Intensified sensitivity and adaptability of zooplankton Bosminidae in subtropical shallow freshwater lakes with increasing trophic level
 Longjuan Cheng1
Longjuan Cheng1  Xinyuan Gao1 Guangwei Wang1
Xinyuan Gao1 Guangwei Wang1  Zhou Ding1 Bin Xue2*
Zhou Ding1 Bin Xue2*  Can Zhang2* Jinliang Liu3
Can Zhang2* Jinliang Liu3  Qingfeng Jiang1
Qingfeng Jiang1- 1School of Geographical Science, Nantong University, Nantong, Jiangsu, China
- 2State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing, Jiangsu, China
- 3School of Environmental Science, Nanjing Xiaozhuang University, Nanjing, Jiangsu, China
The deterioration in lake water environments, especially increasing lake eutrophication, is prevalent all over the world, which has seriously affected the balance and stability of the internal ecosystem of lakes. In this study, modern water and sediment samples were collected from three subtropical freshwater lakes with significant differences in nutrient levels to analyze the concentration of the zooplankton Cladocera Bosminidae and its relationship with lakes’ ecological changes. The results show that the deterioration in lake water environments caused by increasing eutrophication limits the survival of most zooplankton. However, the Bosminidae shows a positive adaptability to eutrophication and high sensitivity to the changes in the lake environment. In addition, the lake eutrophication process caused by the intensification of human activities enhances the survival advantage of Bosminidae with more food sources, which is more conducive to its rapid reproduction.
1. Introduction
Crustacean zooplankton Cladocera are highly abundant in freshwater bodies worldwide. These Cladocera communities are sensitive to changes in the environment and climate (Korhola and Rautio, 2001; Szeroczyńska and Sarmaja-Korjonen, 2007; Pawłowski et al., 2015; Cheng et al., 2020a). The Bosminidae family belongs to Cladocera communities and is characterized by small body sizes (0.2–1.0 mm). They are ubiquitous on lake surfaces where the concentration of nutrients is relatively high (Zaret and Kerfoot, 1975; Jiang and Du, 1979; Zaret and Kerfoot, 1980; Threlkeld, 1981; De Melo and Hebert, 1994). The different species of the Bosminidae family exhibit unique adaptability to different nutritional environments (Jiang and Du, 1979; Szeroczyńska and Sarmaja-Korjonen, 2007). For example, Bosmina longirostris is commonly reported in eutrophic lakes, warming waters, and metal-contaminated lakes (Boucherle and Züllig, 1983; Balcer et al., 1984; Labaj et al., 2015; Adamczuk, 2016). Bosmina (E.) longispina can be used to indicate oligotrophic and mesotrophic environment changes (Deevey, 1942; Boucherle and Züllig, 1983; Miroslaw-Grabowska et al., 2015). However, due to the destruction of Bosminidae chitin fossils in the process of deposition, it is difficult to distinguish all the specific species of Bosminidae. Recent studies showed that the total number of Bosminidae, including both the eutrophic species Bosmina longirostris and oligotrophic and mesotrophic species Bosmina (E.) longispina, shows clear survival advantages in lakes with high trophic levels (Cheng et al., 2019, 2020b). For example, a large number of Bosminidae (a relative abundance greater than 80% in the total concentration of Cladocera) have been found in shallow eutrophic lakes in the subtropical zone (Liu et al., 2008; Wang et al., 2019; Cheng et al., 2020b). Furthermore, the abundance of Bosmina longirostris is usually greater than that of the species Bosmina (E.) longispina (Cheng et al., 2019, 2021) in eutrophic lakes. Therefore, we studied whether the Bosminidae can be used as a new indicator of the evolution of eutrophication in freshwater lakes in a subtropical region.
Since entering the Anthropocene, the impact of human activities on freshwater environments such as lakes has been increasing (Lin et al., 2020). Under the combined effects of anthropogenic activities (such as rapid industrialization and urbanization) and climate change, lakes in the middle and lower reaches of the Yangtze River in eastern China, which is located in a subtropical region with a relatively dense population and developed economy, have undergone drastic deterioration over a few decades (Qin et al., 2013; Zhu et al., 2019). Currently, these freshwater shallow lakes in the middle and lower reaches of the Yangtze River are a prominent area of environmental deterioration in China (Qin et al., 2013; Zhu et al., 2019). Furthermore, most lakes in this area have entered either a mesotrophic stage or a eutrophic stage, or in between, and are suffering serious cyanobacteria outbreaks that the stability and balance in the lake ecosystem are seriously disturbed (Zhu et al., 2019). For example, the biological composition of lakes has changed significantly, including the miniaturization of fishery structure, the reduction of aquatic plant area, cyanobacteria outbreaks, and a significant decline in biodiversity (Mao et al., 2012; Zhang K. et al., 2018; Jenny et al., 2020).
Taihu Lake (severe eutrophication), Nanyi Lake (transition from mesotrophication to eutrophication), and Shengjin Lake (mesotrophication) are located in the middle and lower reaches of the Yangtze River. They show significant differences in trophic level, area, and local economic development level and are representative of the research on the relationship between the evolution of zooplankton community and eutrophication. Therefore, in this study, we chose the three shallow freshwater lakes as our research object. Based on the research methods of paleolimnology and limnology, we focused on the indicative significance of Bosminidae in lake eutrophication in the three lakes since the 1950s. We analyzed the temporal and spatial changes in Bosminidae density and focused on revealing the adaptability and sensitivity of Bosminidae to lake eutrophication.
2. Materials and methods
2.1. Study area and field sampling
Taihu Lake, Nanyi Lake, and Shengjin Lake are located in the middle and lower reaches of the Yangtze River (Figure 1). The annual temperature and precipitation in the lower reaches of the Yangtze River Basin are 15–16°C and 1,010–1,600 mm, respectively (Qin et al., 2007; Wang et al., 2015; Yao and Xue, 2016; Zhang M. et al., 2018). Taihu Lake (mean depth = 1.89 m, area = 2,338 km2) is socioeconomically important and is the third largest freshwater lake in the Yangtze Delta. It supplies drinking water to some of the large cities in the delta, including Shanghai, Suzhou, and Wuxi (Qin et al., 2007). Taihu Lake is divided into different ecological types (Qin et al., 2007). The eastern area of Taihu Lake (called the East Taihu Lake) has an average water depth of < l m. As opposed to the western and central lake areas, it is a grass lake dominated by large areas of aquatic plants (Cheng et al., 2020a). More than 200 rivers flow into Taihu Lake with a combined total length of ~120,000 km (Zhong et al., 2014). The lake has been experiencing prolonged eutrophication over the past 40 years, with increased cyanobacterial (Microcystis) blooms mostly during summer, reducing both water quality and biodiversity (Qin et al., 2007; Yang et al., 2008; Li and Chen, 2010; Dai et al., 2016).
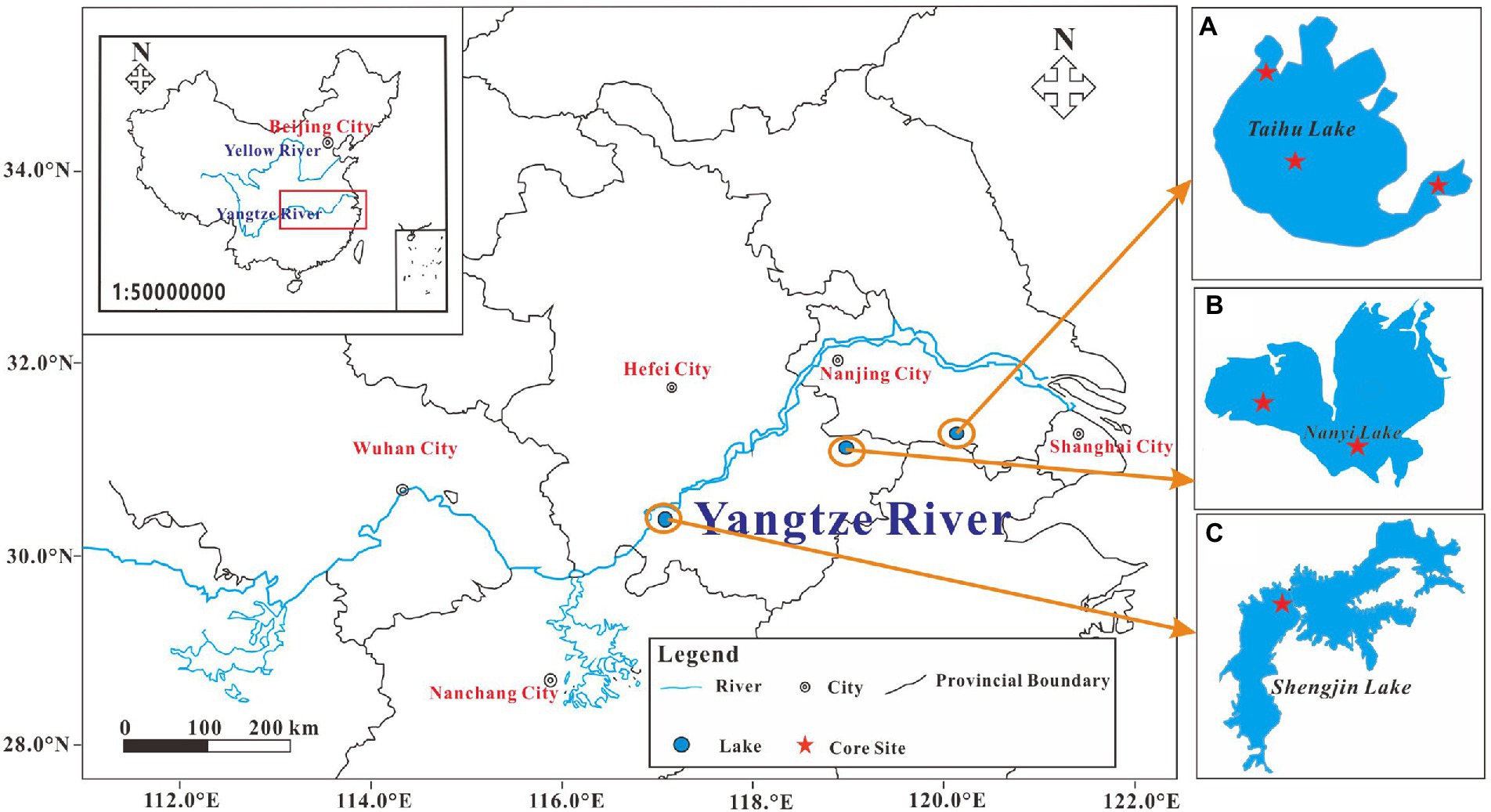
Figure 1. The location of the study area and the sample site. (A) Taihu Lake; (B) Nanyi Lake; and (C) Shengjin Lake.
Nanyi Lake (31°03′-31°10′N, 118°50′-119°02′ E) is located in the Shuiyangjiang River Basin, which belongs to the lower reaches of the Yangtze River. The lake’s surface area is 203 km2 with a mean depth of approximately 2 m (Wang et al., 2013). The Langchuan River and the New Langchuan River flow into the northeastern part of Nanyi Lake (Yao and Xue, 2016). Since the 1970s, significant increases in the trophic level and water level of Nanyi Lake have been observed, and currently, it is in a critical period of ecosystem transformation from mesotrophic to eutrophic status (Cheng et al., 2021).
Shengjin Lake (116° 55′–117° 15′ E and 30° 15′–30° 30′ N), which is an important national nature reserve, has a surface area of 133 km2 and consists of three parts: upper, middle, and lower (Wang et al., 2015). Shengjin Lake serves as a habitat for more than 70,000 wintering water birds each year (Yang, 2011). It also joined the East Asian–Australasian Waterfowl Reserve Network in 2005 and was added to the list of Wetlands of International Importance in 2015 (Cao et al., 2018; Peng et al., 2019; Yang et al., 2019). With the increase in human activities, the trophic state of Shengjin Lake has also been increasing since the 1980s (Cheng et al., 2020c).
2.2. Materials and methods
Modern zooplankton samples were collected in October 2017 and in April and July 2018. Sedimentary cores were collected in October 2016 and 2017 (Table 1). The number of zooplankton in a unit volume of water was counted and the individuals were counted under the microscope at 100–200x magnification. The pretreatment process for subfossil Cladocera and geochemical indices (TN, TP, and δ13C in the Bosminidae (δ13Cbos)) have been described in detail in previous studies (Cheng et al., 2020b,c, 2021). The subsamples for subfossil Cladocera were treated with 100 ml of 10% potassium hydroxide (KOH) solution, heated at 60°C for 45 min, sieved through a 38-μm mesh under running deionized water, and transferred into 15-ml centrifuge tubes (Frey, 1986; Korhola and Rautio, 2001). Most Cladocera remains were identified by their shell, headshield, postabdomen, and claw, and at least 100 individuals from each Cladocera (species) were counted in each sample (Jiang and Du, 1979; Szeroczyńska and Sarmaja-Korjonen, 2007). For the data on dating results and subfossil Bosminidae in the sedimentary cores, we refer to published literature and Supporting Information (Cheng et al., 2019, 2020c, 2021). Bosminidae (Bosmina spp.) includes unidentified Bosmina spp., Bosmina longirostris, Bosmina (E.) longispina, and Bosmina (E.) coregoni.
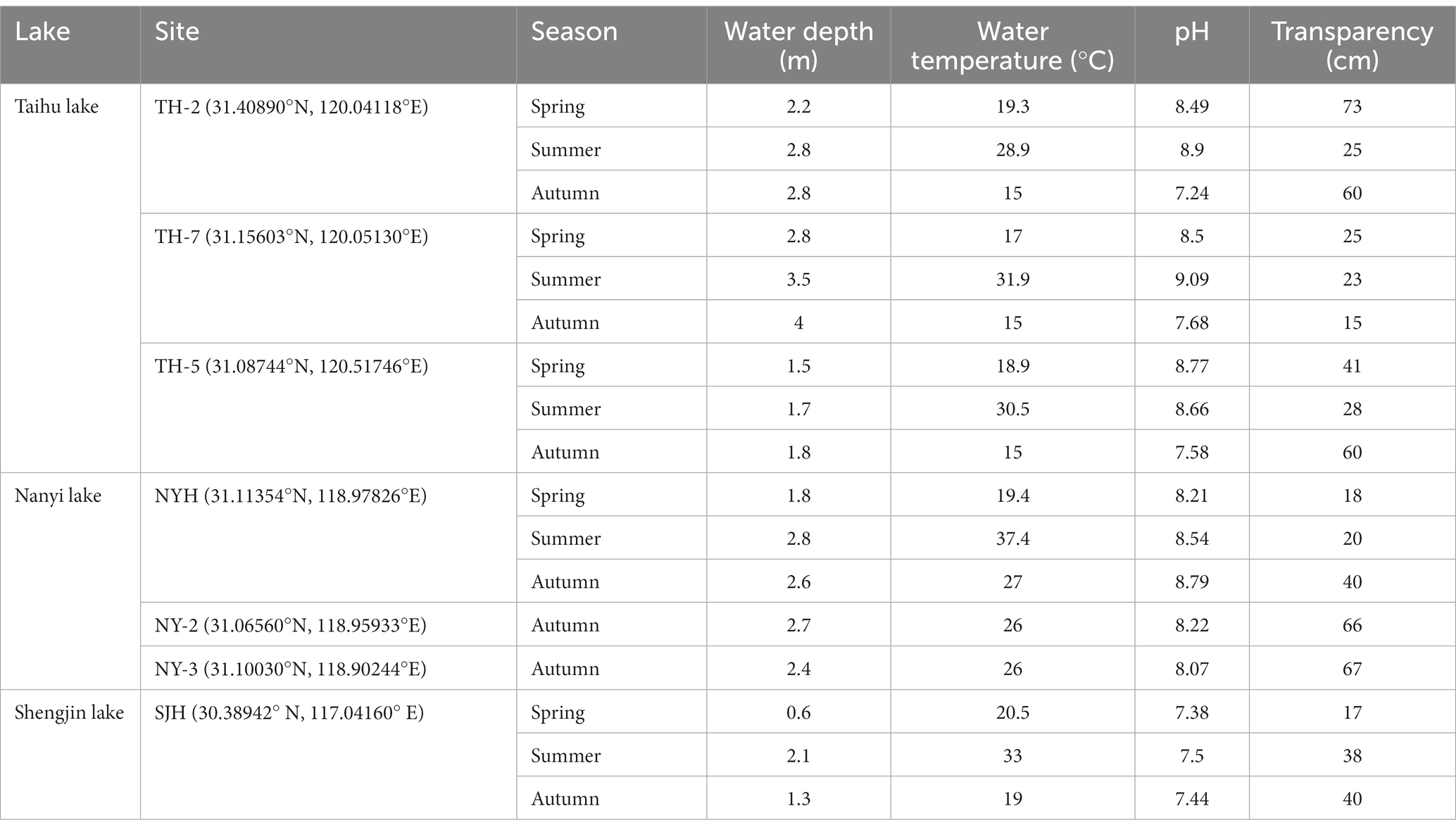
Table 1. The physical and chemical water parameters noted in the sample sites.
2.3. Statistical analysis
Graphical representations of modern zooplankton, Bosminidae, and δ13Cbos records were generated using Origin 8.0 and CorelDRAW 2018. Pearson’s correlation analysis was conducted by IBM SPSS 19.0 (Statistical Graphics Corp, Princeton, United States) and Origin 8.0 using TN, TP, and Bosminidae to investigate the relationships between the biological indicator (Cladocera) and trophic indicators (TN and TP).
3. Results
3.1. Variability in modern Bosminidae abundance
We identified the modern zooplankton in Taihu Lake, Nanyi Lake, and Shengjin Lake. The results showed that Cladocera were the dominant zooplankton species in the three lakes, with the exception of the total number of Copepoda in Shengjin Lake in summer (52 ind./L) exceeding that of Cladocera (44 ind./L) and Rotifera (5 ind./L) (Figure 2). There were higher proportions of Cladocera in zooplankton in the Nanyi Lake (NYH: mean 320 ind./L). Values in the western area of the Taihu Lake (TH-2: mean 127 ind./L) and central area (TH-7: mean 198 ind./L) were much higher than in the Shengjin Lake (SJH: mean 24 ind./L). In spring, the total number of Cladocera identified in TH-7 and NYH reached the maximum values of 478 ind./L and 805 ind./L, respectively (Figure 2). The abundance of zooplankton in the eastern area (TH-5) of the Taihu Lake was far lower than those in the western and central areas (Figure 2). Only the abundance of Cladocera in summer was greater than 50 ind./L in TH-5, while the abundances of Cladocera, Copepoda, and Rotifera in spring and autumn were relatively low and balanced. The total number of zooplankton in SJH was similar to that in TH-5 with a low abundance in each season (Figure 2).
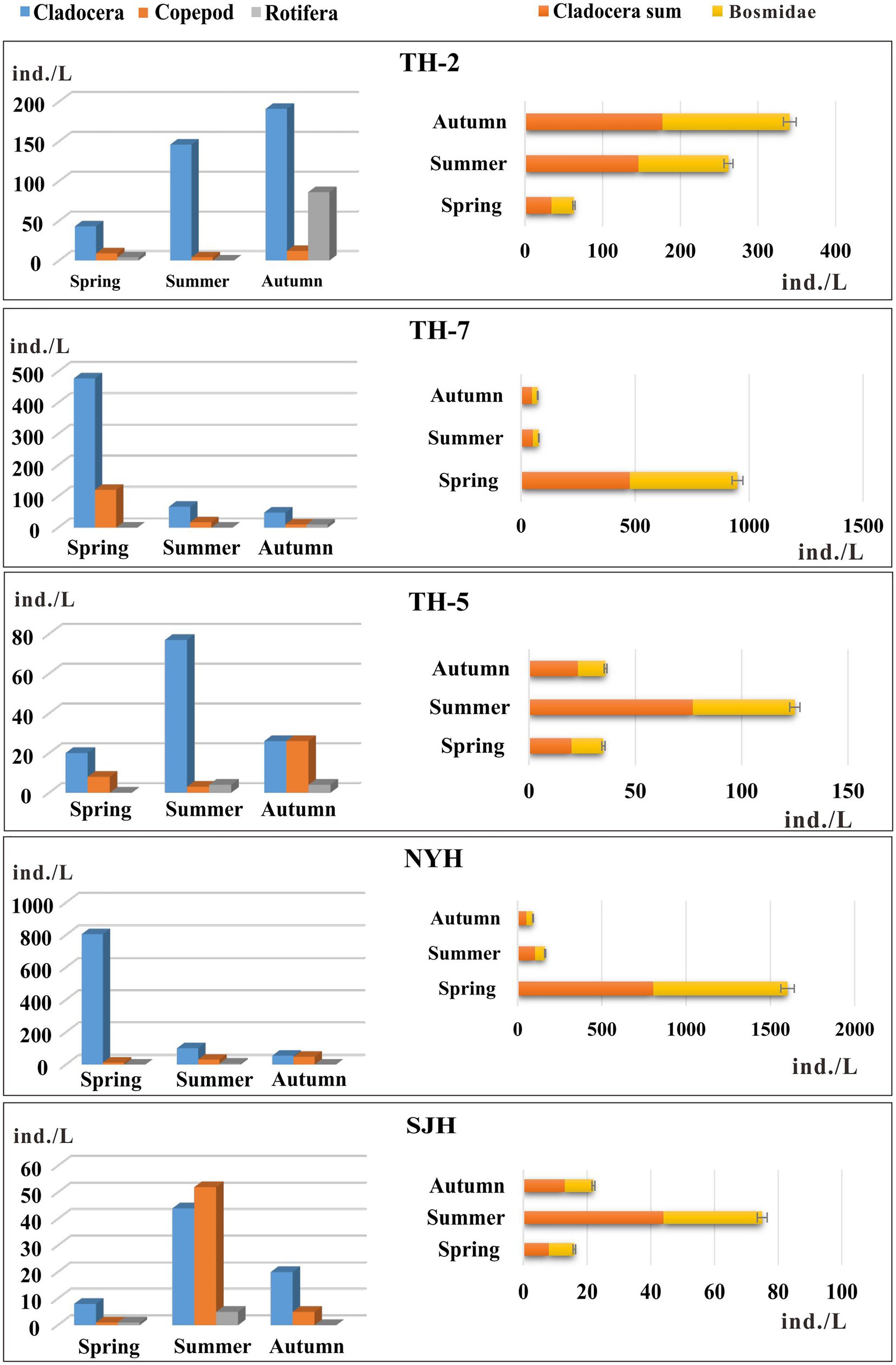
Figure 2. Modern zooplankton and Bosminidae from the Taihu Lake (TH-2, TH-7, and TH-5), Nanyi Lake (NYH), and Shengjin Lake (SJH) in the main living seasons.
The dominant species of the zooplankton composition in different seasons in the three lakes was Cladocera, which mainly consisted of Bosminidae (Figure 2). The composition characteristics of Bosminidae concentrations in each lake and season were consistent with the distribution of the total number of Cladocera. The total abundance of Bosminidae in TH-2 and TH-7 with severe eutrophication and in NYH in the transitional stage from the mesotrophic to the eutrophic status exceeded 400 ind./L, with a relative proportion of more than 95% in spring. The peak values for Bosminidae in other lake areas mostly reached in summer and autumn.
3.2. Changes in trophic levels (TN and TP) and subfossil Bosminidae abundances since the 1950s
Information about species and abundances of subfossil Cladocera and trophic indices (TN and TP) has been described in detail elsewhere (Cheng et al., 2020a,b,c, 2021). For data on Bosminidae, total Cladocera, and chronology, we refer to the “Supporting Information.” The trophic levels (TN-% and TP-mg/g) remained at a high level since the 1950s, with average values of 0.11% and 0.68 mg/g in the TH-2 core, 0.97% and 0.42 mg/g in the TH-5 core, 0.07% and 0.43 mg/g in the TH-7 core, 0.13% and 0.50 mg/g in the NY-2 core, 0.16% and 0.53 mg/g in the NY-3 core, and 0.28% and 0.59 mg/g in the SJH core (Figures 3, 4). The values of TN and TP in the west (TH-2 core) and central (TH-7 core) areas of the Taihu Lake significantly increased, while the values of TN in the east (TH-5 core) tended to decline but were kept at a higher level (0.92% ~ 1.0%) than in the TH-2 core (0.08% ~ 0.18%) and TH-7 core (0.06% ~ 0.10%). The values of TP in the cores of TH-2, TH-7, and TH-5 ranged from 0.12 to 0.48 mg/g, 0.35 to 0.51 mg/g, and 0.40 to 0.45 mg/g, respectively. The abundance of Bosminidae in the Taihu Lake changed consistently with TN and TP in the western and central areas, but there was a significant decreasing trend in the eastern Taihu Lake (Figures 3, 4). The values of TN and TP in the Nanyi Lake and Shengjin Lake clearly increased. The values of TN in the cores of NY-2, NY-3, and SJH ranged from 0.08 to 0.20%, 0.10 to 0.21%, and 0.23 to 0.31%, respectively. The values of TP in the cores of NY-2, NY-3, and SJH changed from 0.31 to 0.69 mg/g, 0.37 to 0.68 mg/g, and 0.45 to 0.64 mg/g, respectively. The abundance of Bosminidae also increased corresponding to the increase in nitrogen and phosphorus concentrations in the Nanyi Lake and Shengjin Lake.
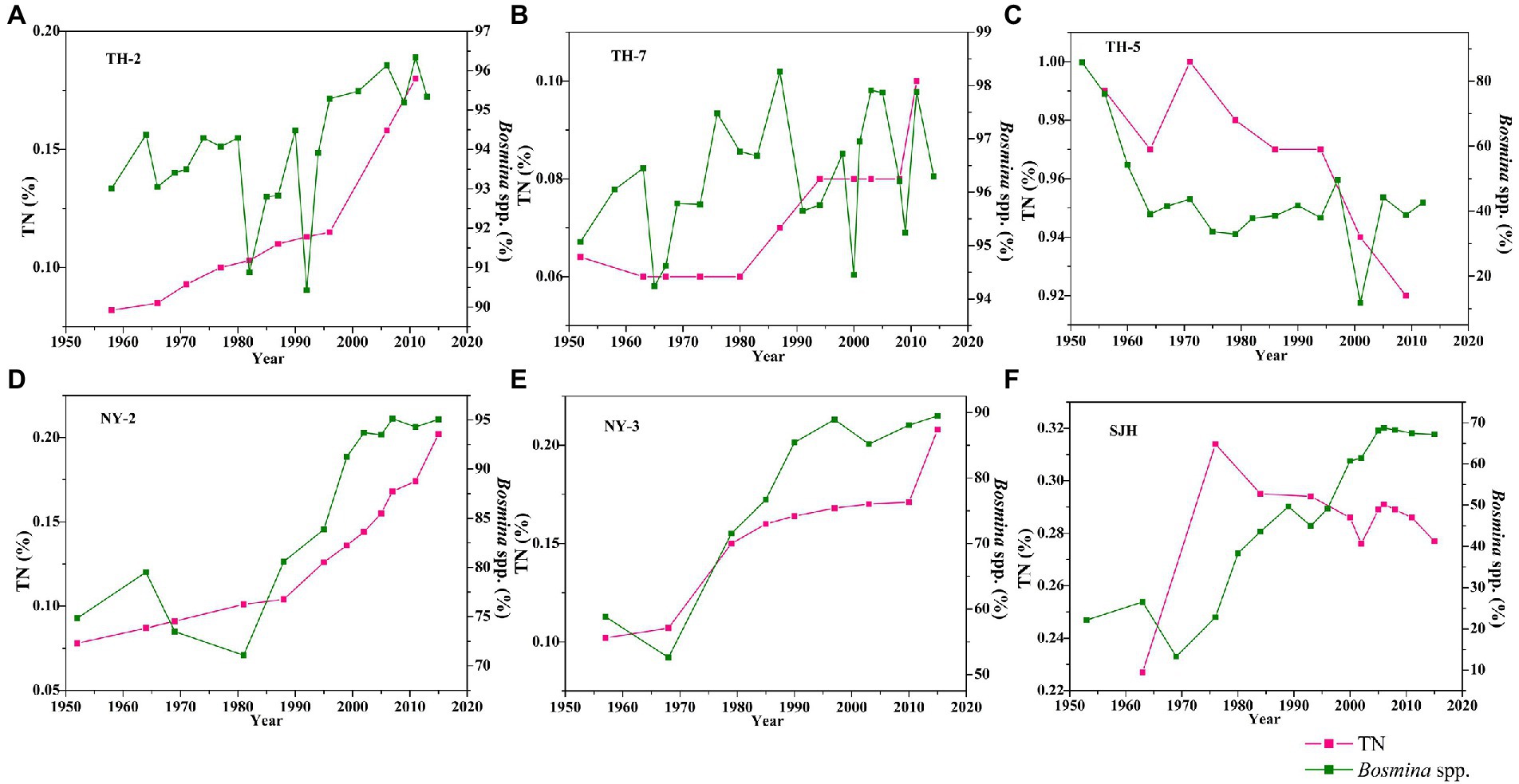
Figure 3. Changes in TN and Bosminidae (Bosmina spp.) in the Taihu Lake (A–C), Nanyi Lake (D,E), and Shengjin Lake (F) since the 1950s.
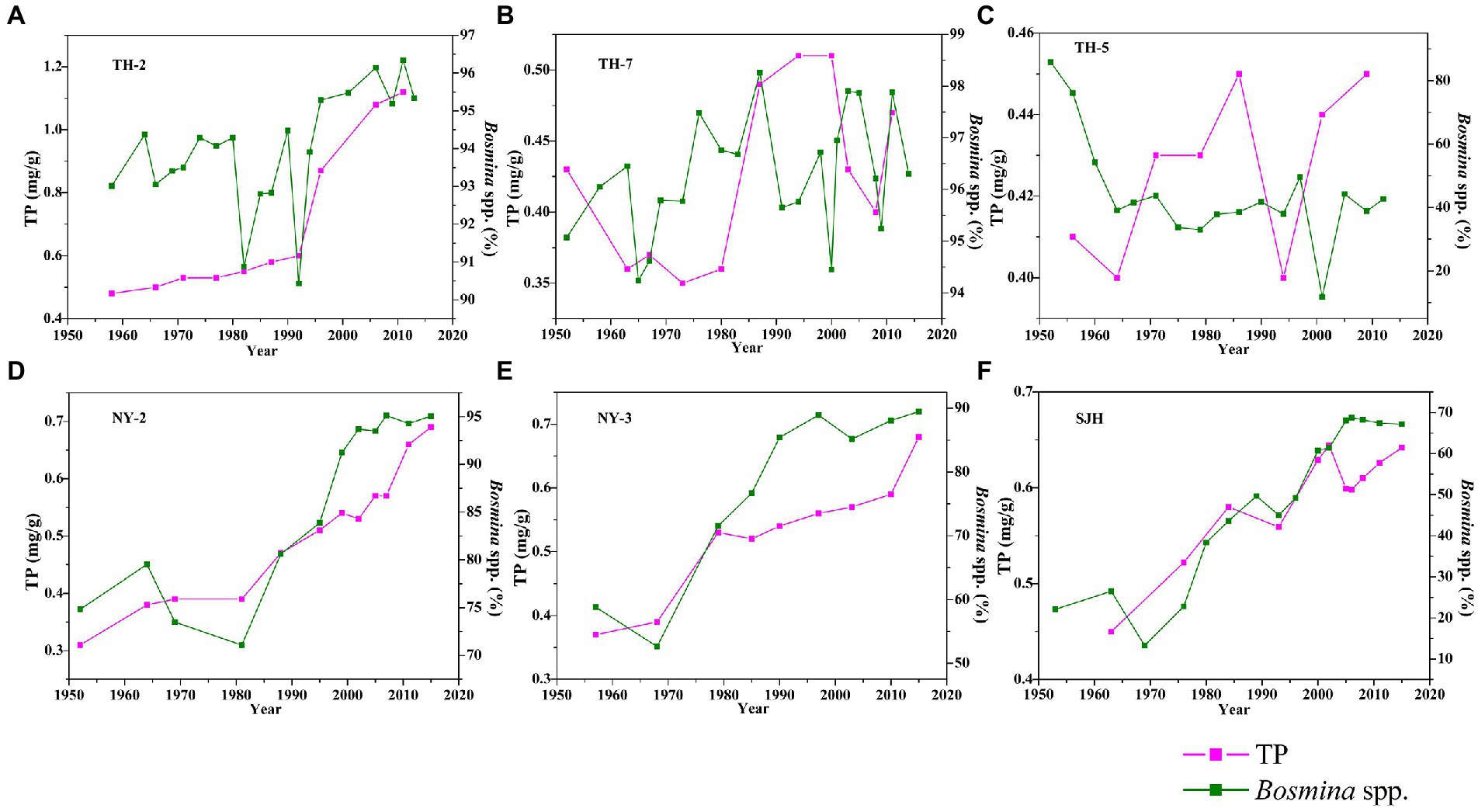
Figure 4. Changes in TP and Bosminidae (Bosmina spp.) in the Taihu Lake (A–C), Nanyi Lake (D,E), and Shengjin Lake (F) since the 1950s.
The abundances of Cladocera sum and Bosminidae in the cores of TH-2 and TH-7 remained at a high level. Among them, the abundances of total Cladocera and Bosminidae in the TH-2 core ranged from 1,080 ~ 5,295 ind./g with a mean value of 3,048 ind./g, i.e., 94% ~ 98% with a mean value of 96%, respectively, while those in the TH-7 core ranged from approximately 2,507 ~ 9,373 ind./g with a mean value of 5,140 ind./g, i.e., 90% ~ 96% with a mean value of 94%, respectively (Figure 5). After the 1950s, the Cladocera sum decreased in the TH-2 core, but the abundance of Bosminidae increased significantly. The change in Cladocera sum and Bosminidae abundances in the TH-7 core was similar to that in the TH-2 core; that is, the fluctuation in Bosminidae increased after the 1950s (Figure 5). The change in Cladocera sum in the TH-5 core corresponding to that of Bosminidae decreased after the 1950s and their changed ranges were 463 ~ 1,127 ind./g with a mean value of 758 ind./g, i.e., 12% ~ 86% with a mean value of 44%, respectively. The change in the abundance of Bosminidae was consistent with that of the Cladocera sum that increased gradually with time in Nanyi Lake and Shengjin Lake (Figure 5). The ranges for Cladocera sum and Bosminidae in the NY-2 core were 111 ~ 901 ind./g with a mean value of 444 ind./g, i.e., 71% ~ 95% with a mean value of 86%, respectively. Values in the NY-3 core were 101 ~ 639 ind./g with a mean value of 400 ind./g, i.e., 53% ~ 89% with a mean value of 77%, respectively. The abundances of Cladocera sum and Bosminidae in Shengjin Lake (SJH core) were relatively lower than those of in the other two lakes, which changed from 241 ~ 487 ind./g with a mean value of 366 ind./g, i.e., 13% ~ 69% with a mean value of 48%, respectively (Figure 5).
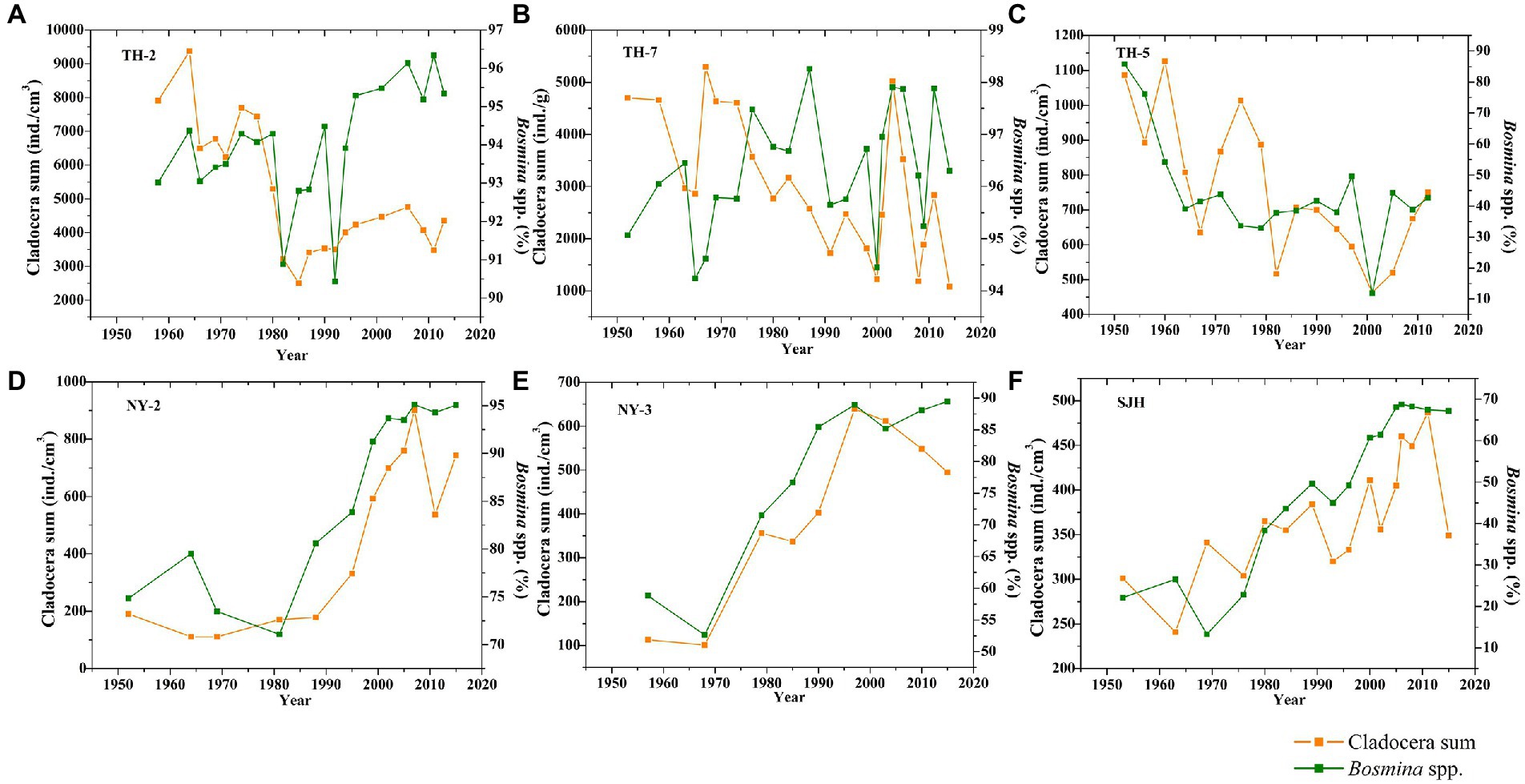
Figure 5. Changes in Cladocera sum and Bosminidae (Bosmina spp.) in the Taihu Lake (A–C), Nanyi Lake (D,E), and Shengjin Lake (F) since the 1950s.
3.3. Changes in the δ13C values of subfossil Bosminidae in Taihu Lake since the 1950s
To accurately track the flow characteristics of carbon sources in primary consumers, the results of the stable carbon isotope analysis of subfossil Bosminidae (δ13Cbos) in the cores of TH-2 and TH-7 from the heavily eutrophic area in Taihu Lake after the 1950s are shown in Figure 6. The mean values of δ13Cbos in the cores of TH-2 and TH-7 showed relatively consistent mean values of −22.45‰ and − 22.51‰, respectively. However, the variation range in the δ13Cbos value of the TH-2 core was higher than that of the TH-7 core (Figure 6A). The δ13Cbos values in the TH-2 core varied in the range of −25.77‰ ~ −19.54‰, while the δ13Cbos values in the TH-7 core changed from −23.40‰ to −20.80‰. After the 1980s, with intensified eutrophication of Taihu Lake, the δ13Cbos values of the two cores shared a common downward trend that was lower than the mean values (Figure 6B).
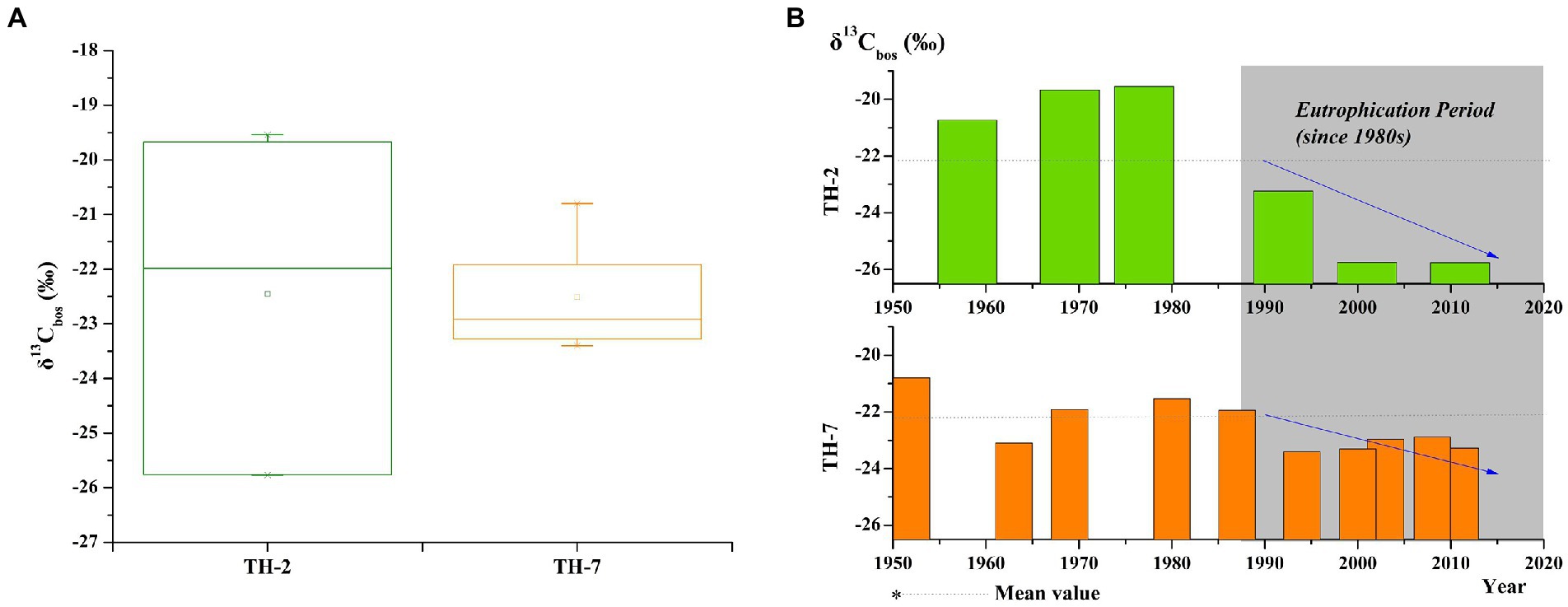
Figure 6. Changes in the δ13C values of subfossil Bosminidae in the Taihu Lake after the 1950s. (A) Statistical distribution of δ13Cbos in the cores of TH-2 and TH-7 and (B) temporal distribution of δ13Cbos in the cores of TH-2 and TH-7.
3.4. Pearson’s correlation analysis between Bosminidae abundance change and trophic level
The Pearson correlation analysis between Bosminidae and trophic indices showed that the abundance of Bosminidae, including B. longirostris and B (E.) longispina, were significantly correlated with TN and TP (Table 2; Cheng et al., 2020a,b,c, 2021). There were significant positive correlations between Bosminidae and TN in the cores of TH-2, NY-2, and NY-3. The Bosminidae were positively correlated with TP in all cores except TH-5, which had a significant negative correlation. In addition, there were significant positive correlations between Bosminidae and TP in the cores of Nanyi Lake and Shengjin Lake. The eutrophic species B. longirostris in the three lakes was significantly positively correlated with TN, except in the TH-5 core. There were significant positive correlations between B. longirostris and TP in the cores of TH-2, NY-2, NY-3, and SJH. The oligotrophic species B (E.) longispina had a significant negative correlation with TN in the cores of TH-2 and TH-7 but a significant positive correlation in the two cores of Nanyi Lake. B (E.) longispina was negatively correlated with TP in the cores of TH-2 and TH-5 and was positively correlated with TP in the cores of NY-2 and NY-3.

Table 2. Pearson’s correlation analysis between the trophic indices TN and TP with the species of Bosminidae. Data sources of TH-2, TH-7, TH-5, NY-2, NY-3, and SJH cores were taken from Cheng et al. (2020a,b,c, 2021).
4. Discussion
The lake trophic status in the lower reaches of the Yangtze River from a recent study showed that most of the lakes have been categorized into the mesotrophic or the eutrophic status (Cheng and Li, 2006; Qin et al., 2013). The trophic levels of the three lakes, located in the middle and lower reaches of the Yangtze River, have also reached the mid-eutrophic level and above and are increasing from Shengjin Lake to Nanyi Lake and then to Taihu Lake, which is closely related to the distribution of economic development, population, and urbanization in each region (Qin et al., 2013; Zhang M. et al., 2018; Shi et al., 2019; Cheng et al., 2020c, 2021). An analysis of modern zooplankton compositions showed that Cladocera was dominant in the three lakes and that the abundance was gradually increasing from Shengjin Lake through Nanyi Lake to Taihu Lake (Figure 2). Furthermore, the abundances of Bosminidae in the Nanyi Lake and in the western and central areas of the Taihu Lake were greater than 90% (Figure 2). This showed that Bosminidae abundance increased corresponding to the increasing trophic level (Li et al., 2016; Cheng et al., 2020b; Li et al., 2021). However, a higher Copepod density was recorded in summer than that of Cladocera and there was a low abundance of Bosminidae in the composition of modern zooplankton in the Shengjin Lake, which might indirectly suggest that the Bosminidae did not have a reproductive advantage under low trophic levels. The subfossil Cladocera in the sedimental cores of the three lakes also showed that the Bosminidae was a broadly dominant species (Figure 5). Especially in Taihu Lake, the density of Bosminidae subfossils was close to 10,000 ind./cm3 (Cheng et al., 2019). In Nanyi Lake and Shengjin Lake, where trophic levels had increased significantly since the 1950s, the abundance of subfossil Bosminidae also increased (Figure 5). In addition, among the modern Cladocera in the three lakes, Bosminidae was the dominant species, which was consistent with the Cladocera subfossil results at the same location and also indirectly proved that Bosminidae subfossils were well preserved in lakes of the middle and lower reaches of the Yangtze River.
In terms of space, there were huge differences in the economic development level, the urbanization level, and population along the Yangtze River from west to east. Taihu Lake is located in the eastern delta of the Yangtze River, which has an extremely developed economy and a very large population (Qin et al., 2007). Because Taihu Lake is located near many developed cities such as Shanghai, Suzhou, and Wuxi, a large quantity of nutrients such as nitrogen and phosphorus flow into Taihu Lake on account of human activities, which are far greater in Taihu Lake than in Nanyi Lake and Shengjin Lake in the Anhui Province (Dai et al., 2016). Agricultural and fisheries development in Nanyi Lake is more adequate than in Shengjin Lake (Wang et al., 2013). As a national nature reserve, Shengjin Lake has a relatively good environment and low nutrient level compared with the other two lakes (Wang et al., 2015). The spatial gradient distributions of Bosminidae abundance in the three lakes were in line with the different economic development levels (Figure 5). Thus, it could be inferred that there were different types of interference of human activities around the three lakes.
The significant positive relationship between Bosminidae and eutrophic species B. longirostris with TN and TP further proved that Bosminidae had an advantage in the eutrophic process (Table 2). In particular, B. longirostris demonstrated strong adaptability and sensitivity in a wide range of water environments, such as the eutrophic status, warming waters, and metal-contaminated lakes in previous studies (Boucherle and Züllig, 1983; De Kluijver et al., 2012; Labaj et al., 2015; Kong et al., 2017; Cheng et al., 2020b). Our study showed that Bosminidae abundance increased with the increase in the outbreak intensity of cyanobacteria in eutrophic lake areas (Figures 3, 4). Especially in the west and central lake areas of Taihu Lake with serious eutrophication, the concentration of Bosminidae has increased to a relatively stable state. While Nanyi Lake and Shengjin Lake are in the stage of rising trophic levels, and the abundance of Bosminidae is also showing a synchronous rising trend (Figure 3). Although previous studies suggested that Bosminidae were too small to feed on Microcystis directly with a large size (Jiang and Du, 1979; Li and Chen, 2010), more studies showed that Microcystis tended to form large suspended aggregates with materials after cell death including dissolved organic carbon, dissolved inorganic carbon, and dissolved inorganic nitrogen and phosphorus in the water column and then were likely the main diet of Bosminidae (De Kluijver et al., 2012; Agasild et al., 2019; Cheng et al., 2020b). It could be speculated that cyanobacteria blooms might change the food preference of the zooplankton Bosminidae. At the same time, the water environment also changed greatly due to excessive reproduction of cyanobacteria; thus, Bosminidae made timely adaptive adjustments to the original living environment and food source changes (Agasild et al., 2019). For example, Cladocera’s size gradually became smaller to better escape the predation of zooplankton-feeding fish (Shurin, 2001; Jeppesen et al., 2003; Liu et al., 2009). Accordingly, in lake areas where cyanobacteria blooms were stronger, the density of small-size species of Bosminidae was greater and other species of zooplankton may have decreased or even gone extinct (Jiang et al., 2013; Li et al., 2016).
The present study revealed that Bosminidae could accelerate reproduction under rich food sources, resulting in excessive abundance (accounting for more than 90% of the total number of Cladocera) (Figure 2). However, water temperature also impacts the body size and community trophic structure of zooplankton (Gao et al., 2021). The three shallow freshwater lakes located in the middle and lower reaches of the Yangtze River in a subtropical zone had suitable temperature conditions that produced a favorable habitat for most lake creatures, including the zooplankton. However, a strong disturbance can easily occur in these shallow freshwater lakes, which can result in the release of sediment nitrogen and phosphorus and a large amount of exogenous nitrogen and phosphorus inputs from catchment by increasing human activities (Zhu et al., 2019). Excessive nitrogen and phosphorus inputs caused cyanobacteria blooms that seriously threatened the survival of most aquatic organisms such as fish. Large aquatic plants and other biodiversity significantly decreased, and this finally led to the imbalance of lake ecosystems (Qin et al., 2013). Surprisingly, the cyanobacteria outbreak provided favorable conditions for the expansion of the Bosminidae (De Kluijver et al., 2012; Cheng et al., 2020b). The changes in δ13Cbos values after 1950s also revealed the flow characteristics of carbon sources in the Bosminidae (Figure 6). In particular, after the 1980s (after China’s Reform and Open Policy), with increasing lake eutrophication, the δ13Cbos values exhibited a significant negative trend (Figure 6), which was consistent with the change in organic carbon isotopes in sediments (δ13Corg; Cheng et al., 2020b). The δ13C value of algae in Taihu Lake was similar to the value of δ13C in modern Bosminidae during the cyanobacteria bloom (Cheng et al., 2020b). This further indicated that the cyanobacteria outbreak provided a rich food source for Bosminidae. Bosminidae propagation in large numbers due to the outbreak of cyanobacteria also showed that Bosminidae had its own unique adaptability to the eutrophic environment and strong competitiveness compared with other zooplankton. The high adaptive ability of Bosminidae to eutrophication could indicate that it is more competitive than other larger Cladocera species.
Our data on subfossil Cladocera in the TH-5 core of East Taihu Lake showed that the total Cladocera and Bosminidae abundances were lower than those found in the core of other lake areas and tended to decrease after the 1950s (Figure 2). This was due to intensification of the swamping trend in East Taihu Lake, the water level reduction, aquatic macrophyte development, and water stagnation for prolonged periods of time caused by enclosed fish cultures and crab farming practices. These are extremely detrimental to the survival of the planktonic species Bosminidae but provide a more suitable habitat for the littoral Cladocera species (Yang et al., 2003; Li, 2004; Cheng et al., 2020a). The significant negative correlation between Bosminidae and TP also indirectly proved that the environment of the East Taihu Lake was not conducive to the survival of Bosminidae (Table. 2). It could be seen that the trophic level was not the only factor limiting the growth of Bosminidae, but it was the key factor behind the rapid expansion of Bosminidae, once certain restrictive conditions were exceeded. As shown in the western and central areas of Taihu Lake, Bosminidae reproduction expanded and abundance increased once the trophic level was enriched and the water level depth met the living conditions of the planktonic species Bosminidae (Figures 2, 5). Additionally, the significant positive correlations between oligotrophic species Bosminidae with TN and TP in Nanyi Lake were opposite of those in Taihu Lake, which was closely related to the fact that Nanyi Lake was in a critical period of transition from the mesotrophic to the eutrophic status (Cheng et al., 2021). Research by Dodson (1992) showed that the abundance of zooplankton usually increased with increasing nutritional levels within a certain range. The current trophic level of the Nanyi Lake is still increasing, but it has not reached the eutrophic level as in the central and western areas of Taihu Lake. Therefore, the abundance of oligotrophic species increased significantly with the increase in trophic level.
5. Conclusion
The spatial distribution of the three subtropical shallow freshwater lakes in the middle and lower reaches of the Yangtze River was characterized by gradually increasing nutrient levels from west to east. An analysis of the composition of zooplankton in the modern lake water of the three lakes and Cladocera subfossils from the sedimental cores revealed that the Cladocera planktonic species Bosminidae changed relatively consistently with the temporal and spatial distributions of the nutrient levels in the three lakes, indicating that the small planktonic Bosminidae species had unique sensitivity and adaptability during lake eutrophication. In addition, the significant correlation between Bosminidae and nutrient indicators such as TN and TP, as well as the gradually negative δ13Cbos values under increasing eutrophication, further proved that the abundance of Bosminidae changed with the change in eutrophication level. This reflected the change in food source preference of the Bosminidae and its strong adaptability to harsh environments during lake eutrophication. Bosminidae was more adaptive to the change in this environment, which showed that the genus was highly sensitive to changes in the lake environment and can indicate the evolution of the lake. This study offered a new alternative index for paleoecology research and also provided a new idea for mechanism research on modern lake eutrophication environments.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
LC: writing–original draft, validation, resources, and formal analysis. XG, GW, and JL: visualization and investigation. ZD: investigation and field work. BX: supervision, project administration, and funding acquisition. CZ: supervision, visualization, investigation, and writing–editing. QJ: software and resources. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by grants from the Natural Science Foundation of China project (42002204 and 42177426), the National Key Research and Development Program of China (2019YFA0607100), and the Jiangsu Special Fund on Technology Innovation of Carbon Dioxide Peaking and Carbon Neutrality (BK20220016).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1121632/full#supplementary-material
References
Adamczuk, M. (2016). Past, present, and future roles of small cladoceran Bosmina longirostris (O. F. Müller, 1785) in aquatic ecosystems. Hydrobiologia 767, 1–11. doi: 10.1007/s10750-015-2495-7
Agasild, H., Panksep, K., Tõnno, I., Blank, K., Kõiv, T., Freiberg, R., et al. (2019). Role of potentially toxic cyanobacteria in crustacean zooplankton diet in a eutrophic lake. Harmful Algae 89:101688. doi: 10.1016/j.hal.2019.101688
Balcer, M. D., Korda, N. L., and Dodson, S. I.. (1984). Zooplankton of the Great Lakes: A Guide to the Identification and Ecology of the Common Crustacean Species. Madison: University of Wisconsin Press.
Boucherle, M. M., and Züllig, H. (1983). Cladoceran remains as evidence of change in trophic state in three Swiss lakes. Hydrobiologia 103, 141–146. doi: 10.1007/BF00028442
Cao, L., Pan, C., and Wang, X. (2018). Study on environmental protection and ecological construction in Shengjinhu lake basin. Guangdong Chem. Ind. 45, 198–199. doi: 10.3969/j.issn.1007-1865.2018.05.091
Cheng, L. J., Giri, K., Xue, B., Yao, S., Li, L., and Liu, J. (2020b). Application of subfossil Bosmina and its δ13C values in tracing the long-term food web dynamics of shallow eutrophic lakes: a case in Taihu Lake, Southeast China. Sci. Total Environ. 730:138909. doi: 10.1016/j.scitotenv.2020.138909
Cheng, X. Y., and Li, S. J. (2006). An analysis on the evolvement processes of lake eutrophication and their characteristics of the typical lakes in the middle and lower reaches of Yangtze River (in Chinese). Chin. Sci. Bull. 51, 1603–1613. doi: 10.1007/s11434-006-2005-4
Cheng, L. J., Xue, B., Yao, S., and Liu, J. (2020c). Response of Cladocera fauna to environmental change based on sediments from Shengjin Lake, a Yangtze River-connected Lake in China. Quat. Int. 536, 52–59. doi: 10.1016/j.quaint.2019.11.044
Cheng, L. J., Xue, B., Zawisza, E., Yao, S., Liu, J., and Li, L. (2020a). Effects of environmental change on subfossil Cladoceran in the subtropical shallow freshwater East Taihu Lake, China. Catena 188:104446. doi: 10.1016/j.catena.2019.104446
Cheng, L. J., Xue, B., Zawisza, E., Yao, S., Liu, J., and Li, S. (2021). Specific species response of Cladocera to the trophic and hydrological environments of lakes: a case study of a typical shallow mesotrophic lake. Catena 207:105630. doi: 10.1016/j.catena.2021.105630
Cheng, L. J., Yao, S., Xue, B., Liu, J., and Li, L. (2019). Long-term change of the assemblages and abundance of cladocerans in different ecotypes of Lake Taihu. J. Lake Sci. 31, 1670–1684. doi: 10.18307/2019.0607
Dai, X. L., Qian, P. Q., Ye, L., and Song, T. (2016). Changes in nitrogen and phosphorus concentrations in Lake Taihu, 1985–2015. J. Lake Sci. 28, 935–943. doi: 10.18307/2016.0502
De Kluijver, A., Yu, J., Houtekamer, M., Middelurg, J. J., and Liu, Z. (2012). Cyanobacteria as a carbon source for zooplankton in eutrophic Lake Taihu, China, measured by 13C labeling and fatty acid biomarkers. Limnol. Oceanogr. 57, 1245–1254. doi: 10.4319/lo.2012.57.4.1245
De Melo, R., and Hebert, P. (1994). A taxonomic reevaluation of north-American Bosminidae. Can. J. Zool. 72, 1808–1825. doi: 10.1139/z94-245
Deevey, E. S. J. (1942). Studies on Connecticut lake sediments. III. The Biostratonomy of Linsley pond. Am. J. Sci. 240, 233–264. doi: 10.2475/ajs.240.4.233
Dodson, S. (1992). Predicting crustacean zooplankton species richness. Limnol. Oceanogr. 37, 848–856. doi: 10.4319/lo.1992.37.4.0848
Frey, D. G. (1986). “Cladocera analysis” in Handbook of Holocene Palaeoecology and Palaeohydrology. ed. B. E. Berglund (New York: John Whiley & sons), 667–692.
Gao, X., Tõnno, I., Vrede, T., Freiberg, R., Nilsson, J. L., and Goedkoop, W. (2021). Both climate trends and short-term fluctuations affected algae–zooplankton interactions in a boreal lake during the late Holocene. Freshw. Biol. 66, 2076–2085. doi: 10.1111/fwb.13815
Jenny, J. P., Anneville, O., Arnaud, F., Baulaz, Y., and Weyhenmeyer, G. A. (2020). Scientists' warning to humanity: rapid degradation of the world's large lakes. J. Great Lakes Res. 46, 686–702. doi: 10.1016/j.jglr.2020.05.006
Jeppesen, E., Jensen, J. P., Lauridsen, T. L., Amsinck, S. L., Christoffersen, K., Sondergaard, M., et al. (2003). Sub-fossils of cladocerans in the surface sediment of 135 lakes as proxies for community structure of zooplankton, fish abundance and lake temperature. Hydrobiologia 491, 321–330. doi: 10.1023/A:1024488525225
Jiang, X., and Du, N.. (1979). Fauna of China Arthropod Phylum Crustacean Freshwater Cladocera. Beijing: Science Press, 41–42.
Jiang, X., Li, Q., Liang, H., Zhao, S., Zhang, L., Zhao, Y., et al. (2013). Clonal variation in growth plasticity within a Bosmina longirostris population: the potential for resistance to toxic cyanobacteria. PLoS One 8:e73540. doi: 10.1371/journal.pone.0073540
Kong, L., Yang, X., Kattel Anderson, G. N. J., and Hu, Z. (2017). The response of Cladocerans to recent environmental forcing in an alpine Lake on the SE Tibetan plateau. Hydrobiologia 784, 171–185. doi: 10.1007/s10750-016-2868-6
Korhola, A., and Rautio, M. (2001). “Cladoceran and other branchiopod crustaceans” in Tracking Environmental Change Using Lake Sediments: Volume 4. Zoological Indicators. eds. J. P. Smol, H. J. B. Birks, and W. M. Last (New York: Kluwer Academic Publishers), 5–41.
Labaj, A. L., Kurek, J., Jeziorski, A., and Smol, J. P. (2015). Elevated metal concentration inhibit biological recovery of Cladocera in previously acidified Boral lakes. Freshw. Biol. 60, 347–359. doi: 10.1111/fwb.12496
Li, W. (2004). “Tendency and reason for evolution toward mash in the east Taihu” in Process and Mechanism of Environmental Changes of Lake Taihu. eds. B. Qin, W. Hu, and W. Chen (Beijing: Science Press), 33–51.
Li, J., and Chen, F. (2010). Preliminary analysis on population declines of daphnia in summer and autumn in Lake Taihu. J. Lake Sci. 22, 552–556. doi: 10.18307/2010.0411
Li, Y., Hu, L., Zhao, Y., Wang, H., Huang, X., Chen, G., et al. (2021). Meltwater-driven water-level fluctuations of Bosten lake in arid China over the past 2,000 years. Geophys. Res. Lett. 48:2020GL090988. doi: 10.1029/2020GL090988
Li, Y., Xie, P., Zhao, D., Zhu, T., Guo, L., and Zhang, J. (2016). Eutrophication strengthens the response of zooplankton to temperature changes in a high-altitude lake. Ecol. Evol. 6, 6690–6701. doi: 10.1002/ece3.2308
Lin, Q., Zhang, K., Liu, E., Sabatier, P., Arnaud, F., and Shen, J. (2020). Deciphering centurial anthropogenic pollution processes in large lakes dominated by socio-economic impacts. Anthropocene 32:100269. doi: 10.1016/j.ancene.2020.100269
Liu, G., Chen, F., and Liu, Z. (2008). Preliminary study on cladoceran microfossils in the sediments of Lake Taihu. J. Lake Sci. 20, 470–476. doi: 10.18307/2008.0410
Liu, G., Liu, Z., Li, Y., Chen, F., Gu, B., and Smoak, J. M. (2009). Effects of fish introduction and eutrophication on the cladoceran community in Lake Fuxian, a deep oligotrophic lake in Southwest China. J. Paleolimnol. 42, 427–435. doi: 10.1007/s10933-008-9286-3
Mao, Z., Gu, X., Zeng, Q., Zhou, L., and Sun, M. (2012). Food web structure of a shallow eutrophic Lake (lake Taihu, China) assessed by stable isotope analysis. Hydrobiologia 683, 173–183. doi: 10.1007/s10750-011-0954-3
Miroslaw-Grabowska, J., Zawisza, E., Jaskolka, A., and Obremska, M. (2015). Natural transformation of the Romoty paleolake (NE Poland) during the late glacial and Holocene based on isotopic, pollen, cladoceran and geochemical data. Quat. Int. 386, 171–185. doi: 10.1016/j.quaint.2015.06.040
Pawłowski, D., Płóciennik, M., Brooks, S. J., Luoto, T. P., Milecka, K., Nevalainen, L., et al. (2015). A multiproxy study of younger Dryas and early Holocene climatic conditions from the Grabia River paleo-oxbow lake (Central Poland). Palaeogeogr. Palaeoclimatol. Palaeoecol. 438, 34–50. doi: 10.1016/j.palaeo.2015.07.031
Peng, L., Dong, B., Wang, P., Sheng, S., Sun, L., Fang, L., et al. (2019). Liping Liu research on ecological risk assessment in land use model of Shengjin Lake in Anhui province. China. Environ. Geochem. Health 1:15. doi: 10.1007/s10653-019-00322-6
Qin, B. Q., Gao, G., Zhu, G. W., Zhang, Y. L., Song, Y. Z., Tang, X. M., et al. (2013). Lake eutrophication and its ecosystem response. Sci. Bull. 58, 961–970. doi: 10.1007/s11434-012-5560-x
Qin, B. Q., Xu, P., Wu, Q., Luo, L., and Zhang, Y. (2007). Environmental issues of Lake Taihu, China. Hydrobiol 581, 3–14. doi: 10.1007/s10750-006-0521-5
Shi, K., Zhang, Y., Zhang, Y., Li, N., Qin, B., Zhu, G., et al. (2019). Phenology of phytoplankton blooms in a trophic lake observed from long-term MODIS data. Environ. Sci. Technol. 53, 2324–2331. doi: 10.1021/acs.est.8b06887
Shurin, J. B. (2001). Interactive effects of predation and dispersal on zooplankton communities. Ecology 82, 3404–3416. doi: 10.1890/0012-9658(2001)082[3404:IEOPAD]2.0.CO;2
Szeroczyńska, K., and Sarmaja-Korjonen, K.. (2007). Atlas of Subfossil Cladoceran from Central and Northern Europe. Swiecie: Friends of the Lower Vistula Society. 10–11.
Threlkeld, S. (1981). The recolonization of Lake Tahoe by Bosmina-longirostris-evaluating the importance of reduced Mysis-relicta population. Limnol. Oceanogr. 26, 433–444. doi: 10.4319/lo.1981.26.3.0433
Wang, L., Wang, C., Deng, D., Zhao, X., and Zhou, Z. (2015). Temporal and spatial variations in phytoplankton: correlations with environmental factors in Shengjin Lake. China. Environ. Sci. Pollut. Res. 22, 14144–14156. doi: 10.1007/s11356-015-4640-2
Wang, Z., Yan, Z., Xu, L., Lu, X., Winemiller, K. O., and Lei, G. (2013). Effects of hydrologic regulation on icefish population dynamics, assemblage structure and fishery yield in Lake Nanyi, China. Ecol. Freshw. Fish 22, 637–644. doi: 10.1111/eff.12068
Wang, L., Zhuang, H., Zhang, Y., and Wei, W. (2019). Diversity of the Bosmina (Cladocera: Bosminidae) in China, revealed by analysis of two genetic markers (mtDNA 16S and a nuclear ITS). BMC Evol. Biol. 19:145. doi: 10.1186/s12862-019-1474-4
Yang, X.. (2011). Research on the Numbers, Distribution, Feeding Behavior and Diet of Great white Fronted Geese at Shengjin Lake, the National Natural Reserves in Anhui Province. Dissertation, University of Science and Technology of China, Hefei, China.
Yang, Y., Li, W., Zhou, Z., and Li, C. (2019). The study on the relationship between wetland landscape pattern and water level in Shengjin Lake. J. Biol. 36, 61–64. doi: 10.3969/j.issn.2095-1736.2019.02.061 (In Chinese)
Yang, G., Pan, H., Liu, Z., Wang, W., and Qin, B. (2008). A comparative study on seasonal variations of crustaceans in the different Lake areas in Lake Taihu. China Environ. Sci. 28, 27–32.
Yang, Z. F., Shi, W. G., Chen, L. Q., Chen, Y., and Zhou, Z. L. (2003). Ecological environment succession and countermeasure of east Taihu Lake. China Environ. Sci. 23, 64–68. (In Chinese)
Yao, S., and Xue, B.. (2016). The Evolution of Lake Environment in the Qingyi River in the Shuiyangjiang River Valley in the Lower Reaches of the Yangtze River. Nanjing: Nanjing University Press, 7–202.
Zaret, R., and Kerfoot, C. (1975). Fish predation on Bosmina-longirostris-body-size selection versus visibility selection. Ecology 56, 232–237. doi: 10.2307/1935317
Zhang, M., Shi, X., Yang, Z., Yu, Y., Shi, L., and Qin, B. (2018). Long-term dynamics and drivers of phytoplankton biomass in eutrophic Lake Taihu. Sci. Total Environ. 645, 876–886. doi: 10.1016/j.scitotenv.2018.07.220
Zhang, K., Yang, X., Xu, M., Lin, Q., Kattel, G., and Shen, J. (2018). Confronting challenges of managing degraded lake ecosystems in the Anthropocene, exemplified from the Yangtze River basin in China. Anthropocene 24, 30–39. doi: 10.1016/j.ancene.2018.11.001
Zhong, J., Liu, M., Wang, Y., Yang, X., Jiang, X., and Xu, C. (2014). Spatial correlation of major water quality indices between the Lake and rivers in Lake Taihu basin. Chinese J. Ecol. 33, 2176–2182.
Keywords: subtropical shallow freshwater lake, zooplankton, Bosminidae, eutrophication, ecological changes
Citation: Cheng L, Gao X, Wang G, Ding Z, Xue B, Zhang C, Liu J and Jiang Q (2023) Intensified sensitivity and adaptability of zooplankton Bosminidae in subtropical shallow freshwater lakes with increasing trophic level. Front. Ecol. Evol. 11:1121632. doi: 10.3389/fevo.2023.1121632
Edited by:
Kunshan Bao, South China Normal University, ChinaReviewed by:
Yuan Li, Chinese Academy of Sciences, ChinaXu Chen, China University of Geosciences Wuhan, China
Copyright © 2023 Cheng, Gao, Wang, Ding, Xue, Zhang, Liu and Jiang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bin Xue, bxue@niglas.ac.cn; Can Zhang, czhang@niglas.ac.cn