Emergence of a Zoonotic Pathogen in a Coastal Marine Sentinel: Capillaria hepatica (syn. Calodium hepaticum)-Associated Hepatitis in Southern Sea Otters (Enhydra lutris nereis)
 Melissa A. Miller1,2*
Melissa A. Miller1,2*  Pádraig J. Duignan3
Pádraig J. Duignan3  Erin Dodd1
Erin Dodd1  Francesca Batac1
Francesca Batac1  Michelle Staedler4 Joseph A. Tomoleoni5
Michelle Staedler4 Joseph A. Tomoleoni5  Michael Murray4 Heather Harris3 Chris Gardiner6
Michael Murray4 Heather Harris3 Chris Gardiner6- 1Marine Wildlife Veterinary Care and Research Center, California Department of Fish and Wildlife, Santa Cruz, CA, United States
- 2Wildlife Health Center, School of Veterinary Medicine, University of California, Davis, Davis, CA, United States
- 3The Marine Mammal Center, Sausalito, CA, United States
- 4Monterey Bay Aquarium, Monterey, CA, United States
- 5Santa Cruz Field Station, Western Ecological Research Center, U. S. Geological Survey, Santa Cruz, CA, United States
- 6JPC Veterinary Pathology Services, The Joint Pathology Center, Silver Spring, MD, United States
Capillaria hepatica is a globally distributed zoonotic nematode parasite that most commonly infects feral and native rats. Soil contact, pica, and living in close proximity to rat populations are risk factors for human infection. Larval nematodes and eggs that were morphologically consistent with C. hepatica were observed microscopically in livers of stranded southern sea otters (Enhydra lutris nereis) from California. Large (90–100 × 45–55 μm), barrel-shaped non-embryonated parasite eggs with large polar prominences and a roughened or striated surface, or 105–120 μm diameter larval aphasmid nematode profiles with a prominent stichosome and hypodermal bands were observed in the livers of three otters. The liver of a fourth animal exhibited serpiginous tracts of necrosis, micro-cavitation and pleocellular inflammation, with intralesional linear eosinophilic material that resembled cuticle from degenerating metazoan parasites. Capillaria hepatica-associated hepatitis and capsular adhesions were the cause of death for one otter, and parasite-associated liver lesions were observed in all cases. All infected otters were adult females that stranded from 2006 through 2016 at multiple sites along the central California coast. All cases stranded from December through May; during and soon after peak seasonal precipitation and land-sea runoff in California. This same seasonal pattern has been reported for other land-based parasites infecting southern sea otters. Neither C. hepatica, nor any similar nematodes have been reported from marine mammals, and southern sea otters are not typical hosts for C. hepatica or any other nematode parasites. The most likely route of exposure was via freshwater runoff containing embryonated eggs liberated from predated or decomposing terrestrial hosts, especially rats. Similar to the land-based parasites Toxoplasma gondii and Sarcocystis neurona, C. hepatica eggs may be concentrated and transmitted through filter-feeding marine invertebrates that serve as southern sea otter prey, which may also pose an unrecognized public health risk for people who consume these species.
Introduction
Capillaria hepatica (syn. Calodium hepaticum) is a globally distributed nematode parasite of mammals, including humans. Wild and feral rodents of the genus Rattus serve as the main source of environmental contamination by parasite eggs in urban environments. Under crowded and unsanitary conditions >70% of rats can be infected, and cannibalism facilitates transmission between successive rodent generations (Fuehrer et al., 2014a). Ingestion of water or food contaminated by embryonated C. hepatica eggs can cause infection in humans and other mammals (Fuehrer et al., 2014b). In an early report of human infection, eggs compatible with C. hepatica were detected in material recovered from a 3rd to 4th century grave in France (Mowlavi et al., 2014).
Capillaria hepatica is the only vertebrate nematode known to require host death for completion of the parasite life cycle (Wright, 1961). Following 3 weeks of hepatic maturation, females begin laying eggs around portal tracts. Nematode migration within the parenchyma can cause necrosis (Redrobe and Patterson-Kane, 2005), and adults ultimately die and disintegrate. Eggs and dying parasites elicit an inflammatory response composed of macrophages, multinucleated giant cells, lymphocytes, plasma cells and eosinophils, often resulting in granuloma formation (Redrobe and Patterson-Kane, 2005). In live animals, eggs remain un-embryonated in the hepatic parenchyma, and are not released through the biliary tract.
Eggs are deposited in the liver in dense masses and may be most concentrated in the subcapsular parenchyma (Moreira et al., 2013). Due to localized egg deposition and exuberant perilesional fibroplasia, affected areas can appear grossly as serpiginous or multinodular yellow-white capsular spots (Moreira et al., 2013). On histopathology the distinctive eggs are barrel-shaped and un-embryonated, with two shallow polar prominences (i.e., polar plugs), and a characteristically rough, pitted shell (Joint Pathology Center [Joint Pathology Center [JPC], 2015]). Once deposited in the liver, eggs can remain viable for months to years (Wright, 1961).
Eggs are released into the environment following either predation, or host death and decomposition. When liver from freshly killed prey is consumed by predators, un-embryonated C. hepatica eggs simply pass via feces into the environment without causing infection. Predators and scavengers serve as paratenic hosts, facilitating egg dissemination and maturation (Redrobe and Patterson-Kane, 2005). Following host death, decomposition and exposure to air and damp soil, liberated eggs embryonate (i.e., become infective) over approximately 30 days, and can overwinter in cold environments (Wright, 1961).
The infection cycle continues when embryonated eggs are consumed by a new mammalian host.
The larvae hatch, penetrate the intestinal mucosa, travel via the portal veins to the liver, invade the parenchyma, and mature over 35–50 days. Adults lay hundreds of eggs, then die 40–59 days later and decompose (Gardiner and Poynton, 1999). Due to their short lifespan, nematodes are less commonly observed on liver histopathology than deposited eggs (Moreira et al., 2013). When present, long, slender C. hepatica nematodes and migration tracks are observed in the parenchyma, often just beneath the hepatic capsule. Infection of other organs is less common (Moreira et al., 2013).
The host range of C. hepatica has been reviewed (Fuehrer et al., 2011, 2014a,b), with only two prior reports of mustelid infection (Solomon and Handley, 1971; Forrester, 1992), and no reports of C. hepatica in marine mammals. Here, we describe C. hepatica infection in a threatened marine mammal from California, the southern sea otter (Enhydra lutris nereis), in which hepatic lesions ranged from mild and incidental, to severe and fatal.
Invertebrate-feeding southern sea otters are not normally parasitized by nematodes, in contrast with their more piscivorous northern counterparts (Margolis et al., 1997). No morphologically similar parasites or eggs have been described from marine mammal livers. Otters are likely infected via ingestion of embryonated C. hepatica eggs reaching the ocean through runoff, or through consumption of eggs in filter-feeding invertebrates, as reported for oocysts and sporocysts of the terrestrial parasites Toxoplasma gondii and Sarcocystis neurona (Arkush et al., 2003; Miller et al., 2008; Michaels et al., 2016). Because C. hepatica can infect humans, our findings could suggest health risks for humans consuming raw or undercooked aquatic invertebrates from areas where rats are abundant, where watersheds are degraded, and where environmental contamination by C. hepatica eggs is high.
Materials and Methods
Stranded southern sea otters from throughout Central California were necropsied by collaborating veterinary pathologists (MM and PD). Necropsy included gross photography and microscopic examination of all major tissues as described (Kreuder et al., 2003). Livers containing suspect C. hepatica eggs, nematodes, and putative parasite migration tracts were further assessed via microscopic examination of formalin-fixed, paraffin-embedded, hematoxylin and eosin (H&E)-stained, 5 μm thick serial sections. The size, location, and morphology of eggs and nematodes were compared with published reports from terrestrial and aquatic mammals, and humans. The stranding location and date, stranding season, sex, and age were assessed to provide insight on the origin and timing of C. hepatica infection. One animal had been implanted with a very high frequency (VHF) radio transmitter and time-depth recorder (TDR) antemortem as part of life history research, so visual resight and electronic monitoring data were compiled by collaborating biologists.
Results
Table 1 summarizes case findings for four sea otters with confirmed or suspected C. hepatica infection.
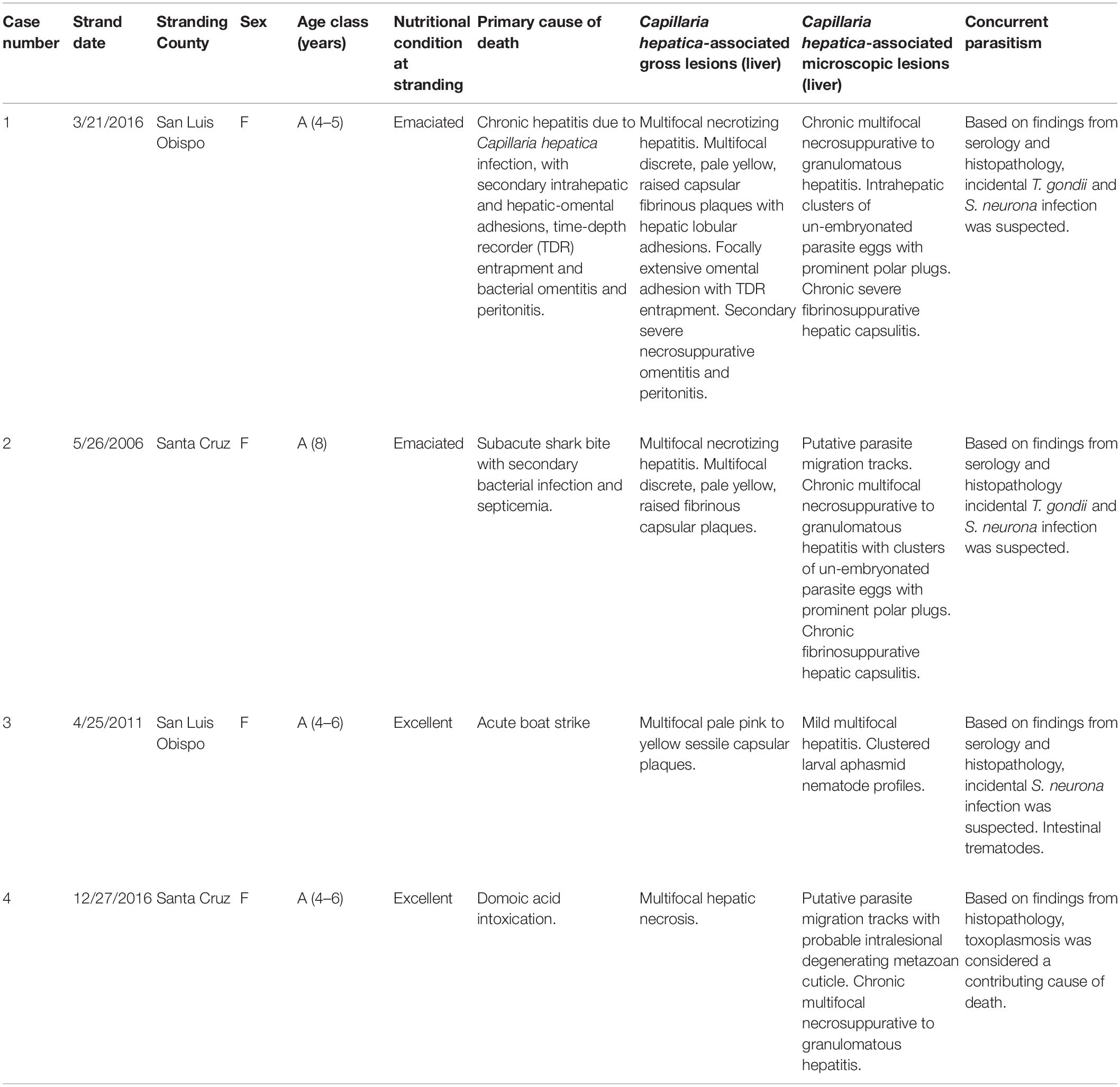
Table 1. Summary of stranding data, gross findings and histopathology for southern sea otters (Enhydra lutris nereis) with putative or confirmed Capillaria hepatica infection.
Case 1 was an emaciated adult female that stranded alive, but died within hours. This animal had been captured, and a VHF radio transmitter and TDR had been surgically implanted in the peritoneal cavity approximately 3.5 years prior to death to facilitate life history research. At the time of capture, the otter weighed 18.5 kg and was in good nutritional condition with no apparent clinical or physical abnormalities. Surgical implantation was uneventful, and parameters on a perioperative complete blood count and blood chemistry panel were unremarkable. Post-capture monitoring revealed that this animal foraged mainly in the vicinity of San Simeon California, and her diet consisted of 46% clams, 18% Cancer spp. crabs, 19% other crab species, and smaller amounts of other prey. Case 1’s TDR data showed body temperature fluctuations suggestive of potential estrus events in 2013 and 2014, but copulation or pupping was not visually confirmed. Data collection by her TDR ceased approximately 18 months prior to death.
At necropsy, the otter’s weight had decreased by 4.5 kg (24%) to 14 kg. Based on mammary, uterine and ovarian morphology, it appeared that abortion had occurred approximately 3 weeks prior to death during the first half of post-implantation pregnancy (Chinn et al., 2016). Although aerobic culture of endometrium grew non-hemolytic Escherichia coli and Vibrio parahaemolyticus, significant endometritis was not noted. Gastric erosions and melena were considered perimortem lesions, reflective of severe stress.
Multifocal pale yellow, raised, 0.5 to >8 cm diameter fibrinous to fibrous plaques were scattered across the hepatic capsule, causing marked contracture of individual hepatic lobes, and extensive adhesions between lobes (Figure 1A). A large adhesion encompassed the visceral surface of the right lateral hepatic lobe and 50% of the omentum, which was bunched into a dense (12 cm × 8 cm × 5 cm) mass (Figure 1A). Adjacent omentum and mesentery were severely congested, thick and opaque. The TDR was recovered from the omental mass, and severe necrosuppurative omentitis with intralesional mixed bacteria was noted adjacent to the TDR entrapment site on histopathology. Aerobic and anaerobic culture of peritoneal exudate yielded E. coli, while V. parahaemolyticus and Clostridium perfringens were identified on fecal culture.
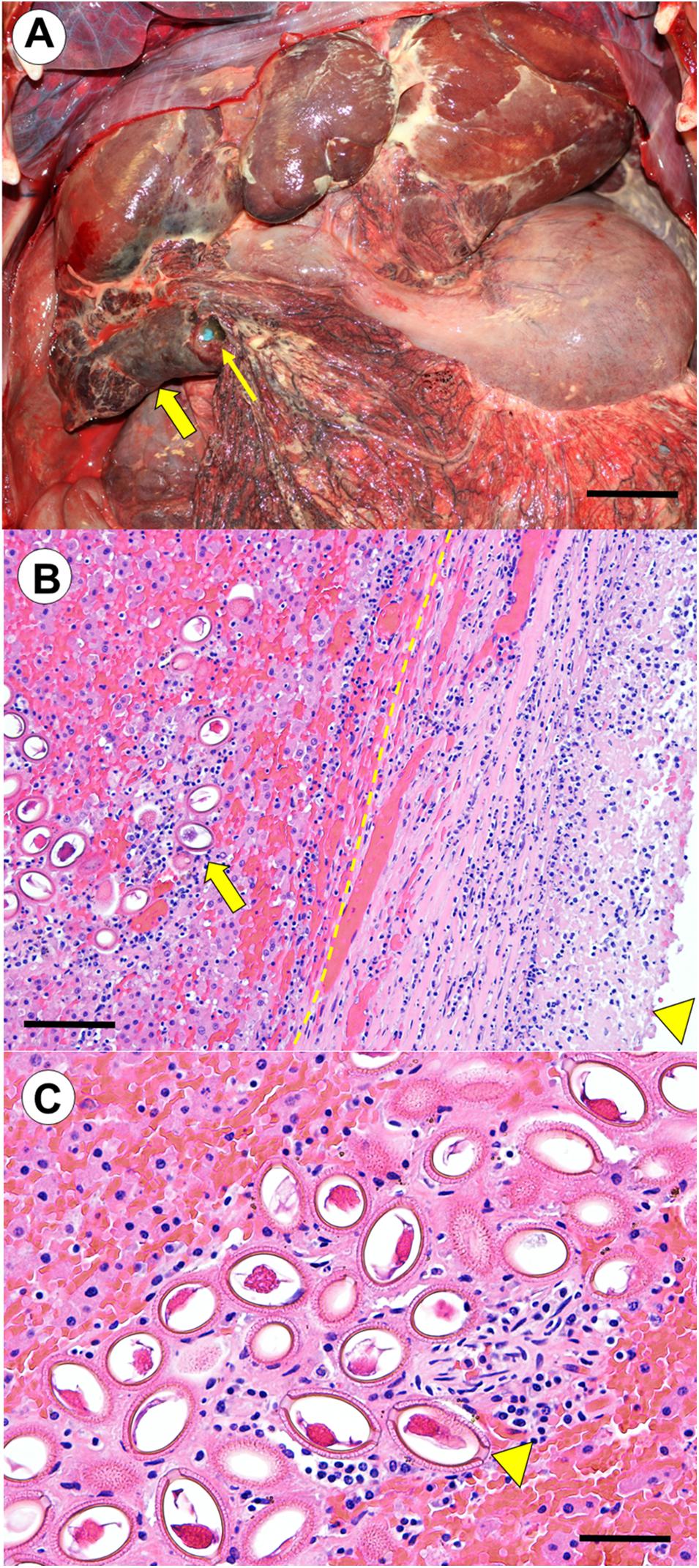
Figure 1. (Case 1): Southern sea otter (Enhydra lutris nereis) with chronic, severe hepatic capillariasis due to Capillaria hepatica, with secondary intraperitoneal instrument entrapment in omentum, leading to chronic bacterial omentitis and peritonitis. (A) Cranioventral peritoneum, showing multifocal fibrinosuppurative to fibrous hepatic lobular adhesions, and a severe, focally extensive hepatic-omental adhesion (broad arrow). The firm, adherent omental mass contained an entrapped time-depth recorder (TDR) that had been surgically implanted in the peritoneal cavity during a capture event 3.5 years previously (a blue lateral edge of the entrapped TDR is indicated by the slender arrow). Also visible is severe secondary fibrinopurulent and necrotizing omentitis and peritonitis. Based on findings from histopathology and bacterial culture, the entrapped TDR appears to have served as a nidus for colonization by bacteria originating from the C. hepatica-associated hepatic lesions (Bar = approximately 6 cm). (B) Transverse histological view of the large hepatic-omental adhesion shown grossly in (A). The hepatic-omental adhesion (arrowhead) is composed of a thick layer of fibrin, granulation tissue, and maturing fibrous connective tissue with admixed neutrophils and granulomatous inflammation. The former hepatic capsular surface (dashed line) lies well below this thick band of surface exudate. Subjacent hepatic parenchyma contains numerous large, elliptical to barrel-shaped nematode eggs (broad arrow), accompanied by mild to moderate pleocellular inflammation (hematoxylin and eosin stain. Bar = 200 μm). (C) Higher magnification view of the nematode eggs. These large, non-pigmented, un-embryonated, barrel-shaped nematode eggs had large polar prominences (arrowhead), and roughened, striated and moderately refractile shell that are characteristic of C. hepatica. Egg masses were encased in a fine fibrovascular stroma admixed with granulomatous inflammation. Inflamed tissue containing parasite eggs were interspersed with large expanses of unaffected hepatic parenchyma with no visible eggs. Linear bands and clusters of parasite eggs were often located immediately subjacent to the hepatic capsule, and were associated with capsular fibrin exudation, inflammation, granulation tissue and fibroplasia, facilitating formation of the severe intrahepatic and hepatic-omental adhesions observed at gross necropsy (hematoxylin and eosin stain. Bar = 100 μm).
On histopathology, multifocal clusters of large (90–100 by 45–55 μm), non-pigmented, un-embryonated, barrel-shaped parasite eggs with large polar prominences at each end, and a roughened, striated and moderately refractile shell were apparent in the subcapsular hepatic parenchyma beneath the omental adhesion (Figure 1B). The hepatic capsule overlying the egg masses was markedly expanded by fibrinosuppurative inflammation, granulation tissue, and fibroplasia (Figure 1B). The egg masses were surrounded by areas of coagulation necrosis and moderate infiltrates of neutrophils, monocytes, macrophages, lymphocytes, eosinophils, hemorrhage, granulation tissue, and fibroplasia (Figure 1C). Egg masses were encased in a fine fibrovascular stroma admixed with granulomatous inflammation, and interspersed with large expanses of unaffected parenchyma (Figure 1C). Low numbers of intramuscular Sarcocystis sp. were also noted on histopathology, but were considered incidental.
Serological testing of serum collected at necropsy showed a high IgG titer to the protozoan parasite T. gondii (10,240; the cutoff for seropositivity is ≥320) (Miller et al., 2002). Mild lymphoplasmacytic meningitis and perivasculitis was observed in the brain on histopathology, accompanied by a focal mild scar in the cerebrum; no parasites compatible with T. gondii were observed on histopathology in any tissue. Serology for the protozoan parasite Sarcocystis neurona was negative (IgG titer 40; the cutoff for seropositivity is presumed to be ≥320) and a single mature intramuscular sarcocyst was observed in the left atrial wall. Protozoan parasite culture, immunohistochemistry, or PCR was not performed. Based on these findings, incidental T. gondii and S. neurona infection was suspected.
Case 2 was an emaciated adult female that died from wound infection and septicemia approximately 2 weeks after a white shark (Carcharodon carcharias) attack. Aerobic bacterial culture of a laceration yielded Streptococcus pyogenes, Streptococcus viridans, Morganella morganii, Vibrio parahaemolyticus, and Shewanella putrefaciens, and heart blood was positive for S. pyogenes and V. parahaemolyticus. Significant health conditions that predated shark bite included severe cardiomyopathy (Kreuder et al., 2003), end-lactation syndrome (ELS) (Chinn et al., 2016), and mild lymphoplasmacytic meningoencephalitis associated with T. gondii infection. Perimortem lesions included dehydration and intestinal melena. Multifocal discrete, pale yellow, raised hepatic capsular plaques and severe necrotizing hepatitis were noted grossly. Several large (90–100 by 45–55 μm), non-pigmented, un-embryonated, barrel-shaped parasite eggs similar to those described for Case 1 were found in hepatic parenchyma subjacent to the capsular plaques (Figure 2A). Parasite eggs were arranged in discrete masses, and were surrounded by large expanses of coagulation necrosis, hemorrhage, inflammation and fibrous connective tissue similar to that described above (Figure 2B). Also present were tiny circular foci of hepatic cavitation with perilesional granulomatous inflammation (putative parasite migration scars) (Figure 2C).
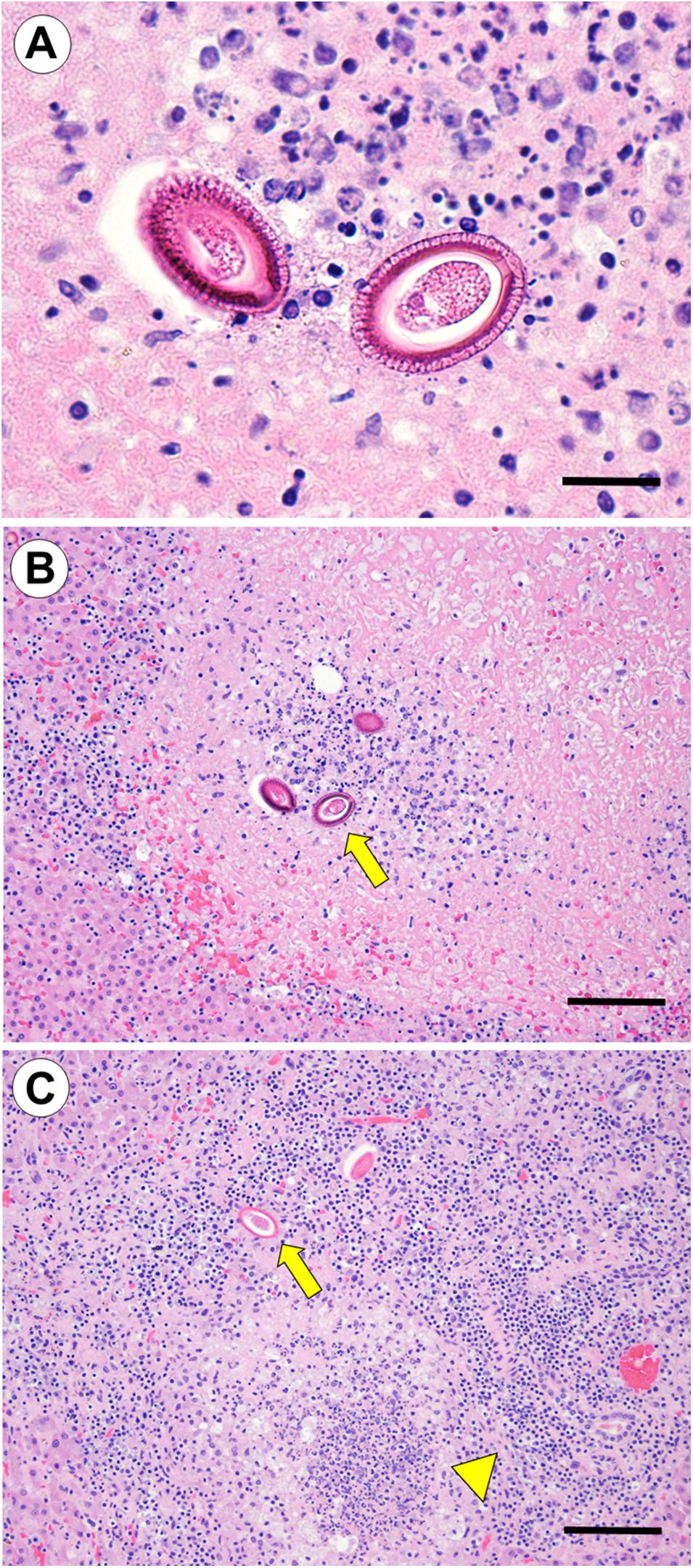
Figure 2. (Case 2): Hepatic capillariasis in a southern sea otter (Enhydra lutris nereis) that died from shark bite with secondary bacterial infection. (A) Microscopic examination of hepatic parenchyma subjacent to grossly apparent hepatic fibrinous to fibrous capsular plaques revealed multifocal clusters of large (90–100 × 45–55 μm), non-pigmented, un-embryonated, barrel-shaped parasite eggs similar to those described for Case 1 above (hematoxylin and eosin stain. Bar = 50 μm). (B) Lower magnification view of the same parasite eggs shown in (A): the parasite eggs were arranged in discrete masses, and were surrounded by areas of coagulation necrosis, hemorrhage, inflammation, and fibrous connective tissue (hematoxylin and eosin stain. Bar = 200 μm). (C) Small circular, linear, or serpiginous bands of hepatic cavitation with perilesional granulomatous inflammation were suggestive of parasite migration tracks (arrowhead). Parasite eggs were often located along the outer edge of the cavitated lesions (broad arrow) (hematoxylin and eosin stain. Bar = 200 μm).
Serological testing revealed a low positive IgG titer to T. gondii (1,280), and this parasite was isolated from brain tissue in cell culture. Brain histopathology showed mild lymphoplasmacytic meningitis, perivasculitis and encephalitis with a single mature T. gondii tissue cyst; the tissue cyst was not present in recuts and immunohistochemistry was negative. Serology found a high IgG titer (10,240) to S. neurona, but no parasites were observed on histopathology or immunohistochemistry. No PCR was attempted. Based on these findings, incidental T. gondii and S. neurona infection was suspected.
Case 3 was an adult female in excellent nutritional condition that died due to boat strike, resulting in multiple limb, rib, and sternal fractures, a lacerated aorta, hemothorax and diaphragmatic rupture, with herniation of portions of the liver and intestines into the thorax. All liver lobes were fractured and hemorrhagic, limiting assessment of lesions that could be attributable to C. hepatica infection, but multiple 1–3 cm diameter pale pink to yellow, sessile capsular foci were noted on histopathology. No bacterial culture was performed.
Serial liver sections contained multiple cross-sectional, oblique and partial longitudinal larval nematode profiles in the parenchyma (Figure 3A). The parasites were 105–120 μm diameter and coelomate with a cuticle, hypodermal bands, a prominent stichosome, and single round to oval intestinal and gonadal profiles (Figures 3B,C). The gonads were small and immature, with no visible ova or spermatozoa. Parasite profiles were clustered within a localized region of subcapsular parenchyma encompassing approximately 50 μm of serial sections. Although small foci of coagulation necrosis and mild suppurative hepatitis were apparent near the parasites, minimal necrosis, host inflammatory response or fibrosis were visible immediately adjacent to the nematodes, and no eggs were observed. Low numbers of intramuscular Sarcocystis sp. were noted on histopathology, but were considered incidental.
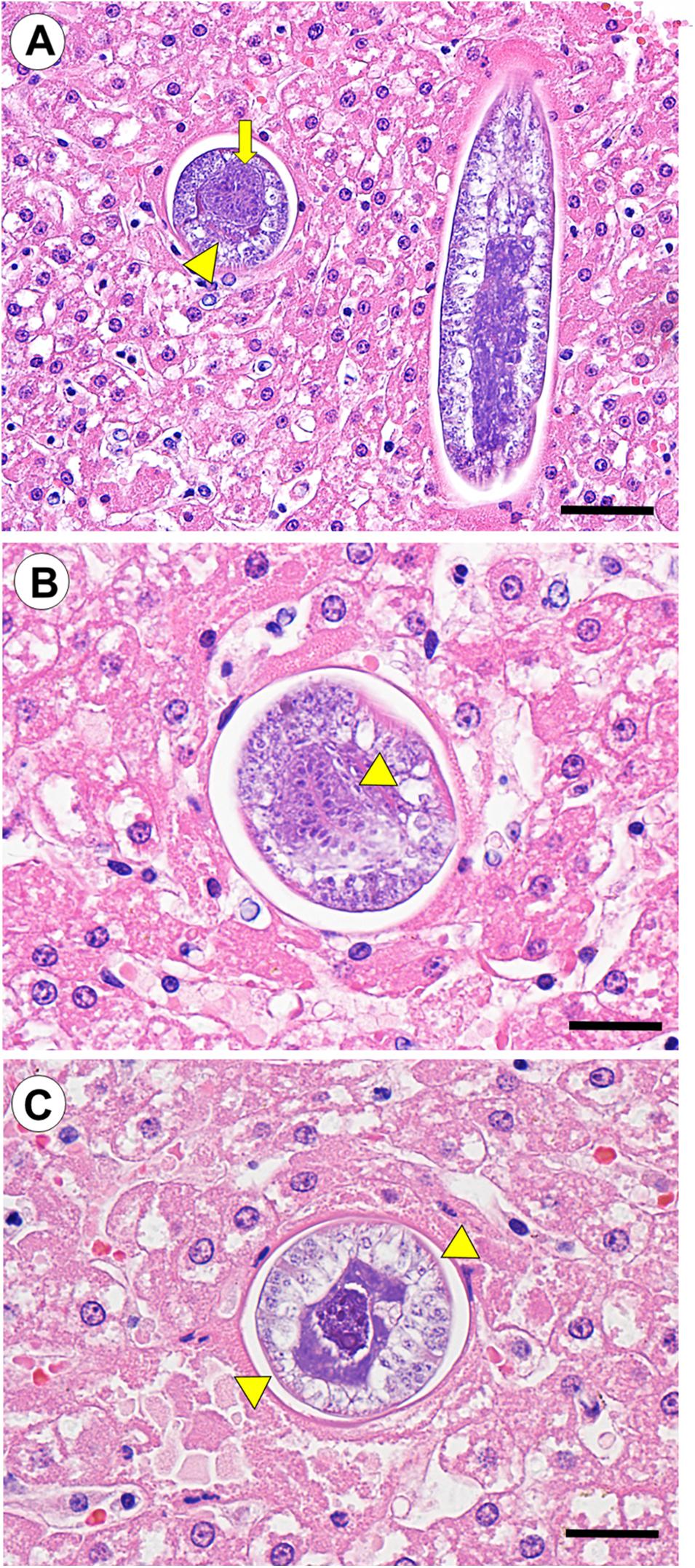
Figure 3. (Case 3): Larval nematode profiles (putative C. hepatica larvae) in the hepatic parenchyma of a southern sea otter (Enhydra lutris nereis). (A) Serial 5 μm thick tissue sections revealed multiple cross-sectional, oblique, and longitudinal nematode profiles that were clustered in a small area of the hepatic parenchyma. Cross-sections of these parasites showed single round to oval intestinal (broad arrow) and gonadal (arrowhead) profiles (hematoxylin and eosin stain. Bar = 100 μm). (B,C) The nematodes were 105–120 μm diameter and coelomate with a cuticle, a prominent stichosome surrounding the esophagus (indicated by arrowhead in B), and paired hypodermal bands (indicated by arrowheads in C); these morphological characteristics are indicative of C. hepatica. The gonads were small with no visible ova or spermatozoa, and no eggs were visible in the adjacent hepatic parenchyma, suggesting that the parasites were immature (hematoxylin and eosin stain. Bar = 50 μm for both B,C).
Serology was negative (IgG titers <40) for both T. gondii and S. neurona. Sparse mature sarcocysts were observed in skeletal myofibers without appreciable inflammation. No PCR or parasite isolation was attempted. Based on these findings, this otter appeared to have incidental S. neurona infection, and was not infected with T. gondii.
Case 4 was a lactating adult female in excellent nutritional condition that was euthanized due to severe neurological disease. At necropsy, a 778 gm, fully haired, meconium-smeared male fetus was recovered from the uterus. On histopathology the most significant lesion was hippocampal necrosis consistent with domoic acid intoxication; maternal and fetal urine were biochemically positive for domoic acid at 317 and 312 ppb, respectively. As with the previous cases, perimortem gastric erosions and melena were acute comorbidities. Multifocal subcapsular hepatic necrosis was randomly distributed within all lobes.
Serial sectioning of liver contained multifocal to coalescing serpiginous bands of necrosuppurative hepatitis, mild tissue cavitation and granulomatous inflammation, especially in the subcapsular hepatic parenchyma (putative parasite migration tracks) (Figure 4A). Multifocal discontinuous, linear to round fragments of eosinophilic hyaline and often striated material resembling partially degenerated nematode cuticle were observed at the center of some lesions (presumptive degenerating metazoan parasites) (Figures 4B,C). No parasite eggs were observed on multiple serial sections.
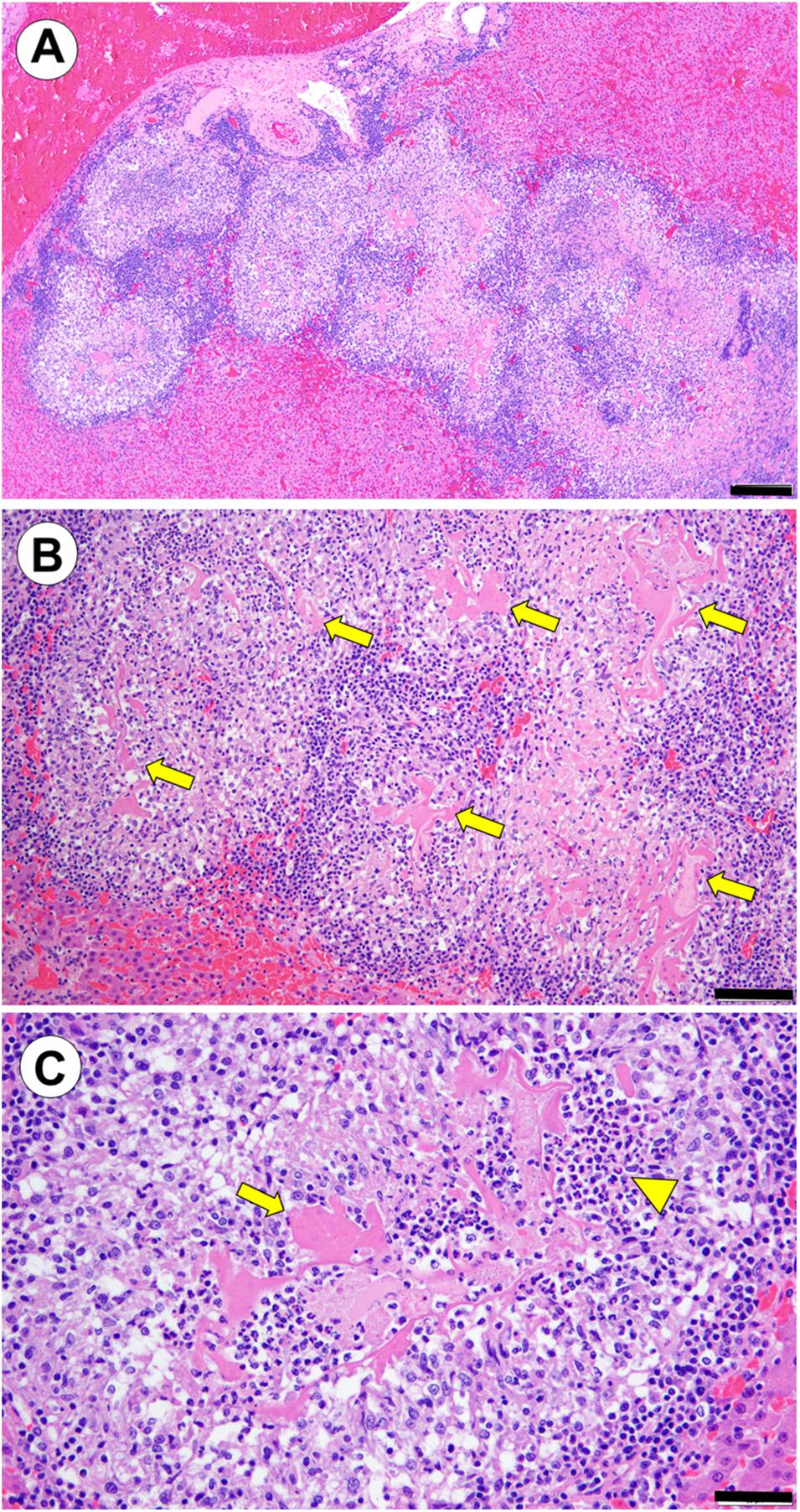
Figure 4. (Case 4): Probable hepatic metazoan parasite migration tracks with intralesional degenerating metazoan cuticle. (A) Linear to serpiginous tracts of necrosuppurative hepatitis, mild tissue cavitation and granulomatous inflammation in the subcapsular hepatic parenchyma (putative parasite migration tracks) (hematoxylin and eosin stain. Bar = 200 μm). (B) Higher magnification view of affected liver. Multifocal discontinuous, linear to round fragments of eosinophilic hyaline, variably striated material (putative degenerating metazoan parasite cuticle) are visible at the center of some lesions (arrows) (hematoxylin and eosin stain. Bar = 100 μm). (C) In this high magnification view of a single inflammatory nodule, putative fragments of metazoan cuticle (arrow) are surrounded by dense clusters of eosinophils (arrowhead), and an outer rim of granulomatous inflammation (hematoxylin and eosin stain. Bar = 50 μm).
On histopathology, multifocal, mild to severe lymphoplasmacytic meningoencephalomyelitis and gliosis was associated with sparse mature T. gondii tissue cysts. No sarcocysts were observed in any tissue and cell culture was negative for all protozoa. Protozoal serology, immunohistochemistry, and PCR were not performed. Toxoplasma gondii-associated meningoencephalomyelitis was considered a contributing cause of death.
Discussion
In the current study, three confirmed, and one suspect case of hepatic capillariasis were identified in wild sea otters stranding along the central California coast. Interestingly, all were adult females (Table 1). Given our small sample, this age and sex bias could represent random chance, or could suggest enhanced risk of C. hepatica infection for sexually mature females. Prior studies report distinct differences in sea otter behavior by sex; adult females have smaller home ranges and greater site fidelity than adult males (Loughlin, 1980; Jameson, 1989; Riedman and Estes, 1990). Females often feed at shallower depths when compared with males, and seek shorelines with calm water and higher kelp density when raising pups (Sandgren et al., 1973; Bodkin et al., 2006; Nicholson et al., 2018). These areas could correspond spatially to a higher density of introduced rats, a key source of C. hepatica infection (Fuehrer et al., 2014a), greater risk of exposure to parasite-contaminated freshwater runoff, and/or recurrent parasite introduction due to infected rodents escaping from moored ships. Infected sea otters stranded at several locations along the Central California coast spanning >250 km, suggesting multiple areas of C. hepatica exposure.
A possible seasonal bias was also noted, with all C. hepatica-infected sea otters stranding during or immediately following the period of peak annual precipitation and freshwater runoff into California coastal marine waters, and the period of highest prevalence and density of C. hepatica infection in Rattus sp. reservoir populations (Conlogue et al., 1979; Rothenburger et al., 2014). A late spring peak has also been reported for sea otter death due to other terrestrial parasites that are spread through dispersal of environmentally resistant stages, such as S. neurona sporocysts (Miller et al., 2008; Shapiro et al., 2012).
Differential diagnoses for gross, microscopic, and clinical pathology findings associated with C. hepatica include hepatic infection or aberrant migration by other nematodes (e.g., Ascaris, Toxocara, Toxascaris, or Baylisascaris spp.), cestodes (e.g., Echinococcus and Mesocestoides sp.), and flukes (e.g., Fasciola hepatica and Fascioloides magna). The range of hepatic macroparasites that could be confused with C. hepatica infection has been reported (Joint Pathology Center [JPC], 2015).
Localization of infection to the hepatic parenchyma, and the unique morphology of the parasites and eggs on histopathology helps distinguish C. hepatica from infection by other parasites. The nematodes are slender, with narrow anterior, and swollen posterior ends (Yadav et al., 2016). Adult C. hepatica average 100–150 μm diameter, and are pseudocoelomate with a cuticle, polymyarian-coelomyarian musculature, and single, round to oval gastrointestinal and gonadal profiles on cross-section (Gardiner and Poynton, 1999; Eberhard, 2014; Joint Pathology Center Joint Pathology Center [JPC], 2015). Other distinctive characteristics that can help distinguish C. hepatica from other nematodes include absence of lateral cords, presence of hypodermal bands (Figure 3C), and presence of a unique glandular structure (stichosome) surrounding the esophagus that is composed of distinctive basophilic cells (stichocytes) (Figure 3B) (Gardiner and Poynton, 1999). The stichosome is diagnostic of C. hepatica (Gardiner and Poynton, 1999; Yadav et al., 2016). Other morphological features include a long esophagus that spans half of the female’s body, and one third for males (Yadav et al., 2016) and a lack of sensory papillae (i.e., phasmids) on their caudal end. Although phasmids are not identifiable histologically, when observed in intact worms they can help differentiate C. hepatica from phasmid nematodes (e.g., strongyles, ascarids, oxyurids, rhabditoids, and spirurids) (Joint Pathology Center [JPC], 2015).
Capillaria hepatica eggs are also distinctive based on their distribution in hepatic parenchyma and morphology; eggs are approximately 66 μm × 36 μm, barrel-shaped, non-pigmented, and un-embryonated, with large polar prominences, and a roughened, striated and moderately refractile shell containing numerous minipores (Yadav et al., 2016) (Figures 1C, 2A). Although some other nematode eggs are elliptical and bioperculate, they differ in size, shape and morphological characteristics, may be embryonated or un-embryonated in tissue, and are unlikely to be observed in dense clusters in the hepatic parenchyma. Trichuris trichiura eggs resemble those of C. hepatica, but differ in size, morphology and tissue location (Cochrane et al., 1957; Li et al., 2010). Although aberrant larval migration is possible, T. trichiura and ascarid eggs are unlikely to be observed in hepatic parenchyma.
Finally, the peri-parasitic, egg-associated and hepatic capsular fibrin exudation and fibrosis that often accompanies C. hepatica infection is distinctive. The host response to C. hepatica can be severe and fatal, forming adhesions between liver lobes, or from the liver to other abdominal structures (Otto et al., 1954). This lesion was apparent for Case 1 (Figure 1A), and the resulting hepatic-omental adhesion with TDR entrapment was the underlying cause of death. The tightly adherent omental pocket containing the TDR appeared to serve as a nidus for chronic secondary bacterial peritonitis, with the bacteria likely originating from the adjacent necrotic hepatic lesions. A pattern of parenchymal fibrosis in association with discrete clusters of subcapsular, bioperculate nematode eggs in the hepatic parenchyma is characteristic of C. hepatica infection in mammals (Joint Pathology Center [JPC], 2015).
In addition to visualization of parasites and eggs, other microscopic lesions can suggest C. hepatica infection, including elongated, linear to serpiginous, tracks that are variably necrotic, cavitated and/or inflamed (putative parasite migration tracts) (Figure 4A), degenerating nematodes associated with these tracts (Figures 4B,C), and multifocal discrete areas of subcapsular hepatitis and fibrosis, with relative sparing of adjacent tissue. The pattern of hepatic fibrosis can also be distinctive: Rats infected with C. hepatica commonly exhibit bridging fibrosis, where hepatic portal tracts are connected to adjacent portal tracts (or rarely centrilobular spaces) by thin strands of connective tissue (Joint Pathology Center [JPC], 2015). Interestingly, C. hepatica-associated septal fibrosis is not necessarily preceded by hepatic necrosis or chronic inflammation (Ferreira and Andrade, 1993; Gaban et al., 2010). In severely affected areas (especially in rats), the hepatic architecture can be reminiscent of porcine liver.
Lymphoplasmacytic, and to a lesser extent, eosinophilic infiltration can also be apparent in portal tracts and near centrilobular veins, and aggregates of eggs may be surrounded by granulomatous inflammation or fibrosis. Liver damage induced by C. hepatica in rodents has been used to study the extensive regenerative capacity of mammalian liver (Santos et al., 2007), and for testing antifibrotic drugs (De Souza et al., 2000). Yi et al. (2010) also noted associations between C. hepatica-induced hepatitis and pulmonary arteriolar hypertrophy in rats, and suggested this model to clarify mechanisms of porto-pulmonary hypertension in humans.
Rodents in the genus Rattus are considered primary C. hepatica hosts (Resendes et al., 2009), and the main source of parasite contamination in urban environments (Redrobe and Patterson-Kane, 2005). In Maryland, Norway rats were far more frequently infected (87.4%) than sympatric house mice (5.4%), and no lesions were observed in meadow voles or white-footed mice (Childs et al., 1988). Rats from urbanized locations had a higher prevalence and intensity of infection, when compared with parkland-associated rats, and higher rat population density and body mass were associated with increased C. hepatica prevalence and lesion severity. Risk factors for C. hepatica infection in feral Norway rats from Vancouver, Canada included older age, sampling during fall and winter, and presence of cutaneous bite wounds (Rothenburger et al., 2014). Higher prevalence of C. hepatica infection in rats sampled during winter was also reported in Connecticut, United States (Conlogue et al., 1979). In studies of C. hepatica transmission ecology, the mean egg count per rat liver was 457,783 (range 11,270–1,400,000) (Farhang-Azad, 1977a, b). Rapid population turnover and cannibalism are considered primary drivers of transmission within rat populations (Farhang-Azad, 1977a, b; Rothenburger et al., 2014).
Other communally living rodents with a high prevalence of C. hepatica infection include muskrats, eastern wood rats (Fuehrer et al., 2014a), and captive prairie dogs (Landolfi et al., 2003; Redrobe and Patterson-Kane, 2005). Along with a high prevalence of C. hepatica infection in native aquatic rodents (i.e., muskrats), cotton rats sampled from freshwater marshes in Florida had higher prevalence of C. hepatica infection (30%), compared with those from saltwater marshes (12%) and upland habitat (5%) (Kinsella, 1974). Capillaria hepatica has also been reported from <140 non-rodent mammals, including humans (Berentsen et al., 2015). The only prior reports of C. hepatica infection in North American mustelids were in a river otter (Lontra canadensis) in Florida (Forrester, 1992), and two long-tailed weasels (Mustela frenata) from Virginia (Solomon and Handley, 1971).
Through 2011, 72 confirmed cases of C. hepatica-associated human hepatic capillariasis were reported worldwide, with the majority affecting children (Fuehrer et al., 2011). Infection was associated with pyrexia, chronic eosinophilia, hepatomegaly, eosinophilic granulomatous hepatitis, hepatic fibrosis, cirrhosis, and rarely, death (Otto et al., 1954; Cochrane et al., 1957; Li et al., 2010). Also reported were abdominal pain, anorexia, emaciation, ascites, cholelithiasis (Ferreira and Andrade, 1993), anemia, and hypergammaglobulinemia (Yadav et al., 2016). Humans can be infected following ingestion of embryonated eggs in food, water, or soil. Residing in areas with poor hygiene, close contact with rats or other rodents, pica, and extensive soil contact are risk factors for human infection (Cochrane et al., 1957; Yadav et al., 2016; Walker et al., 2017). Sub-standard housing conditions may facilitate rat infection rates of up to 85%, increasing risk of human infection (Walker et al., 2017). Common non-native rodents along the central California coast include roof rats (Rattus rattus), Norway rats (Rattus norvegicus) and Eastern fox squirrels (Sciurus niger). Common native rodents include California ground squirrels (Spermophilus beecheyi), Botta’s pocket gophers (Thomomys bottae), California voles (Microtus californicus), western harvest mice (Reithrodontomys megalotis) and dusky-footed woodrats1 (Neotoma fuscipes).
Hepatic capillariasis is often diagnosed in humans by finding C. hepatica nematodes and eggs in biopsy or autopsy specimens, but serodiagnostic tests are now available (Juncker-Voss et al., 2000; Assis et al., 2004; Huang et al., 2001). Because infections are non-patent, detection of C. hepatica eggs in feces indicates spurious passage of ingested eggs, not infection, and has been associated with consumption of wild game (Foster and Johnson, 1939). Options for treating clinical C. hepatica infection in humans include thiobendazole, albendazole, and ivermectin (Yadav et al., 2016).
The potential for parasite co-infection to facilitate C. hepatica infection, or cause more severe disease should be considered. Recent reports have described fatal co-infection of a Korean squirrel (Tamias sibiricus) (Carrasco et al., 2006) and a ring-tailed lemur (Lemur catta) (Siskos et al., 2015) with C. hepatica and T. gondii. In the current study, all four sea otters were concurrently infected with T. gondii, Sarcocystis sp., and/or intestinal trematodes (Table 1). Given the very high prevalence of T. gondii and S. neurona infection in southern sea otters (Miller et al., 2017), and the potential for polyparasitism to cause more severe disease in marine mammals (Gibson et al., 2011), the potential for parasite interactions should be considered as part of future assessment of C. hepatica infections.
Gross lesions suggestive of C. hepatica infection were noted for both cases where C. hepatica eggs were observed on histopathology, but were less clear for the cases where only larval or degenerating worms were present. Although comparatively uncommon, the prevalence of C. hepatica infection in southern sea otters is probably much higher than the four cases reported here. Because the hepatic lesions are often multifocal and are interspersed with areas of normal tissue, C. hepatica infection is very easy to miss unless suspect lesions are collected and examined microscopically. The capsular lesions could also be easily confused with other common peritoneal diseases of sea otters, including acanthocephalan or bacterial peritonitis (Kreuder et al., 2003). Future efforts should include microscopic examination of hepatic parenchyma beneath areas of capsular fibrin deposition and fibrosis. Efforts to more accurately characterize the prevalence of infection could include digesting liver tissue to recover parasite eggs from patchy areas of deposition, and serological testing. It would also be interesting to screen coastal rodents, soil, and runoff for C. hepatica.
Similar to the land-based parasites T. gondii and S. neurona (Miller et al., 2008, 2010), C. hepatica eggs may be concentrated and transmitted through filter-feeding marine invertebrates that serve as southern sea otter prey, which may also pose an unrecognized public health risk for people who consume these species. Although C. hepatica can be transmitted to humans via ingestion of embryonated eggs, risks from handing infected sea otters are negligible unless the carcasses have significantly decomposed, allowing the eggs to become infective. However, because the most likely source of C. hepatica infection for sea otters is consuming water or prey contaminated with embryonated eggs originating from predated or dead, decomposing, land-based rats, sea otter parasitism could suggest un-recognized human health risks for acquiring C. hepatica infection from contaminated water, or raw/undercooked filter-feeding marine and estuarine invertebrates; this pathway for marine wildlife and human exposure to C. hepatica merits further investigation.
Data Availability Statement
All datasets generated for this study are included in the article/supplementary material.
Ethics Statement
Ethical review and approval was not required for the animal study because the study was performed using dead southern sea otters submitted for necropsy to the California Department of Fish and Wildlife as per federal MOU.
Author Contributions
MAM directed the case investigations, conducted gross necropsy and histopathology, completed a literature review on C. hepatica and wrote the manuscript. PD conducted necropsy and histopathology for case 4, contributing photomicrographs and a detailed case history for case 4, and helped substantially with manuscript editing and completion. ED and FB assisted with gross necropsies, case data compilation and review, coordinated completion of serial sections for livers of affected sea otters, and helped with manuscript preparation. MS, JT, MM, and HH contributed live capture and antemortem data for cases in the study and helped with manuscript editing. CG provided expertise to facilitate parasite and egg identification in tissue sections, optimized text related to histological identification of C. hepatica, ensured that key structural features in the figures were labeled correctly, and provided substantial input during manuscript editing and completion.
Funding
This work was funded by the California Department of Fish and Wildlife.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank volunteers and staff at the California Department of Fish and Wildlife – Marine Wildlife Veterinary Care and Research Center, the Monterey Bay Aquarium, and The Marine Mammal Center for their efforts to recover sick and dead-stranded animals along the central California Coast, and for allowing access to archival records and tissue samples. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the United States Government.
Footnotes
References
Arkush, K. D., Miller, M. A., Leutenegger, C. M., Gardner, I. A., Packham, A. E., Heckeroth, A. R., et al. (2003). Molecular and bioassay-based detection of Toxoplasma gondii oocyst uptake by mussels (Mytilus galloprovincialis). Int. J. Parasitol. 33, 1087–1097. doi: 10.1016/S0020-7519(03)00181-4
Assis, B. C. A., Cunha, L. M., Baptista, A. P., and Andrade, Z. A. (2004). A contribution to the diagnosis of Capillaria hepatica infection by indirect immunofluorescence test. Mem. Inst. Oswaldo Cruz. 99, 173–177. doi: 10.1590/S0074-02762004000200010
Berentsen, A. R., Vogt, S., Guzman, A. N., Vice, D. S., Pitt, W. C., Shiels, A. B., et al. (2015). Capillaria hepatica infection in black rats (Rattus rattus) on Diego Garcia, British Indian Ocean Territory. J. Vet. Diagn. Investig. 27, 241–244.
Bodkin, J. L., Esslinger, G. G., and Monson, D. H. (2006). Foraging depths of sea otters and implications to coastal marine communities. Mar. Mamm. Sci. 20, 305–321. doi: 10.1111/j.1748-7692.2004.tb01159.x
Carrasco, L., Raya, A. I., Núñez, A., Gómez-Laguna, J., Hernández, S., and Dubey, J. P. (2006). Fatal toxoplasmosis and concurrent Calodium hepaticum infection in Korean squirrels (Tanias sibericus). Vet. Parasitol. 137, 180–183. doi: 10.1016/j.vetpar.2005.11.026
Childs, J. E., Glass, G. E., and Korch, G. W. Jr. (1988). The comparative epizootiology of Capillaria hepatica (Nematoda) in urban rodents from different habitats of Baltimore, Maryland. Can. J. Zool. 66, 2769–2775. doi: 10.1139/z88-404
Chinn, S. M., Miller, M. A., Tinker, M. T., Staedler, M. M., Batac, F. I., Dodd, E. M., et al. (2016). The high cost of motherhood: end-lactation syndrome in southern sea otters (Enhydra lutris nereis) on the central California coast, USA. J. Wildl. Dis. 52, 307–318. doi: 10.7589/2015-06-158
Cochrane, J. C., Sagokin, L., and Wilcocks, M. G. (1957). Capillaria hepatica Infection in Man. A Syndrome of Extreme Eosinophilia, Hepatomegaly and Hyperglobulinaemia. South Af. Med. J. 31, 751–755.
Conlogue, G., Foreyt, W., Adess, M., and Levine, H. (1979). Capillaria hepatica (Bancroft) in select rat populations of Hartford, Connecticut, with possible public health implications. J. Parasitol. 65, 105–108. doi: 10.2307/3280211
De Souza, M. M., Silva, L. M., Barbosa, A. A., De Oliveira, I. R., Paraná, R., and Andrade, Z. A. (2000). Hepatic capillariasis in rats: a new model for testing antifibrotic drugs. Braz. J. Med. Biol. Res. 33, 1329–1334. doi: 10.1590/S0100-879X2000001100011
Eberhard, M. (2014). “Histopathologic diagnosis,” in Georgi’s Parasitology for Veterinarians, ed. D. Bowman (St. Louis, Missouri: Elsevier-Saunders).
Farhang-Azad, A. (1977a). Ecology of Capillaria hepatica (Bancroft 1893) (Nematoda). 1. Dynamics of infection among Norway rat populations of the Baltimore Zoo, Baltimore, Maryland. J. Parasitol. 63, 117–122. doi: 10.2307/3280114
Farhang-Azad, A. (1977b). Ecology of Capillaria hepatica (Bancroft 1893) (Nematoda). II. Egg-releasing mechanisms and transmission. J. Parasitol. 63, 701–706. doi: 10.2307/3279576
Ferreira, L. A., and Andrade, Z. A. (1993). Capillaria hepatica: a cause of septal fibrosis of the liver. Mem. Inst. Oswaldo Cruz. 88, 441–447. doi: 10.1590/S0074-02761993000300015
Forrester, D. (1992). “Mustelidae,” in Parasites and Diseases of Wild Mammals in Florida, ed. D. Forrester (Gainesville, FL: Univ. Press of Florida).
Foster, A. O., and Johnson, C. M. (1939). An explanation for the occurrence of Capillaria hepatica ova in human faeces suggested by the finding of three new hosts used as food. Trans. R. Soc. Trop. Med. Hyg. 32, 639–644. doi: 10.1016/S0035-9203(39)90027-1
Fuehrer, H. P. (2014a). An overview of the host spectrum and distribution of Calodium hepaticum (syn. Capillaria hepatica): Part 1—Muroidea. Springer Berlin Heidelberg, Berlin/Heidelberg. Parasitol. Res. 113, 619–640. doi: 10.1007/s00436-013-3691-x
Fuehrer, H. P. (2014b). An overview of the host spectrum and distribution of Calodium hepaticum (syn. Capillaria hepatica): Part 2 - Mammalia (excluding Muroidea). Parasitol. Res. 113, 641–651. doi: 10.1007/s00436-013-3692-9
Fuehrer, H. P., Igel, P., and Auer, H. (2011). Capillaria hepatica in man-an overview of hepatic capillariosis and spurious infections. Parasitol. Res. 109, 969–979. doi: 10.1007/s00436-011-2494-1
Gaban, L., Ramos, C. D. L., Barbosa Júnior, A. A., Souza, M. M. D., and Andrade, Z. D. A. (2010). Dynamics of Capillaria hepatica-induced hepatic septal fibrosis in rats. Rev. Soc. Bras. Med. Trop. 43, 643–646. doi: 10.1590/S0037-86822010000600008
Gardiner, C. H., and Poynton, S. L. (1999). An Atlas of Metazoan Parasites in Animal Tissues. Washington, DC: American Registry of Pathology.
Gibson, A. K., Raverty, S., Lambourn, D. M., Huggins, J., Magargal, S. L., and Grigg, M. E. (2011). Polyparasitism is associated with increased disease severity in Toxoplasma gondii-infected marine sentinel species. PLoS Negl. Trop. Dis. 5:e001142. doi: 10.1371/journal.pntd.0001142
Huang, H., Ling, H., Liang, S., Xing, W., and Pan, C. (2001). Diagnosis of Experimental rat hepatic capillariasis by ELISA. Wenzhou Yixueyuan Xuebao 31, 299–302.
Jameson, R. J. (1989). Movements, home range, and territories of male sea otters off central California. Mar. Mamm. Sci. 5, 159–172. doi: 10.1111/j.1748-7692.1989.tb00330.x
Joint Pathology Center [JPC] (2015). “Veterinary Pathology Services,” in Procedings of the Enhanced Wednesday Slide Conference Online, Conference: 05 – 2015 Case: 03 (JPC 4065817), liver – Rat. Available online at: https://www.askjpc.org/wsco/wsc_showconference.php?id=861 (accessed January 5, 2018).
Juncker-Voss, M., Prosl, H., Lussy, H., Enzenberg, U., Auer, H., and Nowotny, N. (2000). Serological detection of Capillaria hepatica by indirect immunofluorescence assay. J. Clin. Microbiol. 38, 431–433.
Kinsella, J. M. (1974). Comparison of Helminth Parasites of the Cotton Rat, Sigmodon Hispidus, from Several Habitats in Florida. Washington, DC: American Museum novitates.
Kreuder, C., Miller, M. A., Jessup, D. A., Lowenstein, L. J., Harris, M. D., Ames, J. A., et al. (2003). Patterns of mortality in southern sea otters (Enhydra lutris nereis) from 1998-2001. J. Wildl. Dis. 39, 495–509. doi: 10.7589/0090-3558-39.3.495
Landolfi, J. A., Karim, B. O., Poynton, S. L., and Mankowski, J. L. (2003). Hepatic Calodium hepaticum (Nematoda) infection in a zoo colony of black-tailed prairie dogs (Cynomys ludovicianus). J. Zoo Wildl. Med. 34, 371–374. doi: 10.1638/02-095
Li, C. D., Yang, H. L., and Wang, Y. (2010). Capillaria hepatica in China. World J. Gastroenterol. 16:698. doi: 10.3748/wjg.v16.i6.698
Loughlin, T. R. (1980). Home range and territoriality of sea otters near Monterey, California. J. Wildl. Manag. 44, 576–582. doi: 10.2307/3808005
Margolis, L., Johnson, S. C., McDonald, T. E., Kent, M. L., and Blaylock, R. B. (1997). Helminth Parasites of Sea Otters (Enhydra lutris) from prince william sound, Alaska: comparisons with other populations of Sea Otters and comments on the origin of their parasites. J. Helminthol. Soc. Washingt. 64, 161–168.
Michaels, L., Rejmanek, D., Aguilar, B., Conrad, P., and Shapiro, K. (2016). California mussels (Mytilus californianus) as sentinels for marine contamination with Sarcocystis neurona. Parasitology 143, 762–769. doi: 10.1017/S0031182016000354
Miller, M. A., Conrad, P. A., Harris, M., Hatfield, B., Langlois, G., Jessup, D. A., et al. (2010). A protozoal-associated epizootic impacting marine wildlife: mass-mortality of southern sea otters (Enhydra lutris nereis) due to Sarcocystis neurona infection. Vet. Parasitol. 172, 183–194. doi: 10.1016/j.vetpar.2010.05.019
Miller, M. A., Gardner, I. A., Packham, A. E., Mazet, J. K., Hanni, K. D., Jessup, D. A., et al. (2002). Evaluation of an indirect fluorescent antibody test (IFAT) for demonstration of antibodies to Toxoplasma gondii in the sea otter (Enhydra lutris). J. Parasitol. 88, 594–599. doi: 10.2307/3285456
Miller, M. A., Miller, W. A., Conrad, P. A., James, E. R., Melli, A. C., Leutenegger, C. M., et al. (2008). Type X Toxoplasma gondii in a wild mussel and terrestrial carnivores from coastal California: new linkages between terrestrial mammals, runoff and toxoplasmosis of sea otters. Int. J. Parasitol. 38, 1319–1328. doi: 10.1016/j.ijpara.2008.02.005
Miller, M. A., Moriarty, M. E., Dodd, E. M., Burgess, T., Tinker, M. T., Batac, F. I., et al. (2017). The Dead do Tell Tales: Investigating Sea Otter Mortality Patterns (1998-2012). Oakland, CA: California Coastal Conservancy.
Moreira, V. L. C., Giese, E. G., Silva, D. C. B. D., Melo, F. T. D. V., Furtado, A. P., Maldonado, A. Jr. et al. (2013). Calodium hepaticum (Nematoda: Capillariidae) in synanthropic rodents (Rattus norvegicus and Rattus rattus) in Eastern Amazonia. Rev. Bras. Parasitol. Vet. 22, 265–269. doi: 10.1590/S1984-29612013000200046
Mowlavi, G., Kacki, S., Dupouy-Camet, J., Mobedi, I., Makki, M., Harandi, M. F., et al. (2014). Probable hepatic capillariosis and hydatidosis in an adolescent from the late Roman period buried in Amiens (France). Parasite 21:9. doi: 10.1051/parasite/2014010
Nicholson, T. E., Mayer, K. A., Staedler, M. M., Fujii, J. A., Murray, M. J., Johnson, A. B., et al. (2018). Gaps in kelp cover may threaten the recovery of California sea otters. Ecography 41, 1751–1762. doi: 10.1111/ecog.03561
Otto, G. P., Berthrong, M., Appleby, E. E., Eawlins, J. C., and Wilbur, O. (1954). Eosinophilia and hepatomegaly due to Capillaria hepatica infection. Bull. Johns Hopkins Hosp. 94, 319–336.
Redrobe, S. P., and Patterson-Kane, J. C. (2005). Calodium hepaticum (syn. Capillaria hepatica) in captive rodents in a zoological garden. J. Comp. Pathol. 133, 73–76. doi: 10.1016/j.jcpa.2005.01.013
Resendes, A. R., Amaral, A. F. S., Rodrigues, A., and Almeria, S. (2009). Prevalence of Calodium hepaticum (Syn. Capillaria hepatica) in house mice (Mus musculus) in the Azores archipelago. Vet. Parasitol. 160, 340–343. doi: 10.1016/j.vetpar.2008.11.001
Riedman, M. L., and Estes, J. A. (1990). The sea otter (Enhydra Lutris): behavior, ecology, and natural history. United States Fish Wildl. Serv. Biol. Rep. 90, 1–126. doi: 10.2307/1382334
Rothenburger, J. L., Himsworth, C. G., Chang, V., LeJeune, M., and Leighton, F. A. (2014). Capillaria hepatica in wild Norway rats (Rattus norvegicus) from Vancouver, Canada. J. Wildl. Dis. 50, 628–633. doi: 10.7589/2013-09-256
Sandgren, F. E., Chu, E. W., and Vandevere, J. E. (1973). Maternal behavior of California sea otter. J. Mammal. 54, 668–679. doi: 10.2307/1378966
Santos, C. C. S., Onofre-Nunes, Z., and Andrade, Z. A. (2007). Role of partial hepatectomy on Capillaria hepatica-induced hepatic fibrosis in rats. Rev. Soc. Bras. Med. Trop. 40, 495–498. doi: 10.1590/S0037-86822007000500001
Shapiro, K., Miller, M. A., and Mazet, J. K. (2012). Temporal association between land-based runoff events and California sea otter (Enhydra lutris nereis) protozoal mortalities. J. Wildl. Dis. 48, 394–404. doi: 10.7589/0090-3558-48.2.394
Siskos, N., Lampe, K., Kaup, F. J., and Mätz-Rensing, K. (2015). Unique case of disseminated toxoplasmosis and concurrent hepatic capillariasis in a ring-tailed lemur: first case description. Primate Biol. 2, 9–12. doi: 10.5194/pb-2-9-2015
Solomon, G., and Handley, C. (1971). Capillaria hepatica (Bancroft, 1893) in Appalachian mammals. J. Parasitol. 57, 1142–1144. doi: 10.2307/3277883
Walker, R., Carvalho-Pereira, T., Serrano, S., Pedra, G., Hacker, K., Taylor, J., et al. (2017). Factors affecting carriage and intensity of infection of Calodium hepaticum within Norway rats (Rattus norvegicus) from an urban slum environment in Salvador, Brazil. Epidemiol. Infect. 145, 334–338. doi: 10.1017/S0950268816002259
Wright, K. A. (1961). Observations on the life cycle of Capillaria hepatica (Bancroft, 1893) with a description of the adult. NRC Res. Press. Can. J. Zool. 39, 167–182. doi: 10.1139/z61-022
Yadav, S., Sathe, P., and Ghodke, R. (2016). Hepatic capillariasis: a rare parasitic infection. Indian J. Pathol. Microbiol. 59, 124–125. doi: 10.4103/0377-4929.174878
Keywords: Calodium hepaticum, Capillaria hepatica, Enhydra lutris nereis, hepatic fibrosis, hepatitis, nematode, Rattus, southern sea otter
Citation: Miller MA, Duignan PJ, Dodd E, Batac F, Staedler M, Tomoleoni JA, Murray M, Harris H and Gardiner C (2020) Emergence of a Zoonotic Pathogen in a Coastal Marine Sentinel: Capillaria hepatica (syn. Calodium hepaticum)-Associated Hepatitis in Southern Sea Otters (Enhydra lutris nereis). Front. Mar. Sci. 7:335. doi: 10.3389/fmars.2020.00335
Received: 18 October 2019; Accepted: 22 April 2020;
Published: 25 May 2020.
Edited by:
Debra Lee Miller, The University of Tennessee, Knoxville, United StatesReviewed by:
Rick Gerhold, The University of Tennessee, Knoxville, United StatesHans-Peter Fuehrer, University of Veterinary Medicine, Vienna, Austria
Copyright © 2020 Miller, Duignan, Dodd, Batac, Staedler, Tomoleoni, Murray, Harris and Gardiner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Melissa A. Miller, melissa.miller@wildlife.ca.gov