 Nikola Y. Gyurchev1,2*
Nikola Y. Gyurchev1,2* Ángela Coral-Medina3,4Susan M. Weening5Salwa Almayouf1Niels G. A. Kuijpers6†
Ángela Coral-Medina3,4Susan M. Weening5Salwa Almayouf1Niels G. A. Kuijpers6† Elke Nevoigt2†Edward J. Louis1*†
Elke Nevoigt2†Edward J. Louis1*†- 1Centre of Genetic Architecture of Complex Traits, Department of Genetics and Genome Biology, University of Leicester, Leicester, United Kingdom
- 2School of Science, Jacobs University Bremen, Bremen, Germany
- 3SPO, Université de Montpellier, INRAE, Institut Agro, Montpellier, France
- 4School of Microbiology, University College Cork, Cork, Ireland
- 5Department of Biotechnology, Delft University of Technology, Delft, Netherlands
- 6Research and Development, HEINEKEN Supply Chain B.V, Zoeterwoude, Netherlands
Non-domesticated, wild Saccharomyces yeasts have promising characteristics for beer diversification, particularly when used in the generation of de novo interspecific hybrids. A major motivation for the current work was the question whether attractive novel Saccharomyces interspecific hybrids can be created for the production of exotic lager beers without using the genomic resources of the ale yeast Saccharomyces cerevisiae. Importantly, maltotriose utilization is an essential characteristic typically associated with domesticated ale/lager brewing strains. A high-throughput screening on nearly 200 strains representing all eight species of the Saccharomyces genus was conducted. Three Saccharomyces mikatae strains were able to aerobically grow on maltotriose as the sole carbon source, a trait until recently unidentified for this species. Our screening also confirmed the recently reported maltotriose utilization of the S. jurei strain D5095T. Remarkably, de novo hybrids between a maltotriose-utilizing S. mikatae or S. jurei strain and the maltotriose-negative Saccharomyces eubayanus strain CBS 12357T displayed heterosis and outperformed both parents with regard to aerobically utilizing maltotriose as the sole source of carbon. Indeed, the maximum specific growth rates on this sugar were comparable to the well-known industrial strain, Saccharomyces pastorianus CBS 1513. In lager brewing settings (oxygen-limited), the new hybrids were able to ferment maltose, while maltotriose was not metabolized. Favorable fruity esters were produced, demonstrating that the novel hybrids have the potential to add to the diversity of lager brewing.
Introduction
Brewing has been connected to human activity for millennia. The long-term unintentional domestication of baker’s yeast (mainly ale yeasts of the species Saccharomyces cerevisiae) has developed its superior fermentation capabilities and the pleasant mixture of aroma and flavor compounds in ale-style beers. Moreover, standardizing and regulating the brewing process in the medieval times to be conducted at lower temperatures (ca. 12°C) gave rise to lager-style beers with their distinguished clean, crisp, and refreshing character (Hornsey, 2003). The yeast responsible for lager fermentation was identified to be different from S. cerevisiae and named Saccharomyces pastorianus, a hybrid between an ale S. cerevisiae and the cold-tolerant Saccharomyces eubayanus. The latter parental wild yeast has only been discovered a decade ago (Libkind et al., 2011). S. eubayanus strains have been isolated from different regions of the world (Libkind et al., 2011; Bing et al., 2014; Peris et al., 2014; Gayevskiy and Goddard, 2016; Nespolo et al., 2020). Two of those isolates have also been used by the brewing industry to brew novel types of lager beer (Lovin Dublin, 2018).
Lager beer brewed with the hybrid yeast S. pastorianus makes up the majority of global beer volume; however, preferences toward novel specialty beers with exotic aroma profiles led to the exponential increase of craft breweries (Garavaglia and Swinnen, 2017; Jaeger et al., 2020). Industrial S. pastorianus strains are genetically related and therefore exhibit limited phenotypic diversity including a narrow range of flavor/aromatic profiles (Dunn and Sherlock, 2008; Okuno et al., 2015; Gallone et al., 2019; Salazar et al., 2019). Therefore, there has been a high interest in increasing aromatic complexity in lager beer by exploiting the biodiversity of wild yeasts of the Saccharomyces clade either individually or after hybridization with other species at the relative cold fermentation temperatures. Indeed, interspecific hybridization techniques have been a powerful tool in this context, particularly since hybrids could outperform both parental species due to heterosis effects as previously demonstrated for de novo S. cerevisiae × S. eubayanus crosses (Hebly et al., 2015; Krogerus et al., 2015; Mertens et al., 2015). Such a novel hybrid with enhanced brewing and aromatic properties has recently been commercialized for the production of lager beer (Lallemand Brewing, 2022).
The Saccharomyces clade currently comprises eight recognized species from various sources and locations: S. cerevisiae, S. paradoxus (Batshinskaya, 1914), S. mikatae (Naumov et al., 2000), S. eubayanus (Libkind et al., 2011), S. kudriavzevii (Naumov et al., 2000), S. uvarum (Beijerinck, 1898), S. arboricola (Wang and Bai, 2008), and the most recently identified S. jurei (Naseeb et al., 2017). Non-domesticated (wild) representatives of the genus have already shown their fermentative and desired fruity aroma potential in lager brewing with several examples recently reported for S. eubayanus (Eizaguirre et al., 2018; Lovin Dublin, 2018; Mardones et al., 2020), S. paradoxus (Nikulin et al., 2020b), and S. jurei (Giannakou et al., 2021; Hutzler et al., 2021). Such studies have been commonly restricted to a single or few tested strains.
Alongside the potential benefits for aroma complexity, there are a few typical traits in wild yeasts that are regarded undesired in lager brewing. These include the production of phenolic off-flavors such as 4-vinyl guaiacol (4-VG; Gallone et al., 2016), lack of cold tolerance (Nikulin et al., 2018), suboptimal flocculation (Hutzler et al., 2021), and most importantly, inefficient fermentation of the most abundant sugars in brewer’s wort, maltose and maltotriose (Steensels and Verstrepen, 2014; Methner et al., 2019; Nikulin et al., 2020a). Maltotriose is typically utilized at the end of the fermentation (Dietvorst et al., 2005). Residual amounts of the trisaccharide could affect both flavor and ethanol yield, an important economical parameter to produce lagers (Zheng et al., 1994). Maltotriose utilization in yeast is complex, being determined by MAL loci harboring three gene families: MALT encoding an oligosaccharide permease, MALS encoding an α-glucosidase and MALR encoding a regulator responsible for the transcriptional induction of MALT and MALS by maltotriose (Charron et al., 1989; Brouwers et al., 2019b).
In general, non-conventional (wild) yeast species of the Saccharomyces clade are thought not to be able to grow on maltotriose as the sole carbon source (Mtt− phenotype). This also holds for the species S. eubayanus since all strains known so far are unanimously maltotriose-negative (Mtt−) (Mardones et al., 2020). In fact, efficient utilization of maltotriose (Mtt+ phenotype) and particularly its fermentation to ethanol was assumed to be generally restricted to ale S. cerevisiae and S. pastorianus maltotriose-positive (Mtt+) strains at the time when the current study was initiated. The ability of the natural hybrid yeast S. pastorianus to utilize maltotriose by fermentation was initially attributed solely to the S. cerevisiae parent (Hebly et al., 2015; Brickwedde et al., 2018). Later, it was demonstrated that the genome of the Mtt− parent, S. eubayanus, contributed as well to this phenotype possibly due to neofunctionalization of the MAL genes (Baker and Hittinger, 2019; Brouwers et al., 2019b) or regulatory crosstalk between the two parental genomes (Brouwers et al., 2019a). Interestingly, a few strains of the S. eubayanus species including the well-studied CBS 12357T have demonstrated evolvability to maltotriose utilization (Baker and Hittinger, 2019; Brouwers et al., 2019b). These results suggest that S. eubayanus might be able to provide the genetic resources for a Mtt+ phenotype in novel hybrids even without S. cerevisiae as a parent. This approach would be particularly promising when using a second wild-type strain that also brings along certain genetic resources enabling maltotriose utilization.
In this study, high-throughput screening (HTS) for growth on maltotriose as the sole carbon source was applied to a wide range of a Saccharomyces isolates and led to the identification of several previously undescribed non-domesticated wild isolates with an Mtt+ phenotype. Newly identified wild strains were hybridized with the cold-tolerant S. eubayanus CBS 12357T leading to novel non-cerevisiae Saccharomyces hybrids. We evaluated the growth performance of the de novo hybrids on maltotriose in comparison to the established domesticated S. pastorianus CBS 1513 strain. We also tested the potential of the new hybrids to ferment brewer’s wort and brew beer with novel aroma profiles under conditions simulating lager brewing in lab scale.
Materials and methods
Strains, plasmids, and media composition
Saccharomyces parental strains for the HTS for growth on maltotriose and S. pastorianus strains were selected from various geographical locations and origins. The complete list of strains screened for this study is shown in Supplementary Table S1 and the strains selected for further use in hybrids are summarized in Table 1. Yeasts were routinely grown on YPD medium (1% (w/v) yeast extract, 2% (w/v) peptone, and 2% (w/v) glucose) and kept in 25% (w/v) glycerol stocks at −80°C. Selection on Hygromycin BR was performed via the addition of 300 μg mL−1 (final concentration).
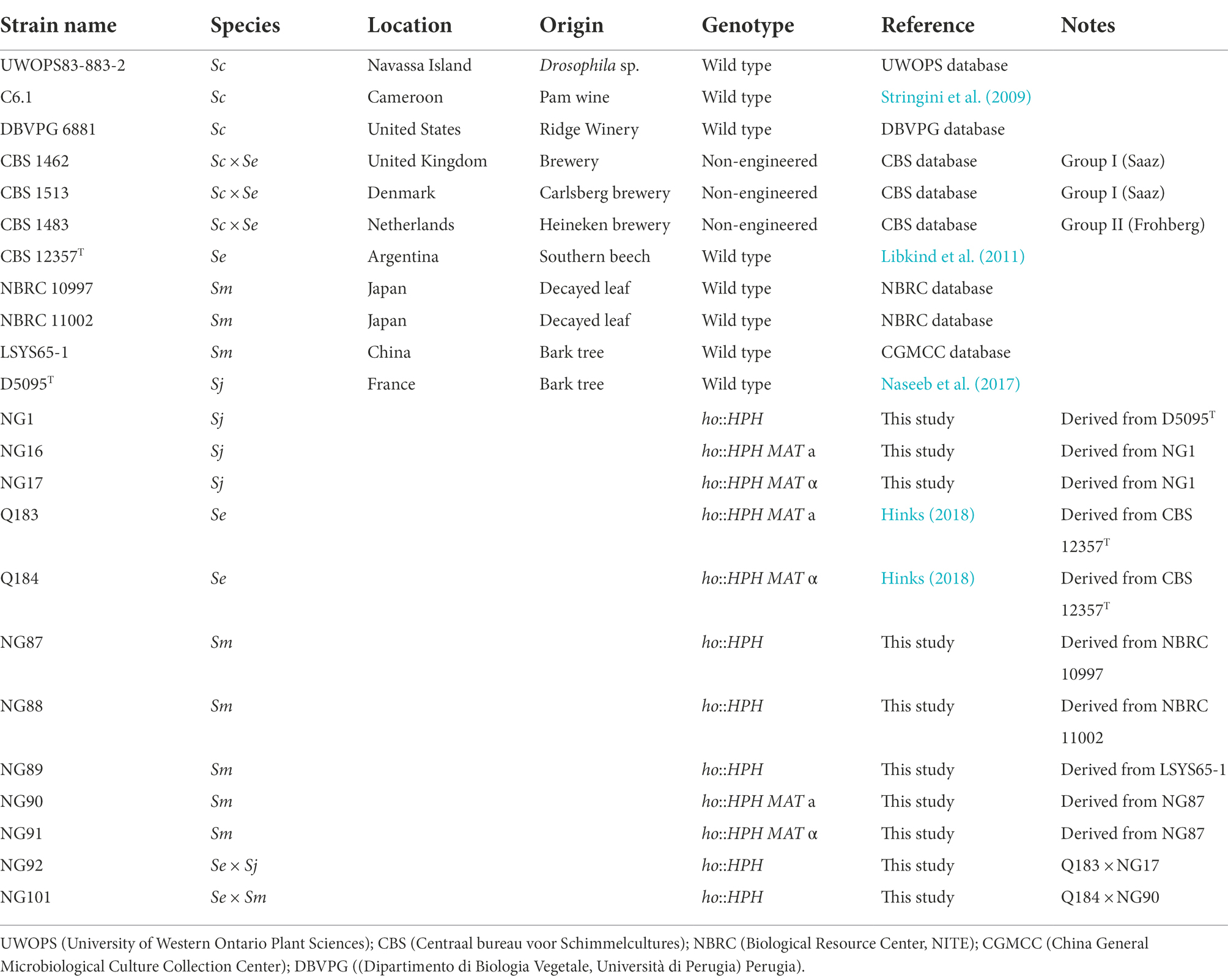
Table 1. List of relevant Saccharomyces strains [S. pastorianus (Sc × Se), S. cerevisiae (Sc), S. eubayanus (Se), S. mikatae (Sm), S. jurei (Sj)] used in this study.
Experiments assessing growth on maltotriose were conducted in either YP medium containing 2% (w/v) maltotriose (YP + Mtt) or synthetic medium (SM) according to Verduyn et al. (1992) with 2% (w/v) maltotriose (SM + Mtt) adjusted to pH 6.5 with 2 M KOH; 2% (w/v) agar was added in solid media. Pre- and intermediate cultures were performed in SM containing 2% (w/v) glucose and cells were washed with SM without carbon source. Wort fermentations were conducted in industrial wort diluted to 12 oP supplemented with 0.6 mg L−1 ZnSO4.7H2O final concentration; sugar content is listed in Supplementary Table S8. To sporulate yeast, potassium acetate (KAc) medium was used containing 2% (w/v) potassium acetate, 0.22% (w/v) yeast extract, and 0.05% (w/v) glucose adjusted at pH 7 with 1 M HCl or 2.5 M NaOH after which 0.087% (w/v) synthetic complete medium powder (Supplementary Table S1) and 2.5% (w/v) agar were added.
General molecular biology techniques
Primers used in this study are listed and detailed in Supplementary Table S2. PCRs for amplifying deletion cassettes, verification, and species-specific identification were performed using OneTaq® DNA Polymerase (New England BioLabs, Germany) or BIOTAQ™ DNA Polymerase (Bioline, UK). Phusion® High-Fidelity DNA Polymerase (New England BioLabs, Germany) was used to amplify fragments for Gibson assembly according to the respective manufacturer’s protocol. Colony PCR was applied via initially suspending cells in 0.02 M NaOH and incubating at 95°C for 10 min and using the supernatant as a DNA template.
Gibson assembly reactions composition and transformation protocols were adapted from the Gibson Assembly® Master Mix instructions (New England BioLabs, Germany) in which 15 μl of Gibson assembly master mix (1.33×; Supplementary Table S1) was used instead of 10 μl and LB medium instead of SOC medium. Transformation of deletion cassettes in Saccharomyces was conducted as described in Gietz et al. (1995) except for the use of lower heat shock temperatures – 37°C for 5 and 20 min applied on S. mikatae and S. jurei strains, respectively.
Screening of Saccharomyces strains for growth on maltotriose
Saccharomyces strains were initially tested for growth on maltotriose via the phenotyping on solid medium (PHENOS) pipeline from Barton et al. (2018). PHENOS produces reproducible proxy growth curves based on automatic absorbance measurements on the colony thickness subtracting the agar absorbance of a rectangular array plate fitting up to 96 strains with four technical replicates. To unequivocally confirm the species designation of the array glycerol stocks, ITS sequencing was performed (White et al., 1990). Strains from Arrays I and II (Supplementary Table S1) were analyzed for growth on YP + Mtt at 30°C. Strains from array III (Supplementary Table S1) were analyzed for growth on SM + Mtt at 25°C. Maltotriose was initially supplied from Sigma-Aldrich, UK with ≥90% purity and added to the YP solid media. Subsequently to decrease impurities, maltotriose was obtained from Glentham Life Sciences, UK, with ≥95% purity and was used instead for further experiments.
Mtt+ candidate strains were tested for growth in liquid SM + Mtt. Initially, 3 ml SM 2% (w/v) glucose pre-cultures were inoculated with single-cell colonies in triplicates in 10 ml glass tubes at 25°C and 250 rpm for ~16 h. Intermediate 3 ml SM 2% (w/v) glucose cultures were prepared by inoculating at 0.2 OD600 and incubating at 25°C at 250 rpm for 24 h. Cells were washed once with SM without carbon source and once with SM + Mtt before inoculating the final 0.7 ml SM + Mtt culture at 0.2 OD600 with two technical replicates in Krystal 24-well clear bottom microplate (Porvair Sciences, Leatherhead, UK) cultivated in the Growth Profiler 1,152 (Enzyscreen, Netherlands) at 200 rpm and 25°C. The Growth Profiler determines the density of the cultures by measuring pixel density from images taken every 40 min of individual wells correlated to equivalent OD600 values via a calibration curve with the equation of the best fit line: OD600 equivalent = 6.1761.10 × Green value3.4784. The calibration curve was based on S. cerevisiae CEN.PK113-1A and was applied to generate OD600 values for all strains tested for a comparative analysis.
Hybrid construction and confirmation
Interspecific hybrids were constructed by firstly disrupting the HO gene in the parental strains to abolish mating-type switching and self-mating and obtain stable haploids (Strathern et al., 1982). The cassettes to integrate the HPH hygromycin marker in S. jurei D5095T, S. mikatae NBRC 10997, S. mikatae NBRC 11002, and S. mikatae LSYS65-1 were amplified from pAG32 (Goldstein and Mccusker, 1999), pUC18_ho::HPH_NBRC_10,997, pUC18_ho::HPH_NBRC_11,002, and pUC18_ho::HPH_LSYS65-1, respectively, (Supplementary Table S1). Homology arms with 45 bp and 500 bp were applied on S. jurei and S. mikatae strains, respectively, targeting the beginning of the HO open reading frame (ORF) to disrupt it and generate strains NG1, NG87, NG88, and NG89.
Sporulation and tetrad dissection were conducted via standard protocols (Sherman and Hicks, 1991) to obtain stable haploids confirmed to be either MAT “a” or “α” via mating-type PCR (Supplementary Table S2) and halo assay (Julius et al., 1983). Stable haploids were not successfully isolated from NG88 (S. mikatae NBRC 11002 ho::HPH) and NG89 (S. mikatae LSYS65-1 ho::HPH). Interspecific hybrids NG92 (Se × Sj) and NG101 (Se × Sm) were constructed via mass mating as described by Sherman and Hicks (1991). Successful hybrid formation was confirmed via initially re-streaking the cell mass for single colonies three times after which they were sporulated to confirm tetrad formation. The production of spores demonstrated their hybrid nature favoring sporulating hybrids in the verification process. Two and four biological replicates of Se × Sj and Se × Sm hybrids, respectively, were successfully isolated. Sporulation efficiency was determined for twenty tetrads per hybrid resulting in ~1% spore viability for each hybrid tested and glycerol stocks were randomly selected and designated as NG92 (Se × Sj) and NG101 (Se × Sm). The interspecific hybrid formation was further verified by individual species-specific PCRs (Muir et al., 2011; Reuben et al., 2013; Naseeb et al., 2021).
Aerobic shake-flask cultivations in synthetic medium with maltotriose as the sole carbon source
Cultivations of 50 ml SM + Mtt in shake flasks were performed in 500 ml Erlenmeyer flasks. Pre- and intermediate cultures and washing steps were conducted as the cultivations in the Growth Profiler upscaled for the intermediate cultures to 10 ml SM 2% (w/v) glucose in 100 ml Erlenmeyer flasks at orbital shaking of 250 rpm at 25°C. Growth was monitored by sampling every 2 h to measure OD600 during the exponential growth phase of each strain. The maximum specific growth rate and lag phase duration were calculated for S. pastorianus CBS 1513 and the interspecific hybrids NG92 (Se × Sj) and NG101 (Se × Sm). Samples for HPLC measurement were taken every 24 h to determine maltotriose consumption. Maltotriose and maltose were measured using an Agilent 1,260 LC equipped with a 1,260 Refractive Index Detector and an Bio-Rad Aminex HPX-42A Column (300 × 7.8 mm, 25 micron) operated at 75°C using MilliQ as eluent at a flow rate of 0.5 ml·min−1.
Wort fermentations and aroma profiling
Pre-cultures of 10 ml YPD in 100 ml Erlenmeyer flasks were inoculated with single-cell colonies in triplicates and incubated at 20°C at 80–120 rpm for 24 h. Intermediate cultures of 30 ml 12 oP wort in 300 ml Erlenmeyer flasks were inoculated at 0.2 OD600 and incubated at 20°C at 80–120 rpm for 48 h. The inoculum was calculated for each replicate by counting cells on a Neubauer Hemocytometer diluted with 50 mM EDTA to avoid flocculation and adjusted to 1.2 × 10^7 cells/mL. Simultaneously, the intermediate culture was gradually cooled to 12°C and the calculated inoculum volumes were mixed with fresh 12 oP wort to a final 200 ml volume with 0.1 ml L−1 autoclaved Antifoam A reagent (Sigma-Aldrich, Germany) in 300 ml infusion bottles with a screw cap and septum (Avantor™, Netherlands) and a 3 cm stirring magnet. Release of CO2 and pressure were controlled via a Microlance needle (Fisher Scientific, UK) covered with cotton to decrease contaminations. Fermentations were carried out at 12°C and the fermentation progress was monitored measuring weight loss through time. Samples were taken with a Microlance needle (Fisher Scientific, UK) and 5 ml were sampled for HPLC measurements of ethanol, glucose, maltose, and maltotriose.
Green beer was analyzed via gas chromatography /mass spectrometry (GC/MS) as described in Rollero et al. (2015) to determine concentrations of volatile compounds of interest. Aroma compound concentrations [mg L−1] were derived from triplicate biological fermentations with high reproducibility including values below and above the sensitivity of the GC/MS analysis per compound listed in Supplementary Table S10. Significant differences between each compound were calculated (p-values <0.05) with one-way ANOVA and Tukey’s multiple comparisons test (Supplementary Table S11). Z-scores were calculated per aroma compound by subtracting the mean of the sample from the observed value, divided by the standard deviation of the sample.
Results
HTS of Saccharomyces strains identified Saccharomyces mikatae strains exhibiting growth on maltotriose
The ability to utilize maltotriose is typically associated with ale S. cerevisiae and S. pastorianus strains, while limited utilization is seen throughout non-domesticated yeast species (Methner et al., 2019; Nikulin et al., 2020a). Nevertheless, relatively few yeast strains have been tested for this phenotype due to lack of appropriate screening tools and the high cost of the oligosaccharide in its pure form. Here, a previously described high-throughput assay for phenotyping microorganisms on solid media - PHENOS (Barton et al., 2018) was adopted to identify non-cerevisiae Saccharomyces strains with the ability to grow on mineral medium with maltotriose as the sole carbon source. Nearly 200 Saccharomyces wild-type strains from various species, geographical locations, and origins were selected for the analysis and were initially tested on solid YP + Mtt at 30°C (Supplementary Figure S1) in two arrays – I and II (Supplementary Table S1). The strains tested comprise all eight currently known species of the Saccharomyces clade (except for S. jurei) and the domesticated Saccharomyces hybrids S. pastorianus and Saccharomyces bayanus. Well-characterized control strains with known maltotriose phenotypes were included, i.e., S. pastorianus CBS 1513 (Mtt+) and S. eubayanus CBS 12357T (Mtt−). The Mtt+ phenotype observed for the Mtt− control strain indicated that the medium required optimization (Supplementary Figure S1). Therefore, synthetic medium with maltotriose (SM + Mtt) and a different maltotriose batch with a higher purity were used in a next step to test the best performers in a single array including two S. jurei strains (Supplementary Table S1: Array III). Performance was ranked based on maximum slope values generated via the PHENOS software (equivalent μmax). The value for the strain S. eubayanus strain CBS 12357T (Mtt−) was set to 1 and the performances of all other strains were shown as fold-change in comparison (Supplementary Figure S2B). The top-performing strains included four non-cerevisiae Saccharomyces strains—the S. jurei D5095T and S. mikatae LSYS65-1 strains which were recently reported as Mtt+ in parallel studies (Giannakou et al., 2021; Peris et al., 2022) and two S. mikatae strains not described yet in literature for the phenotype tested—NBRC 11002 and NBRC 10997 (Supplementary Figure S2).
To eliminate any potential impurities in the components used in our solid medium (i.e., agar) that could have caused the residual growth of Mtt− strains, all superior Mtt+ candidate strains and our Mtt− and Mtt+ control strains were further analyzed in liquid SM + Mtt. To this end, the phenotyping was conducted in small volumes using the Growth Profiler. Under the respective conditions, only minimal growth was observed for CBS 12357T (Mtt−) contrasting the well-performing Mtt+control, i.e., the strain S. pastorianus CBS 1513 (Figure 1). Real quantitative data such as maximum specific growth rates and lag phase durations could not be presented for the newly discovered non-domesticated Mtt+ candidates due to flocculation of all four strains in SM. Nevertheless, all strains successfully grew on maltotriose as the sole carbon source. Three of the four superior Mtt+ strains exhibited a long lag phase while S. mikatae NBRC 11002 interestingly switched metabolism from glucose (pre-culture) to maltotriose in SM without much delay similar to S. pastorianus CBS 1513 (Figure 1).
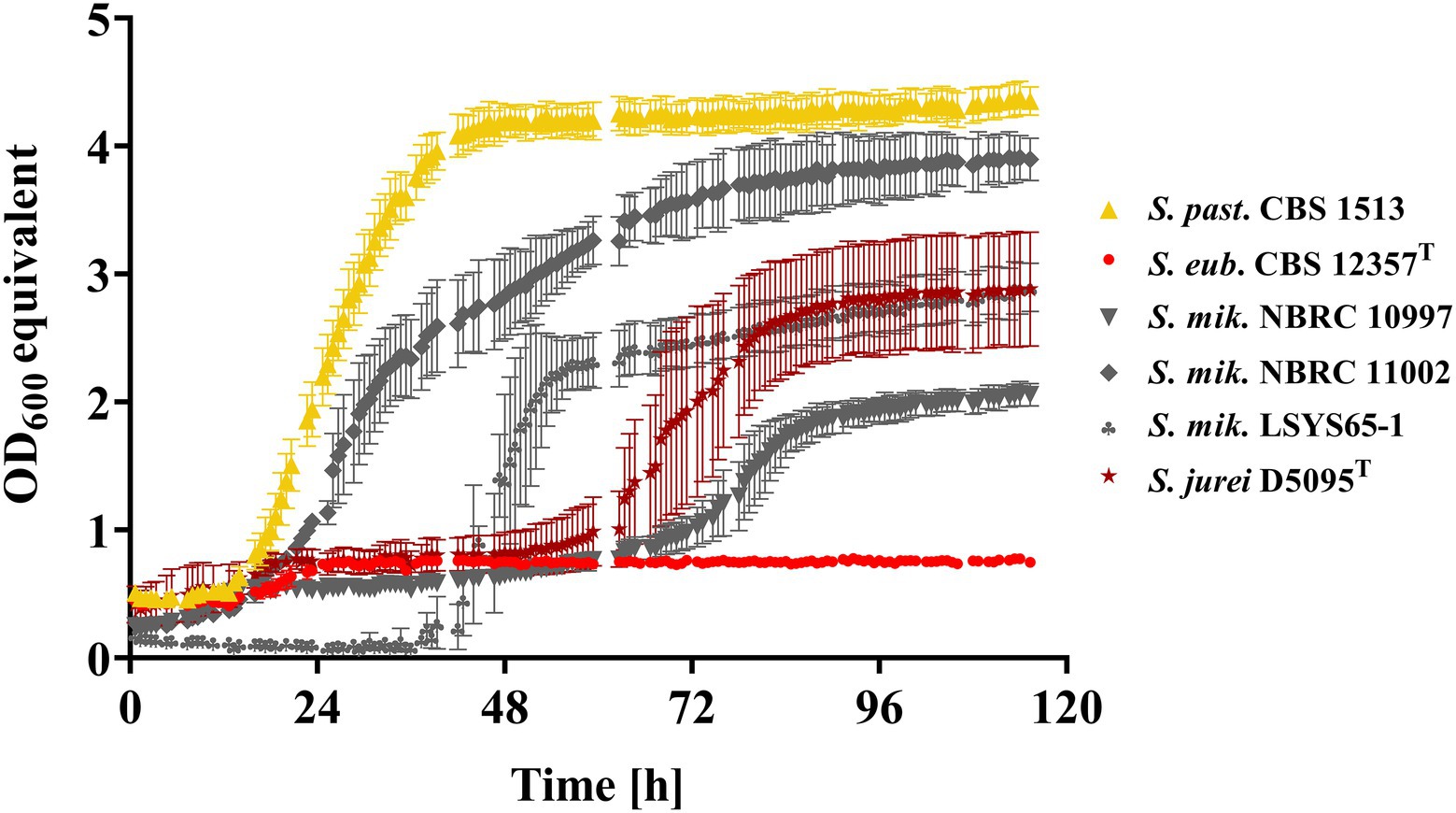
Figure 1. Growth kinetics in synthetic medium containing 2% (w/v) maltotriose (Mtt) of S. pastorianus CBS 1513 (Mtt+; ▲), S. eubayanus CBS 12357T (Mtt−; ●) and the non-S. cerevisiae maltotriose-positive candidates: S. jurei D5095T (★), S. mikatae LSYS65-1 (♣), S. mikatae NBRC 11002 (♦) and S. mikatae NBRC 10997 (▼) recorded via Growth Profiler 1,152. Values are presented as mean and standard deviation of biological triplicates. Raw data are provided in Supplementary Table S3.
Hybridizing Saccharomyces eubayanus CBS 12357T (Mtt−) with Saccharomyces mikatae NBRC 10997 (Mtt+) or Saccharomyces jurei D5095T (Mtt+) resulted in a drastically improved aerobic growth on maltotriose due to heterosis
It was decided to combine the maltotriose utilization capabilities of the newly identified non-cerevisiae Saccharomyces Mtt+ candidates with the Mtt− but cold-tolerant S. eubayanus strain CBS 12357T in a hybrid and check for the heterosis effect in a comparative growth experiment in liquid medium. To this end, it was intended to introduce auxotrophic markers in the parental strains; however, complementary auxotrophs to facilitate mating by complementation were not successfully isolated. Instead, it was decided to rapidly generate stable haploids via genetically engineering the HO endonuclease to abolish self-mating (Strathern et al., 1982). This approach allows the generation of MATa/MATα ho/ho allodiploids which could be further exploited for future quantitative genetic studies on multiple generations of random mating of allotetraploid hybrids with restored fertility (Naseeb et al., 2021). Subsequently, de novo hybrids between S. eubayanus CBS 12357T and S. jurei D5095T and between S. eubayanus CBS 12357T and S. mikatae NBRC 10997, i.e., NG92 (Se × Sj) and NG101 (Se × Sm) were successfully constructed.
To record the kinetics of sugar consumption and ethanol formation in addition to biomass formation of the strains during aerobic growth in liquid SM + Mtt, a higher culture volume was required for frequent sampling of culture supernatants. Thus, the previous Growth Profiler experimental setup (Figure 1) was upscaled to 50 ml cultures in shake flasks which were inoculated with the de novo hybrids - NG92 (Se × Sj) and NG101 (Se × Sm) in parallel with the corresponding parental strains and the S. pastorianus strain CBS 1513 as Mtt+ reference. As expected, the S. pastorianus strain CBS 1513 completed maltotriose consumption after 48 h of cultivation in SM + Mtt (Figure 2A). In contrast, the hybrids NG92 (Se × Sj) and NG101 (Se × Sm) only consumed 25 and 35%, of sugar, respectively, after 120 h of cultivation (Figures 2B,C). Interestingly, the two hybrids did not flocculate in comparison to their respective S. mikatae/S. jurei parental strain allowing growth rate determination during exponential phase. Remarkably, the two de novo hybrids outperformed both parental strains exhibiting growth rates [h−1] comparable to the S. pastorianus strain CBS 1513 and a shorter lag phase [h] compared to the parental Mtt+ strains S. jurei D5095T and S. mikatae NBRC 10997 (Figures 2A–C). NG92 (Se × Sj) displayed a significantly shorter lag phase than NG101 (Se × Sm; Figures 2B,C).
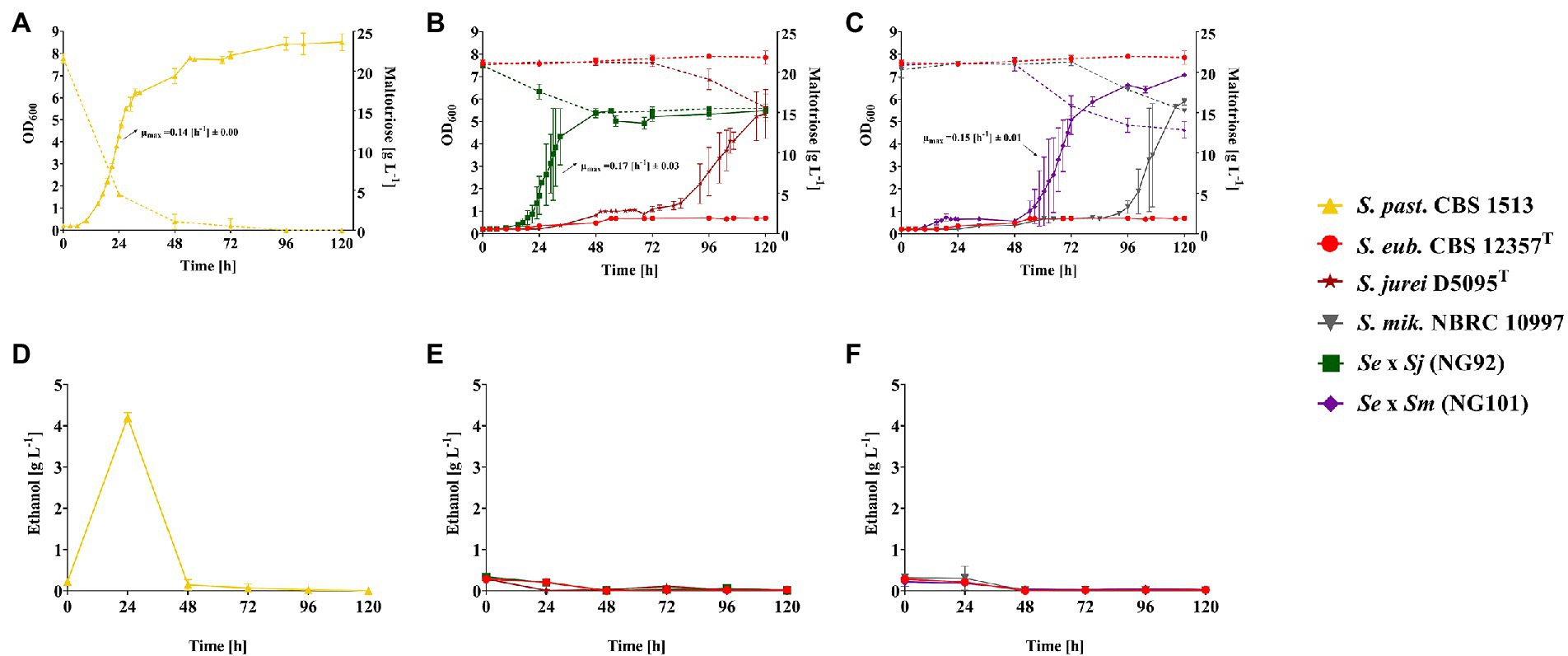
Figure 2. Growth in liquid synthetic medium with 2% (w/v) maltotriose (Mtt) of strains S. pastorianus CBS 1513 (Mtt+; ▲), S. eubayanus CBS 12357T (Mtt−; ●), S. jurei D5095T (Mtt+; ★), S. mikatae NBRC 10997 (Mtt+; ▼) and the constructed de novo hybrids – NG92 (S. eubayanus CBS 12357T × S. jurei D5095T/Se × Sj; ■) and NG101 (S. eubayanus CBS 12357T × S. mikatae NBRC 10997/Se × Sm; ♦). Values are presented as mean and standard deviation of triplicate biological replicates. (A–C) Growth curves depicting OD600 values and maltotriose consumption [g L−1] illustrated with continuous and dotted lines, respectively. Maximum specific growth rates (μmax [h−1]) were calculated for all hybrids but not for the corresponding parental strains due to flocculation. (D–F) Ethanol production [g L−1] measured every 24 h. Raw data are provided in Supplementary Table S7.
Notably, the S. pastorianus strain CBS 1513 produced ethanol from maltotriose under the applied conditions. After 24 h of cultivation, an ethanol concentration of 4 g L−1 was detected (Figure 2D). The fact that there was no remaining ethanol after 48 h demonstrated that the ethanol formed was afterward consumed. However, no ethanol formation could be detected in either of the two de novo hybrids NG92 (Se × Sj) and NG101 (Se × Sm; Figures 2E,F). Probably, these strains indeed did not show any overflow metabolism under the tested conditions or the appropriate time point representing the ethanol production peak was simply missed during sampling.
The de novo hybrids NG92 (Se × Sj) and NG101 (Se × Sm) completely fermented maltose, in contrast to their corresponding Saccharomyces jurei/Saccharomyces mikatae parents, but not maltotriose in lager brewing conditions
Eventually, it was interesting to test the novel hybrids under conditions which are relevant in lager brewing. To this end, brewer’s wort fermentations at 12°C were conducted in lab scale with the constructed de novo hybrids—NG92 (Se × Sj) and NG101 (Se × Sm), all parental strains—S. eubayanus CBS 12357T, S. jurei D5095T, and S. mikatae NBRC 10997 as well as S. pastorianus CBS 1513. The latter strain was added as a reference since it is regularly used in industrial lager beer production (Figure 3). Within the first 48 h, all strains rapidly fermented glucose (Figure 3B) including the residual amounts of fructose and sucrose estimated by the calculated total sugar consumption via weight/CO2 loss measurements (Supplementary Table S8). After this time point, S. pastorianus CBS 1513 and S. eubayanus CBS 12357T, NG92 (Se × Sj), and NG101 (Se × Sm) completely utilized maltose without an apparent lag phase, while the parental S. jurei D5095T and S. mikatae NBRC 10997 strains did not utilize the disaccharide at all. Notably, the de novo hybrids exhibited a slower rate of maltose fermentation compared to S. pastorianus CBS 1513 and S. eubayanus CBS 12357T and required additional 24–48 h to complete the utilization of the sugar (Figure 3C). As expected, the domesticated lager hybrid (S. pastorianus) consumed maltose most rapidly and ethanol formation of 4.4% alcohol by volume (ABV) was achieved. Moreover, S. pastorianus CBS 1513 co-consumed maltose and maltotriose once glucose was depleted as previously described (Magalhães et al., 2016; Figures 3C,D). The two hybrids, NG92 (Se × Sj) and NG101 (Se × Sm), could not utilize maltotriose during the conducted wort fermentations (Figure 3D) similar to S. eubayanus CBS 12357T only reaching an ABV of ~3.5% (Figure 3A) although they efficiently grew on maltotriose as the sole carbon source in the shake-flask cultivations at 25°C, with comparable maximum specific growth rates to S. pastorianus CBS 1513 (Figures 2A–C).
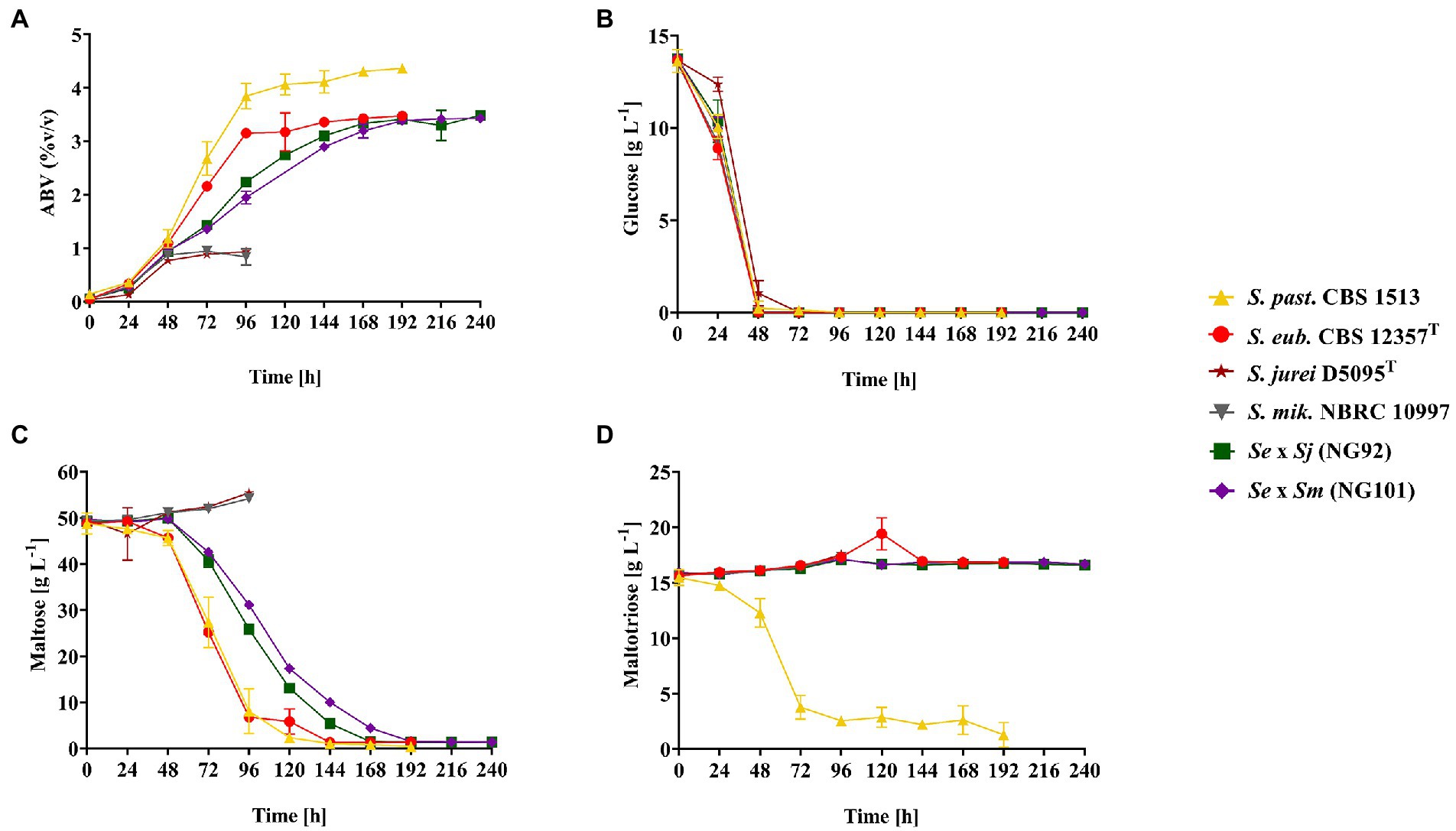
Figure 3. Fermentation in oxygen-limited conditions monitored by ethanol formation represented as alcohol by volume (ABV) [%] at 12°C for 10 days of fermentation of 12 oP wort. The strains tested include the maltotriose-positive (Mtt+) strain S. pastorianus CBS 1513 (▲) as lager reference, the parental strains – S. eubayanus CBS 12357T [maltotriose-negative (Mtt−)] (●), S. jurei D5095T (Mtt+; ★), S. mikatae NBRC 10997 (Mtt+; ▼) and the constructed de novo hybrids - NG92 (S. eubayanus CBS 12357T × S. jurei D5095T/Se × Sj; ■) and NG101 (S. eubayanus CBS 12357T × S. mikatae NBRC 10997/Se × Sm; ♦). Reduced oxygen availability was achieved by using relatively high culture volumes (200 ml in 300 ml infusion bottles). The bottles were equipped with a screw cap and septum with a needle allowing the release of CO2 with reduced shaking frequency. (A) Fermentation progress, (B) Glucose consumption, (C) Maltose consumption and (D) Maltotriose consumption [g L−1] measured every 24 h. Values are presented as mean and standard deviation of triplicate biological fermentations. Raw data are provided in Supplementary Table S8.
Green beer produced by de novo hybrids NG92 (Se × Sj) and NG101 (Se × Sm) showed increased concentrations of fruity flavor esters
The green lager beers produced by the de novo hybrids NG92 (Se × Sj) and NG101 (Se × Sm) as well as their respective S. jurei/S. mikatae parent and the common parent S. eubayanus CBS 12357T (Figure 3) were comparatively analyzed with GC/MS for their aroma profiles. In addition, the lager strain S. pastorianus CBS 1513 was used as a reference. Green beers obtained with the de novo hybrids showed different concentrations of aroma compounds compared to their parental strains (Supplementary Table S9). In fact, heterotic production of desired esters was achieved, especially for NG101 (Se × Sm; Supplementary Table S9). We compare here the actual aroma concentration values detected keeping in mind that the parental strains – S. jurei D5095T and S. mikatae NBRC 10997 could not ferment maltose and maltotriose, therefore, they produced concentrations close or below the lower threshold of measurement. Accurate comparison between the S. eubayanus strain CBS 12357T and the interspecific hybrids is possible reaching an equal final ABV of 3.4–3.5% (Figure 4). Notably, enhanced production of ethyl esters such as ethyl hexanoate (green apple) was observed for NG92 (Se × Sj) which was 2.5-fold higher in comparison to the S. eubayanus CBS 12357T parent, while NG101 (Se × Sm) produced this aroma compound in amounts above its flavor threshold in lager beer (Figure 4). Moreover, ethyl octanoate (apricot) concentration levels were ~2-fold higher for both hybrids compared to CBS 12357T (Figure 4). Comparable concentrations were detected for different acetate esters tested between NG92 (Se × Sj), NG101 (Se × Sm), and CBS 12357T.
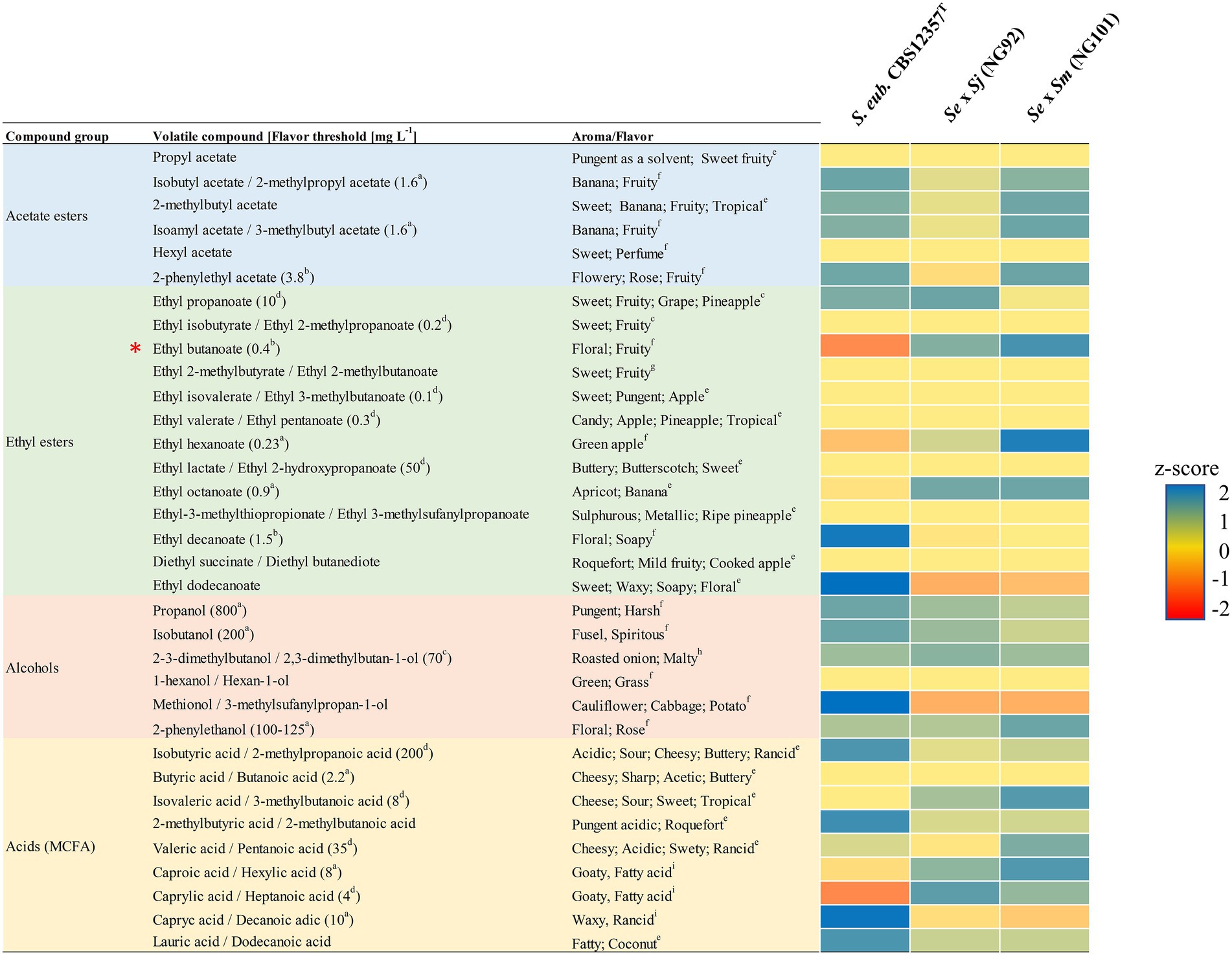
Figure 4. Comparative analysis of aroma production by the generated de novo hybrids NG92 (S. eubayanus CBS 12357T × S. jurei D5095T/Se × Sj) and NG101 (S. eubayanus CBS 12357T × S. mikatae NBRC 10997/Se × Sm) and their common parental strain S. eubayanus CBS 12357T. Colors show the range of the calculated Z-scores per aroma compound indicating production values above average, average, and below average shown with blue, yellow, and red, respectively. The aroma compounds are grouped by acetate esters, ethyl esters, alcohols, and acids (medium-chain fatty acids/MCFA). Flavor thresholds in beer values (shown in brackets) were obtained from Meilgaard (1982) (a); Meilgaard (1975a) (b); Meilgaard (1975b) (c); Harrison (1970) (d). Corresponding aromas/flavors for each volatile compound tested were obtained from The Good Scent Company Information System (2022) (e); Swiegers et al. (2005) (f); PubChem (2022) (g); Dunlevy et al. (2009) (h); Blanco et al. (2016) (i). The calculations are based on aroma compound concentrations [mg L−1] from triplicate biological fermentations with high reproducibility including values below and above the sensitivity of the GC/MS analysis. Detailed quantitative data of the aroma compounds with standard deviation of all strains including the aroma profile of the reference S. pastorianus CBS 1513 strain are presented in Supplementary Table S9. Z-scores generated from concentrations below or just above the threshold of measurement are marked with an asterisk (*).
Interestingly, higher concentrations of isoamyl acetate (banana; 5.22 mg L−1) above the reported flavor threshold in beer and 2-phenylethyl acetate (3.75 mg L−1) close to its threshold of perception were determined for the NG101 (Se × Sm) hybrid compared to the industrial lager hybrid – S. pastorianus CBS 1513 (Supplementary Table S9). Similar amounts were identified between all hybrids and S. eubayanus CBS 12357T for favorable higher alcohols such as 2-phenylethanol (rose, floral) and propanol (spicy, hard). However, the off-flavor 2,3-dimethylbutan-1-ol (roasted onion) associated typically with wine fermentations was produced with a slightly higher concentration in the de novo hybrids (Supplementary Table S9). Undesired medium-chain fatty acids (MCFA) were detected for all strains tested with significantly higher concentrations of caprylic acid (goaty) produced by S. pastorianus CBS 1513 albeit below its taste threshold (Supplementary Table S9). Although we present here a diverse set of aroma compounds with interesting heterotic production of desired fruity esters, the combined aroma profile could not be assessed in a sensory analysis due to the use of genetic engineering to construct the strains.
Discussion
The use of wild yeasts and de novo hybrids in brewing is an attractive approach toward diversifying the lager beer style. However, the challenge is to meet pleasant taste with fermentation performance at the relative cold fermentation temperatures during lager beer brewing. The consumption of the wort sugar maltotriose is an important trait; however, non-conventional Saccharomyces strains able to utilize maltotriose have not been reported at the time when the current study was initiated.
Our HTS identified so far uncharacterized S. mikatae Mtt+ wild strains as well as the recently reported Mtt+ S. mikatae LSYS65-1 strain (Peris et al., 2022). Parallel studies conducted by other authors demonstrated that the two S. jurei strains TUM 629 and D5095T are also able to utilize wort maltotriose to a certain extent (Giannakou et al., 2021; Hutzler et al., 2021) and that some S. mikatae strains are able to grow on this sugar (Peris et al., 2022). The current study included the strain S. jurei D5095T and confirmed its Mtt+ phenotype. The S. jurei D5095T (Mtt+) strain and one of the identified Mtt+ S. mikatae strains in this study (NBRC 10997) were used to construct novel hybrids with the S. eubayanus strain CBS 12357T. We demonstrated several clear heterosis effects in the hybrids as discussed below.
Our HTS of ~200 strains of the clade Saccharomyces for growth on maltotriose as the sole source of carbon revealed limitations when conducted on solid medium. In fact, the negative control strain S. eubayanus CBS 12357T showed significant growth in the PHENOS phenotyping. Still, the latter method successfully identified superior Mtt+ isolates of non-domesticated yeasts. Their Mtt+ phenotypes were afterward confirmed in a liquid medium via the Growth Profiler.
Our finding that S. mikatae strains exhibit growth on maltotriose as the sole carbon source provides an example of the high intraspecies phenotypic diversity uncovered recently for the species despite its low genetic diversity (Peris et al., 2022). However, only a limited number of strains have been tested so far for wort fermentability. In fact, Nikulin et al. (2018) showed that neither α-glucoside (maltose and maltotriose) in brewer’s wort was utilized by the S. mikatae type strain (IFO 1815). Moreover, authors only tested the strain’s performance at 12°C which, in the current study, turned out to be unfavorable for the utilization of the respective wort sugars by S. mikatae.
So far, S. mikatae strains have only been isolated in East Asia even though evidence of the species’ presence in Europe was provided by sequencing the ITS1 region from DNA obtained from the mycobiome of soils surrounding trees in the Italian alps (Alsammar et al., 2019). Interestingly, S. jurei is the most closely related species to S. mikatae (Naseeb et al., 2018) and strains of S. jurei have already been isolated in Europe. The existence of Mtt+ isolates in both species seems to be in contrast to all other non-domesticated Saccharomyces species. Although more data are required to verify this, one might hypothesize that the ability to grow on maltotriose could have been obtained before the evolutionary split of the two species.
Phylogenetic distribution of MAL genes throughout the Saccharomyces clade suggests that maltotriose transport is the limiting factor (Day et al., 2002; Baker and Hittinger, 2019), mediated by Agt1 (Alves et al., 2008) and Mtt1 (Dietvorst et al., 2005), while certain diastatic yeasts strains can degrade the sugar extracellularly prior to transport (Alves et al., 2018). Hutzler et al. (2021) confirmed maltotriose transmembrane transport in the S. jurei strain TUM 629 employing radiolabeled substrate. The authors conducted a genetic analysis on the respective strain and identified a gene with 82.6% sequence identity to S. cerevisiae AGT1, which can potentially encode for a maltotriose permease. This might also apply to S. jurei D5095T, considering the low sequence divergence of the two strains (0.1%). The three new Mtt+ S. mikatae strains identified here (LSYS65-1, NBRC 11002, and NBRC 10997) are interesting candidates to further study the genetic basis underlying the Mtt+ phenotypes in the species S. jurei and S. mikatae. For whole-genome sequencing, it is recommended to use third-generation sequencing to properly assemble sub-telomeric regions which cannot be dissected by Illumina sequencing. As known from other Mtt+ strains of the Saccharomyces clade, the sub-telomeric regions usually contain the MAL genes.
One might further question whether the genetic resources causing the Mtt+ trait of the ale yeast S. cerevisiae can be replaced by the newly discovered Mtt+ wild strains (S. jurei and S. mikatae) when hybridized with the cold-tolerant S. eubayanus CBS 12357T. Enhancing traits relevant in brewing via interspecific Saccharomyces hybrids has been successfully applied in several previous studies (Hebly et al., 2015; Krogerus et al., 2015; Mertens et al., 2015; Nikulin et al., 2018; Giannakou et al., 2021). We hypothesized that heterosis regarding the Mtt+ phenotype could be achieved in such hybrids via synergistic actions of structural and regulatory proteins involved in maltotriose metabolism. Regarding the two sub-genomes of S. pastorianus (S. cerevisiae and S. eubayanus), a regulatory crosstalk between MALR and MALT genes has previously been demonstrated in a de novo S. pastorianus hybrid and provided a possible explanation for the efficient maltotriose fermentation of the domesticated lager yeast (Brouwers et al., 2019a). The common parent, S. eubayanus CBS 12357T, does not contain functional maltotriose transporters but has been shown to exhibit evolvability to the trisaccharide indicating the existence of genetic resources (Baker and Hittinger, 2019; Brouwers et al., 2019b). Interestingly, the de novo hybrids between Mtt− S. eubayanus CBS 12357T and Mtt+ S. jurei D5095T/S. mikatae NBRC 10997 successfully displayed heterosis with regard to their aerobic growth performance on maltotriose as the sole carbon source. Both hybrids, NG92 (Se × Sj) and NG101 (Se × Sm) outperformed the respective parents in terms of this trait, and both exhibited maximum specific growth rates comparable to the industrial S. pastorianus CBS 1513 strain. We presume that, in the de novo hybrids, an increased expression of MALR regulatory genes originating from the S. eubayanus CBS 12357T genome could positively regulate potential transporter genes in the sub-genomes of S. mikatae NBRC 10997 and S. jurei D5095T. Notably, there was a significant difference in the lag phase duration between NG92 (Se × Sj) and NG101 (Se × Sm) upon this switch to maltotriose which might have been caused by differences in the sub-genome interactions between the common parent S. eubayanus CBS 12357T and the S. jurei and S. mikatae parent, respectively.
Regardless of the remarkable maximum growth rates in SM + Mtt, it remains unknown why biomass formation of the de novo hybrids ended before maltotriose was depleted which was in clear contrast to S. pastorianus CBS 1513, which completely consumed the trisaccharide exhibiting a comparable growth rate. The latter partially assimilated maltotriose in a fermentative manner (as visible by ethanol production) in the aerobic shake-flask experiments. Simple shake-flask cultivations are known for limited oxygen supply. Moreover, no control over important variables such as pH is possible (Link and Weuster-Botz, 2011). Oxygen limitations might have resulted in a limited supply of ATP required to transport maltotriose through the cell membrane via the presumed maltotriose transporter in the two hybrids NG92 (Se × Sj) and NG101 (Se × Sm). Notably, the Agt1 permease has been shown to be an active maltotriose H+ symporter (Stambuk et al., 1999). A low substrate affinity of maltotriose transporters might contribute to the sudden growth termination of the de novo hybrids.
While aerobic growth on maltotriose as the sole carbon source is an interesting trait in the context of phylogeny, successful fermentation of the sugar to alcohol is of much greater importance for brewers. While strains of the domesticated species S. pastorianus strains are well known for the latter trait, the de novo hybrids NG92 (Se × Sj) and NG101 (Se × Sm) did not ferment maltotriose in spite of their ability to aerobically assimilate maltotriose. Nevertheless, the hybrids completely fermented maltose which contrasted with their corresponding S. jurei/ S. mikatae parents even though the rates were lower compared to that of the other co-parent, S. eubayanus CBS 12357T. An improvement of the wort sugar fermentation rates could be achieved via prior adaptation of the strains to wort. In fact, re-pitching in maltose-rich medium combined with cell viability monitoring were vital requirements to adapt S. paradoxus for wort fermentations (Nikulin et al., 2020b). Moreover, pre-adaptation to maltotriose was shown to be essential to achieve partial fermentation of the trisaccharide in wort by S. jurei TUM 629 (Hutzler et al., 2021). These findings suggest that further adaptive evolutionary experiments with NG92 (Se × Sj) and NG101 (Se × Sm) could be a promising approach to fully exploit their potential to ferment brewer’s wort. Nevertheless, the genome stability of the genetically engineered hybrids must be evaluated and directed evolution might be necessary to reach an industrial potential. The use of genetic modifications gave a convenient and rapid opportunity to study the fermentative and aromatic potential of such novel de novo hybrids in the lab environment. Still, alternative mating methods without genetic manipulations are strongly preferred if hybrids will be used in large-scale industrial applications in beer (Nakatomi and Gunge, 1972; Alexander et al., 2016; Gorter De Vries et al., 2019).
The most valuable traits of using wild strains and novel hybrids for beer consumers are the exotic aromas and flavors present in the beer produced with such yeast strains. In the hybrids constructed here with S. eubayanus, the parental wild S. jurei and S. mikatae strains contributed to the aroma profile. There was a heterotic production of desired fruity aroma compounds. Both NG92 (Se × Sj) and NG101 (Se × Sm) revealed favorable heterotic production of ethyl octanoate (apricot) and ethyl hexanoate (green apple). The latter was already outlined in the study of Hutzler et al. (2021) for contributing to the tropical final profile caused by S. jurei TUM 629 as detected via a sensory panel. Other desirable aromatic compounds in brewing such as 3-methylbutyl acetate (banana) and 2-phenylethyl acetate (rose, fruity) exhibited higher concentration in both hybrids compared to the commercial S. pastorianus CBS 1513 (Supplementary Table S9). The concentration of 2-phenylethanol was also increased in both de novo hybrids and increased concentrations of this compound have been highlighted to mask unwanted aroma compounds such as 4-VG notes typical for wild strains (Bamforth, 2020). The determination of important off-flavors such as diacetyl (butter-like; Duong et al., 2011) or 4-VG (clove-like; Coghe et al., 2004) was not in the scope of this study. Although the genetically modified organism (GMO)-nature of the constructed hybrids hampered any sensory evaluations, the measured concentrations of aroma compounds suppose a potential exotic tropical final aroma profile.
Conclusion
The increase in the consumer demand for aromatic diversity in beer can be met in lager beer through interspecific hybridization of wild Saccharomyces yeast species. In such attempts, the utilization of maltotriose by the generated hybrids is a relevant trait for brewers. Here, we report maltotriose-utilizing strains from the species S. mikatae and confirmed the Mtt+ phenotype recently identified by others in strains of S. jurei (D5095T) and S. mikatae (LSYS65-1). We constructed two de novo hybrids: (i) between an Mtt+ S. mikatae strain isolated in the current study (NBRC 10997) and the Mtt− but cold-tolerant S. eubayanus CBS 12357T and (ii) between the Mtt+ S. jurei strain D5095T and the S. eubayanus strain CBS 12357T. Both hybrids displayed heterosis on maltotriose and outperformed both parents regarding aerobic growth on maltotriose synthetic medium achieving growth rates comparable to the industrial reference strain S. pastorianus CBS 1513. In contrast to their S. mikatae/S. jurei parents, the de novo hybrids completely fermented maltose in wort fermentations in a lager brewing setting and produced enhanced amounts of favored fruity esters. Although the hybrids failed regarding the fermentation of maltotriose, there is the potential to improve this relevant trait as well as the rate of maltose fermentation by future adaptive laboratory evolution. The current study might therefore pave the way toward non-cerevisiae Saccharomyces hybrids able to completely ferment all wort sugars and brew exotic lager beers. By using complementary auxotrophic mutants and/or different physiological traits of the partners, non-GMO hybrids could be produced which could contribute to the diversification of lager beers in the future.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
NG conducted the experimental work and drafted the manuscript. ÁC-M, SW, and SA provided experimental assistance. EL and EN provided support in experimental setup and writing guidance. NK provided support in experimental setup and critical revision of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
The project was funded by the European Union’s Horizon 2020 Research and Innovation Program under the Marie Skłodowska-Curie Grant Agreement No. 764927.
Acknowledgments
We express our enormous gratitude to Dave Barton (University of Leicester), Rafael Álvarez (INRAE Montpellier/University of Leicester), Tom Elink Schuurman (R&D, HEINEKEN), and Viktor Boer (R&D, HEINEKEN) for advisory support throughout the project. Strains were obtained from various culture collections referenced in the text. Strains from the CGMCC database were obtained under Material Transfer Agreement (MTA) for academic research use.
Conflict of interest
NK was employed by HEINEKEN Supply Chain B.V.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.1025132/full#supplementary-material
References
Alexander, W. G., Peris, D., Pfannenstiel, B. T., Opulente, D. A., Kuang, M., and Hittinger, C. T. (2016). Efficient engineering of marker-free synthetic allotetraploids of saccharomyces. Fungal Genet. Biol. 89, 10–17. doi: 10.1016/j.fgb.2015.11.002
Alsammar, H. F., Naseeb, S., Brancia, L. B., Gilman, R. T., Wang, P., and Delneri, D. (2019). Targeted metagenomics approach to capture the biodiversity of saccharomyces genus in wild environments. Environ. Microbiol. Rep. 11, 206–214. doi: 10.1111/1758-2229.12724
Alves, S. L., Herberts, R. A., Hollatz, C., Trichez, D., Miletti, L. C., de Araujo, P. S., et al. (2008). Molecular analysis of maltotriose active transport and fermentation by Saccharomyces cerevisiae reveals a determinant role for the AGT1 permease. Appl. Environ. Microbiol. 74, 1494–1501. doi: 10.1128/AEM.02570-07
Alves, S. L., Thevelein, J. M., and Stambuk, B. U. (2018). Extracellular maltotriose hydrolysis by Saccharomyces cerevisiae cells lacking the AGT1 permease. Lett. Appl. Microbiol. 67, 377–383. doi: 10.1111/lam.13048
Baker, E. C. P., and Hittinger, C. T. (2019). Evolution of a novel chimeric maltotriose transporter in Saccharomyces eubayanus from parent proteins unable to perform this function. PLoS Genet. 15:e1007786. doi: 10.1371/journal.pgen.1007786
Bamforth, C. W. (2020). The Horace Brown medal. Forever in focus: researches in malting and brewing sciences. J. Inst. Brew. 126, 4–13. doi: 10.1002/jib.594
Barton, D. B. H., Georghiou, D., Dave, N., Alghamdi, M., Walsh, T. A., Louis, E. J., et al. (2018). PHENOS: a high-throughput and flexible tool for microorganism growth phenotyping on solid media. BMC Microbiol. 18:9. doi: 10.1186/s12866-017-1143-y
Batshinskaya, A. A. (1914). Entwicklungsgeschichte und Kultur des neuen Hefepilzes Saccharomyces paradoxus. J. Microbiol. Epidemiol. Immunobiol. 1, 231–247.
Beijerinck, M. (1898). Uber Regeneration der Sporenbildung bei Alkohol Hefen, wo diese Funktion im Verchwinden begriffen ist. Cent. Bakt 4, 657–731.
Bing, J., Han, P. J., Liu, W. Q., Wang, Q. M., and Bai, F. Y. (2014). Evidence for a far east Asian origin of lager beer yeast. Curr. Biol. 24, R380–R381. doi: 10.1016/j.cub.2014.04.031
Blanco, C. A., Andrés-Iglesias, C., and Montero, O. (2016). Low-alcohol beers: flavor compounds, defects, and improvement strategies. Crit. Rev. Food Sci. Nutr. 56, 1379–1388. doi: 10.1080/10408398.2012.733979
Brickwedde, A., Brouwers, N., van den Broek, M., Gallego Murillo, J. S., Fraiture, J. L., Pronk, J. T., et al. (2018). Structural, physiological and regulatory analysis of maltose transporter genes in Saccharomyces eubayanus CBS 12357T. Front. Microbiol. 9:1786. doi: 10.3389/fmicb.2018.01786
Brouwers, N., Brickwedde, A., Gorter de Vries, A. R., van den Broek, M., Weening, S. M., van den Eijnden, L., et al. (2019a). Himalayan Saccharomyces eubayanus genome sequences reveal genetic markers explaining heterotic maltotriose consumption by Saccharomyces pastorianus hybrids. Appl. Environ. Microbiol. 85. doi: 10.1128/AEM.01516-19
Brouwers, N., Gorter De Vries, A. R., Van Den Broek, M., Weening, S. M., Elink Schuurman, T. D., Kuijpers, N. G. A., et al. (2019b). In vivo recombination of Saccharomyces eubayanus maltose-transporter genes yields a chimeric transporter that enables maltotriose fermentation. PLoS Genet. 15, 1–30. doi: 10.1371/journal.pgen.1007853
Charron, M. J., Read, E., Haut, S. R., Michels, C. A., Dubin, R. A., Perkins, E., et al. (1989). Molecular evolution of the telomere-associated MAL loci of Saccharomyces. Genetics 122, 307–316. doi: 10.1093/genetics/122.2.307
Coghe, S., Benoot, K., Delvaux, F., Vanderhaegen, B., and Delvaux, F. R. (2004). Ferulic acid release and 4-Vinylguaiacol formation during Brewing and fermentation: indications for Feruloyl esterase activity in Saccharomyces cerevisiae. J. Agric. Food Chem. 52, 602–608. doi: 10.1021/jf0346556
Day, R. E., Rogers, P. J., Dawes, I. W., and Higgins, V. J. (2002). Molecular analysis of maltotriose transport and utilization by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 68, 5326–5335. doi: 10.1128/AEM.68.11.5326-5335.2002
Dietvorst, J., Londesborough, J., and Steensma, H. Y. (2005). Maltotriose utilization in lager yeast strains: MTTI encodes a maltotriose transporter. Yeast 22, 775–788. doi: 10.1002/yea.1279
Dunlevy, J. D., Kalua, C. M., Keyzers, R. A., and Boss, P. K. (2009). Grapevine molecular physiology and biotechnology : Second Edition (Springer, Dordrecht), 293–340.
Dunn, B., and Sherlock, G. (2008). Reconstruction of the genome origins and evolution of the hybrid lager yeast Saccharomyces pastorianus. Genome Res. 18, 1610–1623. doi: 10.1101/gr.076075.108
Duong, C. T., Strack, L., Futschik, M., Katou, Y., Nakao, Y., Fujimura, T., et al. (2011). Identification of Sc-type ILV6 as a target to reduce diacetyl formation in lager brewers’ yeast. Metab. Eng. 13, 638–647. doi: 10.1016/j.ymben.2011.07.005
Eizaguirre, J. I., Peris, D., Rodríguez, M. E., Lopes, C. A., de Los Ríos, P., Hittinger, C. T., et al. (2018). Phylogeography of the wild lager-brewing ancestor (Saccharomyces eubayanus) in Patagonia. Environ. Microbiol. 20, 3732–3743. doi: 10.1111/1462-2920.14375
Gallone, B., Steensels, J., Mertens, S., Dzialo, M. C., Gordon, J. L., Wauters, R., et al. (2019). Interspecific hybridization facilitates niche adaptation in beer yeast. Nat. Ecol. Evol. 3, 1562–1575. doi: 10.1038/s41559-019-0997-9
Gallone, B., Steensels, J., Prahl, T., Soriaga, L., Saels, V., Herrera-Malaver, B., et al. (2016). Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cells 166, 1397–1410.e16. doi: 10.1016/j.cell.2016.08.020
Garavaglia, C., and Swinnen, J. (2017). The craft beer revolution: an international perspective. Choices 32, 1–8. doi: 10.22004/ag.econ.261900
Gayevskiy, V., and Goddard, M. R. (2016). Saccharomyces eubayanus and Saccharomyces arboricola reside in North Island native New Zealand forests. Environ. Microbiol. 18, 1137–1147. doi: 10.1111/1462-2920.13107
Giannakou, K., Visinoni, F., Zhang, P., Nathoo, N., Jones, P., Cotterrell, M., et al. (2021). Biotechnological exploitation of Saccharomyces jurei and its hybrids in craft beer fermentation uncovers new aroma combinations. Food Microbiol. 100:103838. doi: 10.1016/j.fm.2021.103838
Gietz, R. D., Schiestl, R. H., Willems, A. R., and Woods, R. A. (1995). Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast 11, 355–360. doi: 10.1002/yea.320110408
Goldstein, A. L., and Mccusker, J. H. (1999). Three new dominant drug resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast 15, 1541–1553. doi: 10.1002/(SICI)1097-0061(199910)15:14<1541::AID-YEA476>3.0.CO;2-
Gorter De Vries, A. R., Koster, C. C., Weening, S. M., Luttik, M. A. H., Kuijpers, N. G. A., Geertman, J. M. A., et al. (2019). Phenotype-independent isolation of interspecies Saccharomyces hybrids by dual-dye fluorescent staining and fluorescence-activated cell sorting. Front. Microbiol. 10:871. doi: 10.3389/fmicb.2019.00871
Harrison, G. A. F. (1970). The flavour of beer – a review. Instr. Brew. 76, 486–495. doi: 10.1002/j.2050-0416.1970.tb03333.x
Hebly, M., Brickwedde, A., Bolat, I., Driessen, M. R. M., de Hulster, E. A. F., van den Broek, M., et al. (2015). S. cerevisiae × S. eubayanus interspecific hybrid, the best of both worlds and beyond. FEMS Yeast Res. 15, 1–14. doi: 10.1093/femsyr/fov005
Hinks, A. (2018). Saccharomyces hybrids: Generation and analysis. Dissertation, Leicester, UK: University of Leicester.
Hornsey, I. (2003). A history of Beer and Brewing. London, United Kingdom: Royal Society of Chemistry.
Hutzler, M., Michel, M., Kunz, O., Kuusisto, T., Magalhães, F., Krogerus, K., et al. (2021). Unique Brewing-relevant properties of a strain of Saccharomyces jurei isolated from ash (Fraxinus excelsior). Front. Microbiol. 12:645271. doi: 10.3389/fmicb.2021.645271
Jaeger, S. R., Worch, T., Phelps, T., Jin, D., and Cardello, A. V. (2020). Preference segments among declared craft beer drinkers: perceptual, attitudinal and behavioral responses underlying craft-style vs. traditional-style flavor preferences. Food Qual. Pref. 82:103884. doi: 10.1016/j.foodqual.2020.103884
Julius, D., Blair, L., Brake, A., Sprague, G., and Thorner, J. (1983). Yeast alpha factor is processed from a larger precursor polypeptide: the essential role of a membrane-bound dipeptidyl aminopeptidase. Cells 32, 839–852. doi: 10.1016/0092-8674(83)90070-3
Krogerus, K., Magalhães, F., Vidgren, V., and Gibson, B. (2015). New lager yeast strains generated by interspecific hybridization. J. Ind. Microbiol. Biot. 42, 769–778. doi: 10.1007/s10295-015-1597-6
Lallemand Brewing (2022). Available at: https://www.lallemandbrewing.com/en/united-states/product-details/lalbrew-novalager/ (Accessed September 27, 2022).
Lovin Dublin (2018). Available at: https://lovindublin.com/sponsored/limited-edition-wild-lager-himalayas-h32-heineken-dubling (Accessed January 18, 2022).
Libkind, D., Hittinger, C. T., Valeŕio, E., Goncaalves, C., Dover, J., Johnston, M., et al. (2011). Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast. Proc. Natl. Acad. Sci. U. S. A. 108, 14539–14544. doi: 10.1073/pnas.1105430108
Link, H., and Weuster-Botz, D. (2011). “Medium formulation and development,” in Comprehensive Biotechnology. ed. M. Moo-Young (Amsterdam, Netherland: Elsevier), 119–134.
Magalhães, F., Vidgren, V., Ruohonen, L., and Gibson, B. (2016). Maltose and maltotriose utilisation by group I strains of the hybrid lager yeast Saccharomyces pastorianus. FEMS Yeast Res. 16:fow053. doi: 10.1093/femsyr/fow053
Mardones, W., Villarroel, C. A., Krogerus, K., Tapia, S. M., Urbina, K., Oporto, C. I., et al. (2020). Molecular profiling of beer wort fermentation diversity across natural Saccharomyces eubayanus isolates. Microb. Biotechnol. 13, 1012–1025. doi: 10.1111/1751-7915.13545
Meilgaard, M. C. (1975a). Flavor chemistry of beer: part II: flavor and threshold of 239 aroma volatiles. Tech. Q. MBAA 12, 151–168.
Meilgaard, M. C. (1975b). Flavor chemistry of beer. I. Flavor interaction between principal volatiles. Tech. Q. MBAA 12, 107–117.
Meilgaard, M. C. (1982). Prediction of flavor differences between beers from their chemical composition. J. Agric. Food Chem. 30, 1009–1017. doi: 10.1021/jf00114a002
Mertens, S., Steensels, J., Saels, V., de Rouck, G., Aerts, G., and Verstrepen, K. J. (2015). A large set of newly created interspecific Saccharomyces hybrids increases aromatic diversity in lager beers. Appl. Environ. Microbiol. 81, 8202–8214. doi: 10.1128/AEM.02464-15
Methner, Y., Hutzler, M., Matoulková, D., Jacob, F., and Michel, M. (2019). Screening for the brewing ability of different non-saccharomyces yeasts. Fermentation 5, 1–23. doi: 10.3390/fermentation5040101
Muir, A., Harrison, E., and Wheals, A. (2011). A multiplex set of species-specific primers for rapid identification of members of the genus Saccharomyces. FEMS Yeast Res. 11, 552–563. doi: 10.1111/j.1567-1364.2011.00745.x
Nakatomi, Y., and Gunge, N. (1972). Genetic mechanisms of rare matings of the yeast Saccharomyces cerevisiae heterozygous for mating. Genetics 70, 41–58. doi: 10.1093/genetics/70.1.41
Naseeb, S., Alsammar, H., Burgis, T., Donaldson, I., Knyazev, N., Knight, C., et al. (2018). Whole genome sequencing, de novo assembly and phenotypic profiling for the new budding yeast species Saccharomyces jurei. G3 8, 2967–2977. doi: 10.1534/g3.118.200476
Naseeb, S., James, S. A., Alsammar, H., Michaels, C. J., Gini, B., Nueno-Palop, C., et al. (2017). Saccharomyces jurei sp. Nov., isolation and genetic identification of a novel yeast species from Quercus robur. Int. J. Syst. Evol. Microbiol. 67, 2046–2052. doi: 10.1099/ijsem.0.002013
Naseeb, S., Visinoni, F., Hu, Y., Hinks Roberts, A. J., Maslowska, A., Walsh, T., et al. (2021). Restoring fertility in yeast hybrids: breeding and quantitative genetics of beneficial traits. Proc. Natl. Acad. Sci. U. S. A. 118, 1–12. doi: 10.1073/pnas.2101242118/
Naumov, G. I., James, S. A., Naumova, E. S., Louis, E. J., and Roberts, I. N. (2000). Three new species in the Saccharomyces sensu stricto complex: Saccharomyces cariocanus, Saccharomyces kudriavzevii and Saccharomyces mikatae. Int. J. Syst. Evol. Microbiol. 50, 1931–1942. doi: 10.1099/00207713-50-5-1931
Nespolo, R. F., Villarroel, C. A., Oporto, C. I., Tapia, S. M., Vega-Macaya, F., Urbina, K., et al. (2020). An out-of-Patagonia migration explains the worldwide diversity and distribution of Saccharomyces eubayanus lineages. PLoS Genet. 16:e1008777. doi: 10.1371/journal.pgen.1008777
Nikulin, J., Eerikäinen, R., Hutzler, M., and Gibson, B. (2020a). Brewing characteristics of the maltotriose-positive yeast Zygotorulaspora florentina isolated from oak. Beverages 6, 1–19. doi: 10.3390/beverages6040058
Nikulin, J., Krogerus, K., and Gibson, B. (2018). Alternative Saccharomyces interspecies hybrid combinations and their potential for low-temperature wort fermentation. Yeast 35, 113–127. doi: 10.1002/yea.3246
Nikulin, J., Vidgren, V., Krogerus, K., Magalhães, F., Valkeemäki, S., Kangas-Heiska, T., et al. (2020b). Brewing potential of the wild yeast species Saccharomyces paradoxus. Eur. Food Res. Technol. 246, 2283–2297. doi: 10.1007/s00217-020-03572-2
Okuno, M., Kajitani, R., Ryusui, R., Morimoto, H., Kodama, Y., and Itoh, T. (2015). Next-generation sequencing analysis of lager brewing yeast strains reveals the evolutionary history of interspecies hybridization. DNA Res. 23, dsv037–dsv080. doi: 10.1093/dnares/dsv037
Peris, D., Sylvester, K., Libkind, D., Gonçalves, P., Sampaio, J. P., Alexander, W. G., et al. (2014). Population structure and reticulate evolution of Saccharomyces eubayanus and its lager-brewing hybrids. Mol. Ecol. 23, 2031–2045. doi: 10.1111/mec.12702
Peris, D., Ubbelohde, E. J., Kuang, M. C., Kominek, J., Langdon, Q. K., Adams, M., et al. (2022). Macroevolutionary diversity of traits and genomes in the model yeast genus Saccharomyces. BioRxiv [Preprint]. doi: 10.1101/2022.03.30.486421
PubChem (2022). Available at: https://pubchem.ncbi.nlm.nih.gov (Accessed March 12, 2022).
Reuben, J., Pengelly, W., and Alan, E. (2013). Rapid identification of Saccharomyces eubayanus and its hybrids. FEMS Yeast Res. 13, 156–161. doi: 10.1111/1567-1364.12018
Rollero, S., Bloem, A., Camarasa, C., Sanchez, I., Ortiz-Julien, A., Sablayrolles, J. M., et al. (2015). Combined effects of nutrients and temperature on the production of fermentative aromas by Saccharomyces cerevisiae during wine fermentation. Appl. Microbiol. Biotechnol. 99, 2291–2304. doi: 10.1007/s00253-014-6210-9
Salazar, A. N., Gorter De Vries, A. R., van den Broek, M., Brouwers, N., de La Torre Cortès, P., Kuijpers, N. G. A., et al. (2019). Chromosome level assembly and comparative genome analysis confirm lager-brewing yeasts originated from a single hybridization. BMC Genomics 20:916. doi: 10.1186/s12864-019-6263-3
Sherman, F., and Hicks, J. (1991). Micromanipulation and dissection of asci. Meth. Enzymol. 194, 21–37. doi: 10.1016/0076-6879(91)94005-w
Stambuk, B. U., da Silva, M. A., Panek, A. D., and de Araujo, P. S. (1999). Active-glucoside transport in Saccharomyces cerevisiae. FEMS Microbiol. Lett. 170, 105–110. doi: 10.1111/j.1574-6968.1999.tb13361.x
Steensels, J., and Verstrepen, K. J. (2014). Taming wild yeast: potential of conventional and nonconventional yeasts in industrial fermentations. Annu. Rev. Microbiol. 68, 61–80. doi: 10.1146/annurev-micro-091213-113025
Strathern, J. N., Klar, A. J. S., Hicks, J. B., Abraham, J. A., Ivy, J. M., Nasmyth, K. A., et al. (1982). Cassettes is initiated by a cut in the MAT locus homothallic switching of yeast mating type double-stranded cut in the MAT locus. Cells 31, 183–192. doi: 10.1016/0092-8674(82)90418-4
Stringini, M., Comitini, F., Taccari, M., and Ciani, M. (2009). Yeast diversity during tapping and fermentation of palm wine from Cameroon. Food Microbiol. 26, 415–420. doi: 10.1016/j.fm.2009.02.006
Swiegers, J. H., Bartowsky, E., and Pretorius, I. S. (2005). Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 11, 139–173. doi: 10.1111/j.1755-0238.2005.tb00285.x
The Good Scent Company Information System (2022). Available at: http://www.thegoodscentscompany.com (Accessed March 12, 2022).
Verduyn, C., Postma, E., Scheffers, W. A., and van Dijken, J. P. (1992). Effect of benzoic acid on metabolic fluxes in yeasts: a continuous-culture study on the regulation of respiration and alcoholic fermentation. Yeast 8, 501–517. doi: 10.1002/yea.320080703
Wang, S. A., and Bai, F. Y. (2008). Saccharomyces arboricolus sp. nov., a yeast species from tree bark. Int. J. Syst. Evol. Microbiol. 58, 510–514. doi: 10.1099/ijs.0.65331-0
White, T. J., Bruns, T. D., Lee, S. B., and Taylor, J. W. (1990). Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. (New York: Academic Press), 315–322.
Keywords: lager brewing, yeast, de novo hybrids, diversity, maltotriose utilization, heterosis, novel aroma profiles, non-cerevisiae
Citation: Gyurchev NY, Coral-Medina Á, Weening SM, Almayouf S, Kuijpers NGA, Nevoigt E and Louis EJ (2022) Beyond Saccharomyces pastorianus for modern lager brews: Exploring non-cerevisiae Saccharomyces hybrids with heterotic maltotriose consumption and novel aroma profile. Front. Microbiol. 13:1025132. doi: 10.3389/fmicb.2022.1025132
Edited by:
Pavol Sulo, Comenius University, SlovakiaReviewed by:
David Peris Navarro, University of Oslo, NorwayMatthias Sipiczki, University of Debrecen, Hungary
Copyright © 2022 Gyurchev, Coral-Medina, Weening, Almayouf, Kuijpers, Nevoigt and Louis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nikola Y. Gyurchev, n.gyurchev@gmail.com; Edward J. Louis, ejl21@leicester.ac.uk
†These authors share last authorship