A Gut Feeling in Amyotrophic Lateral Sclerosis: Microbiome of Mice and Men
 Sarah Martin
Sarah Martin Carolina Battistini
Carolina Battistini Jun Sun
Jun Sun- 1Division of Gastroenterology and Hepatology, Department of Medicine, University of Illinois at Chicago, Chicago, IL, United States
- 2Department of Microbiology and Immunology, University of Illinois at Chicago, Chicago, IL, United States
- 3University of Illinois at Chicago (UIC) Cancer Center, University of Illinois at Chicago, Chicago, IL, United States
- 4Jesse Brown VA Medical Center, Chicago, IL, United States
Amyotrophic lateral sclerosis (ALS) is a severely debilitating disease characterized by progressive degeneration of motor neurons. ALS etiology and pathophysiology are not well understood. It could be the consequences of complex interactions among host factors, microbiome, and the environmental factors. Recent data suggest the novel roles of intestinal dysfunction and microbiota in ALS etiology and progression. Although microbiome may indeed play a critical role in ALS pathogenesis, studies implicating innate immunity and intestinal changes in early disease pathology are limited. The gastrointestinal symptoms in the ALS patients before their diagnosis are largely ignored in the current medical practice. This review aims to explore existing evidence of gastrointestinal symptoms and progress of microbiome in ALS pathogenesis from human and animal studies. We discuss dietary, metabolites, and possible therapeutic approaches by targeting intestinal function and microbiome. Finally, we evaluate existing evidence and identify gaps in the knowledge for future directions in ALS. It is essential to understanding the microbiome and intestinal pathogenesis that determine when, where, and whether microbiome and metabolites critical to ALS progression. These studies will help us to develop more accurate diagnosis and better treatment not only for this challenging disease, but also for other neurodegenerative diseases.
Introduction
ALS is a highly fatal neuromuscular disease currently with no cure. ALS has a median incidence of about 2.8 cases per 100,000 persons per year and a median prevalence about 5.4 cases per 100,000 persons for a median age at 61.8 ± 3.8 years (Chio et al., 2013). Although relatively rare, this public health impact of ALS is significant due to its morbidity and mortality.
The causes of ALS are largely unknown, only 10–15% of ALS cases are familial amyotrophic lateral sclerosis (fALS), with the remainder considered sporadic ALS (sALS) (Turner et al., 2017). Monogenic mutations in over 30 genes are associated with about ALS cases, including C9orf72, SOD1, FUS, and TARDBP/TDP43 (Nicolas et al., 2018; Volk et al., 2018). However, these monogenic forms explain only 15% of sporadic cases and 66% of familial cases (Turner et al., 2017). ALS pathogenesis is multifactorial and involves complex host-environment interactions. The risk factors include nutritional status, lower body mass index (BMI), smoking, high level of physical fitness, and military service (Wang et al., 2017). Environmental neurotoxins might contribute to ALS, e.g. β-Methylamino-L-alanine (BMAA) in people in Guam (Banack and Cox, 2003; Murch et al., 2004) and formaldehyde in factory workers (Weisskopf et al., 2009; Roberts et al., 2016).
Although ALS is a neurodegenerative disorder, progressive loss of motor neurons. Little is known about the gastrointestinal (GI) changes in ALS patients before and after the diagnosis, although autonomic dysfunction is reported in ALS. On one hand, in the literature, we see the comments that ALS patients do not complain of gastrointestinal symptoms and there is no GI symptom in ALS. On the other hand, there are cases that celiac disease with neurologic manifestations was misdiagnosed as ALS (Turner et al., 2007; Brown et al., 2010; Ham et al., 2017). There is report that gastrointestinal motor dysfunction occurred in ALS with delayed gastric emptying and delayed colonic transit times in patients (Wingate, 1999). Pancreatic and parotid deficiencies were observed in ALS patients (Charchaflie et al., 1974). There is a possible link between ALS and sensitivity to gluten based on an Israel study (Gadoth et al., 2015). In certain cases, an ALS syndrome might be associated with autoimmunity and gluten sensitivity. These reports suggest that ALS patients show GI symptoms, maybe at the early stage of their disease or before their diagnoses.
Growing evidence indicates that gut microbiome may actively contribute to ALS pathogenesis. ‘Microbiome’ includes all the microorganisms living (e.g., bacteria, virus, fungi) in the body. The definition of microbiome proposed by Whipps et al. is a broad one which describes microbiome as a “characteristic microbial community” in a “reasonably well-defined habitat which has distinct physio-chemical properties” as their “theatre of activity” (Whipps et al., 1988). Microbes inhabit the human body mediating key metabolic, physiological, and immune functions (Baquero and Nombela, 2012) and can be considered as another human organ. The last decade has witnessed a rebirth in interest in the microbes because of their novel and potential roles in disease pathogenesis and treatment. We are the first to report the elevated intestinal inflammation, reduced beneficial bacteria, and shift of microbiome profile in ALS (Wu et al., 2015; Rowin et al., 2017; Zhang et al., 2017; Zhang et al., 2021). Later, studies in human ALS and experimental animal models also reported the altered microbiome in the ALS (Fang, 2016; Labarre et al., 2017; Blacher et al., 2019; Figueroa-Romero et al., 2019; Burberry et al., 2020). Antibiotic usage, especially repeated use of antibiotics, are reported to be associated with a higher subsequent risk of ALS (Sun et al., 2019). The novel impact of gut microbiota on ALS development could explain some of the disease features beyond the influence of the genetic factor. However, microbiome studies in ALS are still very limited.
In the current review, we summarize the altered GI functions and roles of the gut microbiota in ALS, based on human data and experimental models. We discuss possible therapeutic approaches by targeting intestinal function and microbiome. Our goal is to evaluate existing evidence and identify gaps in the knowledge for future directions in ALS and other neurodegenerative diseases.
Gastrointestinal (GI) Disorders in Human ALS
Pancreatic function was studied in ALS patients. In a 1967 study (Quick and Greer, 1967) that examined the pancreatic function of 5 ALS patients and 5 patients with non-ALS neurological diseases, they found that all 5 ALS patients displayed elevations in leucine aminopeptidase. 4 of the 5 ALS patients had abnormal results to tolbutamide testing. In the 4 ALS patients that underwent secretin testing, there were abnormalities in the response the secretin. The results of this study suggested that there is a disturbance of the function of the exocrine gland in patients with ALS, a finding that was consistent across patients. This is believed to be the first study that observed changes with this gland, as previous studies had only observed alterations of the endocrine gland. This study raised the question if there is a direct relationship between ALS and the pancreas or if alterations of the pancreas are due to secondary complications.
Utterback et al. (1970) analyzed the pancreatic function of 10 ALS patients and 1 patient with chronic pancreatitis. They found that the patients experienced a variety of intestinal symptoms. One patient experienced small bowel abnormalities which consisted of segmentation and small bowel coarsening. Another patient underwent rapid transit time of the small bowel. A third patient experienced small sliding hernia while another had a potential duodenal ulcer. Yet another patient had varices and a traction diverticulum of the esophagus. The authors determined that the only system with abnormalities was endocrine function. They concluded that abnormalities with glucose function resulted in malnutrition, diminished physical activity, and decreased muscle mass. In another study looking at the pancreatic function of ALS patients, Charchaflie et al. (1974), found that ALS patients experienced an insufficient pancreatic exocrine response. Additionally, there was no pathological structural changes observed, specifically organic lesions. The ALS patients experienced a marked decrease in amylase concentration. Post stimulation with citric acid, the parotid secretory response was significantly lower in the ALS patients, compared to the control group (Charchaflie et al., 1974). Additionally, the bicarbonate concentration in the saliva was significantly lower in the ALS patients. There was no difference in plasma osmolarity between ALS patients and normal controls. These results suggest a functional impairment of the exocrine gland. The pancreatic deficiencies were potentially a result of modifications of neuroendocrine mechanisms. They also raised the question whether ALS pathogenesis could be a result of digestive gland functional insufficiencies. However, there is a report on no abnormal pancreatic, but abnormal in glucose utilization in ALS. The authors believed host factors (e.g., age, malnutrition, diminished physical activity, and decreased muscle mass) may explain the observed abnormal glucose utilization.
Wingate (1999) reported GI motor dysfunction with delayed gastric emptying and colonic transit times in patients with ALS (Wingate, 1999). Nubling et al. investigated disease severity, intestinal dysfunction, and lower urinary tract symptoms (LUTS) in 43 patients with ALS (Nubling et al., 2014). Urinary incontinence was increased in patients with ALS aged ≥ 60 years compared to the EPIC cohort. Intake of muscle relaxants and anticholinergics was associated with both urinary incontinence and severity of symptoms (Nubling et al., 2014). Furthermore, a high prevalence of constipation (46%), but stool incontinence was only reported in 9% of the group. Overall, the increased prevalence of urge incontinence and high GI symptom burden imply in patients with ALS. There are cases that celiac disease with neurologic manifestations was misdiagnosed as ALS (Turner et al., 2007; Brown et al., 2010; Ham et al., 2017). An ALS syndrome may be associated with autoimmunity and gluten sensitivity (Group, 2016).
In 1987, Nakano et al., found an increase in the size of pale hepatocytes, mild fatty infiltration, and variability in the nuclear size of these hepatocytes in ALS patients (Nakano et al., 1987). Additionally, fibrosis around the hepatocytes was present, as well as abnormalities of the hepatocellular mitochondria. This was one of the first studies to observe this fibrosis and was found in 76.2% of ALS patients versus 42.3% of controls. However, there was no relationship between these abnormal findings in the livers of ALS patients with the progression of the disease. This study suggest that abnormalities of the liver potentially occur in ALS and that the presence of these abnormalities would not be secondary to disease symptoms.
Jonsson et al., 2008 looked at the liver pathology from a single ALS patient in a case report. They found granular mutant SOD1 inclusions in the hepatocytes and kidney tubular epithelium and the absolute levels of G127X SOD1 were greater in the liver than in the temporal lobe. Staining of the hepatocytes was more widespread and uneven. The liver had inactive micronodular cirrhosis with some steatosis. Blood plasma levels indicated a moderate form of liver disease. From these results, the authors asked the question whether G127X SOD1 could contribute to these alterations to the liver.
The enteric nervous system (ENS) provides neural control in the gastrointestinal tract. When analyzing human ALS patient tissue, Luesma et al., 2014 (Luesma et al., 2014) found that there was evidence of the presence of adult enteric neurons stained with c-Ret (a RET kinase inhibitor) in the myenteric and submucosal ganglia of the adult duodenum. It suggested that c-Ret was necessary for the human mature ENS. Further studies would be needed to look at this connection with neurodegenerative diseases such as ALS.
Our study (Rowin et al., 2017) evaluated infection and markers of intestinal inflammation and the human gut microbiome in stool samples from ALS patients. A majority of patients had signs of intestinal inflammation. This is the first comprehensive examination of inflammatory markers in the stool of ALS patients. A previous study (Zhang et al., 2009) reported that the level of plasma lipopolysaccharides (LPS), a bacterial endotoxin, significant increased and had a positive correlation with activation of blood monocyte/macrophage in sALS groups, and LPS was most elevated in patients with advanced sALS disease (Zhang et al., 2009). Circulating endotoxin and systemic immune activation in sALS suggested intestinal leakage and local inflammation in these sALS patients.
Clearly, there are GI disorders in patients with ALS. We summarized the current literature in Table 1. These reports suggest that ALS patients show GI symptoms, maybe at the early stage of their disease or before their diagnoses. However, GI symptoms for pre-diagnosis of ALS are largely ignored. It is challenging to perform systemic studies before the diagnoses.
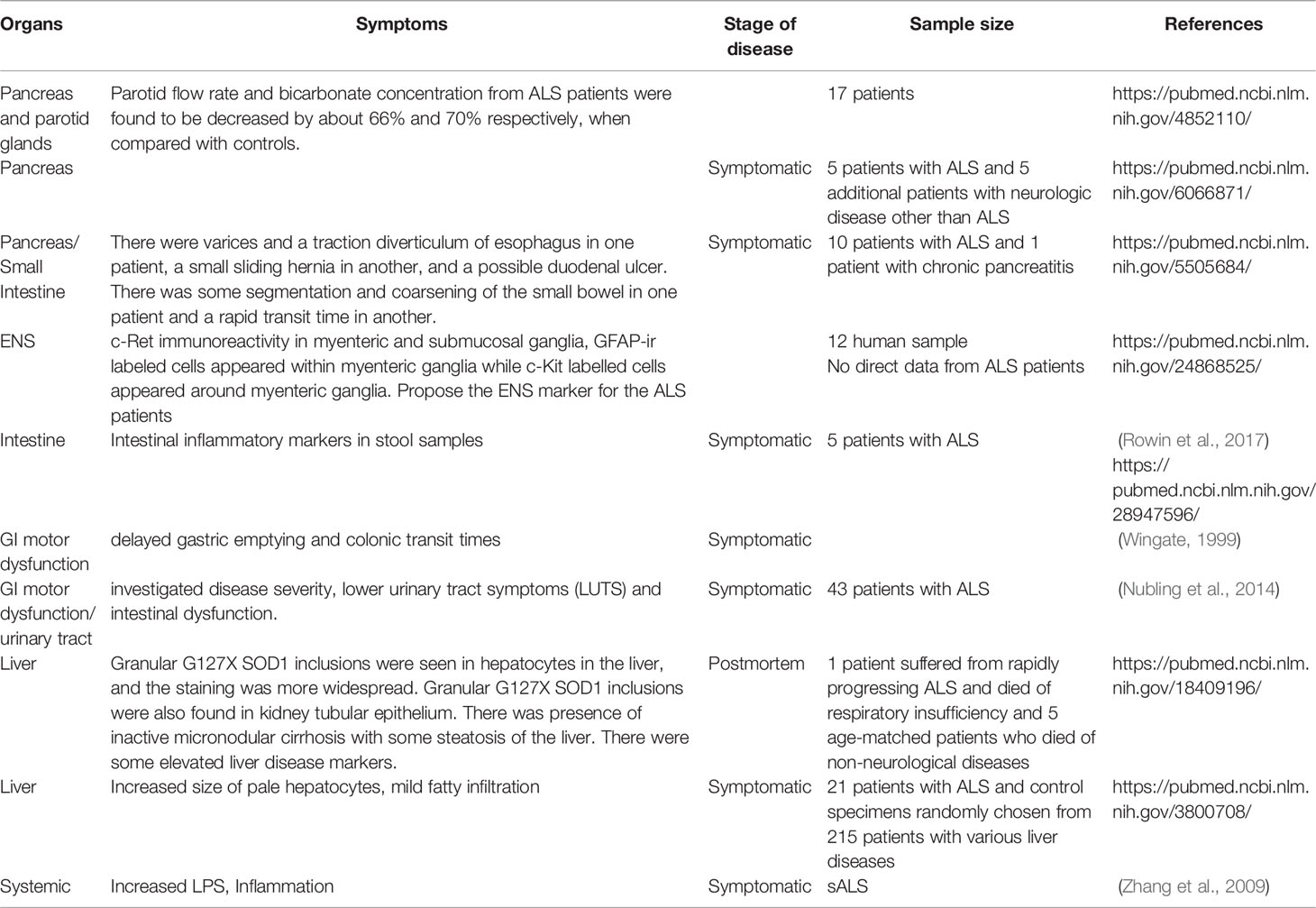
Table 1 Involved organs of GI tract in patients with ALS.
Gut Pathophysiology in ALS Mouse Models
Transgenic animals are expected to imitate the key features of ALS. There are different ALS experimental models, with their advantages and disadvantages (Philips and Rothstein, 2015). In patients with ALS, there are spinal cord and muscle pathology. About 10% of familial ALS cases are attributed to mutations in Cu, Zn superoxide dismutase (SOD). Normal SOD catalyzes the dismutation of superoxide radicals into hydrogen peroxide and molecular oxygen. The ALS pathology can be modeled in hSOD1G93A mice, which carry the human point mutant SOD1 G93A.
Similar to patients with ALS, the hSOD1G93A mice showed liver problems, e.g., hepatic abnormalities and lymphocytic infiltration (Lee and Yang, 2018). Lee and Yang (2018) investigated the liver of hSOD1G93A mice transgenic mice, changes in liver protein expression over the course of disease progression were found. This included a significant increase in inflammatory cytokines (TNF-α, IL-1β, and CD11b), oxidative stress-related proteins (NQ01 and ferritin), and regulators of cell death (Bax and pJNK). For pre-symptomatic mice hSOD1G93A mice, there was a significant increase in fibrosis-related proteins (HDAC4, GADD45α, and PDGFb) in the liver. These findings suggested that liver dysfunction may lead to liver fibrosis in hSOD1G93A mice. Additionally, liver dysfunction in hSOD1G93A mice could potentially be mediated by increased inflammation and oxidative stress, in addition to the upregulation of fibrosis-related proteins.
We found the damaged tight junctions (TJs) and increased intestinal permeability in the hSOD1G93A mice (Wu et al., 2015). TJs act as barriers in the intestinal epithelial cells to protect host from pathogens, inflammation, and other injuries (Zhang et al., 2013; Zhang et al., 2019). We studied hSOD1G93A and age matched wild type mice and found that TJ ZO-1 protein expression was significantly decreased in the intestine of G93A, compared to WT mice at 2-month-old. The G93A mice do not show ALS symptoms until 3 months old. The adherent junction protein E-cadherin was significantly decreased in the intestines of G93A mice, compared to the age-matched WT mice. ZO-1 was a tight junction protein restricted to cellular borders in a smooth and organized pattern at the apical side of the healthy colons. We identified abnormal and disrupted ZO-1 distribution in the membrane of intestinal epithelial cells in ALS mice at the age of 2 months. We also showed a significant increase in abnormal Paneth cells and increased inflammatory cytokine IL-17 in hSOD1G93A mice (Wu et al., 2015). As the changes in intestine function occurs early, it suggests the possibility that altered intestinal homeostasis may take place before the decline of neuromuscular function in ALS mice. This study demonstrated that impaired gut-neuromuscular crosstalk could potentially contribute to the progression of ALS.
Intestinal mobility is a key physiologic parameter governing digestion and absorption of nutrients affected by the ENS, microbiome, and host genetics (Rao and Gershon, 2016; Niesler et al., 2021). Few studies in general have been conducted looking at the connection between the ENS in the SOD1G39A ALS model. Our recent study (Zhang et al., 2021) reported the dysfunction of ENS in SOD1G39A. Before ALS onset, 2-month-old G93A mice had significant lower intestinal motility, decreased grip strength, and reduced time in the rotarod. We observed increased GFAP, a marker for ENS, and decreased SMMHC expression, a marker for smooth muscle (Zhang et al., 2021). These ENS changes are significantly correlated with increased aggregation of hSOD1G93A in the spinal cord, small intestine, and colon (Zhang et al., 2021).
In a 2011 study by Finkelstein et al, the authors discovered that the liver parenchyma underwent significant atrophy and lipid redistribution, while an increase in natural killer T cells in both the lymphoid organs and spinal cord of the mSOD1 mice compared to WT mice was also observed. While the liver itself decreased in size as the disease progressed, the proportions of natural killer T cells in the liver increased by 4-fold at the end-stage of the disease. In mSOD1 mice with the C57B1 background, the natural killer T cells had a similar fate. T cells were significantly enhanced in the spinal cord and spleen of mSOD1 mice. Additionally, natural killer cells were significantly enhanced in the spinal cord of mSOD1 mice, but not in the spleen and liver. This study showed that changes to liver are present during disease progression in transgenic mice and therapeutic interventions that target these changes are needed.
Some animal models may not always exhibit similar co-morbidity phenotypes to human patients with ALS. The transgenic mice overexpressing a mutant human TDP43 gene (Hatzipetros et al., 2014), the Prp-TDP43A315T mice, exhibited decreased survival rates that may be associated with neurodegeneration, occurring in the myenteric plexus of the colon. In these Prp-TDP43A315T mice, death coincided with severe GI pathology. In the lower GI tract, distal to the cecum, there were pathological changes, such as swelling, intraintestinal coagulated blood, and necrotic tissue. It is most likely a result of overexpression of the transgene in the lower GI tract, as opposed to the upper GI where there is no expression of the transgene and no neurodegeneration. The neurodegeneration of the lower GI tract is a feature that is unique to the Prp-TDP43A315T mice. Similar findings have been reported by others (Guo et al., 2012). TDP43 A315T mice exhibited not only a reduction in food ingestion and defecation, but also a thinned colon and swollen small intestine. The swollen small intestine and thinned large bowel also indicate peristaltic malfunction in the TDP43 A315T mice. In the colon, the accumulated TDP43 A315T in the myenteric nerve plexus and myenteric neuron degeneration could result in phenotypes similar to hypoganglionosis. These results, in addition to no limb paralysis present at the end stage of disease progression, suggested that intestinal dysfunction contributed more to their death than severe muscle weakness in the TDP43 A315T mice. The authors believed that while the TDP43 A315T model can be used to analyze GI tract degeneration, it did not display a phenotype that was acceptable to study ALS therapeutics.
Herdewyn et al. (2014) found a significant reduction in the ability of the transgenic TDP43 A315T mice to generate propulsive contractions. There was degeneration of nitric oxide synthase (NOS) (Fleck et al., 2021) neurons in the enteric nervous system, but no difference in choligeneric neurons. Because NOS and excitatory neurons both regulate intestinal peristalsis, lack of both excitation and inhibition would result in a failure to generate contractile force and the prevention of peristalsis. The degenerated NOS neurons suggested the loss of inhibitory control that led to abnormal intestinal propulsion, dysmotility and pseudo-obstruction, and sudden death (Herdewyn et al., 2014). There was downregulation of endogenous TDP-43 in the spinal cord and brain of the TDP43 A315T mice prior to the observation of neurodegeneration. The feeding of jellified food prevented intestinal obstruction, allowing the motor phenotype to develop, and significantly extending survival. These results suggested that the degeneration of NOS neurons in the myenteric plexus resulted in intestinal dysmotility in the TDP43 A315T mice. In yet another study using the TDP43 A315T model (Esmaeili et al., 2013), the authors found intestinal dilation with the caecum and lower ileum mildly to moderately dilated and contained dry, firm fecal material. One animal had a fibrous band at the ileocecocolic junction. The stomachs of the animals were all empty. The authors concluded that the animals were dying due to a reduction in motility that originates in the ileocaecal area of the GI tract. A limitation of this study is a general limit of the TDP43 A315T animal model itself –lacking lower motor neuron degeneration in the ventral horn of the spinal cord. Toxicity from TDP43 overexpression may lead to a reduction in GI motility, though it is still not fully understood how TDP-43 overexpression leads to GI tract abnormalities in the TDP43 A315T mice. These studies suggest that gastrointestinal complications may have been the cause of death for TDP43 A315T mice and not spinal motor neuron loss.
Studies in the animal models have shown the GI petrophysical changes before the motor neuron degeneration was observed. We summarized these findings in Table 2. Although the transgenic animal models cannot sufficiently represent the development of human ALS, especially the sALS, studies in these models still provide valuable data and mechanisms during the disease progression.
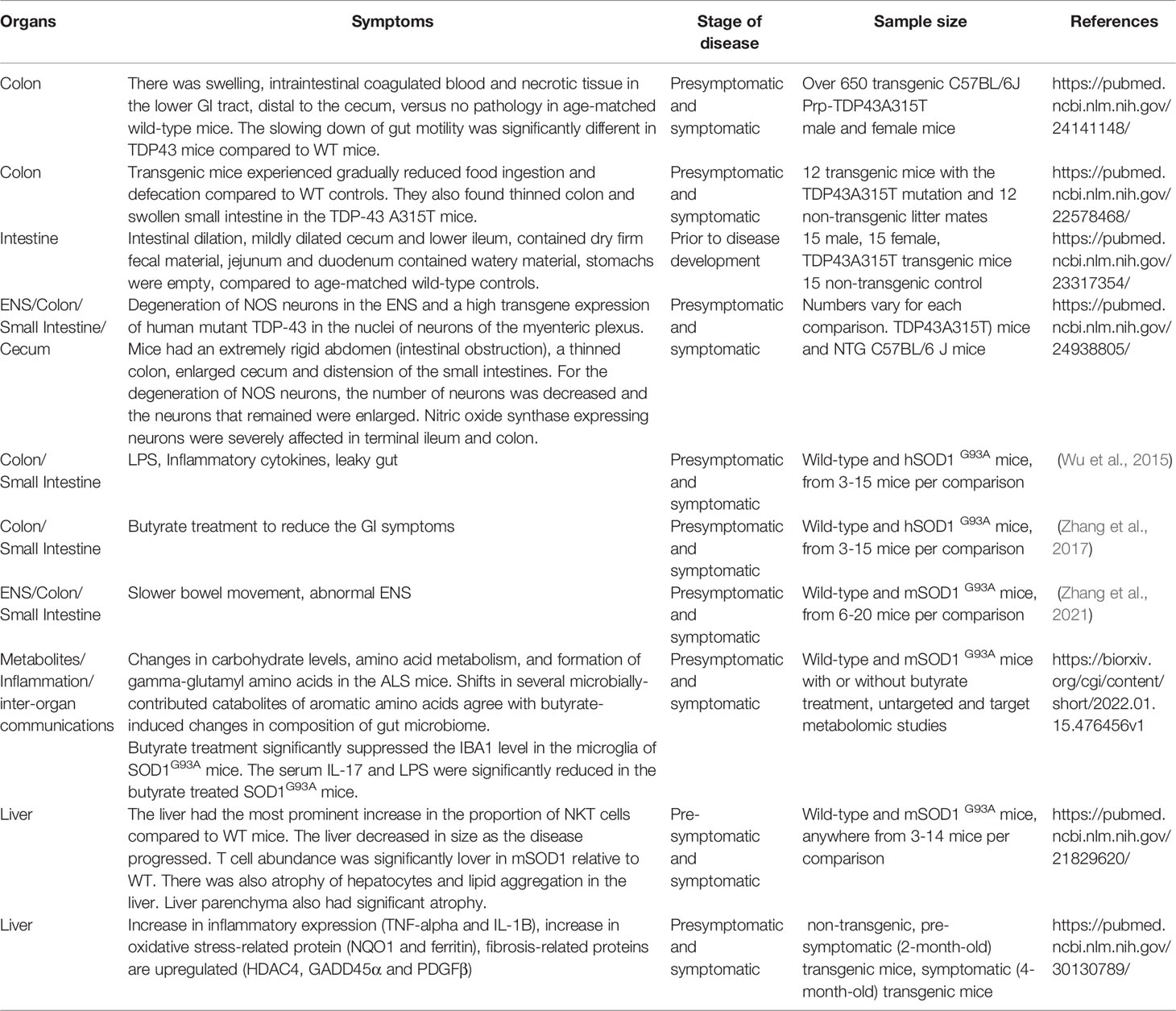
Table 2 Involved organs of GI tract in ALS animal models.
Altered Gut Microbiome in ALS Mouse Models
There are known active host-microbiome interactions in the GI tract. Our 2015 study has demonstrated the altered bacterial abundance, reduced levels of butyrate-producing bacteria in the ALS SOD1G93A animals (Wu et al., 2015). The intestinal microbiome was altered as butyrate producing bacteria (e.g., Butyrivibrio Fibrisolvens) were reduced in ALS mice at the age of 2 months before the onset of disease. The cellular and physiological changes mirror the population and function changes in the gut microbiome of the SOD1G93A mice. With an interventional study attempting to alleviate symptoms through treatment with sodium butyrate (Zhang et al., 2017). SOD1G93A mice chronically treated with sodium butyrate fared better than those not given the compound, showing improved intestinal barrier function and delays in both weight loss and death of 50 and 38 days respectively. Further longitudinal studies (Zhang et al., 2021) demonstrated the early stage microbial changes in the mice at 1-month old. The altered bacterial community is related to autoimmunity (e.g., Clostridium sp. ASF502, Lachnospiraceae bacterium A4), inflammation (e.g., Enterohabdus Muris), and metabolism (e.g., Desulfovibrio fairfieldensis) at 1- and 2-month-old SOD1G93A mice, suggesting the early microbial contribution to the ALS pathological changes (Zhang et al., 2021).
A 2019 study (Blacher et al., 2019) confirmed the significant differences in the fecal microbiome composition of the SOD1G93A mice, compared with wild-type littermate controls. In the SOD1-Tg mice (generally known as SOD1G93A mice), antibiotic treatment was associated with a significant exacerbation of motor abnormalities in the mice throughout the progression of the disease. Motor neuron cell death increased after chronic exposure to antibiotics. In antibiotic treated SOD1G93A mice, the addition of P. distastonis and R. torques exacerbated disease progression. Treatment with L. gasseri and P. melaninogenica displayed disease promoting effects in a some but not all behavioral tests. None of the 11 bacterial strains that were tested affected motor abilities in the wild type mice. There was a slow reduction in the levels of A. muciniphila in SOD1G93A mice with progression of the disease, while its levels remained constant across time in the wild-type mice. A. muciniphila treatment significantly and substantially prolonged the lifespan of SOD1G93A mice, compared to vehicle-treated mice or SOD1G93A mice treated with other commensal microbiome species. A. muciniphila favorably modulated the course and severity of the disease in mice, whereas P. distastonis, R. torques, and possibly L. gasseri, P. melaninogenica adversely modulated the course and severity of ALS in mice. Overall, these results confirm the involvement of the gut microbiome in SOD1G93A mice.
Another study detected differences in the gut microbiome between SOD1G93A mice and controls at multiple time points (Figueroa-Romero et al., 2019). Interestingly, they noted that the ALS phenotype in the mice, including survival, varied depending on the animal facility used to house the mice; the authors suggested this curious observation was a result of differences in gut microbiota (Figueroa-Romero et al., 2019).
Intestine acts as a barrier to harmful molecules. A range of dietary toxins has been implicated in the cause of ALS, and dysbiosis could facilitate their entry because dysbiosis leads to increased intestinal permeability, as we observed in the previous report (Wu et al., 2015).
The ALS C9orf72 mouse model harbors loss of function mutations in the orthologous gene C9orf72 (Philips and Rothstein, 2015). A 2020 study (Burberry et al., 2020) found that the murine norovirus, Helicobacter spp., Pasteurella pneumotropica and Tritrichomonas muris were significantly more abundant in C9orf72Harvard mice than in C9orf72Broad mice. The increase in Helicobacter spp. levels suggested that gut microbiota changes may be the underlying mechanism of increased rate of mortality and inflammatory phenotypes in C9orf72Harvard mice. Treatment with antibiotics significantly decreased both the abundance and diversity of bacterial species without affecting the levels of murine norovirus, compared to vehicle treated controls. The bacterial species that decreased included Helicobacter spp. Administering lifetime antibiotics to the C9orf72Harvard -/- resulted in the complete suppression of inflammatory or autoimmune phenotypes, suggesting that when C9orf72 function is decreased, gut bacteria signals and promote autoimmunity and inflammation. Acute administration of broad-spectrum antibiotics reduced inflammatory and autoimmune phenotypes and improved rotarod performance in the mutant mice.
We summarized the existing studies on gut microbiome in ALS models in Table 3. Different ALS experimental models show altered microbiome, and the pathophysiological changes could by modulated by bacterial metabolites or antibiotic treatment. These results suggested that gut microbiota and environment modify immunity, neuroinflammation, and motor deficits in the development of ALS.
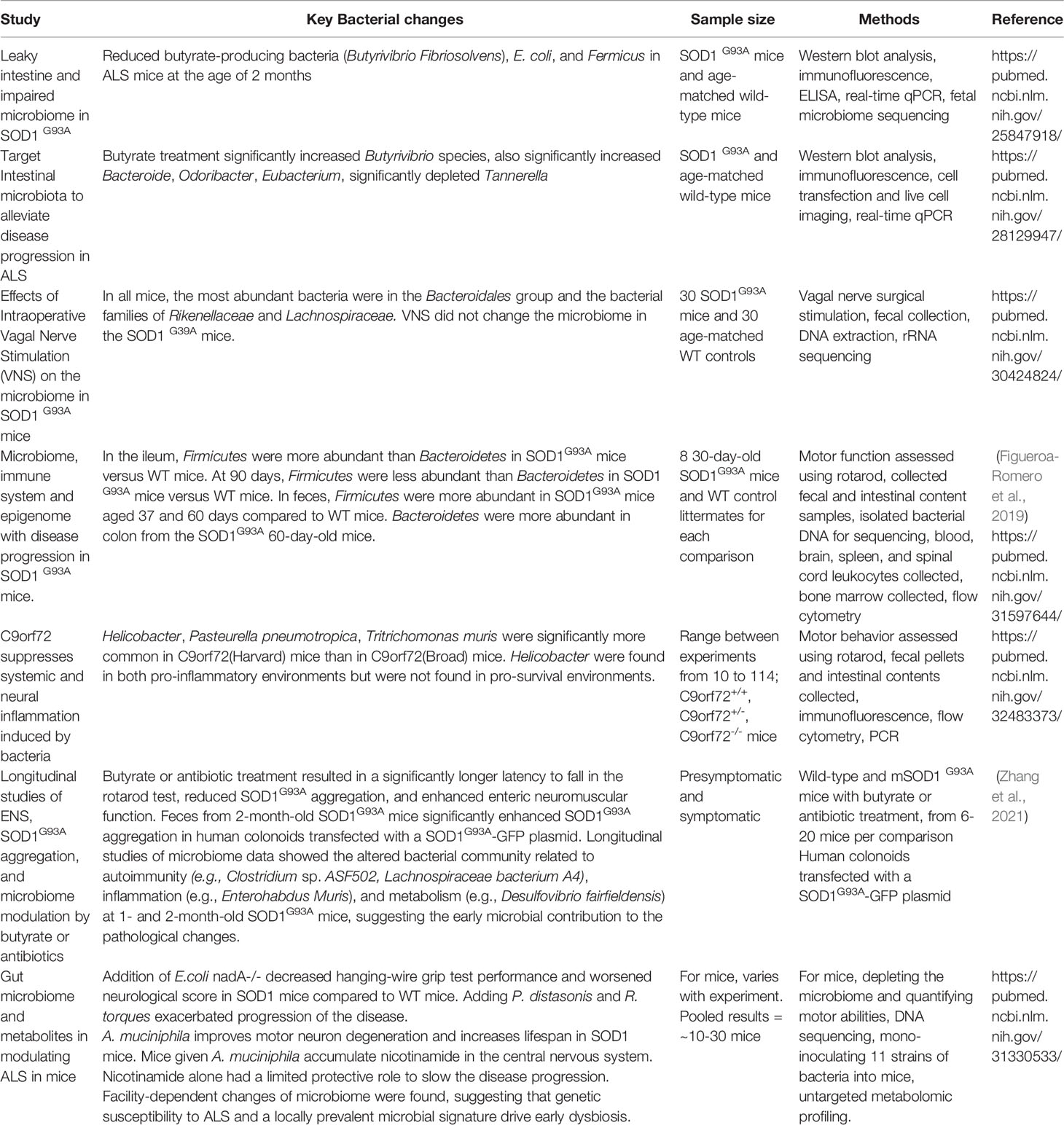
Table 3 Microbiome studies in ALS experimental models.
Human Studies of the Gut Microbiome in ALS
When examining 6 ALS patients and 5 healthy people without ALS, a 2016 study (Fang et al., 2016) found microbiomes of ALS showed a significant difference in the global bacterial gene content compared to healthy controls. Phylum Bacteroidetes, class Bacteroidia), order Bacteriodales, and genus Dorea were significantly higher in ALS patients than in the healthy controls. Lachnospiraceae (at family level), Firmicutes (at phylum level) and Clostridia (at class level), Oscillibacter (at genus level), Family XII (at family level), Anaerostipes (at genus level), Lachnospiraceae (at genus level), and Clostridiales (at order level) were significantly higher in the healthy controls than in the ALS patients. In the healthy controls, the 3 dominant phyla in 80% of the fecal microbiomes was Bacteroidetes, Firmicutes, and Actinobacteria. There was a significant decrease of Oscillibacter, Anaerostipes, and Lachnospiraceae in ALS patients.
The gastrointestinal complaints of the ALS patients predated neurological symptoms. In 2017 (Rowin et al., 2017), we studied the medical tests of 5 patients with ALS. The GI symptoms included gastroesophageal reflux, disease, chronic constipation, intermittent diarrhea, and abdominal pain and bloating. All ALS patients showed dysbiosis indicated by a decreased diversity of the microbiome compared to the healthy individuals. Patients 1, 3, and 5 had reduced benefit bacteria at the phylum level. Three patients (#1, 3 and 5) had a low Firmicutes/Bacteroidetes (F/B) ratio. Low Ruminococcus spp. occurred in all three ALS patients with a low F/B ratio, while Clostridium spp. and Roseburia spp. were low in patient 1. Patient 5 had high Bacteroides- Prevotella, Odoribacter spp., Barnesiella spp. and Bacteroides vulgatus. Patient 3 had high Bacteroides vulgatus, which could be the cause of a low F/B ratio. Patient 2 did not have a low F/B ratio, but the patient was taking probiotic supplements at the time of stool sample collection. All the ALS patients had a change to their gut microbiome that was characterized by a low diversity of the microbiome compared to the healthy individuals who had intact abundance within their gut microbiome. Our results suggested a potential role of intestinal inflammation and microbiome in the development and/or progression of human ALS.
In an observational study of 37 human ALS patients versus 29 healthy BMI- and aged- matched family members as controls (Blacher et al., 2019), the microbiome composition of ALS patients was significantly different from those of the healthy controls. There was only a small significant difference in the abundance of certain bacterial species in ALS patients. The microbiomes of the ALS patients showed a significant difference in the global bacterial gene content (Blacher et al., 2019). Additionally, there was a significant reduction in the abundance of several genes that map to the Akkermansia muciniphila genome.
A 2017 study (Brenner et al., 2018) analyzed the microbiome of 25 human patients with ALS and 32 healthy age- and gender- matched controls. They primarily found different proportions of uncultured Ruminococcaceae, However, aside from this finding and a difference in the overall number of microbial species, there was no significant differences in the quantity, diversity, and relative abundance of the fecal microbiota of the ALS patients. The range of different types of species of the intestinal microbiota did not differ between the ALS patients and the healthy controls. The F/B did not differ significantly between the ALS patients and healthy controls. The authors raised the possibility that while the gut microbiota of ALS patients experienced a change in metabolite production, ALS may not be associated with a significantly altered gut microbiota composition. This conclusion may be because the authors used a strict selection of ALS patient that had a high revised ALS functional rating scale without bulbar or respiratory symptoms, in order to avoid confounding factors. They analyzed a larger group of patients that were clinically well characterized. However, when ALS patients are at late stages of the disease, they may suffer from a variety of conditions that can affect the composition of the gut microbiota, influencing results. Revisiting this study, we actually know that F/B at the phylum level is a not realizable readout to indicate the functional changes of microbiome and the changes of microbiome should be investigated in-depth. The different proportion of Ruminococcaceae was an indicator of altered microbiome in these ALS patients. The authors only selected patients that were at a high functional level and did not have any overt bulbar or respiratory symptoms, therefore producing small and possibly inconsequential significant results. To generate more meaningful correlation, studies that involve all stages of the disease would need to be conducted. A secondary limitation is that altered metabolite production may be present in ALS patients, further disrupting the gut microbiota.
A 2018 study (Mazzini et al., 2018) analyzed the microbiota in 50 ALS patients and 50 healthy controls that were matched for sex, age, and origin. They found that there was a clear division between the bacterial profiles of ALS patients and the healthy controls. There was a lower DNA concentration in ALS patients compared to the controls. There was a low abundance of Clostridium and yeast, and there was a higher abundance of E. coli and Enterobacteria. The results of this study confirmed the ongoing hypothesis that the gut microbiota is altered in ALS patients and could potentially play a role in the pathogenesis of ALS.
In a 2020 study (Zeng et al., 2020) that examined the gut microbiota associated with ALS in 20 human patients and 20 age-matched healthy controls, there was a significant increase in the relative abundance of Bacteroidetes at the phylum level, Kineothrix, Parabacteroides, Odoribacter, Sporobacter, Eisenbergiella, Mannheimia, Anaerotruncus, and unclassified Porphyromonadaceae at the genus level and a significant reduction in Firmicutes at the phylum level and Megamonas at the genus level. Additionally, Bacteria at the domain level, Bacteroidetes at the phylum level, Bacteroidia at the class level, Bacteroidales at the order level, and Porphyromonadaceae at the family level were higher in ALS patients compared to controls. Microbes at the species level were significantly higher in ALS patients, including Sulfuricurvum kujiense, Cyanothece sp. and Haladaptatus paucihalophilus, while Enterococcus columbae was significantly decreased. These results indicated that the community diversity and species composition of the fecal microbiota is changed in patients with ALS. Limitations of this study include the fact that the authors could not determine whether the correlation between the intestinal dysbacteriosis and ALS occurred before or after the onset of ALS, so it still cannot be determined whether a causal relationship exists.
Another 2020 study (Di Gioia et al., 2020) examined 50 human patients with probable or defined sporadic and 50 age-matched case controls. The longitudinal study addressed the microbiota in patients with ALS and the impact of a probiotic supplementation on the gut microbiota and ALS progression. There was a significantly lower amount of Clostridium cluster I and yeasts and a higher concentration of E. coli in the ALS patients. A lower DNA concentration was present in the ALS patients, while the number of total bacteria was not significantly different between the ALS patients and the control group. Enterobacteriaceae were higher in ALS patients. The more representative phyla were Bacteroidetes and Firmicutes in both groups, which showed an abundance of 40-45%. Cyanobacteria phylum members were significantly higher in ALS patients versus control patients. At the genus level, Lactobacillus, Citrobacter, Coprococcus, and Ruminococcaceae were more abundant in the ALS patient group. Veillonellaceae and Lachnospiraceae families, Parasutterella, Ruminococcus and Subdogranulum were reduced in ALS group, compared to the controls. Overall, these results suggested the existence of differences in the bacterial composition between healthy controls and patients with ALS. This study demonstrated the imbalance of intestinal bacteria that may play important roles in modulating the central nervous system (CNS). Limitations of this study include the restricted group size - 20 ALS patients were not able to complete the study and 1 died. Additionally, intra-individual variability was not incorporated into the study.
In a 2021 study (Niccolai et al., 2021), they looked at the gut microbiota and its SCFA composition and the intestinal and systemic inflammatory response in ALS patients compared to their healthy controls. In 19 ALS patients and 9 healthy family caregivers that were matched for sex and were close in age, there were significant differences in cytokine levels. When looking at the cytokine levels in the serum, they found that IL-8, IL-15, MCP-1 and VEGF-A levels were significantly lower in ALS patients compared to controls. When looking at the cytokine levels in the fecal samples, they found that IL-2 was higher in ALS patients compared to controls, although the difference was not significant. Additionally, IL-21 was lower in ALS patients that had a faster progressing form of the disease, compared to patients that has medium or slow progressions. Patients with a slower progression of ALS had lower microbial diversity than other variations of progression. Similar results were displayed for ALS patients with the classical form of the disease compared to patients with various clinical phenotypes. Control samples displayed a higher Firmicutes/Bacteroides ratio than the ALS patients with the Firmicutes and Bacteroides being the most represented phyla in all samples. ALS patients that had a medium progression form of the disease displayed a higher Firmicutes/Bacteroides ratio than both ALS patients with other forms of progression and healthy controls. However, these differences did not appear to be statistically significant. At the genus level, the healthy controls displayed an enrichment of Erysipelotrichaceae_UCG-003, Fusicatenibacter, and Subdoligranulum compared to the ALS patients. There appeared to be an increase of Streptococcus in the samples from ALS patients with a slow progression of the disease. From this study, the difference in serum profile between the ALS patients and healthy controls suggests that there may be an intestinal inflammatory aspect to ALS. Since IL-21 was expression more in patients with a slow form of ALS compared to a fast progression, it could potentially be developed into a prognostic biomarker. The study confirmed that there is dysregulation occurring in both a systemic and intestinal manner in ALS patients. Limitations of this study include small sample sizes and the presence of various confounding factors that can include diet and secondary disease effects.
A 2021 study (Niccolai et al., 2021) found the differences in 15 bacterial species between the ALS patients and healthy controls. This included butyrate-prodiucing bacteria Roseburia intestinalis and Eubacterium rectale. The total relative abundance of several butyrate producers was significantly lower in ALS patients compared to healthy controls. Bilophila (unclassified), Clostridiaceae bacterium JC1118, Coprobacter fastidiosus, Eubacterium eligens, and Ruminococcus sp 5 1 39 BFAA was lower in ALS patients compared to controls, while the relative abundance of Escherichia (unclassified) and Streptocossus salivarius was higher in ALS patients. To avoid the potential changes that occur in late term ALS, they compared the samples from PALS that were collected within one year after diagnosis, the relative abundance of both Roseburia intestinalis and Eubacterium rectale were significantly lower in the ALS patients compared to healthy controls. The levels of total butyrate-producing bacteria were lower in the PALS patients compared to healthy controls, but the difference was not statistically significant. In patients that were not using Riluzole at the time of sample collection, they found that the results were similar – Roseburia intestinalis, Eubacterium rectale, and total abundance of butyrate-producing bacteria were lower in these ALS patients compared to healthy controls. The results were the same when excluding ALS patients that were using a gastrostomy tube. Overall, they found reduced levels of anti-inflammatory SCFA-producing bacteria in patients with ALS. A limitation of this study could be that patients unintentionally changed their diet after developing ALS, therefore altering the gut microbiota composition. Dietary differences were unfortunately not able to be accounted for. This study further indicates that gut microbiome could act as an integral component in the pathophysiology of ALS.
A 2022 study (Zhang et al., 2022) investigated the connection between the gut microbiota and ALS in a GWAS of ALS that involved 20,806 patients and 59,804 control individuals. They found that OTU10032 unclassified Enterobacteriaceae (2 independent SNPs) and unclassified Acidaminococcaceae (4 uncorrelated SNPs) were associated with a higher risk of ALS. Gamma-glutamyl amino acids demonstrated possible negative effects of the risk of developing ALS. Gamma-glutamylphenylalanine specifically displayed an increased risk of developing ALS that was significant. 1-arachidonoyl-GPI and 3-methyl-2-oxobutyrate, two metabolites, were connected with an increased risk of developing ALS. An increase in the levels of 4-acetamidobutanoate may lower the risk of developing ALS. Additionally, an increase in the relative abundance of OTU4607_Sutterella was associated with genetically predicted ALS. An increase in the relative abundance of Lactobacillalesorder was associated with genetically predicted ALS. This study showed that a bidirectional relationship between the gut microbiota and ALS. Limitations of this study include weak instrument bias and population stratification, though this bias was somewhat reduced by restricting the dataset to individuals of European ancestry.
A 2022 study (Hertzberg et al., 2022) analyzed the gut microbiota of ALS patients while using the patients’ spouses as the healthy controls. The authors found that the ALS patients did not have any butanoate metabolic pathway enzymes. They also displayed lower amounts of enzymes that are utilized in various metabolic pathways. These include pathways involving response regulators and carbon metabolism pathways. When comparing the 25 stool and inflammatory markers, including IL-2, IL-4, tumor necrosis factor, and interferon, there were no statistically significant differences between the ALS patients and the healthy controls. Additionally, ALS patients did not have amplicon sequence variants of the genus Prevotella, specifically P. timonensis, compared to the healthy controls. This specific result was consistent with other studies that suggest that gut microbiome dysbiosis is associated with neurodegenerative diseases. The authors suggested that the difference in P. timonensis in ALS patients versus the controls may have contributed to the lack of differences in pro-inflammatory cytokine levels. In contrast to previous studies, they did not find a lack of butyrate-producing microbes. However, they found a decrease in the enzymes associated with butyrate metabolism, which is consistent with previous studies, suggesting an association between gut butyrate availability and progression of ALS. Further studies looking at this association would be needed. Limitations of this study include the small sample size and the lack of diet information. To overcome the small sample size, larger scaled studies would need to be conducted.
In a 2019 study (Zhai et al., 2019) looking at 8 patients with ALS and 8 healthy controls, they found that the richness and evenness of bacterial and archaeal communities of healthy individuals were healthier than those of the ALS patients. They also found that the average relative abundance of Firmicutes in ALS patients was 4.7% higher than the abundance in healthy controls. On the class level, the relative abundance of Negativicutes and Bacili were decreased compared to healthy controls. The relative abundance of phylum Euryarchaeota, class Methanobacteria, and genus Methanobrevibacter were all significantly increased in ALS patients compared to the healthy controls. While there were no significant differences in metabolites between ALS patients and healthy controls, there was an increase in SCFA, NO2-N/NO3N, and γ-aminobutyric acid in ALS patients compared to healthy controls. The results of this study suggested that the biodiversity and composition of intestinal microflora in ALS patients is of lower quality than that of controls. The increase in the Firmicutes/Bacteroidetes ratio and Methanobrevibacter indicates that there is a gut flora composition imbalance in patients with ALS. One limitation of this study was that metagenomic sequencing was not performed in order to analyze microbial function variation. Another limitation would be the need for microbial community acclimation in order to better understand the composition of the microbiota in ALS patients.
In a 2020 study (Ngo et al., 2020) that looked at the relationship of the fecal microbiota and prognosis of ALS, they found that ALS patients with a higher richness and evenness of their fecal microbiome had a decreased survival from their onset of symptoms than the patients who had less richness and evenness. There was no significant difference in Proteobacteria between ALS patients and controls. There were also no significant differences in the proportional abundance in the two most abundant phyla, Firmicutes and Bacteriodetes, between ALS patients and controls. Overall, they found the fecal microbiota in a group of ALS patients is not significantly different than those of healthy controls. They believe that clinical and metabolic features of ALS are not connected with the fecal microbiome composition. However, the diversity and richness of the fecal microbiome may be associated with an increased risk of an earlier death. The results suggest that the fecal microbiota of ALS patients does not differ from those of healthy individuals. One limitation of this study is that they included both patients who were newly diagnosed and patients who had already progressed and were attending clinics. Other limitations would be a small sample size and the amplifying of variability due to the patients’ dietary habits.
A potential proxy for the effect of the gut microbiome on disease is the effect of oral antibiotics which significantly modify the balance of gut microbial species. A 2019 study demonstrated that any use of antibiotics, especially repeated use, was associated with increased risk of ALS in the native Swedish population (2006-2013 period) (Sun et al., 2019). Similarly, SOD1G93A mice repeatedly exposed to antibiotics develop a more severe motor phenotype and increased motor neuron loss (Blacher et al., 2019),
We summarized the studies on gut microbiome in ALS patients in Table 4. These results suggested novel roles of intestinal microbiome in the development of ALS and the potential benefits of decreasing the number of pathogens and increasing probiotics. The challenge of early studies of microbiome in ALS lies in the following aspects: 1) limited sample size; 2) preliminary data analysis; and 3) reluctance from neurologists to pay attention the intestinal changes in the early stage of ALS. In the earlier time of microbiome research, a F/B ratio was used. Although it indicated the evidence of dysbiosis in patients with ALS. The F/B ratio only reflect the relative abundance of bacteria at the phylum level and may miss the changes at the species, thus, it is not a reliable readout for the in-depth analysis and advanced studies of microbiome data.
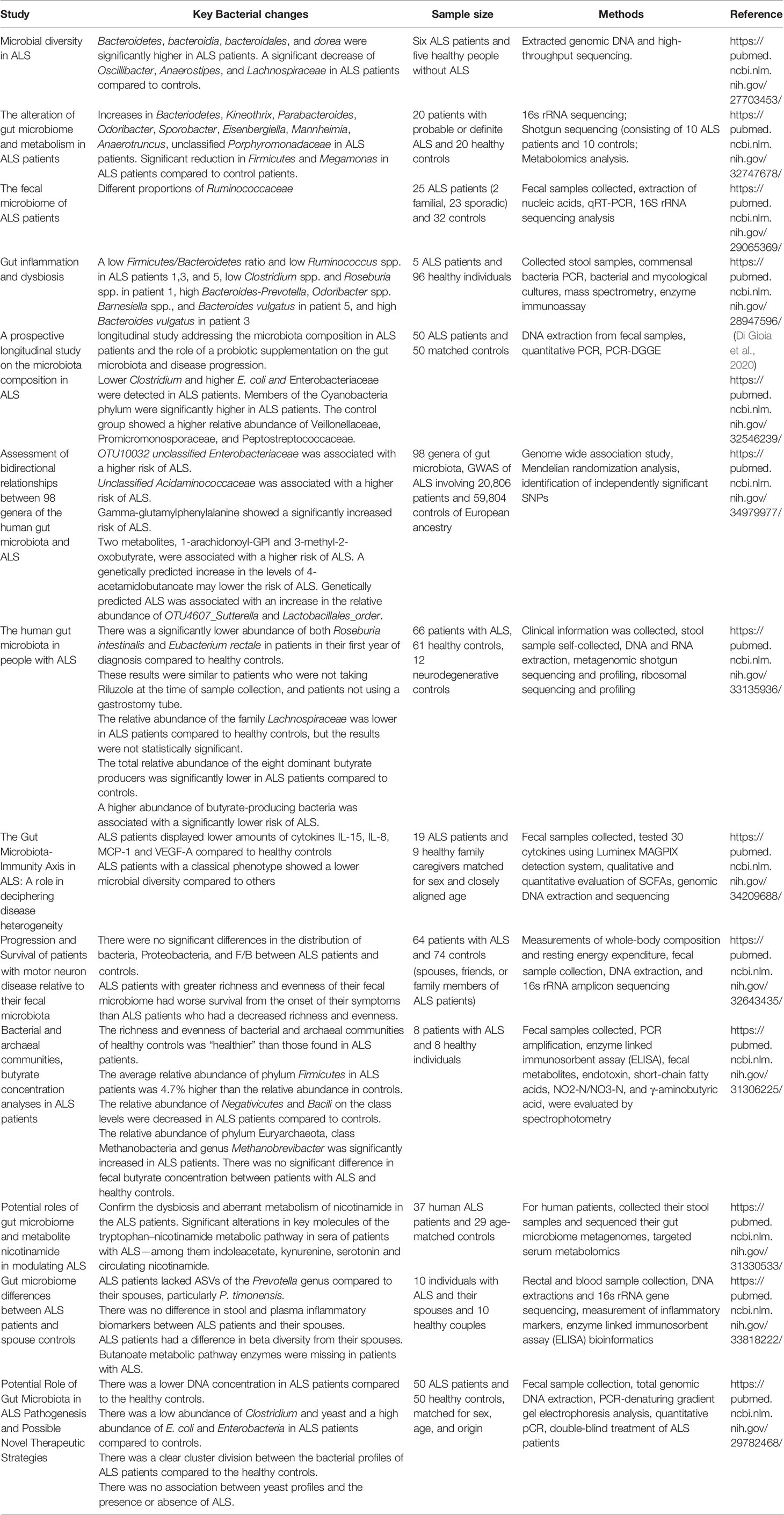
Table 4 Microbiome studies in patients with ALS.
Intestinal Inflammation in ALS
There is abundant evidence many substances involved in the promotion of inflammatory processes are present in the CNS of patients. Previous reviews have discussed the role of inflammation in ALS and the possibility of treating ALS by immune modulation (McCombe and Henderson, 2011; Kwon and Koh, 2020). Dysregulation of the peripheral immune system and reduced levels of regulatory T cells are associated with disease progression. Monocytes are activated in ALS and dendritic cells play a role in pathogenesis. In the brain and spinal cord of patients with ALS, microglia respond to injury in a beneficial way, but chronic microglial activation is thought to contribute to the pathogenesis of the neurodegenerative disease. There is activation of microglia (‘neuroinflammation’) in ALS. Microglia can have opposite roles at different times. However, the local inflammation in other organs, e.g GI, was less explored in the ALS research.
Intestinal Paneth cells in the small intestine produce most of the antimicrobial peptides (AMPs) and play an important role in regulating the innate immunity and inflammation (Deretic et al., 2008; Lu et al., 2021). The Paneth cell is a unique that can sense commensals and secrete AMPs, thereby playing critical roles in the maintenance of homeostasis at the intestinal-microbial interface. There was a decreased number of normal Paneth cells in the small intestine of SOD1G93A mice, whereas abnormal Paneth cells were significantly increased in SOD1G93A mice. There was a decreased amount of the AMP defensin 5 alpha in the intestine of the 3-month-old SOD1G93A mice with disease symptoms. There was a significant reduction in lysozyme 1 in the G93A intestine, but lysozyme 2 was not altered. There were no pathological changes in the small intestine of G93A mice. There were increased serum IL-17 levels in the young G93A mice at 2 months of age. IL 17 was increased in the intestine of G93A mice, suggesting a preinflammatory state in ALS mice before disease onset. There were higher FITC readings that indicated higher permeability of the intestine. Our study has demonstrated Paneth cell dysfunction and inflammation in the intestine of G93A mice. The intestinal inflammation may occur earlier than the systemic inflammation.
A recent study (Figueroa-Romero et al., 2019) in the SOD1G93A model showed that dysbiosis was observed in the 37 day-old mice. Moderate differences in the peripheral system beginning at 90 days, with elevated CD8T cells in the spleen, and then at 120 days with CD4T cell counts in peripheral blood. The CNS inflammation occurs during late-stage disease in the SOD1G93A mice. This study provides a roadmap to the chronological changes that occur in the intestinal microbiome and immune system relative to disease onset and progression in the SOD1G93A mouse model. It further supports the critical role of microbiome in the early stage of ALS pathogenesis (Zhang et al., 2021). Limitations of this study include that there may have been some variation introduced by a separate cohort of 120-day-old SOD1 G93A mice. Additionally, the number of males versus females was not always equal during the course of the study.
A previous study (Zhang et al., 2009) reported that the level of plasma LPS showed a significant increase, correlating with blood monocyte/macrophage activation in sALS groups, especially those with advanced sALS disease (Zhang et al., 2009). A report (Tortelli et al., 2020) studied a panel of five cytokines (IL-2, IL-6, IL-10, IFN-gamma, and TNF-alpha) in plasma in ALS (79 patients with ALS and 79 age- and sex-matched healthy controls). All the five cytokines were significantly increased in plasma samples of patients compared with controls, with IL-6 having the highest median concentration (10.11 pg/ml) in the ALS group. Furthermore, IL-6 was the plasma cytokine with the highest discrimination ability between patients and controls according to the receiver operating characteristic analysis. The elevated inflammatory cytokines in ALS support a possible role of inflammation in disease progression. Recent animal experimental studies (Butovsky et al., 2012) have tried to modulate of inflammatory monocytes with microRNAs in ALS mice.
In the animal model of C9orf72 study (Burberry et al., 2020), transplantation of pro-survival intestinal microbiome in the C9orf72Harvard resulted in a significant decrease of inflammatory and autoimmune phenotypes, compared with transplantation of pro-inflammatory microbiome. These results suggested that signals from gut microbiota can help maintain the suppression of certain inflammatory and autoimmune phenotypes. Helicobacter spp. was found in both pro-inflammatory environments, but not in pro-survival environments. The abundance of 62 of 301 bacterial species were significantly changed when comparing the two pro-survival environments to the two-pro-inflammatory environments.
Gut microbes can influence the innate and adaptive immune systems, including immune cells in the gut and the systemic immunity.
Neuroinflammation contributes to the pathogenesis of ALS through various mechanisms including enhanced reactive oxygen species (ROS), activation of innate immune systems, increased proinflammatory cytokines, and infiltration of immune cells. Dysbiosis leads to an inflammatory state and altered gut microbiota could influence the immune system in ALS. Moreover, the nutrition has a direct impact on gut microbiome, which shapes local intestinal immune responses and in turn affects even autoimmune responses. Conjugated linoleic acid (CLA), a naturally occurring fatty acid in meat and dairy products of ruminants. A recent study (Fleck et al., 2021) identified CLA as a potent modulator of the gut–CNS axis by targeting myeloid cells in the intestine, which in turn controlled encephalitogenic T-cell responses in mouse model of multiple sclerosis and a pilot human study. However, microbiome, nutrients, immune cells, and cytokines in early disease pathology of human ALS are still unclear. The precise role of inflammation in various organs, include GI, in ALS needs to be further investigated on larger samples and with more mechanistic studies.
Targeting Microbiome and Metabolites in ALS Treatment
Significant alterations in the levels of various metabolites were identified in the sera of ALS patients. Bacterial product butyrate is beneficial and slows down the progression of ALS, based on our studies in the SOD1G93A mice (Zhang et al., 2017; Zhang et al., 2021). There is a trial (Paganoni et al., 2020; Turnbull, 2020) using sodium phenylbutyrate-taurursodiol in ALS patients. It reported Sodium phenylbutyrate-taurursodiol resulted in slower functional decline than placebo as measured by the ALSFRS-R score over a period of 24 weeks.
There were alterations in the levels of key components of the tryptophan-nicotinamide (NAM) pathway in some ALS patients. Specifically, the levels of the metabolite NAM in the CSF of 14 patients with ALS were significantly lower than the levels in 17 healthy controls. These results suggested that ALS does interact in some manner with the intestinal microbiota of patients and creates a cycle in which the increase in pathogens and decrease in probiotic organisms in the intestines of ALS patients can upregulate or downregulate the production of NO, GABA, and SCFAs, increasing the pathogenesis of ALS which furthers the imbalance of the intestinal microbiota. The gut microbiota produces metabolites of tryptophan that play a role in regulating astrocyte and microglial activation. Combined, these findings suggest a possible role for microbial metabolites in the control of neuroinflammation during ALS disease progression. Targeting the gut-brain axis could modify the neuronal function (Huang et al., 2019; Kang et al., 2019) and possibly contribute to some of the non-motor cognitive and behavioral symptoms in ALS, but this would require further study.
Riluzole is FDA proved disease-modifying treatment for ALS and has the potential to extend life by only a few months (Miller et al., 2012). Interestingly, Riluzole was significantly metabolized by 40 of the bacteria, based on a 2019 study of on drug metabolism by microbiome (Zimmermann et al., 2019). Many of these bacteria are known to vary in prevalence in the human population. However, the role of the microbiome has not been linked to the drug metabolism and management of ALS treatment.
The use of proton pump inhibitor (PPI) drugs has been associated with a decrease in gastric pH, leading to gastrointestinal dysbiosis (Minalyan et al., 2017). It has also been suggested that PPI use may be related to an increased risk of neurodegenerative diseases (Erber et al., 2020). A retrospective cohort study conducted in Austria reported that 97.5% of ALS patients use other drugs together with riluzole, including PPI and centrally acting muscle relaxants (CAMR). A significant trend in PPI in reducing the survival of ALS was noted; however, after statistical corrections were applied, this effect was no longer observed. On the other hand, the use of CAMR in association with riluzole showed a beneficial effect on the survival of these patients (Cetin et al., 2015). In a case-control study conducted with 2,484 ALS patients in Sweden, no correlation between PPI use and risk of ALS was observed in a lag window of 1-3 years (Cetin et al., 2020). Thus, it is still lacking evidence to support the detrimental effect of PPI in ALS; nevertheless, 50% of ALS patients report using PPI, raising the importance of further investigations to better understand this correlation.
The increase in the abundance of Rikenellaceae was significant in ALS patients who underwent a 6-month probiotic treatment administered daily (Di Gioia et al., 2020). The probiotic formulation is a mixture of five lactic acid bacteria: Streptococcus thermophilus ST10–DSM 25246, Lactobacillus fermentum LF10–DSM 19187, and Lactobacillus delbrueckii subsp. delbrueckii LDD01–DSM 22106, Lactobacillus plantarum LP01–LMG P-21021, and Lactobacillus salivarius LS03–DSM 22776. No adverse events attributed to probiotic supplementation. The significant increase in Cyanobacteria at the phylum, family, and genus level in ALS patients compared to controls, suggesting that cyanobacteria play a critical role in the pathogenesis of neurodegenerative diseases. The abundance of cyanobacteria decreased overtime in both the probiotic and placebo groups, although the difference was not significant.
Fecal microbiota transplantation (FMT) could improve gastrointestinal and behavioral symptoms in neurological diseases (Kang et al., 2017; Kang et al., 2019). Parkinson’s disease patients after one-week treatment of FMT improved constipation and motor symptoms such as leg tremors. The tremors recurred 2 months after FMT, whereas constipation was relieved even after 3 months (Huang et al., 2019). Intestinal bacteria as an external trigger could explain the rare cases of ALS in spouses or in some clusters (Sabel et al., 2003). To reconstruct the intestinal microbiome, FMT has been used to transfer the gut microbiota from healthy individuals to ALS patients (Xu et al., 2021). ClinicalTrials.gov (https://clinicaltrials.gov/ct2/show/NCT03766321) has an ongoing FMT trial for a year with 42 ALS patients, at an early stage (28 FMT-treated patients vs. 14 controls). This will promote a further understanding of microbiota restoration and GI function.
Diet, Microbiome, and ALS
Many factors are involved in the gut microbiota composition and function, including genetics, gender, age, geographic location, lifestyle, and diet (Ballan et al., 2020; de Vos et al., 2022). Also, the gut microbiota is responsible for metabolization of compounds from the diet producing several metabolites (Ballan et al., 2020; de Vos et al., 2022). As a consequence, different gut microbiota compositions will result in different metabolite profiles, that will interact with cell receptors, having a direct impact on different signaling pathways and on the host health (de Vos et al., 2022).
Diet is associated with the ALS risk, onset, and progression, providing potential modifiers of disease (Pape and Grose, 2020). Two mutant SOD1(G86R and G93A) mice exhibited an altered metabolic status (Dupuis et al., 2004). This study provided evidence that transgenic ALS mice suffer from a dramatic defect in energy homeostasis, likely linked to a hypermetabolism mainly of muscular origin. High-energy-diet improved the altered metabolic phenotype of G86R mice and delays disease onset (Dupuis et al., 2004). Several epidemiological studies have identified diets that positively affect ALS patients, including various high-calorie fat or sugar-based diet, summarized in a recent review (Pape and Grose, 2020).
Gluten-induced autoimmunity was speculated to trigger ALS. However, the data supporting this link are weak (Group, 2016). Leptin is an adipokine involved in food intake regulation and energy balance. Lower level of circulating leptin is associated with fat mass loss, and thus, body weight loss in ALS. Moreover, it has been suggested that fat mass loss occurs prior to muscle atrophy. Thus, leptin could be a potential biomarker of adipose tissue wasting in the early stages of ALS. The appetite-stimulating hormone ghrelin also seems to be downregulated in an ALS mice model, compared with their WT controls, contributing to lower food intake and body weight loss (Ferrer-Donato et al., 2021).
Earlier studies suggested that polyphenols (e.g., resveratrol, curcumin, epigallocatechin gallate, quercetin, and phenolic acids), which can be found in fruits, vegetables, coffee, tea, and whole grains, may have a promising neuroprotective effect in ALS. It was observed, in vivo and in vitro, that these bioactive compounds may have the potential to regulate mitochondrial biogenesis, improve energy metabolism, reduce toxic protein aggregation, reduce microglia and astrocytes inflammation, and improve motor functions and survival (Solanki et al., 2015; Novak et al., 2021).
Resveratrol, an antioxidant compound found in grapes, has been widely studied due to its neuroprotective properties. Resveratrol may reduce the in vitro neurotoxicity of cerebrospinal fluid (CSF) from ALS patients, preventing neuronal loss and improving Ca2+ homeostasis, which seems to be related to the antioxidant capacity of resveratrol. Curiously, co-incubation with riluzole inhibited this protective effect (Yanez et al., 2011). In fact, Ca2+ dyshomeostasis is related to impaired autophagy mechanisms and toxic protein aggregation in neurodegenerative disorders, including ALS (Tedeschi et al., 2019). Therapeutic interventions aiming to modulate autophagy pathways seemed to be an interesting approach to reduce protein aggregates, mainly in the early stages of ALS (Cipolat Mis et al., 2016).
A prospective study reported that probiotics may promote beneficial effects in ALS patients. The administration of probiotics to ALS patients increased the relative abundance of groups related to propionate and butyrate production in gut microbiota, which may improve energy provision. Also, it has been speculated a possible metabolization of neurotoxic compounds (Di Gioia et al., 2020).
In our previous work, we observed the administration of butyrate, a beneficial microbial metabolite, delayed the disease progress and significantly extended the survival time of the SOD1 G93A mice and prolonged the life span by 38 days on average (Zhang et al., 2017). Butyrate treatment improved the gut microbiome and restored the Paneth cells and the signaling of lysozyme 1 and anti-microbial peptide defensin 5 alpha in the SOD1 G93A mice. Meanwhile, the intestinal protein aggregation of SOD1G93A was significantly decreased by butyrate treatment (Zhang et al., 2017). Furthermore, butyrate treatment was able to enhance healthy metabolites by longitudinal untargeted metabolomic analysis in ALS (Destiny Ogbu and Claud, 2022).
Nicotinamide showed slightly delay of the disease progression in the ALS SOD1G93A mice (Blacher et al., 2019). The survival curve showed only several days difference between the SOD1G93A mice with or without nicotinamide treatment. Aberrant metabolism of nicotinamide was observed in the sera of patients with ALS. There was a trial of nicotinamide/pterostilbene supplement in ALS (https://clinicaltrials.gov/ct2/show/NCT04562831). The results have not been reported yet.
Emerging evidence support the roles of gut-neuron-microbiome in various human diseases (Ogbu et al., 2020; Fang et al., 2020). For examples, in a human pilot study in patients with multiple sclerosis, dietary conjugated linoleic acid-supplementation for 6 months significantly enhanced the anti-inflammatory profiles and functional signatures of circulating myeloid cells (Fleck et al., 2021). To move forward, we hypothesize that dietary interventions with functional foods (e.g., prebiotics, probiotics, polyphenols) or the administration of probiotic metabolites (e.g., butyrate) aiming the gut microbiota modulation may enhance intestinal barrier functions and autophagy regulation, restore healthy host-microbial interactions, and reduce toxic protein aggregation and inflammation, thus, delaying the progression of ALS.
Conclusion and Future Directions
ALS is a progressive neurodegenerative disorder involving brain and spinal cord motor neuron death resulting in weakness and wasting of musculature and leaning inexorably to death. There are GI abnormalities involved liver, intestine, and pancreas in ALS. Subclinical gastrointestinal motor dysfunction, delayed colonic transit time, delayed gastric emptying an increased prevalence of constipation and anal sphincter abnormalities have been noted, but the mechanism of these issues is unclear. In various mouse models of ALS, the altered microbiome and GI issues are also observed. However, factors such as changes of diet, GI issues, inflammation, and infection were not well considered for the ALS diagnosis.
Emerging evidence has demonstrated the novel roles of microbiome and metabolites in the early stage of disease development and progression, suggesting the potential biomarkers (Chatterjee et al., 2020; Donatti et al., 2020). SALS and fALS forms of ALS manifest similar pathological and clinical phenotypes, suggesting that different initiating causes lead to a mechanistically similar neurodegenerative pathway. ALS patients have abnormal GI systems, elevated intestinal inflammation and dysbiosis. The ENS and smooth muscle automatism are unable to modulate the motor functions of the digestive tract, which provide an anatomical explanation for these clinical manifestations. Studies on microbiome changes with pro-inflammatory serum cytokines and LPS in patients will help with the early diagnosis of the disease. To treat gut microbiota dysbiosis through microbiota restoration would have the potential to interfere and slow ALS progression (Mandrioli et al., 2019). The therapeutic methods to target microbiome and intestinal functions, e.g, FMT, prebiotics, and probiotics, will help both SALS and FALS patients. ALS studies should be performed by considering diet, microbiome, lifestyle, and gender difference. Studies that examine the microbiome together with intestinal pathogenesis will help to determine when, where, and whether microbiome and metabolites are critical to disease progression of ALS. Understanding the pathogenesis of ALS GI will provide innovative strategies for accurate diagnosis and better treatment for this challenging disease.
Author Contributions
SM and JS performed literature search and detailed analyses and summary of related literature, prepared the draft text. CB contributed to literature search of ALS mouse models, nutrition, and the draft text. JS designed the study/project, obtained funds, summarize literature, and directed the project. All authors contributed to the article and approved the submitted version.
Funding
We would like to acknowledge the VA Merit Award 1 I01BX004824-01, the NIDDK/National Institutes of Health grant R01 DK105118, and R01DK114126 to JS. The study sponsors play no role in the study design, data collection, analysis, and interpretation of data. The contents do not represent the views of the United States Department of Veterans Affairs or the United States Government.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ballan, R., Battistini, C., Xavier-Santos, D., Saad, S. M. I. (2020). Interactions of Probiotics and Prebiotics With the Gut Microbiota. Prog. Mol. Biol. Transl. Sci. 171, 265–300. doi: 10.1016/bs.pmbts.2020.03.008
Banack, S. A., Cox, P. A. (2003). Biomagnification of Cycad Neurotoxins in Flying Foxes: Implications for ALS-PDC in Guam. Neurology 61, 387–389. doi: 10.1212/01.WNL.0000078320.18564.9F
Baquero, F., Nombela, C. (2012). The Microbiome as a Human Organ. Clin. Microbiol. Infect. 18, 2–4. doi: 10.1111/j.1469-0691.2012.03916.x
Blacher, E., Bashiardes, S., Shapiro, H., Rothschild, D., Mor, U., Dori-Bachash, M., et al. (2019). Potential Roles of Gut Microbiome and Metabolites in Modulating ALS in Mice. Nature 572, 474–480. doi: 10.1038/s41586-019-1443-5
Brenner, D., Hiergeist, A., Adis, C., Mayer, B., Gessner, A., Ludolph, A. C., et al. (2018). The Fecal Microbiome of ALS Patients. Neurobiol. Aging 61, 132–137. doi: 10.1016/j.neurobiolaging.2017.09.023
Brown, K. J., Jewells, V., Herfarth, H., Castillo, M. (2010). White Matter Lesions Suggestive of Amyotrophic Lateral Sclerosis Attributed to Celiac Disease. AJNR Am. J. Neuroradiol. 31, 880–881. doi: 10.3174/ajnr.A1826
Burberry, A., Wells, M. F., Limone, F., Couto, A., Smith, K. S., Keaney, J., et al. (2020). C9orf72 Suppresses Systemic and Neural Inflammation Induced by Gut Bacteria. Nature 582, 89–94. doi: 10.1038/s41586-020-2288-7
Butovsky, O., Siddiqui, S., Gabriely, G., Lanser, A. J., Dake, B., Murugaiyan, G., et al. (2012). Modulating Inflammatory Monocytes With a Unique microRNA Gene Signature Ameliorates Murine ALS. J. Clin. Invest. 122, 3063–3087. doi: 10.1172/JCI62636
Cetin, H., Klickovic, U., Rath, J., Zulehner, G., Fuzi, J., Reichardt, B., et al. (2015). Associations Between Co-Medications and Survival in ALS-A Cohort Study From Austria. J. Neurol. 262, 1698–1705. doi: 10.1007/s00415-015-7767-7
Cetin, H., Sun, J., Almqvist, C., Reichardt, B., Tomschik, M., Zimprich, F., et al. (2020). No Association Between Proton Pump Inhibitor Use and ALS Risk: A Nationwide Nested Case-Control Study. Sci. Rep. 10, 13371. doi: 10.1038/s41598-020-70373-8
Charchaflie, R. J., Bustos Fernandez, L., Perec, C. J., Gonzalez, E., Marzi, A. (1974). Functional Studies of the Parotid and Pancreas Glands in Amyotrophic Lateral Sclerosis. J. Neurol. Neurosurg. Psychiatry 37, 863–867. doi: 10.1136/jnnp.37.7.863
Chatterjee, I., Lu, R., Zhang, Y., Zhang, J., Dai, Y., Xia, Y., et al. (2020). Vitamin D Receptor Promotes Healthy Microbial Metabolites and Microbiome. Sci. Rep. 10, 7340. doi: 10.1038/s41598-020-64226-7
Chio, A., Logroscino, G., Traynor, B. J., Collins, J., Simeone, J. C., Goldstein, L. A., et al. (2013). Global Epidemiology of Amyotrophic Lateral Sclerosis: A Systematic Review of the Published Literature. Neuroepidemiology 41, 118–130. doi: 10.1159/000351153
Cipolat Mis, M. S., Brajkovic, S., Frattini, E., Di Fonzo, A., Corti, S. (2016). Autophagy in Motor Neuron Disease: Key Pathogenetic Mechanisms and Therapeutic Targets. Mol. Cell Neurosci. 72, 84–90. doi: 10.1016/j.mcn.2016.01.012
Deretic, V., Master, S., Singh, S. (2008). Autophagy Gives a Nod and a Wink to the Inflammasome and Paneth Cells in Crohn’s Disease. Dev. Cell 15, 641–642. doi: 10.1016/j.devcel.2008.10.009
Destiny Ogbu, Y. Z., Claud, K. (2022). Yinglin Xia, Un Sun, Dietary Butyrate Treatment Enhances Healthy Metabolites by Longitudinal Untargeted Metabolomic Analysis in Amyotrophic Lateral Sclerosis Mice. Biorxiv. doi: 10.1101/2022.01.15.476456
de Vos, W. M., Tilg, H., Van Hul, M., Cani, P. D. (2022). Gut Microbiome and Health: Mechanistic Insights. Gut. doi: 10.1136gutjnl-2021-326789
Di Gioia, D., Bozzi Cionci, N., Baffoni, L., Amoruso, A., Pane, M., Mogna, L., et al. (2020). A Prospective Longitudinal Study on the Microbiota Composition in Amyotrophic Lateral Sclerosis. BMC Med. 18, 153. doi: 10.1186/s12916-020-01607-9
Donatti, A., Canto, A. M., Godoi, A. B., da Rosa, D. C., Lopes-Cendes, I. (2020). Circulating Metabolites as Potential Biomarkers for Neurological Disorders-Metabolites in Neurological Disorders. Metabolites 10 (10), 389. doi: 10.3390/metabo10100389
Dupuis, L., Oudart, H., Rene, F., Gonzalez de Aguilar, J. L., Loeffler, J. P. (2004). Evidence for Defective Energy Homeostasis in Amyotrophic Lateral Sclerosis: Benefit of a High-Energy Diet in a Transgenic Mouse Model. Proc. Natl. Acad. Sci. U.S.A. 101, 11159–11164. doi: 10.1073/pnas.0402026101
Erber, A. C., Cetin, H., Berry, D., Schernhammer, E. S. (2020). The Role of Gut Microbiota, Butyrate and Proton Pump Inhibitors in Amyotrophic Lateral Sclerosis: A Systematic Review. Int. J. Neurosci. 130, 727–735. doi: 10.1080/00207454.2019.1702549
Esmaeili, M. A., Panahi, M., Yadav, S., Hennings, L., Kiaei, M. (2013). Premature Death of TDP-43 (A315T) Transgenic Mice Due to Gastrointestinal Complications Prior to Development of Full Neurological Symptoms of Amyotrophic Lateral Sclerosis. Int. J. Exp. Pathol. 94, 56–64. doi: 10.1111/iep.12006
Fang, X. (2016). Potential Role of Gut Microbiota and Tissue Barriers in Parkinson’s Disease and Amyotrophic Lateral Sclerosis. Int. J. Neurosci. 126, 771–776. doi: 10.3109/00207454.2015.1096271
Fang, P., Kazmi, S. A., Jameson, K. G., Hsiao, E. Y. (2020). The Microbiome as a Modifier of Neurodegenerative Disease Risk. Cell Host Microbe 28, 201–222. doi: 10.1016/j.chom.2020.06.008
Fang, X., Wang, X., Yang, S., Meng, F., Wang, X., Wei, H., et al. (2016). Evaluation of the Microbial Diversity in Amyotrophic Lateral Sclerosis Using High-Throughput Sequencing. Front. Microbiol. 7, 1479. doi: 10.3389/fmicb.2016.01479
Ferrer-Donato, A., Contreras, A., Frago, L. M., Chowen, J. A., Fernandez-Martos, C. M. (2021). Alterations in Leptin Signaling in Amyotrophic Lateral Sclerosis (ALS). Int. J. Mol. Sci. 22 (19), 10305. doi: 10.3390/ijms221910305
Figueroa-Romero, C., Guo, K., Murdock, B. J., Paez-Colasante, X., Bassis, C. M., Mikhail, K. A., et al. (2019). Temporal Evolution of the Microbiome, Immune System and Epigenome With Disease Progression in ALS Mice. Dis. Model Mech. 13 (2), dmm041947. doi: 10.1242/dmm.041947
Fleck, A. K., Hucke, S., Teipel, F., Eschborn, M., Janoschka, C., Liebmann, M., et al. (2021). Dietary Conjugated Linoleic Acid Links Reduced Intestinal Inflammation to Amelioration of CNS Autoimmunity. Brain 144 (4), 1152–1166. doi: 10.1093/brain/awab040
Gadoth, A., Nefussy, B., Bleiberg, M., Klein, T., Artman, I., Drory, V. E. (2015). Transglutaminase 6 Antibodies in the Serum of Patients With Amyotrophic Lateral Sclerosis. JAMA Neurol. 72, 676–681. doi: 10.1001/jamaneurol.2015.48
Group, A. L. S. U. (2016). ALSUntangled No. 32: Gluten-Free Diet. Amyotroph. Lateral Scler. Frontotemporal Degener. 17, 307–309. doi: 10.3109/21678421.2015.1096496
Guo, Y., Wang, Q., Zhang, K., An, T., Shi, P., Li, Z., et al. (2012). HO-1 Induction in Motor Cortex and Intestinal Dysfunction in TDP-43 A315T Transgenic Mice. Brain Res. 1460, 88–95. doi: 10.1016/j.brainres.2012.04.003
Ham, H., Lee, B. I., Oh, H. J., Park, S. H., Kim, J. S., Park, J. M., et al. (2017). A Case of Celiac Disease With Neurologic Manifestations Misdiagnosed as Amyotrophic Lateral Sclerosis. Intest. Res. 15, 540–542. doi: 10.5217/ir.2017.15.4.540
Hatzipetros, T., Bogdanik, L. P., Tassinari, V. R., Kidd, J. D., Moreno, A. J., Davis, C., et al. (2014). C57BL/6J Congenic Prp-TDP43A315T Mice Develop Progressive Neurodegeneration in the Myenteric Plexus of the Colon Without Exhibiting Key Features of ALS. Brain Res. 1584, 59–72. doi: 10.1016/j.brainres.2013.10.013
Herdewyn, S., Cirillo, C., Van Den Bosch, L., Robberecht, W., Vanden Berghe, P., Van Damme, P. (2014). Prevention of Intestinal Obstruction Reveals Progressive Neurodegeneration in Mutant TDP-43 (A315T) Mice. Mol. Neurodegener. 9, 24. doi: 10.1186/1750-1326-9-24
Hertzberg, V. S., Singh, H., Fournier, C. N., Moustafa, A., Polak, M., Kuelbs, C. A., et al. (2022). Gut Microbiome Differences Between Amyotrophic Lateral Sclerosis Patients and Spouse Controls. Amyotroph Lateral Scler Frontotemporal Degener 23, 91–99. doi: 10.1080/21678421.2021.1904994
Huang, H., Xu, H., Luo, Q., He, J., Li, M., Chen, H., et al. (2019). Fecal Microbiota Transplantation to Treat Parkinson’s Disease With Constipation: A Case Report. Med. (Baltimore) 98, e16163. doi: 10.1097/MD.0000000000016163
Kang, D. W., Adams, J. B., Coleman, D. M., Pollard, E. L., Maldonado, J., McDonough-Means, S., et al. (2019). Long-Term Benefit of Microbiota Transfer Therapy on Autism Symptoms and Gut Microbiota. Sci. Rep. 9, 5821. doi: 10.1038/s41598-019-42183-0
Kang, D. W., Adams, J. B., Gregory, A. C., Borody, T., Chittick, L., Fasano, A., et al. (2017). Microbiota Transfer Therapy Alters Gut Ecosystem and Improves Gastrointestinal and Autism Symptoms: An Open-Label Study. Microbiome 5, 10. doi: 10.1186/s40168-016-0225-7
Kwon, H. S., Koh, S. H. (2020). Neuroinflammation in Neurodegenerative Disorders: The Roles of Microglia and Astrocytes. Transl. Neurodegener. 9, 42. doi: 10.1186/s40035-020-00221-2
Labarre, A., Guitard, E., Parker, J. A. (2017). Variation of Gut Microbiome Rescues Paralysis and Neurodegeneration Profiles in C. Elegans ALS Models. Amyotrophic Lateral Scler Frontotemporal Degener, 245–246.
Lee, S. H., Yang, E. J. (2018). Relationship Between Liver Pathology and Disease Progression in a Murine Model of Amyotrophic Lateral Sclerosis. Neurodegener. Dis. 18, 200–207. doi: 10.1159/000491392
Luesma, M. J., Cantarero, I., Álvarez-Dotu, J. M., Santander, S., Junquera, C. (2014). New Insights Into C-Ret Signalling Pathway in the Enteric Nervous System and its Relationship With ALS. BioMed. Res. Int. 2014, 328348. doi: 10.1155/2014/328348
Lu, R., Zhang, Y. G., Xia, Y., Zhang, J., Kaser, A., Blumberg, R., et al. (2021). Paneth Cell Alertness to Pathogens Maintained by Vitamin D Receptors. Gastroenterology 160, 1269–1283. doi: 10.1053/j.gastro.2020.11.015
Mandrioli, J., Amedei, A., Cammarota, G., Niccolai, E., Zucchi, E., D’Amico, R., et al. (2019). FETR-ALS Study Protocol: A Randomized Clinical Trial of Fecal Microbiota Transplantation in Amyotrophic Lateral Sclerosis. Front. Neurol. 10, 1021. doi: 10.3389/fneur.2019.01021
Mazzini, L., Mogna, L., De Marchi, F., Amoruso, A., Pane, M., Aloisio, I., et al. (2018). Potential Role of Gut Microbiota in ALS Pathogenesis and Possible Novel Therapeutic Strategies. J. Clin. Gastroenterol. 52, S68–S70. Proceedings from the 9th Probiotics, Prebiotics and New Foods, Nutraceuticals and Botanicals for Nutrition & Human and Microbiota Health Meeting, held in Rome, Italy from September 10 to 12, 2017. doi: 10.1097/MCG.0000000000001042
McCombe, P. A., Henderson, R. D. (2011). The Role of Immune and Inflammatory Mechanisms in ALS. Curr. Mol. Med. 11, 246–254. doi: 10.2174/156652411795243450
Miller, R. G., Mitchell, J. D., Moore, D. H. (2012). Riluzole for Amyotrophic Lateral Sclerosis (ALS)/motor Neuron Disease (MND). Cochrane Database Syst. Rev., CD001447. doi: 10.1002/14651858.CD001447.pub3
Minalyan, A., Gabrielyan, L., Scott, D., Jacobs, J., Pisegna, J. R. (2017). The Gastric and Intestinal Microbiome: Role of Proton Pump Inhibitors. Curr. Gastroenterol. Rep. 19, 42. doi: 10.1007/s11894-017-0577-6
Murch, S. J., Cox, P. A., Banack, S. A., Steele, J. C., Sacks, O. W. (2004). Occurrence of Beta-Methylamino-L-Alanine (BMAA) in ALS/PDC Patients From Guam. Acta Neurol Scand 110, 267–269. doi: 10.1111/j.1600-0404.2004.00320.x
Nakano, Y., Hirayama, K., Terao, K. (1987). Hepatic Ultrastructural Changes and Liver Dysfunction in Amyotrophic Lateral Sclerosis. Arch. Neurol. 44, 103–106. doi: 10.1001/archneur.1987.00520130079022
Ngo, S. T., Restuadi, R., McCrae, A. F., Van Eijk, R. P., Garton, F., Henderson, R. D., et al. (2020). Progression and Survival of Patients With Motor Neuron Disease Relative to Their Fecal Microbiota. Amyotroph. Lateral Scler. Frontotemporal Degener. 21, 549–562. doi: 10.1080/21678421.2020.1772825
Niccolai, E., Di Pilato, V., Nannini, G., Baldi, S., Russo, E., Zucchi, E., et al. (2021). The Gut Microbiota-Immunity Axis in ALS: A Role in Deciphering Disease Heterogeneity? Biomedicines 9, 1–17. doi: 10.3390/biomedicines9070753
Nicolas, A., Kenna, K. P., Renton, A. E., Ticozzi, N., Faghri, F., Chia, R., et al. (2018). Genome-Wide Analyses Identify KIF5A as a Novel ALS Gene. Neuron 97, 1268–126+. doi: 10.1016/j.neuron.2018.02.027
Niesler, B., Kuerten, S., Demir, I. E., Schafer, K. H. (2021). Disorders of the Enteric Nervous System - a Holistic View. Nat. Rev. Gastroenterol. Hepatol. 18, 393–410. doi: 10.1038/s41575-020-00385-2
Novak, V., Rogelj, B., Zupunski, V. (2021). Therapeutic Potential of Polyphenols in Amyotrophic Lateral Sclerosis and Frontotemporal Dementia. Antioxidants (Basel) 10 (8), 1328. doi: 10.3390/antiox10081328
Nubling, G. S., Mie, E., Bauer, R. M., Hensler, M., Lorenzl, S., Hapfelmeier, A., et al. (2014). Increased Prevalence of Bladder and Intestinal Dysfunction in Amyotrophic Lateral Sclerosis. Amyotroph. Lateral Scler. Frontotemporal. Degener. 15, 174–179. doi: 10.3109/21678421.2013.868001
Ogbu, D., Xia, E., Sun, J. (2020). Gut Instincts: Vitamin D/vitamin D Receptor and Microbiome in Neurodevelopment Disorders. Open Biol. 10, 200063. doi: 10.1098/rsob.200063
Paganoni, S., Macklin, E. A., Hendrix, S., Berry, J. D., Elliott, M. A., Maiser, S., et al. (2020). Trial of Sodium Phenylbutyrate-Taurursodiol for Amyotrophic Lateral Sclerosis. N Engl. J. Med. 383, 919–930. doi: 10.1056/NEJMoa1916945
Pape, J. A., Grose, J. H. (2020). The Effects of Diet and Sex in Amyotrophic Lateral Sclerosis. Rev. Neurol. (Paris) 176, 301–315. doi: 10.1016/j.neurol.2019.09.008
Philips, T., Rothstein, J. D. (2015). Rodent Models of Amyotrophic Lateral Sclerosis. Curr. Protoc. Pharmacol. 69, 5 67 1–5 67 21. doi: 10.1002/0471141755.ph0567s69
Quick, D. T., Greer, M. (1967). Pancreatic Dysfunction in Patients With Amyotrophic Lateral Sclerosis. Neurology 17, 112–116. doi: 10.1212/WNL.17.2.112
Rao, M., Gershon, M. D. (2016). The Bowel and Beyond: The Enteric Nervous System in Neurological Disorders. Nat. Rev. Gastroenterol. Hepatol. 13, 517–528. doi: 10.1038/nrgastro.2016.107
Roberts, A. L., Johnson, N. J., Cudkowicz, M. E., Eum, K. D., Weisskopf, M. G. (2016). Job-Related Formaldehyde Exposure and ALS Mortality in the USA. J. Neurol. Neurosurg Psychiatry 87, 786–788. doi: 10.1136/jnnp-2015-310750
Rowin, J., Xia, Y., Jung, B., Sun, J. (2017). Gut Inflammation and Dysbiosis in Human Motor Neuron Disease. Physiol. Rep. 5, 1–6. doi: 10.14814/phy2.13443
Sabel, C. E., Boyle, P. J., Loytonen, M., Gatrell, A. C., Jokelainen, M., Flowerdew, R., et al. (2003). Spatial Clustering of Amyotrophic Lateral Sclerosis in Finland at Place of Birth and Place of Death. Am. J. Epidemiol. 157, 898–905. doi: 10.1093/aje/kwg090
Solanki, I., Parihar, P., Mansuri, M. L., Parihar, M. S. (2015). Flavonoid-Based Therapies in the Early Management of Neurodegenerative Diseases. Adv. Nutr. 6, 64–72. doi: 10.3945/an.114.007500
Sun, J., Zhan, Y., Mariosa, D., Larsson, H., Almqvist, C., Ingre, C., et al. (2019). Antibiotics Use and Risk of Amyotrophic Lateral Sclerosis in Sweden. Eur. J. Neurol. 26, 1355–1361. doi: 10.1111/ene.13986
Tedeschi, V., Petrozziello, T., Secondo, A. (2019). Calcium Dyshomeostasis and Lysosomal Ca(2+) Dysfunction in Amyotrophic Lateral Sclerosis. Cells 8 (10), 1216. doi: 10.3390/cells8101216
Tortelli, R., Zecca, C., Piccininni, M., Benmahamed, S., Dell’Abate, M. T., Barulli, M. R., et al. (2020). Plasma Inflammatory Cytokines Are Elevated in ALS. Front. Neurol. 11, 552295. doi: 10.3389/fneur.2020.552295
Turnbull, J. (2020). Sodium Phenylbutyrate-Taurursodiol for ALS. N. Engl. J. Med. 383, 2293–2294. doi: 10.1056/NEJMc2030710
Turner, M. R., Al-Chalabi, A., Chio, A., Hardiman, O., Kiernan, M. C., Rohrer, J. D., et al. (2017). Genetic Screening in Sporadic ALS and FTD. J. Neurol Neurosurg Psychiatry 88, 1042–1044. doi: 10.1136/jnnp-2017-315995
Turner, M. R., Chohan, G., Quaghebeur, G., Greenhall, R. C., Hadjivassiliou, M., Talbot, K. (2007). A Case of Celiac Disease Mimicking Amyotrophic Lateral Sclerosis. Nat. Clin. Pract. Neurol. 3, 581–584. doi: 10.1038/ncpneuro0631
Utterback, R. A., Cummins, A. J., Cape, C. A., Goldenberg, J. (1970). Pancreatic Function in Amyotrophic Lateral Sclerosis. J. Neurol. Neurosurg Psychiatry 33, 544–547. doi: 10.1136/jnnp.33.4.544
Volk, A. E., Weishaupt, J. H., Andersen, P. M., Ludolph, A. C., Kubisch, C. (2018). Current Knowledge and Recent Insights Into the Genetic Basis of Amyotrophic Lateral Sclerosis. Med. Genet-Berlin 30, 252–258. doi: 10.1007/s11825-018-0185-3
Wang, M. D., Little, J., Gomes, J., Cashman, N. R., Krewski, D. (2017). Identification of Risk Factors Associated With Onset and Progression of Amyotrophic Lateral Sclerosis Using Systematic Review and Meta-Analysis. Neurotoxicology 61, 101–130. doi: 10.1016/j.neuro.2016.06.015
Weisskopf, M. G., Morozova, N., O’Reilly, E. J., McCullough, M. L., Calle, E. E., Thun, M. J., et al. (2009). Prospective Study of Chemical Exposures and Amyotrophic Lateral Sclerosis. J. Neurol. Neurosurg. Psychiatry 80, 558–561. doi: 10.1136/jnnp.2008.156976
Whipps, J., Lewis, K., Cooke, R. (1988). “Mycoparasitism and Plant Disease Control,” in Fungi in Biological Control Systems. Ed. M, B. (Manchester: Manchester University Press), 161–187.
Wingate, D. (1999). Gastrointestinal Dysfunction in ALS. Amyotroph Lateral Scler Other Motor Neuron Disord. 1, 2.
Wu, S., Yi, J., Zhang, Y. G., Zhou, J., Sun, J. (2015). Leaky Intestine and Impaired Microbiome in an Amyotrophic Lateral Sclerosis Mouse Model. Physiol. Rep. 3, 1–10. doi: 10.14814/phy2.12356
Xu, H. M., Huang, H. L., Zhou, Y. L., Zhao, H. L., Xu, J., Shou, D. W., et al. (2021). Fecal Microbiota Transplantation: A New Therapeutic Attempt From the Gut to the Brain. Gastroenterol. Res. Pract. 2021, 6699268. doi: 10.1155/2021/6699268
Yanez, M., Galan, L., Matias-Guiu, J., Vela, A., Guerrero, A., Garcia, A. G. (2011). CSF From Amyotrophic Lateral Sclerosis Patients Produces Glutamate Independent Death of Rat Motor Brain Cortical Neurons: Protection by Resveratrol But Not Riluzole. Brain Res. 1423, 77–86. doi: 10.1016/j.brainres.2011.09.025
Zeng, Q., Shen, J., Chen, K., Zhou, J., Liao, Q., Lu, K., et al. (2020). The Alteration of Gut Microbiome and Metabolism in Amyotrophic Lateral Sclerosis Patients. Sci. Rep. 10, 12998. doi: 10.1038/s41598-020-69845-8
Zhai, C. D., Zheng, J. J., An, B. C., Huang, H. F., Tan, Z. C. (2019). Intestinal Microbiota Composition in Patients With Amyotrophic Lateral Sclerosis: Establishment of Bacterial and Archaeal Communities Analyses. Chin. Med. J. (Engl) 132, 1815–1822. doi: 10.1097/CM9.0000000000000351
Zhang, Y. G., Lu, R., Xia, Y., Zhou, D., Petrof, E., Claud, E. C., et al. (2019). Lack of Vitamin D Receptor Leads to Hyperfunction of Claudin-2 in Intestinal Inflammatory Responses. Inflamm. Bowel Dis. 25, 97–110. doi: 10.1093/ibd/izy292
Zhang, R., Miller, R. G., Gascon, R., Champion, S., Katz, J., Lancero, M., et al. (2009). Circulating Endotoxin and Systemic Immune Activation in Sporadic Amyotrophic Lateral Sclerosis (sALS). J. Neuroimmunol. 206, 121–124. doi: 10.1016/j.jneuroim.2008.09.017
Zhang, Y. G., Ogbu, D., Garrett, S., Xia, Y., Sun, J. (2021). Aberrant Enteric Neuromuscular System and Dysbiosis in Amyotrophic Lateral Sclerosis. Gut Microbes 13, 1996848. doi: 10.1080/19490976.2021.1996848
Zhang, Y. G., Wu, S., Sun, J. (2013). Vitamin D, Vitamin D Receptor, and Tissue Barriers. Tissue Barriers 1 (1). doi: 10.4161/tisb.23118
Zhang, Y. G., Wu, S., Yi, J., Xia, Y., Jin, D., Zhou, J., et al. (2017). Target Intestinal Microbiota to Alleviate Disease Progression in Amyotrophic Lateral Sclerosis. Clin. Ther. 39, 322–336. doi: 10.1016/j.clinthera.2016.12.014
Zhang, L., Zhuang, Z., Zhang, G., Huang, T., Fan, D. (2022). Assessment of Bidirectional Relationships Between 98 Genera of the Human Gut Microbiota and Amyotrophic Lateral Sclerosis: A 2-Sample Mendelian Randomization Study. BMC Neurol. 22, 8. doi: 10.1186/s12883-021-02522-z
Keywords: amyotrophic lateral sclerosis (ALS), gastrointestinal digestion, enteric neuron, metabolite, microbiome, bacteria, CNS - central nervous system, motor neurodegeneration
Citation: Martin S, Battistini C and Sun J (2022) A Gut Feeling in Amyotrophic Lateral Sclerosis: Microbiome of Mice and Men. Front. Cell. Infect. Microbiol. 12:839526. doi: 10.3389/fcimb.2022.839526
Received: 20 December 2021; Accepted: 14 February 2022;
Published: 11 March 2022.
Edited by:
Gislane Lelis Vilela de Oliveira, São Paulo State University, BrazilReviewed by:
Yuishin Izumi, Tokushima University, JapanJessica Mandrioli, University of Modena and Reggio Emilia, Italy
Copyright © 2022 Martin, Battistini and Sun. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jun Sun, junsun7@uic.edu