 Yumeng Tian
Yumeng Tian Ping Li
Ping Li- Department of Endocrinology, Shengjing Hospital of China Medical University, China Medical University, Shenyang, China
Diabetes mellitus is a chronic disease caused by the interaction of genetics and the environment that can lead to chronic damage to many organ systems. Genome-wide association studies have identified accumulating single-nucleotide polymorphisms related to type 2 diabetes mellitus and gestational diabetes mellitus. Genetic risk score (GRS) has been utilized to evaluate the incidence risk to improve prediction and optimize treatments. This article reviews the research progress in the use of the GRS in diabetes mellitus in recent years and discusses future prospects.
Gestational diabetes mellitus (GDM) refers to varying degrees of glucose intolerance that first appear during pregnancy (1). It has been associated with poor maternal and offspring outcomes, including preeclampsia, polyhydramnios, operative delivery, shoulder dystocia, birth canal lacerations, and fetal overgrowth (also called macrosomia) (2). The patient’s blood glucose is normal before pregnancy, but during pregnancy, blood glucose quietly and gradually increases, which may cause adverse effects on the mother and the baby before diagnosis and recognition. Therefore, early prediction of GDM has been explored to facilitate the prevention, early diagnosis, and treatment of the disease. Currently, the pathogenesis of GDM is believed to be closely related to the glycemic effect of placental hormones, but some studies have shown that GDM has an obvious genetic background and a similar genetic pathway with type 2 diabetes mellitus (T2DM) (3, 4). Therefore, the genetic association between T2DM and GDM has been analyzed in recent years to seek breakthroughs. Genome-wide association studies (GWAS) are one way to do that. GWAS have analyzed at least hundreds of single-nucleotide polymorphisms (SNPs) to determine their associations with complex clinical conditions and phenotypes. Similar to type 2 diabetes, GDM is closely related to both genetic and environmental factors, each of which may increase the risk of developing the disease (5). The effect of a single or small number of SNPs is too weak to accurately predict the disease; thus, the cumulative effect of more SNPs needs to be determined. The genetic risk score (GRS) is not only a common strategy but also a new stage in the detection of genetic susceptibility to complex diseases. The GRS is a useful gene analysis strategy that integrates genetic information from multiple loci to predict an individual’s likelihood of developing a disease (or a specific clinical trait) and to assess the effectiveness of associated variations in predicting the disease. This paper reviews the research status and the application value of the GRS in GDM by summarizing the progress in domestic and foreign research. This paper aims to help clinicians understand the role of genetic background in the pathogenesis of GDM and explore the value of the GRS in the diagnosis and treatment strategy of GDM.
Introduction to GRS
A GRS is used to evaluate the effect of genetic susceptibility factors in risk prediction models. A GRS allows for the assessment of the contributions of numerous factors to disease development and outcomes, including disease susceptibility, progression, and response to treatment (6). At present, the clinical application of such evaluations is the polygenetic risk score (PRS), which can condense a large amount of genomic variation information into scores that can measure individual susceptibility, calculated as a totality of their genome-wide genotypes and (or) weighted by the corresponding genotype effect size estimates based on GWAS statistical data (7). A PRS is an extension of a large number of GRSs that may be related to markers. It is aimed to predict all genetic variations measured by correlated markers (6). Taking T2DM as an example, the purpose of constructing a GRS model and risk score for the T2DM population is divided into two aspects: 1) to estimate the risk of an individual developing T2DM based on existing information, usually genetic, clinical, demographic, or a combination, or to predict risk only for a disease characteristic, such as insulin resistance, simple fasting blood glucose, and HbA1c; and 2) to assess the predictive power of a single SNP for the risk of T2DM and thus establish the contribution of this SNP to the etiology of T2DM. The prediction accuracy of the GRS is most often valued by measuring the area under the curve (AUC) of the receiving operator characteristic, an indicator of model accuracy (6).
The methods that most studies used included simple count GRS [SC-GRS, sGRS, or unweighted GRS (uGRS)], weighted GRS (wGRS), pruning and thresholding (P + T), LDpred GRS, GraBLD GRS, and direct logistic regression GRS (DL-GRS). The SC-GRS and wGRS are the most commonly used. The wGRS is calculated based on the number of risk alleles weighed by their effect sizes reported in previous GWAS. The SC-GRS is calculated as the sum of the risk alleles (0, 1, 2) of each of the variants. The formulas are shown in Table 1.

Table 1 The formulas of the three most commonly used GRS.
GRS used in studies on T2DM
GRS for predicting the risk for T2DM
GWAS have increasingly identified more SNPs that are related to the risk factors of T2D. Considerable research has demonstrated that the GRS improves the accuracy of T2D risk prediction, including research conducted in Europe, Asia, and North America. A study showed that GRST2D, fasting plasma glucose GRS (GRSFPG), insulin secretion GRS (GRSIS), and 2-h plasma glucose GRS (GRS2hPG) were associated with increased glucose AUC, increased FPG, decreased insulin secretion, and increased 2hPG, respectively (p< 0.0017 for all models) (8). Miranda-Lora et al. found that for non-diabetic Mexican Americans, the beta-cell GRS was related to reduced insulinogenic index, the insulin sensitivity GRS was related to the corresponding effect, the lipodystrophy GRS was related to reduced adiposity, and the body mass index plus lipid GRS was related to increased insulin clearance. All GRSs were strongly associated with T2D (9). Another study showed that the GRS of the associated variants of NF-E2-related factor 2 expression is prone to be a useful indicator of T2D development in the normal human population. In addition, linear regression analyses showed positive associations between the GRS and fasting glucose (p-value = 0.028, β = 0.62), 2-h glucose (p-value = 0.0004, β = 1.13), and HbA1c (p-value = 0.033, β = 0.03) (10). Inaishi et al. revealed that among pediatric-onset T2D cases, more than 95% of the cases had six or more risk alleles, the average GRS was higher compared with the controls, and the joint construction of the model may have a highly suggestive effect on the risk of T2D (11). Furthermore, the erythrocyte phospholipid ALA and unweighted GRS can jointly predict the risk of T2D in a Chinese population (12).
Some studies have combined GRSs with other risk factors to predict T2D. For instance, one study found a significant interaction between family history (FH) and lifestyle factors, and T2D genetic risk combined with the GRS and FH had the strongest interaction with lifestyle scores (13). There is a relationship observed between the beta-cell dysfunction GRS and young onset age with the risk of glycemic progression among people with type 2 diabetes, even though a linear relationship was observed between the GRS and the risk of glycemic progression (14). A multiple GRS (AUC: 0.58; p = 1.37 × 10−17) presented a greater AUC for T2D prevalence in non-obese than in obese individuals (AUC: 0.53; p = 1.4 × 10−4). At the same time, the obGRS showed a greater AUC for T2D in obese subjects (15). Likewise, Chikowore et al. found that the GRS, which consisted of variants that had significant associations in their population, was significantly associated with increased T2D risk as indicated by an OR of 1.21 (1.02–1.43). Due to the limitations of the GRS in predicting the risk of T2D, there are few clinical applications in the black population of southern Africa (16).
GRS for predicting the risk of developing T2DM complications
Moreover, GRSs can be used to predict T2D complications and explore their associations with other diseases. A study used the GRS to assess the T2D susceptibility limit based on 41 of the best-known T2D susceptibility variants. Cumulatively, subjects with higher GRSs had an approximately 40% higher risk of developing severe coronary artery disease (CAD) compared with those with lower scores (17). The APN level and the GRS were two independent risk factors for diabetic retinopathy. The GRS was calculated based on 10 SNPs for each subject, and the cumulative effect of genes was observed (18). Another study found that the higher the mother’s GRS, the higher the offspring’s risk of weight gain (19). These studies indicate that the GRS plays a certain role in the prediction of complications of T2D.
GRS used in studies on GDM
The genetic background of GDM is similar to that of T2D, so the SNPs related to the two diseases and the constructed GRS will be inevitably overlapped. However, as a special period of the body, the abnormal blood glucose metabolism is associated with not only the mother’s diabetes susceptibility but also the secretion of the various hormones from the placenta, which help to maintain blood glucose. Also, the mother’s changes in weight, blood lipid, and intestinal flora and obstetric complications in pregnancy are all closely related to maternal blood glucose metabolism (2).
Therefore, the pathogenesis of GDM is more complicated, and the SNPs involved and the GRS constructed therefrom are also more diversified, forming a genetic background different from T2D. In this paper, the research progress in this field during recent years will be described.
Relationship between GRS and the risk of GDM
As shown in Table 2, Lamri et al. found that for GDM patients in South Asia, the predictive effect of the PRS can increase by integrating GWAS data, and extracting aggregate statistics from an enormous multi-ethnic genome-wide meta-analysis. Participants with the highest PRSs had an increased risk of GDM compared with the other groups (20). Another study used the SC-GRS and DL-GRS to investigate the association between SNPs and the identified risk factors [age and body mass index (BMI)]. The study showed that the GRS of GDM pregnant women was significantly higher than that of the control group (p< 0.001) (21). Moen et al. found that SNPs, which have been proven to be associated with hyperglycemia in Norway’s non-pregnant population, also had the ability to predict the risk of hyperglycemia during pregnancy in the pregnant population by analyzing the known and theoretically related variation of the GRS. Furthermore, SNPs, which have been shown to be associated with glucose metabolism parameters in non-pregnant women, are also related to the same glucose metabolism parameters in pregnant women in the pregnant population, albeit with low statistical power (22).
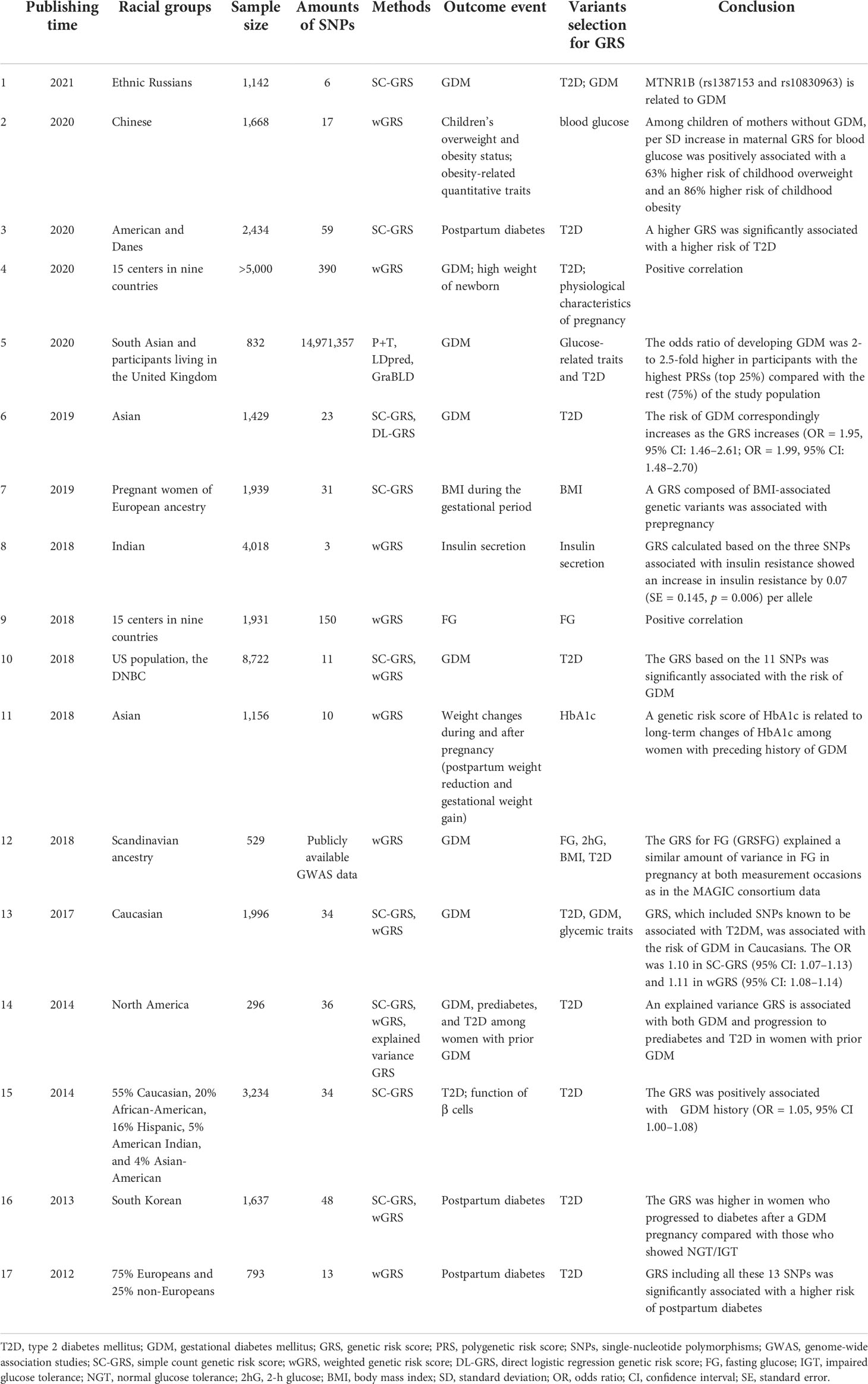
Table 2 Genetic risk score in the studies on GDM.
Kwak et al. found that the uGRS and wGRS were robustly associated with clinical parameters such as the effect of insulin treatment, fasting glucose, glucose AUC, and decreased insulin AUC. Moreover, they were significantly associated with a decreased insulin secretion/insulin resistance disposition index, which indicated impaired insulin secretory capacity (β = −0.025; p = 7.5 × 10−5). The wGRS can increase predictive power combined with other risk factors. Although the GRS has a limited value in identifying GDM at the population level in general, it can more precisely stratify women’s risk of GDM (23). The GRS could show genetic information of women at high risk for GDM to improve interventions so as to reduce the impact of metabolic disorders in pregnancy (24). A study identified eight variants associated with GDM based on the GRS (25). Nevertheless, a study showed that the GRS expresses a limited value in the identification of GDM cases (26).
Relationship between GRS and the risk of T2DM in women with a history of GDM
A meta-analysis reported that women with a prior history of GDM had a more than sevenfold increased risk of developing T2DM 10 years after delivery compared with those with normal blood glucose (27). A study with long-term follow-up of two independent populations of White women with a history of GDM showed that the higher the GRS, the higher the T2D risk. Furthermore, the association may be related to dietary quality; the association between GRSs and the risk of T2D was stronger in pregnant women with a poor diet (28). Kwak et al. found that among women with a history of GDM, the GRSs of women who progressed to diabetes 1 year after delivery were significantly higher than those of women who progressed to normal or impaired glucose tolerance. Incorporating the wGRS into the model made up of risk factors such as age can improve the predictive power of the model (24). A GRS consisting of three previously reported SNPs associated with insulin secretion was not associated with the risk of GDM. Notably, the diagnostic criteria of insulin resistance are the WHO 2013 criteria (29). In women diagnosed with prediabetes, a GRS containing 34 diabetes-related loci was used to assess the genetic risk of diabetes, and it was found that women with a history of GDM had a significantly higher GRS than women without GDM (30).
Relationship between GRS and adverse pregnancy outcomes
In addition, the GRS is associated with adverse pregnancy outcomes. Song et al. found that in the offspring of mothers with normal blood glucose during pregnancy, the risk of overweight and obesity increased by 63% and 86%, respectively, for each SD increase in the GRS. However, in children of GDM mothers, the same GRS model did not predict the risk of offspring being overweight or obese. The authors also found similarly significant interactions between genetically determined maternal blood glucose levels and GDM status on other obesity-related outcomes in children, such as body fat percentage (31). Han et al. found that compared with women with higher GRSs, women with lower GRSs showed more weight loss and better HbA1c improvement after delivery (32).
Most common SNPs used in the GRS studies on GDM
To perform a comprehensive review, we searched PubMed using the terms “SNP” OR “Single nucleotide polymorphism” AND “gestational diabetes mellitus” OR “GDM,” “Genetic risk score” OR “GRS” OR “polygenetic risk score” OR “PRS” OR “risk score” AND “gestational diabetes mellitus” OR “GD” and included articles from January 2012 to May 2021. After excluding unrelated articles, only 17 reviews were included. We retrieved these articles that investigated the correlation between the GRS and GDM, from which we extracted all SNPs used to build the GRS model for each study. We sorted them according to the occurrence or frequency of each gene and found that the top 4 “high-frequency genes” most commonly used to build the GRS were SLC30A8, TCF7L2, MTNR1B, and KCNJ11.
Transcription factor 7-like 2
Transcription factor 7-like 2 (TCF7L2) is a key transcriptional effector of the Wnt/beta-catenin signaling pathway, an important developmental signaling pathway that negatively regulates fat formation. It regulates insulin secretion and islet beta-cell proliferation/apoptosis and is a key gene in the regulation of glucose homeostasis (33).
Women carrying the TCF7L2 rs7903146 risk alleles exhibit impaired insulin secretion and proinsulin conversion compared with non-carrier women (34). It is reported that even after adjusting for confounding factors such as BMI and age, the T risk allele in TCF7L2 rs7903146 is associated with early postprandial glucose control failure and insulin treatment needs in women with gestational diabetes (35). The genetic variant rs7903146 (C > T) in TCF7L2 presents a strong association with GDM risk (36). Another article presented evidence that TCF7L2 rs290487, rs6585194, and rs7094463 polymorphisms were associated with insulin resistance and insulin secretion in women with GDM (37). Franzago et al. demonstrated an important correlation between the TCF7L2 rs7903146 variant and GDM with a more than fivefold risk in the TT genotype (38).
On the contrary, another study found no statistically significant difference in glucose, cholesterol levels, and different TCF7L2 SNP alleles in the GDM group. It is reported that TCF7L2 SNPs are not different among women with GDM, although significantly higher incidences of TCF7L2 rs7901695 SNP CC/CT, rs7903146 SNP CT/TT, and rs12255372 GT/TT were observed compared with the general female population (39). Similarly, Anghebem-Oliveira et al. found that TCF7L2 rs7901695 was not associated with GDM in a Brazilian population (40).
Melatonin receptor 1B gene
MTNR1B, the melatonin receptor 1B gene, encodes one of the receptors for melatonin and not only regulates circadian rhythm but also plays an important role in glucose metabolism, which is considered a bridge between circadian rhythm regulation and glucose metabolism (41).
Research has identified some SNPs utilizing GRS, including the risk SNPs rs10830963 (22, 25, 29, 32), rs1387153 (19, 22, 29), 2166706 (19), and rs7936247 (22) and two protective SNPs (rs1447352 and rs4753426) (19). Of these SNPs, rs10830963 is the most studied. A study indicated that MTNR1B rs10830963 is associated with GDM susceptibility and that women carrying the G allele have an increased risk of developing GDM (42). Among Chinese women with a history of GDM, Nisa et al. found that there was a bidirectional effect between MTNR1B gene variation and gestational weight gain in 2-h OGTT changes 1–5 years after delivery (43). It is suggested that the risk of GDM correlated with the rs10830963 risk allele G cannot be changed by other successful lifestyle interventions (44). Another article revealed that rs10830963, rs1387153, and rs2166706 interact with GDM risk in a southern Chinese population. rs1447352 and rs4753426, in particular, are associated with the reduction of the risk of GDM (45).
Another study revealed a strong association between rs1387153 and rs10830963 with GDM susceptibility in the dominant genotype (p = 0.006 and 0.007, respectively) and allelic (p = 0.008 and 0.013, respectively) models. Rosta et al. found that the MTNR1B rs10830963/G allele had the most robust association with GDM, as well as with glycemic traits, including both the FPG and the post-challenge (2 h) PG values at 75 g OGTT (46). Li et al. found that MTNR1B rs10830963 and its protein expression levels in the placenta are associated with an increased risk of developing GDM. Then, it may be a genetic factor leading to insulin resistance in Han Chinese women with GDM (47). The findings of another study suggested that rs10830963 and rs7936247 may be markers for susceptibility to GDM in a Chinese population (48). Firneisz et al. found that the rs10830963/G risk allele in MTNR1B was associated with a notable increase in the OR for antenatal insulin therapy initiation (OR = 5.2) in Hungarian women with GDM and pre-pregnancy BMI ≥29 kg/m2 (49).
Zinc transporter-8 gene
The zinc transporter-8 gene (SLC30A8), which encodes insulin gland-specific zinc transporter-8, is an important factor in the regulation of insulin secretion in islet cells that can promote zinc accumulation in the vesicles of islet β cells. A study showed that in women with gestational weight gain of GDM, SLC30A8 rs13266634 has a certain influence on long-term blood glucose changes, and women carrying the rs13266634 C allele will be limited in weight gain during pregnancy, which will reduce the occurrence of postpartum hyperglycemia (50). In the population of the district of Lund, the rs13266634 C allele in SLC30A8 was associated with an increased risk of developing GDM (51). Another study suggested that SLC30A8 rs13266634/T is a protective variant against the development of GDM among European women (46). SLC30A8 rs13266634/T also showed significant associations with GDM among Filipinos (52). Furthermore, SLC30A8 rs3802177 was also significantly associated with the risk of GDM (25). In Han Chinese, the CC genotype of SLC30A8 rs2466293 was significantly correlated with an increased risk of GDM with an OR of 1.455 (95% CI: 1.077, 1.966; p-value = 0.014) after adjustment for age. It was also significantly associated with FPG and 2hPG during OGTT (53).
Potassium channel inwardly rectifying subfamily J member 1
Potassium channel inwardly rectifying subfamily J member 11, also called KCNJ11, located on chromosome 11p15.1, is an open-reading frame encoding an inward rectifying potassium channel Kir6.2 protein composed of 390 amino acids. It is important for insulin secretion (54). A study revealed that KCNJ11 rs5219 was associated with GDM (OR = 1.15; 95% CI: 1.06, 1.24; p = 0.0004) (41). Another meta-analysis presented that the T allele of rs5219 was associated with an increased risk of GDM (pooled OR = 1.15; 95% CI: 1.06, 1.26) (55). A study showed that in Chinese women, gestational glycemic traits, such as HOMA-IR, insulin disposition index, early-phase insulin release, and 2-h postprandial proinsulin conversion, were associated with the risk allele of KCNJ11 rs5219 (p = 0.001, 0.006, 0.001, and 4.2 × 10−12, respectively) (56). KCNJ11 rs5215 is related to the development of postpartum abnormal glucose tolerance in Japanese women (OR = 1.82, 95% CI: 1.05, 3.14; p = 0.032) (57). However, no evidence was found on the association between KCNJ11 rs5219 and susceptibility to GDM or to other relevant metabolic features in a Greek population (58). Similarly, a non-significant association was found between KCNJ11 and the risk of GDM in a Chinese population (59).
Conclusion and future prospects
As GDM has become a common but difficult complication of pregnancy, it has received increasing research attention. Its harm to the puerpera and newborns cannot be ignored. Early detection and prevention are key to managing GDM. The GRS provides a way to solve this problem. While a large number of studies have shown that for such complex diseases, GDM-related risk genes can be identified through genetic methods such as GWAS and GRSs, the predictive value of relevant prediction models is limited; more clinical factors and indicators may need to be combined to obtain sufficient results. Some pregnant women develop GDM even without traditional risk factors such as family history. Therefore, the combinations of clinical characteristics that can build a reliable prediction model of GDM-related diseases are an urgent research problem to be explored and solved. The studies described in this article used GRSs to predict the development of T2D and GDM, determine their associations with disease diagnosis and prognosis, provide new evidence for risk assessment and early diagnosis of GDM, and identify their adverse consequences. However, it is shown that the development of GRSs is associated with some risks, such as incorrect risk estimation for individuals, failure to convey uncertainty in the assessment, and aggravating genetic discrimination (60). The cost-effectiveness of carrying out GRSs is also not negligible. Currently, most GWAS are conducted using Caucasian individuals, the utilization of these SNPs in other populations may result in a poor predictive power of GRSs, and then LD differences may be a major challenge to overcome (61).
Utilizing GRSs appropriately is full of challenges and opportunities. Improving the diagnosis and treatment of GDM solely based on the genetic background is insufficient; thus, the accuracy of predictions can be improved by combining other known risk factors, such as age and diet, among others. The gaps in GRS research need to be filled in order to use GRSs more effectively and safely before they can be used on a large scale. The epidemiology of large population-based cohort studies and the evaluation of the effectiveness of predictions in large populations will advance personalized medicine and ultimately improve health among susceptible populations.
Author contributions
YT wrote the first draft of the manuscript and edited it. PL performed critical revision of the literature and edited the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This article was funded by the Natural Science Foundation of Liaoning Province (2020-MS-153).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
GRS, genetic risk score; PRS, polygenetic risk score; SNPs, single-nucleotide polymorphisms; GWAS, genome-wide association studies; SC-GRS, simple count genetic risk score; OR, odds ratio; P + T, pruning and thresholding; DL-GRS, direct logistic regression GRS; 2hG, 2-h glucose; LD, linkage disequilibrium; FG, fasting glucose; IGT, impaired glucose tolerance; NGT, normal glucose tolerance; BMI, body mass index; SD, standard deviation; CI, confidence interval; UGRS, unweighted GRS; GraBLD GRS, gradient boosted and linkage disequilibrium adjusted GRS; SE, standard error.
References
1. American diabetes association. Classification and diagnosis of diabetes: Standards of medical care in diabetes. Diabetes Care (2019) 42(Suppl 1):S13–28. doi: 10.2337/dc19-S002
2. David McIntyre H, Catalano P, Zhang C, Desoye G, Mathiesen ER, Damm P, et al. Gestational diabetes mellitus. Nat Rev Dis Primers (2019) 5(1):47. doi: 10.1038/s41572-019-0098-8
3. Bao W, Yeung E, Tobias DK, Hu FB, Vaag AA, Chavarro JE, et al. Long-term risk of type 2 diabetes mellitus in relation to BMI and weight change among women with a history of gestational diabetes mellitus: a prospective cohort study. Diabetologia (2015) 58(6):1212–9. doi: 10.1007/s00125-015-3537-4
4. Shaat N, Ekelund M, Lernmark A, Ivarsson S, Almgren P, Berntorp K, et al. Association of the E23K polymorphism in the KCNJ11 gene with gestational diabetes mellitus. Diabetologia (2005) 48(12):2544–51. doi: 10.1007/s00125-005-0035-0
5. Welter D, MacArthur J, Morales J, Burdett T, Hall P, Junkins H, et al. The NHGRI GWAS catalog, a curated resource of SNP-trait associations. Nucleic Acids Res (2014) 42:D1001–6. doi: 10.1093/nar/gkt1229
6. Igo RP Jr., Kinzy TG, Cooke Bailey JN. Genetic risk scores. Curr Protoc Hum Genet (2019) 104(1):e95. doi: 10.1002/cphg.95
7. Choi SW, Mak TS, O'Reilly PF. Tutorial: a guide to performing polygenic risk score analyses. Nat Protoc (2020) 15(9):2759–72. doi: 10.1038/s41596-020-0353-1
8. Stančáková A, Kuulasmaa T, Kuusisto J, Mohlke KL, Collins FS, Boehnke M, et al. Genetic risk scores in the prediction of plasma glucose, impaired insulin secretion, insulin resistance and incident type 2 diabetes in the METSIM study. Diabetologia (2017) 60(9):1722–30. doi: 10.1007/s00125-017-4313-4
9. Shin JH, Lee K, Shin J, Kang KD, Nho CW, Cho YS, et al. A genetic risk score improves the prediction of type 2 diabetes mellitus in Mexican youths but has lower predictive utility compared with non-genetic factors. Front Endocrinol (Lausanne) (2021) 12:647864. doi: 10.3389/fendo.2021.647864
10. Inaishi J, Hirakawa Y, Horikoshi M. Genetic risk score combining six genetic variants associated with the cellular NRF2 expression levels correlates with type 2 diabetes in the human population. Genes Genomics (2019) 41(5):537–45. doi: 10.1007/s13258-019-00791-0
11. Inaishi J, Hirakawa Y, Horikoshi M. Association between genetic risk and development of type 2 diabetes in a general Japanese population: The hisayama study. J Clin Endocrinol Metab (2019) 104(8):3213–22. doi: 10.1210/jc.2018-01782
12. Zheng J-S, Li K, Huang T, Chen Y, Xie H, Xu D, et al. Genetic risk score of nine type 2 diabetes risk variants that interact with erythrocyte phospholipid alpha-linolenic acid for type 2 diabetes in Chinese Hans: A case-control study. Nutrients (2017) 9(4):376. doi: 10.3390/nu9040376
13. Ding M, Ahmad S, Qi L, Hu Y, Bhupathiraju SN, Guasch-Ferré M, et al. Additive and multiplicative interactions between genetic risk score and family history and lifestyle in relation to risk of type 2 diabetes. Am J Epidemiol (2020) 189(5):445–60. doi: 10.1093/aje/kwz251
14. Liu J-J, Gurung RL, Liu S, Yiamunaa M, Lee J, Ang K, et al. Associations of young onset age and genetic risk of beta cell dysfunction with glycaemic progression in individuals with type 2 diabetes. Diabetes Metab (2021) 47(6):101238. doi: 10.1016/j.diabet.2021.101238
15. Corella D, Coltell O, Sorlí JV, Estruch R, Quiles L, Martínez-González MA, et al. Polymorphism of the transcription factor 7-like 2 gene (TCF7L2) interacts with obesity on type-2 diabetes in the PREDIMED study emphasizing the heterogeneity of genetic variants in type-2 diabetes risk prediction: Time for obesity-specific genetic risk scores. Nutrients (2016) 8(12):793. doi: 10.3390/nu8120793
16. Chikowore T, Zyl T, Feskens EJM, Conradie KR. Predictive utility of a genetic risk score of common variants associated with type 2 diabetes in a black south African population. Diabetes Res Clin Pract (2016) 122:1–8. doi: 10.1016/j.diabres.2016.09.019
17. Zheng QW, Jiang J, Huo Y, Chen DF. Genetic predisposition to type 2 diabetes is associated with severity of coronary artery disease in patients with acute coronary syndromes. Cardiovasc Diabetol (2019) 18(1):131. doi: 10.1186/s12933-019-0930-1
18. Liao W-L, Chen Y-H, Chen C-C, Huang Y-C, Lin H-J, Chen Y-T, et al. Effect of adiponectin level and genetic variation of its receptors on diabetic retinopathy: A case-control study. Med (Baltimore) (2019) 98(11):e14878. doi: 10.1097/MD.0000000000014878
19. Rahman ML, Shrestha D, Workalemahu T, Wu J, Zhu C, Zhang C, et al. Maternal and offspring genetic risk of type 2 diabetes and offspring birthweight among African ancestry populations. J Clin Endocrinol Metab (2019) 104(11):5032–42. doi: 10.1210/jc.2018-02756
20. Lamri A, Mao S, Desai D, Gupta M, Paré G, Anand SS. Fine-tuning of genome-wide polygenic risk scores and prediction of gestational diabetes in south Asian women. Sci Rep (2020) 10(1):8941. doi: 10.1038/s41598-020-65360-y
21. Shen Y, Yulong J, Li Y, Gu X, Wan G, Zhang P, et al. Genetic determinants of gestational diabetes mellitus: a case-control study in two independent populations. Acta Diabetol (2020) 57(7):843–52. doi: 10.1007/s00592-020-01485-w
22. Moen G-H, LeBlanc M, Sommer C, Prasad RB, Lekva T, Normann KR, et al. Genetic determinants of glucose levels in pregnancy: genetic risk scores analysis and GWAS in the Norwegian STORK cohort. Eur J Endocrinol (2018) 179(6):363–72. doi: 10.1530/EJE-18-0478
23. Kwak SH, Choi SH, Kim K, Jung HS, Cho YM, Lim S, et al. Prediction of type 2 diabetes in women with a history of gestational diabetes using a genetic risk score. Diabetologia (2013) 56(12):2556–63. doi: 10.1007/s00125-013-3059-x
24. Kawai VK, Levinson RT, Adefurin A, Kurnik D, Collier SP, Conway D, et al. A genetic risk score that includes common type 2 diabetes risk variants is associated with gestational diabetes. Clin Endocrinol (Oxf) (2017) 87(2):149–55. doi: 10.1111/cen.13356
25. Ding M, Chavarro J, Olsen S, Tkachuk AS, Vasukova E, Anopova AD, et al. Genetic variants of gestational diabetes mellitus: a study of 112 SNPs among 8722 women in two independent populations. Diabetologia (2018) 61(8):1758–68. doi: 10.1007/s00125-018-4637-8
26. Popova PV, Klyushina AA, Vasilyeva LB, Lin Y, Ley SH, Bao W, et al. Association of common genetic risk variants with gestational diabetes mellitus and their role in GDM prediction. Front Endocrinol (Lausanne) (2021) 12:628582. doi: 10.3389/fendo.2021.628582
27. Leanne B, Casas J, Hingorani AD, Williams D, et al. Type 2 diabetes mellitus after gestational diabetes: a systematic review and meta-analysis. Lancet (2009) 373(9677):1773–9. doi: 10.1016/S0140-6736(09)60731-5
28. Li M, Rahman ML, Wu J, Ding M, Chavarro JE, Lin Y, et al. Genetic factors and risk of type 2 diabetes among women with a history of gestational diabetes: findings from two independent populations. BMJ Open Diabetes Res Care (2020) 8(1):e000850. doi: 10.1136/bmjdrc-2019-000850
29. Arora GP, Almgren P, Brøns C, Thaman RG, Vaag AA, Groop L, et al. Association between genetic risk variants and glucose intolerance during pregnancy in north Indian women. BMC Med Genomics (2018) 11(1):64. doi: 10.1186/s12920-018-0380-8
30. Kong X, Xing X, Hong J, Zhang W, Yang X. Association of a type 2 diabetes genetic risk score with insulin secretion modulated by insulin sensitivity among Chinese Hans. Clin Genet (2017) 91(6):832–42. doi: 10.1111/cge.12817
31. Song Q, Wang L, Liu H, Liang Z, Chen Y, Sun D, et al. Maternal GDM status, genetically determined blood glucose, and offspring obesity risk: An observational study. Obes (Silver Spring) (2021) 29(1):204–12. doi: 10.1002/oby.23047
32. Han L, Duan D, Zhang S, Li W, Wang L, Liu H, et al. Effects of the interaction between glycated haemoglobin genetic risk score and postpartum weight reduction on glycaemic changes: A gene-weight interaction analysis. Diabetes Obes Metab (2018) 20(12):2733–9. doi: 10.1111/dom.13452
33. Dias S, Pheiffer C, Abrahams Y, Rheeder P, Adam S. Molecular biomarkers for gestational diabetes mellitus. Int J Mol Sci (2018) 19(10):2926. doi: 10.3390/ijms19102926
34. Fritsche L, Sarief M, Wagner R, Stefan N, Lehmann R, Häring H-U, et al. Genetic variation in TCF7L2 rs7903146 and history of GDM negatively and independently impact on diabetes-associated metabolic traits. Diabetes Res Clin Pract (2018) 146:251–7. doi: 10.1016/j.diabres.2018.11.003
35. Potasso L, Perakakis N, Lamprinou A, Polyzou E, Kassanos D, Peter A, et al. Clinical impact of the TCF7L2 gene rs7903146 type 2 diabetes mellitus risk polymorphism in women with gestational diabetes mellitus: Impaired glycemic control and increased need of insulin therapy. Exp Clin Endocrinol Diabetes (2020) 128(10):663–6. doi: 10.1055/a-1008-9223
36. Franzago M, Fraticelli F, Marchetti D, Celentano C, Liberati M, Stuppia L, et al. Nutrigenetic variants and cardio-metabolic risk in women with or without gestational diabetes. Diabetes Res Clin Pract (2018) 137:64–71. doi: 10.1016/j.diabres.2018.01.001
37. Ye D, Fei Y, Ling Q, Xu W, Zhang Z, Shu J, et al. Polymorphisms in TCF7L2 gene are associated with gestational diabetes mellitus in Chinese han population. Sci Rep (2016) 6:30686. doi: 10.1038/srep30686
38. Franzago M, Fraticelli F, Nicolucci A, Celentano C, Liberati M, Stuppia L, et al. Molecular analysis of a genetic variants panel related to nutrients and metabolism: Association with susceptibility to gestational diabetes and cardiometabolic risk in affected women. J Diabetes Res 2017. (2017) p:4612623. doi: 10.1155/2017/4612623
39. Fd A, Lesauskaite V, Tamosiunas A, Jasukaitiene A, Velickienė D. Genetic variants of TCF7L2 gene and its coherence with metabolic parameters in Lithuanian (Kaunas district) women population with previously diagnosed gestational diabetes mellitus compared to general population. Diabetes Res Clin Pract (2021) 172:108636. doi: 10.1016/j.diabres.2020.108636
40. Anghebem-Oliveira MI, Martins BR, Alberton D, Ramos EAS, Picheth G, Rego FGM. Type 2 diabetes-associated genetic variants of FTO, LEPR, PPARg, and TCF7L2 in gestational diabetes in a Brazilian population. Arch Endocrinol Metab (2017) 61(3):238–48. doi: 10.1590/2359-3997000000258
41. Hongyan M, Li Q, Shujun. G. Meta-analysis of the relationship between common type 2 diabetes risk gene variants with gestational diabetes mellitus. PloS One (2012) 7(9):e45882. doi: 10.1371/journal.pone.0045882
42. Tarnowski M, Malinowski D, Safranow K, Dziedziejko V, Pawlik A. MTNR1A and MTNR1B gene polymorphisms in women with gestational diabetes. Gynecol Endocrinol (2017) 33(5):395–8. doi: 10.1080/09513590.2016.1276556
43. Nisa H, Malinowski D, Safranow K, Dziedziejko V, Pawlik A. The circadian rhythm-related MTNR1B genotype, gestational weight gain, and postpartum glycemic changes. J Clin Endocrinol Metab (2018) 103(6):2284–90. doi: 10.1210/jc.2018-00071
44. Grotenfelt NE, Wasenius NS, Rönö K, Laivuori H, Stach-Lempinen B, Orho-Melander M, et al. Interaction between rs10830963 polymorphism in MTNR1B and lifestyle intervention on occurrence of gestational diabetes. Diabetologia (2016) 59(8):1655–8. doi: 10.1007/s00125-016-3989-1
45. Jia Y, Shen Y, Shi X, Gu X, Zhang P, Liu Y, et al. MTNR1B gene on susceptibility to gestational diabetes mellitus: a two-stage hospital-based study in southern China. Mol Genet Genomics (2020) 295(6):1369–78. doi: 10.1007/s00438-020-01706-5
46. Rosta K, Al-Aissa Z, Hadarits O, Harreiter J, Nádasdi Á, Kelemen F, et al. Association study with 77 SNPs confirms the robust role for the rs10830963/G of MTNR1B variant and identifies two novel associations in gestational diabetes mellitus development. PloS One (2017) 12(1):e0169781. doi: 10.1371/journal.pone.0169781
47. Li C, Zhou Y, Qiao B, Xu L, Li Y, Li C. Association between a melatonin receptor 1B genetic polymorphism and its protein expression in gestational diabetes mellitus. Reprod Sci (2018) 26(10):1382–8. doi: 10.1177/1933719118765983
48. Xie K, Zhang Y, Wen J, Chen T, Kong J, Zhang J, et al. Genetic predisposition to gestational glucose metabolism and gestational diabetes mellitus risk in a Chinese population. J Diabetes (2019) 11(11):869–77. doi: 10.1111/1753-0407.12923
49. Firneisz G, Rosta K, Al-Aissa Z, Hadarits O, Harreiter J, Nádasdi Á, et al. The MTNR1B rs10830963 variant in interaction with pre-pregnancy BMI is a pharmacogenetic marker for the initiation of antenatal insulin therapy in gestational diabetes mellitus. Int J Mol Sci (2018) 19(12):3734. doi: 10.3390/ijms19123734
50. Wang T, Liu H, Wang L, Huang T, Li W, Zheng Y, et al. Zinc-associated variant in SLC30A8 gene interacts with gestational weight gain on postpartum glycemic changes: a longitudinal study in women with prior gestational diabetes. Diabetes (2016) 65(12):3786–93. doi: 10.2337/db16-0730
51. Dereke J, Palmqvist S, Nilsson C, Landin-Olsson M, Hillman M. The prevalence and predictive value of the SLC30A8 R325W polymorphism and zinc transporter 8 autoantibodies in the development of GDM and postpartum type 1 diabetes. Endocrine (2016) 53(3):740–6. doi: 10.1007/s12020-016-0932-7
52. Benny P, Ahn HJ, Burlingame J, Lee M-J, Miller C, Chen J, et al. Genetic risk factors associated with gestational diabetes in a multi-ethnic population. PloS One (2021) 16(12):e0261137. doi: 10.1371/journal.pone.0261137
53. Wang X, Li W, Ma L, Ping F, Liu J, Wu X, et al. Investigation of miRNA-binding site variants and risk of gestational diabetes mellitus in Chinese pregnant women. Acta Diabetol (2017) 54(3):309–16. doi: 10.1007/s00592-017-0969-y
54. Jakub R, Szostak B, Machaj F, Pawlak A. The role of genetics and epigenetics in the pathogenesis of gestational diabetes mellitus. Ann Hum Genet (2020) 84(2):114–24. doi: 10.1111/ahg.12356
55. Zhang C, Bao W, Rong Y, Yang H, Bowers K, Yeung E, et al. Genetic variants and the risk of gestational diabetes mellitus: a systematic review. Hum Reprod Update. Jul-Aug (2013) 19(4):376–90. doi: 10.1093/humupd/dmt013
56. Liao S, Liu Y, Chen X, Tan Y, Mei J, Song W, et al. The impact of genetic variants for different physiological characterization of type 2 diabetes loci on gestational insulin signaling in nondiabetic pregnant Chinese women. Reprod Sci (2015) 22(11):1421–8. doi: 10.1177/1933719115580995
57. Kasuga Y, Miyakoshi K, Tajima A, Saisho Y, Ikenoue S, Ochiai D, et al. Clinical and genetic characteristics of abnormal glucose tolerance in Japanese women in the first year after gestational diabetes mellitus. J Diabetes Investig (2019) 10(3):817–26. doi: 10.1111/jdi.12935
58. Pappa KI, Gazouli M, Economou K, Daskalakis G, Anastasiou E, Anagnou NP, et al. Gestational diabetes mellitus shares polymorphisms of genes associated with insulin resistance and type 2 diabetes in the Greek population. Gynecol Endocrinol (2011) 27(4):267–72. doi: 10.3109/09513590.2010.490609
59. Wang K, Chen Q, Feng Y, Yang H, Wu W, Zhang P, et al. Single nucleotide polymorphisms in CDKAL1 gene are associated with risk of gestational diabetes mellitus in Chinese population. J Diabetes Res (2019) 2019:3618103. doi: 10.1155/2019/3618103
60. Polygenic risk score task force of the international common disease alliance, responsible use of polygenic risk scores in the clinic: potential benefits, risks and gaps. Net Med (2021) 27(11):1876–84. doi:10.1038/s41591-021-01549-6
Keywords: diabetes mellitus, type 2 diabetes mellitus, single-nucleotide polymorphism, genetic risk score, endocrinology
Citation: Tian Y and Li P (2022) Genetic risk score to improve prediction and treatment in gestational diabetes mellitus. Front. Endocrinol. 13:955821. doi: 10.3389/fendo.2022.955821
Received: 29 May 2022; Accepted: 29 September 2022;
Published: 19 October 2022.
Edited by:
Alok Raghav, Ganesh Shankar Vidyarthi Memorial Medical College, IndiaReviewed by:
Cheng Hu, Shanghai Jiao Tong University, ChinaYao Qiying, Dalian Medical University, China
Copyright © 2022 Tian and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ping Li, lipingsj@163.com