón, M., Link, A. J., Furlong, D., and Kolter, R. (1992). A novel DNA-binding protein with regulatory and protective roles in starved Escherichia coli. Genes Dev. 6, 2646–2654. doi: 10.1101/gad.6.12b.2646
Anantharaman, V., Iyer, L. M., and Aravind, L. (2012). Ter-dependent stress response systems: novel pathways related to metal sensing, production of a nucleoside-like metabolite, and DNA-processing. Mol. Biosyst. 8, 3142–3165. doi: 10.1039/c2mb25239b
Antelmann, H., Engelmann, S., Schmid, R., and Hecker, M. (1996). General and oxidative stress responses in Bacillus subtilis: cloning, expression, and mutation of the alkyl hydroperoxide reductase operon. J. Bacteriol. 178, 6571–6578. doi: 10.1128/jb.178.22.6571-6578.1996
Aslund, F., and Beckwith, J. (1999). Bridge over troubled waters: sensing stress by disulfide bond formation. Cell 96, 751–753. doi: 10.1016/s0092-8674(00)80584-x
Bajerski, F., Wagner, D., and Mangelsdorf, K. (2017). Cell membrane fatty acid composition of Chryseobacterium frigidisoli PB4T, isolated from antarctic glacier forefield soils, in response to changing temperature and pH Conditions. Front. Microbiol. 8:677. doi: 10.3389/fmicb.2017.00677
Baross, J., and Morita, R. (1978). “Microbial life at low temperatures: ecological aspects,” in Microbial life in Extreme Environments, ed. D. J. Kushner (London: Academic Press), 9–71.
Beckerman, M. (2005). “Protein folding and binding,” in Molecular and Cellular Signaling, ed. T. Abl (New York, NY: Springer), 89–110.
Benz, R., Maier, E., and Gentschev, I. (1993). TolC of Escherichia coli functions as an outer membrane channel. Zentralbl Bakteriol. 278, 187–196. doi: 10.1016/s0934-8840(11)80836-4
Bowman, J. P., Cavanagh, J., Austin, J. J., and Sanderson, K. (1996). Novel Psychrobacter species from Antarctic ornithogenic soils. Int. J. Syst. Bacteriol. 46, 841–848. doi: 10.1099/00207713-46-4-841
Bromfield, E. G., and Nixon, B. (2013). The function of chaperone proteins in the assemblage of protein complexes involved in gamete adhesion and fusion processes. Reproduction 145, R31–R42. doi: 10.1530/rep-12-0316
Caldas, T., Laalami, S., and Richarme, G. (2000). Chaperone properties of bacterial elongation factor EF-G and initiation factor IF2. J. Biol. Chem. 275, 855–860. doi: 10.1074/jbc.275.2.855
Calhoun, L. N., and Kwon, Y. M. (2011). Structure, function and regulation of the DNA-binding protein Dps and its role in acid and oxidative stress resistance in Escherichia coli: a review. J. Appl. Microbiol. 110, 375–386. doi: 10.1111/j.1365-2672.2010.04890.x
Camberg, J. L., Doyle, S. M., Johnston, D. M., and Wickner, S. (2013). “Molecular Chaperones,” in Brenner’s Encyclopedia of Genetics, 2nd Edn, eds S. Maloy and K. Hughes (San Diego, CA: Academic Press), 456–460.
Cascales, E., Bernadac, A., Gavioli, M., Lazzaroni, J. C., and Lloubes, R. (2002). Pal lipoprotein of Escherichia coli plays a major role in outer membrane integrity. J. Bacteriol. 184, 754–759. doi: 10.1128/jb.184.3.754-759.2002
Caspeta, L., and Nielsen, J. (2015). Thermotolerant yeast strains adapted by laboratory evolution show trade-off at ancestral temperatures and preadaptation to other stresses. mBio 6:e00431. doi: 10.1128/mBio.00431-15
Cavicchioli, R., Siddiqui, K. S., Andrews, D., and Sowers, K. R. (2002). Low-temperature extremophiles and their applications. Curr. Opin. Biotechnol. 13, 253–261. doi: 10.1016/s0958-1669(02)00317-8
Chang, Y. F., and Carman, G. M. (2008). CTP synthetase and its role in phospholipid synthesis in the yeast Saccharomyces cerevisiae. Prog. Lipid Res. 47, 333–339. doi: 10.1016/j.plipres.2008.03.004
Chasteen, T. G., Fuentes, D. E., Tantaleán, J. C., and Vásquez, C. C. (2009). Tellurite: history, oxidative stress, and molecular mechanisms of resistance. FEMS Microbiol. Rev. 33, 820–832. doi: 10.1111/j.1574-6976.2009.00177.x
Chattopadhyay, M. K., Raghu, G., Sharma, Y. V., Biju, A. R., Rajasekharan, M. V., and Shivaji, S. (2011). Increase in oxidative stress at low temperature in an antarctic bacterium. Curr. Microbiol. 62, 544–546. doi: 10.1007/s00284-010-9742-y
Che, S., Song, W., and Lin, X. (2013). Response of heat-shock protein (HSP) genes to temperature and salinity stress in the antarctic psychrotrophic bacterium Psychrobacter sp. G. Curr. Microbiol. 67, 601–608. doi: 10.1007/s00284-013-0409-3
Chen, J., Tan, W., Wang, W., Hou, S., Chen, G., Xia, L., et al. (2019). Identification of common antigens of three pathogenic Nocardia species and development of DNA vaccine against fish nocardiosis. Fish Shellfish Immunol. 95, 357–367. doi: 10.1016/j.fsi.2019.09.038
Chu, J., Liu, J., and Hoover, T. R. (2020). Phylogenetic distribution, ultrastructure, and function of bacterial flagellar sheaths. Biomolecules 10:363. doi: 10.3390/biom10030363
Cid, C., Garcia-Descalzo, L., Casado-Lafuente, V., Amils, R., and Aguilera, A. (2010). Proteomic analysis of the response of an acidophilic strain of Chlamydomonas sp. (Chlorophyta) to natural metal-rich water. Proteomics 10, 2026–2036. doi: 10.1002/pmic.200900592
Cloutier, P., and Coulombe, B. (2013). Regulation of molecular chaperones through post-translational modifications: decrypting the chaperone code. Biochim. Biophys. Acta 1829, 443–454. doi: 10.1016/j.bbagrm.2013.02.010
Cooper, G. M., and Hausman, R. E. (2000). “Protein folding and processing,” in The Cell: A Molecular Approach, 2nd Edn (Sunderland, MA: Sinauer Associates; Oxford University Press).
Cray, J. A., Stevenson, A., Ball, P., Bankar, S. B., Eleutherio, E. C. A., Ezeji, T. C., et al. (2015). Chaotropicity: a key factor in product tolerance of biofuel-producing microorganisms. Curr. Opin. Biotechnol. 33, 228–259. doi: 10.1016/j.copbio.2015.02.010
Daigle, F., Lerat, S., Bucca, G., Sanssouci, É, Smith, C. P., Malouin, F., et al. (2015). A terD domain-encoding gene (SCO2368) is involved in calcium homeostasis and participates in calcium regulation of a DosR-like regulon in Streptomyces coelicolor. J. Bacteriol. 197, 913–923. doi: 10.1128/jb.02278-14
D’Amico, S., Collins, T., Marx, J.-C., Feller, G., Gerday, C., and Gerday, C. (2006). Psychrophilic microorganisms: challenges for life. EMBO Rep. 7, 385–389. doi: 10.1038/sj.embor.7400662
De Maayer, P., Anderson, D., Cary, C., and Cowan, D. A. (2014). Some like it cold: understanding the survival strategies of psychrophiles. EMBO Rep. 15, 508–517. doi: 10.1002/embr.201338170
De Martino, M., Ershov, D., van den Berg, P. J., Tans, S. J., and Meyer, A. S. (2016). Single-cell analysis of the dps response to oxidative stress. J. Bacteriol. 198, 1662–1674. doi: 10.1128/jb.00239-16
deLivron, M. A., Makanji, H. S., Lane, M. C., and Robinson, V. L. (2009). A novel domain in translational GTPase BipA mediates interaction with the 70S ribosome and influences GTP hydrolysis. Biochemistry 48, 10533–10541. doi: 10.1021/bi901026z
dos Santos, M. F., Muniz de Pádua, V. L., de Matos Nogueira, E., Hemerly, A. S., and Domont, G. B. (2010). Proteome of Gluconacetobacter diazotrophicus co-cultivated with sugarcane plantlets. J. Proteomics 73, 917–931. doi: 10.1016/j.jprot.2009.12.005
Doyle, S. M., Hoskins, J. R., and Wickner, S. (2007). Collaboration between the ClpB AAA+ remodeling protein and the DnaK chaperone system. Proc. Natl. Acad. Sci. U. S. A. 104, 11138–11144. doi: 10.1073/pnas.0703980104
Doyle, S. M., Shastry, S., Kravats, A. N., Shih, Y. H., Miot, M., Hoskins, J. R., et al. (2015). Interplay between E. coli DnaK, ClpB and GrpE during protein disaggregation. J. Mol. Biol. 427, 312–327. doi: 10.1016/j.jmb.2014.10.013
Dulyaninova, N. G., Podlepa, E. M., Toulokhonova, L. V., and Bykhovsky, V. Y. (2000). Salvage pathway for NAD biosynthesis in Brevibacterium ammoniagenes: regulatory properties of triphosphate-dependent nicotinate phosphoribosyltransferase. Biochim. Biophys. Acta 1478, 211–220. doi: 10.1016/s0167-4838(00)00045-5
Endrizzi, J. A., Kim, H., Anderson, P. M., and Baldwin, E. P. (2004). Crystal structure of Escherichia coli cytidine triphosphate synthetase, a nucleotide-regulated glutamine amidotransferase/ATP-dependent amidoligase fusion protein and homologue of anticancer and antiparasitic drug targets. Biochemistry 43, 6447–6463. doi: 10.1021/bi0496945
Fatima, K., Naqvi, F., and Younas, H. (2021). A review: molecular chaperone-mediated folding, unfolding and disaggregation of expressed recombinant proteins. Cell Biochem. Biophys. 79, 153–174. doi: 10.1007/s12013-021-00970-5
Fourie, K. R., and Wilson, H. L. (2020). Understanding GroEL and DnaK stress response proteins as antigens for bacterial diseases. Vaccines 8:773. doi: 10.3390/vaccines8040773
Fralick, J. A. (1996). Evidence that TolC is required for functioning of the Mar/AcrAB efflux pump of Escherichia coli. J. Bacteriol. 178, 5803–5805. doi: 10.1128/jb.178.19.5803-5805.1996
García-Descalzo, L., Alcazar, A., Baquero, F., and Cid, C. (2011). Identification of in vivo HSP90-interacting proteins reveals modularity of HSP90 complexes is dependent on the environment in psychrophilic bacteria. Cell Stress Chaperones 16, 203–218. doi: 10.1007/s12192-010-0233-7
García-Descalzo, L., García-López, E., Alcázar, A., Baquero, F., and Cid, C. (2014). Proteomic analysis of the adaptation to warming in the Antarctic bacteria Shewanella frigidimarina. Biochim. Biophys. Acta 1844, 2229–2240. doi: 10.1016/j.bbapap.2014.08.006
Gerdes, S. Y., Kurnasov, O. V., Shatalin, K., Polanuyer, B., Sloutsky, R., Vonstein, V., et al. (2006). Comparative genomics of NAD biosynthesis in cyanobacteria. J. Bacteriol. 188, 3012–3023. doi: 10.1128/jb.188.8.3012-3023.2006
Goulhen, F., Dé, E., Pagès, J. M., and Bolla, J. M. (2004). Functional refolding of the Campylobacter jejuni MOMP (major outer membrane protein) porin by GroEL from the same species. Biochem. J. 378(Pt 3), 851–856. doi: 10.1042/bj20031239
Gragerov, A., Nudler, E., Komissarova, N., Gaitanaris, G. A., Gottesman, M. E., and Nikiforov, V. (1992). Cooperation of GroEL/GroES and DnaK/DnaJ heat shock proteins in preventing protein misfolding in Escherichia coli. Proc. Natl. Acad. Sci. U. S. A. 89, 10341–10344. doi: 10.1073/pnas.89.21.10341
Hamill, P. G., Stevenson, A., McMullan, P. E., Williams, J. P., Lewis, A. D. R., Sudharsan, S., et al. (2020). Microbial lag phase can be indicative of, or independent from, cellular stress. Sci. Rep. 10:5948. doi: 10.1038/s41598-020-62552-4
Hartl, F. U., and Hayer-Hartl, M. (2002). Molecular chaperones in the cytosol: from nascent chain to folded protein. Science 295, 1852–1858. doi: 10.1126/science.1068408
Hébraud, M., and Potier, P. (1999). Cold shock response and low temperature adaptation in psychrotrophic bacteria. J. Mol. Microbiol. Biotechnol. 1, 211–219.
Hecker, M., and Völker, U. (1990). General stress proteins in Bacillus subtilis. FEMS Microbiol. Lett. 74, 197–213. doi: 10.1111/j.1574-6968.1990.tb04065.x
Herruzo-Cabrera, R., Vizcaino-Alcaide, M. J., and Fernández-Aceñero, M. J. (2004). The influence of laboratory adaptation on test strains, such as Pseudomonas aeruginosa, in the evaluation of the antimicrobial efficacy of ortho-phthalaldehyde. J. Hosp. Infect. 57, 217–222. doi: 10.1016/j.jhin.2004.01.031
Hochachka, P. W., and Somero, G. N. (2002). Biochemical Adaptation: Mechanism and Process in Physiological Evolution. Oxford: Oxford University Press.
Horst, S. A., Jaeger, T., Denkel, L. A., Rouf, S. F., Rhen, M., and Bange, F. C. (2010). Thiol peroxidase protects Salmonella enterica from hydrogen peroxide stress in vitro and facilitates intracellular growth. J. Bacteriol. 192, 2929–2932. doi: 10.1128/jb.01652-09
Irwin, J. A., and Baird, A. W. (2004). Extremophiles and their application to veterinary medicine. Ir. Vet. J. 57, 348–354. doi: 10.1186/2046-0481-57-6-348
Jeong, K. C., Baumler, D. J., and Kaspar, C. W. (2006). dps expression in Escherichia coli O157:H7 requires an extended −10 region and is affected by the cAMP receptor protein. Biochim. Biophys. Acta 1759, 51–59. doi: 10.1016/j.bbaexp.2006.02.001
Jones, P. G., VanBogelen, R. A., and Neidhardt, F. C. (1987). Induction of proteins in response to low temperature in Escherichia coli. J. Bacteriol. 169, 2092–2095. doi: 10.1128/jb.169.5.2092-2095.1987
Jørgensen, C. M., Hammer, K., Jensen, P. R., and Martinussen, J. (2004). Expression of the pyrG gene determines the pool sizes of CTP and dCTP in Lactococcus lactis. Eur. J. Biochem. 271, 2438–2445. doi: 10.1111/j.1432-1033.2004.04168.x
Karas, V. O., Westerlaken, I., and Meyer, A. S. (2015). The DNA-binding protein from starved cells (Dps) utilizes dual functions to defend cells against multiple stresses. J. Bacteriol. 197, 3206–3215. doi: 10.1128/jb.00475-15
Kedzierska, S., Akoev, V., Barnett, M. E., and Zolkiewski, M. (2003). Structure and function of the middle domain of ClpB from Escherichia coli. Biochemistry 42, 14242–14248. doi: 10.1021/bi035573d
Kim, J. J., and Miura, R. (2004). Acyl-CoA dehydrogenases and acyl-CoA oxidases. Structural basis for mechanistic similarities and differences. Eur. J. Biochem. 271, 483–493. doi: 10.1046/j.1432-1033.2003.03948.x
Kiss, E., Huguet, T., Poinsot, V., and Batut, J. (2004). The typA gene is required for stress adaptation as well as for symbiosis of Sinorhizobium meliloti 1021 with certain Medicago truncatula lines. Mol. Plant Microbe Interact. 17, 235–244. doi: 10.1094/mpmi.2004.17.3.235
Knöppel, A., Knopp, M., Albrecht, L. M., Lundin, E., Lustig, U., Näsvall, J., et al. (2018). Genetic adaptation to growth under laboratory conditions in Escherichia coli and Salmonella enterica. Front. Microbiol. 9:756. doi: 10.3389/fmicb.2018.00756
Krajewska, J., Modrak-Wójcik, A., Arent, Z. J., Więckowski, D., Zolkiewski, M., Bzowska, A., et al. (2017). Characterization of the molecular chaperone ClpB from the pathogenic spirochaete Leptospira interrogans. PLoS One 12:e0181118. doi: 10.1371/journal.pone.0181118
Krewulak, K. D., and Vogel, H. J. (2011). TonB or not TonB: is that the question? Biochem. Cell Biol. 89, 87–97. doi: 10.1139/o10-141
Krishnan, K., and Flower, A. M. (2008). Suppression of DeltabipA phenotypes in Escherichia coli by abolishment of pseudouridylation at specific sites on the 23S rRNA. J. Bacteriol. 190, 7675–7683. doi: 10.1128/jb.00835-08
Langer, T., Lu, C., Echols, H., Flanagan, J., Hayer, M. K., and Hartl, F. U. (1992). Successive action of DnaK, DnaJ and GroEL along the pathway of chaperone-mediated protein folding. Nature 356, 683–689. doi: 10.1038/356683a0
Leblanc, L., Leboeuf, C., Leroi, F., Hartke, A., and Auffray, Y. (2003). Comparison between NaCl tolerance response and acclimation to cold temperature in Shewanella putrefaciens. Curr. Microbiol. 46, 157–162. doi: 10.1007/s00284-002-3837-z
Lee, S., Sowa, M. E., Choi, J. M., and Tsai, F. T. (2004). The ClpB/Hsp104 molecular chaperone-a protein disaggregating machine. J. Struct. Biol. 146, 99–105. doi: 10.1016/j.jsb.2003.11.016
Li, W., Trabuco, L. G., Schulten, K., and Frank, J. (2011). Molecular dynamics of EF-G during translocation. Proteins 79, 1478–1486. doi: 10.1002/prot.22976
Liao, C. H., Chang, C. L., Huang, H. H., Lin, Y. T., Li, L. H., and Yang, T. C. (2021). Interplay between OmpA and RpoN regulates flagellar synthesis in Stenotrophomonas maltophilia. Microorganisms 9:1216. doi: 10.3390/microorganisms9061216
Liberek, K., Lewandowska, A., and Zietkiewicz, S. (2008). Chaperones in control of protein disaggregation. EMBO J. 27, 328–335. doi: 10.1038/sj.emboj.7601970
Lin, Y. S. (2008). Using a strategy based on the concept of convergent evolution to identify residue substitutions responsible for thermal adaptation. Proteins 73, 53–62. doi: 10.1002/prot.22049
Liochev, S. I., Hausladen, A., and Fridovich, I. (1999). Nitroreductase A is regulated as a member of the soxRS regulon of Escherichia coli. Proc. Natl. Acad. Sci. U. S. A. 96, 3537–3539. doi: 10.1073/pnas.96.7.3537
Lu, J., and Holmgren, A. (2014). The thioredoxin antioxidant system. Free Radic. Biol. Med. 66, 75–87. doi: 10.1016/j.freeradbiomed.2013.07.036
Lüders, S., Fallet, C., and Franco-Lara, E. (2009). Proteome analysis of the Escherichia coli heat shock response under steady-state conditions. Proteome Sci. 7:36. doi: 10.1186/1477-5956-7-36
Maier, T., Schmidt, A., Güell, M., Kühner, S., Gavin, A. C., Aebersold, R., et al. (2011). Quantification of mRNA and protein and integration with protein turnover in a bacterium. Mol. Syst. Biol. 7:511. doi: 10.1038/msb.2011.38
Makhnevych, T., and Houry, W. A. (2013). The control of spindle length by Hsp70 and Hsp110 molecular chaperones. FEBS Lett. 587, 1067–1072. doi: 10.1016/j.febslet.2013.02.018
Masullo, M., Arcari, P., de Paola, B., Parmeggiani, A., and Bocchini, V. (2000). Psychrophilic elongation factor Tu from the antarctic Moraxella sp. Tac II 25: biochemical characterization and cloning of the encoding gene. Biochemistry 39, 15531–15539. doi: 10.1021/bi0018133
Mayer, M. P. (2021). The Hsp70-chaperone machines in bacteria. Front. Mol. Biosci. 8:694012. doi: 10.3389/fmolb.2021.694012
Metpally, R. P., and Reddy, B. V. (2009). Comparative proteome analysis of psychrophilic versus mesophilic bacterial species: insights into the molecular basis of cold adaptation of proteins. BMC Genomics 10:11. doi: 10.1186/1471-2164-10-11
Miller, I., Crawford, J., and Gianazza, E. (2006). Protein stains for proteomic applications: which, when, why? Proteomics 6, 5385–5408. doi: 10.1002/pmic.200600323
Miot, M., Reidy, M., Doyle, S. M., Hoskins, J. R., Johnston, D. M., Genest, O., et al. (2011). Species-specific collaboration of heat shock proteins (Hsp) 70 and 100 in thermotolerance and protein disaggregation. Proc. Natl. Acad. Sci. U. S. A. 108, 6915–6920. doi: 10.1073/pnas.1102828108
Mirus, O., Strauss, S., Nicolaisen, K., von Haeseler, A., and Schleiff, E. (2009). TonB-dependent transporters and their occurrence in cyanobacteria. BMC Biol. 7:68. doi: 10.1186/1741-7007-7-68
Miyake, R., Kawamoto, J., Wei, Y. L., Kitagawa, M., Kato, I., Kurihara, T., et al. (2007). Construction of a low-temperature protein expression system using a cold-adapted bacterium. Shewanella sp. strain Ac10, as the host. Appl. Environ. Microbiol. 73, 4849–4856. doi: 10.1128/aem.00824-07
Mogk, A., Schlieker, C., Strub, C., Rist, W., Weibezahn, J., and Bukau, B. (2003). Roles of individual domains and conserved motifs of the AAA+ chaperone ClpB in oligomerization. ATP hydrolysis, and chaperone activity. J. Biol. Chem. 278, 17615–17624. doi: 10.1074/jbc.M209686200
Morita, R. Y. (1975). Psychrophilic bacteria. Bacteriol. Rev. 39, 144–167. doi: 10.1128/br.39.2.144-167.1975
Muga, A., and Moro, F. (2008). Thermal adaptation of heat shock proteins. Curr. Protein Pept. Sci. 9, 552–566. doi: 10.2174/138920308786733903
Nagano, T., Kojima, K., Hisabori, T., Hayashi, H., Morita, E. H., Kanamori, T., et al. (2012). Elongation factor G is a critical target during oxidative damage to the translation system of Escherichia coli. J. Biol. Chem. 287, 28697–28704. doi: 10.1074/jbc.M112.378067
Nagy, M., Guenther, I., Akoyev, V., Barnett, M. E., Zavodszky, M. I., Kedzierska-Mieszkowska, S., et al. (2010). Synergistic cooperation between two ClpB isoforms in aggregate reactivation. J. Mol. Biol. 396, 697–707. doi: 10.1016/j.jmb.2009.11.059
Nair, S., and Finkel, S. E. (2004). Dps protects cells against multiple stresses during stationary phase. J. Bacteriol. 186, 4192–4198. doi: 10.1128/JB.186.13.4192-4198.2004
Nedwell, D. B., and Rutter, M. (1994). Influence of temperature on growth rate and competition between two psychrotolerant Antarctic bacteria: low temperature diminishes affinity for substrate uptake. Appl. Environ. Microbiol. 60, 1984–1992. doi: 10.1128/aem.60.6.1984-1992.1994
Niwa, T., Kanamori, T., Ueda, T., and Taguchi, H. (2012). Global analysis of chaperone effects using a reconstituted cell-free translation system. Proc. Natl. Acad. Sci. U. S. A. 109, 8937–8942. doi: 10.1073/pnas.1201380109
Palmer, S. O., Rangel, E. Y., Montalvo, A. E., Tran, A. T., Ferguson, K. C., and Bullard, J. M. (2013). Cloning and characterization of EF-Tu and EF-Ts from Pseudomonas aeruginosa. Biomed. Res. Int. 2013:585748. doi: 10.1155/2013/585748
Paredes, D. I., Watters, K., Pitman, D. J., Bystroff, C., and Dordick, J. S. (2011). Comparative void-volume analysis of psychrophilic and mesophilic enzymes: structural bioinformatics of psychrophilic enzymes reveals sources of core flexibility. BMC Struct. Biol. 11:42. doi: 10.1186/1472-6807-11-42
Parsot, C., Hamiaux, C., and Page, A. L. (2003). The various and varying roles of specific chaperones in type III secretion systems. Curr. Opin. Microbiol. 6, 7–14. doi: 10.1016/s1369-5274(02)00002-4
Perrody, E., Cirinesi, A.-M., Desplats, C., Keppel, F., Schwager, F., Tranier, S., et al. (2012). A bacteriophage-encoded J-domain protein interacts with the DnaK/Hsp70 chaperone and stabilizes the heat-shock factor σ32 of Escherichia coli. PLoS Genet. 8:e1003037. doi: 10.1371/journal.pgen.1003037
Pfennig, P. L., and Flower, A. M. (2001). BipA is required for growth of Escherichia coi K12 at low temperature. Mol. Genet. Genomics 266, 313–317. doi: 10.1007/s004380100559
Phadtare, S. (2004). Recent developments in bacterial cold-shock response. Curr. Issues Mol. Biol. 6, 125–136.
Piette, F., D’Amico, S., Mazzucchelli, G., Danchin, A., Leprince, P., and Feller, G. (2011). Life in the cold: a proteomic study of cold-repressed proteins in the antarctic bacterium pseudoalteromonas haloplanktis TAC125. Appl. Environ. Microbiol. 77, 3881–3883. doi: 10.1128/aem.02757-10
Piette, F., D’Amico, S., Struvay, C., Mazzucchelli, G., Renaut, J., Tutino, M. L., et al. (2010). Proteomics of life at low temperatures: trigger factor is the primary chaperone in the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125. Mol. Microbiol. 76, 120–132. doi: 10.1111/j.1365-2958.2010.07084.x
Prüss, B. M., and Wolfe, A. J. (1994). Regulation of acetyl phosphate synthesis and degradation, and the control of flagellar expression in Escherichia coli. Mol. Microbiol. 12, 973–984. doi: 10.1111/j.1365-2958.1994.tb01085.x
Raimo, G., Lombardo, B., Masullo, M., Lamberti, A., Longo, O., and Arcari, P. (2004). Elongation factor Ts from the Antarctic eubacterium Pseudoalteromonas haloplanktis TAC 125: biochemical characterization and cloning of the encoding gene. Biochemistry 43, 14759–14766. doi: 10.1021/bi048949b
Ray, M. K., Kumar, G. S., and Shivaji, S. (1994). Phosphorylation of membrane proteins in response to temperature in an Antarctic Pseudomonas syringae. Microbiology 140(Pt 12), 3217–3223. doi: 10.1099/13500872-140-12-3217
Rigal, A., Bouveret, E., Lloubes, R., Lazdunski, C., and Benedetti, H. (1997). The TolB protein interacts with the porins of Escherichia coli. J. Bacteriol. 179, 7274–7279. doi: 10.1128/jb.179.23.7274-7279.1997
Robin, S., Togashi, D. M., Ryder, A. G., and Wall, J. G. (2009). Trigger factor from the psychrophilic bacterium Psychrobacter frigidicola is a monomeric chaperone. J. Bacteriol. 191, 1162–1168. doi: 10.1128/jb.01137-08
Rodrigues, D. F., and Tiedje, J. M. (2008). Coping with our cold planet. Appl. Environ. Microbiol. 74, 1677–1686. doi: 10.1128/AEM.02000-07
Ruggiero, I., Raimo, G., Palma, M., Arcari, P., and Masullo, M. (2007). Molecular and functional properties of the psychrophilic elongation factor G from the Antarctic Eubacterium Pseudoalteromonas haloplanktis TAC 125. Extremophiles 11, 699–709. doi: 10.1007/s00792-007-0088-8
Russell, N. J. (1990). Cold adaptation of microorganisms. Philos. Trans. R. Soc. Lond. B Biol. Sci. 326, 595–608, discussion 608-611. doi: 10.1098/rstb.1990.0034
Savelsbergh, A., Rodnina, M. V., and Wintermeyer, W. (2009). Distinct functions of elongation factor G in ribosome recycling and translocation. RNA 15, 772–780. doi: 10.1261/rna.1592509
Schumann, W. (2009). Temperature sensors of eubacteria. Adv. Appl. Microbiol. 67, 213–256. doi: 10.1016/s0065-2164(08)01007-1
Siliakus, M. F., van der Oost, J., and Kengen, S. W. M. (2017). Adaptations of archaeal and bacterial membranes to variations in temperature, pH and pressure. Extremophiles 21, 651–670. doi: 10.1007/s00792-017-0939-x
Somprasong, N., Jittawuttipoka, T., Duang-Nkern, J., Romsang, A., Chaiyen, P., Schweizer, H. P., et al. (2012). Pseudomonas aeruginosa thiol peroxidase protects against hydrogen peroxide toxicity and displays atypical patterns of gene regulation. J. Bacteriol. 194, 3904–3912. doi: 10.1128/jb.00347-12
Sousa, M. C., and McKay, D. B. (2001). Structure of the universal stress protein of Haemophilus influenzae. Structure 9, 1135–1141. doi: 10.1016/s0969-2126(01)00680-3
Steven, B., Léveillé, R., Pollard, W. H., and Whyte, L. G. (2006). Microbial ecology and biodiversity in permafrost. Extremophiles 10, 259–267. doi: 10.1007/s00792-006-0506-3
Storz, G., and Imlay, J. A. (1999). Oxidative stress. Curr. Opin. Microbiol. 2, 188–194. doi: 10.1016/s1369-5274(99)80033-2
Summers, M. L., Denton, M. C., and McDermott, T. R. (1999). Genes coding for phosphotransacetylase and acetate kinase in Sinorhizobium meliloti are in an operon that is inducible by phosphate stress and controlled by phoB. J. Bacteriol. 181, 2217–2224. doi: 10.1128/jb.181.7.2217-2224.1999
Takeshita, D., and Tomita, K. (2010). Assembly of Q{beta} viral RNA polymerase with host translational elongation factors EF-Tu and -Ts. Proc. Natl. Acad. Sci. U. S. A. 107, 15733–15738. doi: 10.1073/pnas.1006559107
Taylor, D. E. (1999). Bacterial tellurite resistance. Trends Microbiol. 7, 111–115. doi: 10.1016/s0966-842x(99)01454-7
Thomas, T., and Cavicchioli, R. (2000). Effect of temperature on stability and activity of elongation factor 2 proteins from Antarctic and thermophilic methanogens. J. Bacteriol. 182, 1328–1332. doi: 10.1128/jb.182.5.1328-1332.2000
Thompson, A. D., Bernard, S. M., Skiniotis, G., and Gestwicki, J. E. (2012). Visualization and functional analysis of the oligomeric states of Escherichia coli heat shock protein 70 (Hsp70/DnaK). Cell Stress Chaperones 17, 313–327. doi: 10.1007/s12192-011-0307-1
Tkaczuk, K. L., Shumilin, I. A., Chruszcz, M., Evdokimova, E., Savchenko, A., and Minor, W. (2013). Structural and functional insight into the universal stress protein family. Evol. Appl. 6, 434–449. doi: 10.1111/eva.12057
Tortora, G., Funke, B., and Case, C. (2010). Microbiology An Introduction, 10th Edn. San Francisco, CA: Benjamin Cummings.
Turkovicova, L., Smidak, R., Jung, G., Turna, J., Lubec, G., and Aradska, J. (2016). Proteomic analysis of the TerC interactome: novel links to tellurite resistance and pathogenicity. J. Proteomics 136, 167–173. doi: 10.1016/j.jprot.2016.01.003
Venkateswaran, K., Moser, D. P., Dollhopf, M. E., Lies, D. P., Saffarini, D. A., MacGregor, B. J., et al. (1999). Polyphasic taxonomy of the genus Shewanella and description of Shewanella oneidensis sp. nov. Int. J. Syst. Bacteriol. 49(Pt 2), 705–724. doi: 10.1099/00207713-49-2-705
Ventura, M., Canchaya, C., Zink, R., Fitzgerald, G. F., and van Sinderen, D. (2004). Characterization of the groEL and groES loci in Bifidobacterium breve UCC 2003: genetic, transcriptional, and phylogenetic analyses. Appl. Environ. Microbiol. 70, 6197–6209. doi: 10.1128/aem.70.10.6197-6209.2004
Wang, Q., Hou, Y., Qu, J., Hong, Y., Lin, Y., and Han, X. (2013). Molecular cloning, expression, purification and characterization of thioredoxin from Antarctic sea-ice bacteria Pseudoalteromonas sp. AN178. Mol. Biol. Rep. 40, 6587–6591. doi: 10.1007/s11033-013-2771-4
Yang, Y., Harris, D. P., Luo, F., Wu, L., Parsons, A. B., Palumbo, A. V., et al. (2008). Characterization of the Shewanella oneidensis Fur gene: roles in iron and acid tolerance response. BMC Genomics 9, (Suppl. 1):S11. doi: 10.1186/1471-2164-9-s1-s11
Keywords: psychrophiles, proteomics, adaptation, chaperone protein, protein complex
Citation: García-Descalzo L, García-López E and Cid C (2022) Comparative Proteomic Analysis of Psychrophilic vs. Mesophilic Bacterial Species Reveals Different Strategies to Achieve Temperature Adaptation. Front. Microbiol. 13:841359. doi: 10.3389/fmicb.2022.841359
Received: 22 December 2021; Accepted: 08 March 2022;
Published: 03 May 2022.
Edited by:
Gloria Paz Levicán, University of Santiago, ChileReviewed by:
Andrés Esteban Marcoleta, University of Chile, ChileCatherine Duport, University of Avignon, France
Copyright © 2022 García-Descalzo, García-López and Cid. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Laura García-Descalzo, garciadl@cab.inta-csic.es