Hedgehog signaling and the glioma-associated oncogene in cancer radioresistance
 Li Zhang
Li Zhang Yuhan Zhang2
Yuhan Zhang2  Shuai Xue
Shuai Xue- 1Nephrology Department, The 1st Hospital of Jilin University, Changchun, China
- 2General Surgery Center, Department of Thyroid Surgery, The 1st Hospital of Jilin University, Changchun, China
Tumor radioresistance remains a key clinical challenge. The Hedgehog (HH) signaling pathway and glioma-associated oncogene (GLI) are aberrantly activated in several cancers and are thought to contribute to cancer radioresistance by influencing DNA repair, reactive oxygen species production, apoptosis, autophagy, cancer stem cells, the cell cycle, and the tumor microenvironment. GLI is reported to activate the main DNA repair pathways, to interact with cell cycle regulators like Cyclin D and Cyclin E, to inhibit apoptosis via the activation of B-cell lymphoma-2, Forkhead Box M1, and the MYC proto-oncogene, to upregulate cell stemness related genes (Nanog, POU class 5 homeobox 1, SRY-box transcription factor 2, and the BMI1 proto-oncogene), and to promote cancer stem cell transformation. The inactivation of Patched, the receptor of HH, prevents caspase-mediated apoptosis. This causes some cancer cells to survive while others become cancer stem cells, resulting in cancer recurrence. Combination treatment using HH inhibitors (including GLI inhibitors) and conventional therapies may enhance treatment efficacy. However, the clinical use of HH signaling inhibitors is associated with toxic side effects and drug resistance. Nevertheless, selective HH agonists, which may relieve the adverse effects of inhibitors, have been developed in mouse models. Combination therapy with other pathway inhibitors or immunotherapy may effectively overcome resistance to HH inhibitors. A comprehensive cancer radiotherapy with HH or GLI inhibitor is more likely to enhance cancer treatment efficacy while further studies are still needed to overcome its adverse effects and drug resistance.
1 Hedgehog (HH) signaling and the glioma-associated oncogene (GLI) protein
The evolutionarily conserved Hedgehog (HH) signaling pathway plays important roles during embryonic development and tumorigenesis by influencing various processes, including cell proliferation, differentiation, angiogenesis, and apoptosis (Jiang, 2022). Although HH signaling is generally inactive in adult tissues and organs, its activation maintains somatic cell renewal and regeneration through the induction of the differentiation of stem cells and pluripotent cells for tissue repair and epithelium replacement (wound healing) in various tissues, including lungs, prostate, exocrine pancreatic cells, the skin, and neural organs (Skoda et al., 2018). Moreover, in primary cilia, HH signaling is vital for the reception of thermal, chemical, and mechanical signals (Fattahi et al., 2018). The presence of most factors involved in canonical HH signaling in the cilia is dynamic. HH signaling activation downregulates the levels of some factors, such as the twelve-span transmembrane protein, Patched (PTCH), in the ciliary tips, whereas others like SMO (a seven-span transmembrane G protein-coupled receptor), GLI2, GLI3, suppressor of fused homolog (SUFU), and the microtubule-associated atypical kinesin family member 7 (KIF7), are upregulated. KIF7 regulates Sonic Hedgehog (SHH) signaling by modifying cilial structure (Liem et al., 2009). HH signaling also depletes the levels of G protein-coupled receptor 161 (GPR161) in primary cilia, and this process depends on SMO and β-arrestin (Pal et al., 2016). β-arrestins, which are adaptor proteins, are recruited to the proximal C-terminus of GPR161 through G protein-coupled receptor kinase (GRK) 2. Moreover, GRK2 and GRK3 can transmit high-level SHH signals independently of GPR161. This suggests that multiple factors contribute to the dynamic regulation of HH signaling components in the cilia. HH signaling has also been reported to influence tumorigenesis, tumor progression, and treatment resistance through mechanisms that act on various cell types, such as basal, pancreatic, esophageal, gastric, prostate, and breast cancer cells (Briscoe and Thérond, 2013; Bakshi et al., 2017; Xu et al., 2019). In cancer, aberrant HH signaling activation is classified as type I, II, or III. Type I, which is autonomous and ligand-independent, usually occurs in meningiomas and rhabdomyosarcomas (Clark et al., 2013). Type II, the ligand-dependent oncogenic form, acts in an autocrine/juxtacrine manner and mostly occurs in breast and lung cancers. Type III, which is ligand-dependent and involves paracrine or reverse paracrine factors, is usually activated in prostate and pancreatic cancers (Jiang and Hui, 2008; Efstathiou et al., 2013; Steele et al., 2021).
HH signaling pathway components include HH ligands, PTCH (a twelve-span transmembrane protein), SMO (a seven-span transmembrane member of the G protein-coupled receptor family), transcription factors (GLI1, GLI2, and GLI3), SUFU, and several HH signaling target genes, including Cyclin D, Cyclin E, Forkhead box M1 (FoxMI), and MYC proto-oncogene (c-Myc) (Suchors and Kim, 2022) (Figure 1).
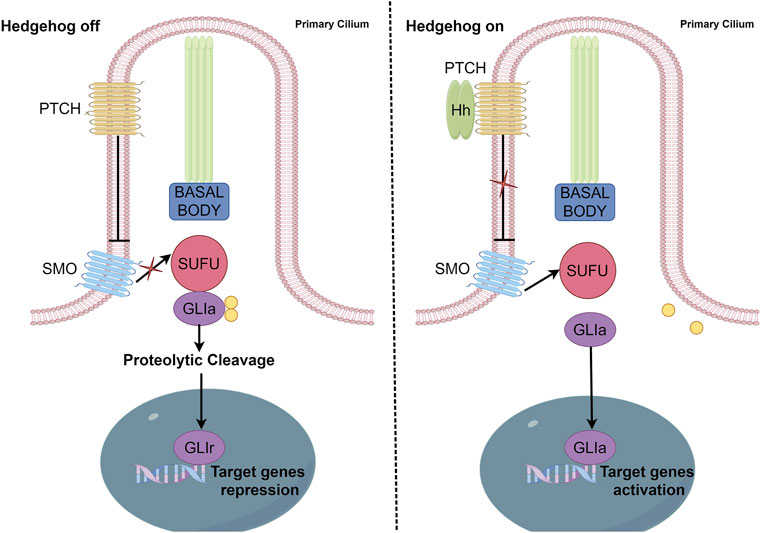
FIGURE 1. Canonical activation of the HH pathway. In canonical HH signaling, the transcription of HH target genes is suppressed in the absence of the HH ligand (left panel) and activated in its presence (right panel). Critical components of this network include PTCH, SMO, GLI activator (GL1a), GLI repressor (GLIr), and various negative regulators, including SUFU.
GLI is a zinc finger protein named after glioblastoma, from which it was first identified. Vertebrates possess three GLI genes, GLI1, GLI2, and GLI3, which are C2H2 Kruppel-type transcription factors with five zinc finger domains (ZF1–ZF5) (Niewiadomski et al., 2019). GLI proteins regulate the expression of target genes by directly binding to their promoters. Specifically, GLI1 and GLI3 recognize the 5′-GACCACCCA-3′ sequence within the promoters of their target genes, whereas GLI2 recognizes the almost identical, 5′-GAACCACCCA-3′, sequence. In the absence of HH ligands, PTCH suppresses SMO signaling (Matise and Joyner, 1999). Phosphorylation of GLI2 and GLI3 by protein kinase A (PKA), glycogen synthase kinase 3 (GSK3), and casein kinase 1 (CK1) causes the cleavage of their N-terminal domains, resulting in C-terminally truncated repressor forms, termed GLIr. These truncated forms then translocate into the nucleus, where they suppress the transcription of downstream genes. The binding of HH ligands to PTCH relieves SMO inhibition, which then inhibits the phosphorylation of GLI2 and GLI3, thereby allowing their full-length forms to translocate into the nucleus and drive target gene transcription (Matissek and Elsawa, 2020). However, GLI1 proteins that lack the N-terminal inhibitory region only function as transcriptional activators. SUFU is a key regulator of the HH pathway (Ruiz i Altaba, 1999) and the activation of HH ligands and SMO can cause SUFU ubiquitination and proteasomal degradation. In addition to canonical GLI activation through the HH–PTCH–SMO pathway, which is commonly observed in normal cells, accumulating evidence indicates the existence of SMO-independent GLI function induction in cancer. Such non-canonical activation involves the transcriptional activation of GLI genes and the post-translational modification of GLI proteins, resulting in various modifications that contribute to the progression of various cancers characterized by GLI signaling elevation. Studies have also reported non-canonical signaling activation in which the expression and activation of GLI transcription factors are regulated by other signaling pathways and proteins, including transforming growth factor beta (TGF-β), epidermal growth factor receptor (EGFR), ribosomal protein S6 kinase beta-1(S6K1)/mammalian target of rapamycin (mTOR), mitogen-activated protein kinase (MAPK), and fibroblast growth factor (FGF) signaling. Additionally, GLI is regulated by bromodomain and extra-terminal domain (BET) proteins and histone deacetylases (HDACs) (Ruiz i Altaba et al., 2002; Lauth and Toftgård, 2007; Canettieri et al., 2010; Long et al., 2014; Huang et al., 2016) (Figure 2).
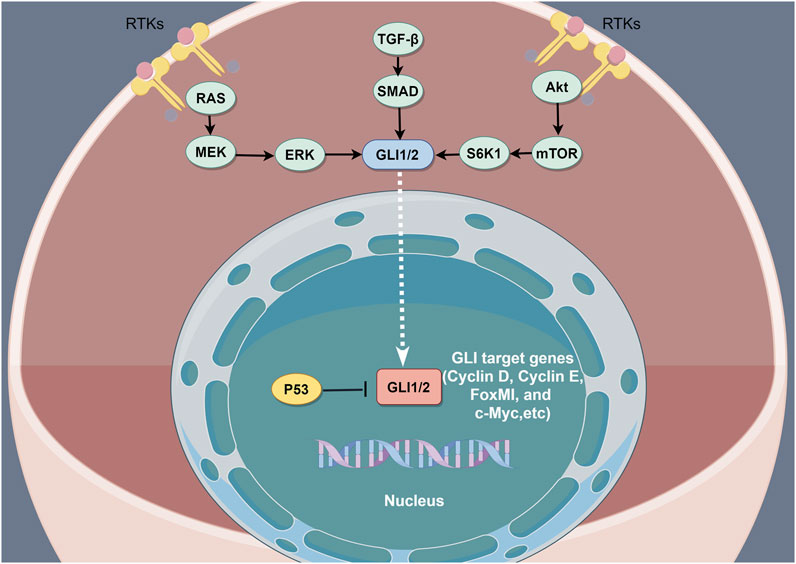
FIGURE 2. The non-canonical activation of the HH pathway by oncogenic pathways. The non-canonical activation of GLI by other pathways is a key driver of resistance to HH inhibition. The combined inhibition of the non-canonical HH activation pathway is considered an effective method of overcoming resistance to HH inhibitors.
2 Mechanisms of cancer radioresistance
Radiotherapy (RT) is the most common method of cancer treatment and is widely used in combination with other treatments, such as surgery, chemotherapy, or immunotherapy. Based on various factors, such as tumor radiosensitivity, RT can be defined as curative, adjuvant, or palliative (Schaue and McBride, 2015). When some radiosensitive tumors, such as lymphomas, carcinomas of the larynx, prostate, or cervix, and several central nervous system neoplasms are treated at an early stage, RT can potentially kill all tumor cells and cure the cancer. For adjuvant purposes, RT is usually used together with surgery. RT can be used preoperatively to shrink tumors, thereby facilitating subsequent R0 resection. Additionally, RT can be used intraoperatively to precisely deliver large doses of radioactive particles to the tumor site while minimizing adverse effects on normal tissues. Postoperatively, RT can be used to lower the risk of tumor recurrence, such as the use of radioactive iodine for differentiated thyroid carcinoma after total thyroidectomy. For palliative purposes, RT can be used to reduce or eliminate the compression symptoms of unresectable tumors in the central or peripheral nervous system, as well as in blood vessels (Citrin, 2017). However, cancer radioresistance is a key clinical challenge (Gong et al., 2021).
2.1 DNA repair and reactive oxygen species (ROS) production
In general, the use of RT for cancer treatment involves high-energy photon radiation, such as X-rays, gamma (g) rays, and particle radiation. The particles used for RT include alpha (a) and beta (b) particles, electrons (e), protons, carbon ions, or neutron beams. These types of radiation can be used for various therapeutic purposes (Santivasi and Xia, 2014). The principle behind the use of RT is based on how ionizing radiation interacts with the molecular components of tumor cells, which can be direct or indirect. Direct ionizing radiation damages biomolecules within the cells, such as proteins, lipids, and DNA. Of these, radiation impacts DNA the most. DNA damage can stop cell division and proliferation, resulting in cell death via necrosis or apoptosis. The indirect effects of radiation involve the destruction of biomolecules through free radicals, particularly ROS, which is mainly caused by water radiolysis. ROS contain unpaired electrons that can damage biomolecules through various chemical reactions, including hydrogen extraction, addition, disproportionation, and electron capture. Such reactions may cause structural biomolecule damage, including single-strand or double-strand DNA breaks and DNA–DNA or DNA–protein cross-linking, which result in cell death. ROS are crucial for RT because they damage biomolecules and activate signaling pathways that promote tumor cell apoptosis (Nikjoo et al., 2016; Srinivas et al., 2019; Li, 2023).
DNA breakage is the main mechanism through which RT kills tumor cells and the ability of tumor cells to repair DNA damage determines their likelihood of developing radioresistance. In cancer cells, DNA damage is mainly repaired through non-homologous end joining (NHEJ) and homologous recombination (Sinha and Häder, 2002). Currently, NHEJ is considered the main DNA repair mechanism. In NHEJ, first, the heterodimer Ku70/Ku80 binds to the ends of the broken DNA double strand, enveloping it. It then recruits DNA protein kinase-dependent catalytic subunits (DNA PKcs). The mutual binding of DNA PKcs not only promotes the reaggregation of the broken ends but also allows two DNA PKcs molecules to interact and repair the DNA (Sinha and Häder, 2002). With the help of the NHEJ-specific nuclease, Artemis, the irreparable terminal DNA sequence is excised. Finally, X-rays are used to stagger the complementary repair proteins, which stabilize DNA ligase IV, thereby stimulating its activity and the ligation of the DNA ends. Homologous recombination mainly generates Rad51 nuclear protein filaments, which then recruit double-stranded DNA molecules to identify homologous sequences and form replacement rings (Shibata and Jeggo, 2020). DNA resynthesis then begins and progresses, forming a Holliday linker. Through the activity of various enzymes, precise DNA repair can be achieved.
2.2 Apoptosis and autophagy
Apoptosis is a form of programmed cell death. Tumor cell radiosensitivity depends on the capacity of radiation to induce apoptosis in as many tumor cells as possible. Radioresistant tumor cells often lack apoptosis activation mechanisms (Jiao et al., 2022). However, an analysis by Japanese researchers, in which tumor cells were irradiated and observed daily using contrast microscopy, found that most irradiated tumor cells did not exhibit obvious apoptotic features, such as pyknosis, chromatin breakage, and apoptotic bodies. However, they all had LC3-positive autophagosomes, suggesting that autophagy is more crucial in the regulation of tumor cell radiosensitivity than apoptosis (Kuwahara et al., 2018). Autophagy is mainly regulated by phosphatidylinositol 3-kinase protein kinase B (a rapamycin target protein), unfolded protein response, and mitogen-activated protein kinase signaling. Therefore, drugs or molecular techniques that act on one or more of the genes in these signaling pathways are frequently used to manipulate autophagy in irradiated tumor cells, thereby altering their radiosensitivity (Patel et al., 2020). Some studies have shown that autophagy is deployed as a protective mechanism by irradiated cells and that autophagy inhibition can radiosensitize tumor cells (Digomann et al., 2019; Patel et al., 2021). However, mounting evidence has confirmed that inducing autophagy through drugs or gene regulation promotes radiosensitivity (Huang et al., 2014; Sannigrahi et al., 2015). Hence, researchers now sensitize cancer cells via autophagy induction (Yang et al., 2015; Taylor et al., 2018).
2.3 Cancer stem cells (CSCs)
CSCs are a small population of cancer cells with the ability to self-renew and differentiate into multiple lineages (Hoque et al., 2023). Studies have found that CSCs not only maintain tumor growth but also play critical roles in chemotherapy and radioresistance (Abad et al., 2020; Olivares-Urbano et al., 2020). When compared with other tumor cells, CSCs are more radioresistant. Moreover, radiation triggers additional gene mutations or epigenetic regulation in CSCs, which may enhance radioresistance. Therefore, regulating signaling pathways that maintain the self-renewal and multi-directional differentiation of CSCs, such as Wnt, HH, and Notch signaling, may enhance tumor radiosensitivity (Koch et al., 2010; Kim et al., 2015; Schulz et al., 2019). In some tumors, especially breast cancer and glioma, signaling pathways that regulate CSC maintenance can influence tumor radiosensitivity. Moreover, the correlation between CSCs and cancer radiosensitivity has been well summarized. CSCs have been implicated in the development of primary and secondary cancer resistance to RT (Rycaj and Tang, 2014). This has been attributed to various radioresistance regulatory mechanisms (Ogawa et al., 2013; Krause et al., 2017; Abad et al., 2020), including the following: 1) CSCs having superior DNA damage repair ability (Abad et al., 2020), 2) CSCs being able to activate cell cycle checkpoint kinases earlier and more intensely while delaying the cell cycle and prolonging DNA repair time (Caglar and Biray Avci, 2020), 3) CSCs expressing higher levels of anti-apoptotic proteins and autophagy factors while suppressing radiation-induced apoptosis and enhancing radioresistance via autophagy (Babaei et al., 2021), 4) CSCs having the ability to self-renew and differentiate into several lineages, with mutations or epigenetic modifications that promote the development of secondary radiation resistance (Zhu et al., 2020), and 5), CSCs having a more efficient ROS-scavenging system and therefore being able to withstand radiation-induced oxidative stress (Biswas et al., 2022).
2.4 The cell cycle
The eukaryotic cell cycle is mainly regulated by cyclin and cyclin-dependent kinases. Cells in different cell cycle stages have different levels of radiosensitivity. Generally, cells in the gap 2 (G2) and mitotic (M) phases are more sensitive to radiation than those in the synthesis (S) phase (Pawlik and Keyomarsi, 2004). Therefore, effective cell cycle control and the accumulation of tumor cells in the late G2/M stage is an important strategy for improving tumor cell radiosensitivity (Wilson, 2004). Additionally, DNA double-strand breaks activate cell cycle checkpoints, thereby acting on cell cycle regulatory protein/kinase complexes to slow down cell cycle progression and provide sufficient time for effective DNA repair before the cell enters the S or M phases (Alexandrou and Li, 2014). Thus, treating tumor cells with cell cycle checkpoint inhibitors can reduce the time available for tumor cells to repair DNA after RT, thereby improving their radiosensitivity (Ishikawa et al., 2006).
2.5 The tumor microenvironment (TME)
Tumors not only contain a diverse pool of cancer cells, but also a range of cellular and non-cellular factors, including fibroblasts, immune cells, soluble growth factors, the extracellular matrix, and the vasculature (Arneth, 2019), which are collectively referred to as the TME. The nature of the interaction between tumor cells and their microenvironment significantly impacts tumor initiation, progression, metastasis, and eradication (Suwa et al., 2021). For instance, hypoxic conditions elevate the levels of hypoxia-inducible factor 1-alpha (HIF-1α), which, when present in sufficient quantities, activates HH signaling in pancreatic cancer cells, thereby promoting epithelial–mesenchymal transition and invasion (Jarosz-Biej et al., 2019; Zhao et al., 2021; Taeb et al., 2022). Additionally, an acidic microenvironment caused by tumor-derived lactic acid inhibits lactic acid excretion from cytotoxic T-cell lymphocytes, which adversely affects their metabolism and immune functions, including cytokine production, infiltration, and proliferation (Fischer et al., 2007; Barker et al., 2015). The loss of T-cell function suppresses the clearance of tumor cells or damaged cells by the immune system. Activated macrophages stimulate the expression of cyclooxygenase-2 (COX-2), an inflammatory prostaglandin synthase, through the activation of nuclear factor κB (NF-κB). The resulting prostaglandin E2 (PGE2) expedites tumor growth and radioresistance in various cancers (Logan et al., 2007; Dewhirst and Chi, 2013). Furthermore, radiation-mediated macrophage activation can induce radioresistance in cancer cells and enable tumor recurrence after radiotherapy by upregulating tumor necrosis factor-α (TNF-α) and angiogenesis (Logan et al., 2007). The interplay between macrophages and cancer cells is a crucial factor in tumor growth and radioresistance. Macrophage stimulation by cancer cell-secreted interleukin 6 (IL6), macrophage colony-stimulating factor (M-CSF), and PGE2 triggers their polarization into the M2 subtype (Chen et al., 2019; Xiao and Yu, 2021). Additionally, the secretion of proangiogenic factors, such as vascular endothelial growth factor (VEGF) and platelet-derived growth factor (PDGF) by M2 macrophages is reported to establish a tumor-conducive microenvironment that promotes tumor growth even after RT (Meng et al., 2010; Chen et al., 2019). Moreover, the secretion of C–C motif chemokine ligand 22 (CCL22) by M2 macrophages triggers regulatory T cell infiltration, which suppresses antitumor immunity, thereby boosting tumor growth and radioresistance (Xiao and Yu, 2021; Vattai et al., 2023). Additionally, the secretion of PDGF and TGF-β by cancer cells induces stromal fibroblast trans-differentiation into myofibroblasts, resulting in the formation of “cancer-associated fibroblasts” in the tumor tissue (Caja et al., 2018). Furthermore, it has been reported that macrophage-produced TNF-α stimulates fibroblast proliferation during wound healing. This mechanism has also been reported to create a fibroblast-rich TME (Wu and Dai, 2017). Because the cancer-associated fibroblast-rich microenvironment is known to occur in the hypoxic regions of the tumor, it is thought to induce radioresistance (Wang et al., 2019; Khosravi et al., 2020).
3 HH/GLI signaling and radioresistance
HH signaling is a developmental pathway that is active during embryonal development as well as in adult stem cell maintenance, and tissue repair and regeneration. Dysregulated HH signaling has been implicated in the development of several cancers. Moreover, HH signaling drives the transcription of cancer radioresistance genes (Table 1), including those involved in DNA repair, the cell cycle, apoptosis, angiogenesis, and the regulation of CSCs and the tumor microenvironment (Figure 3).
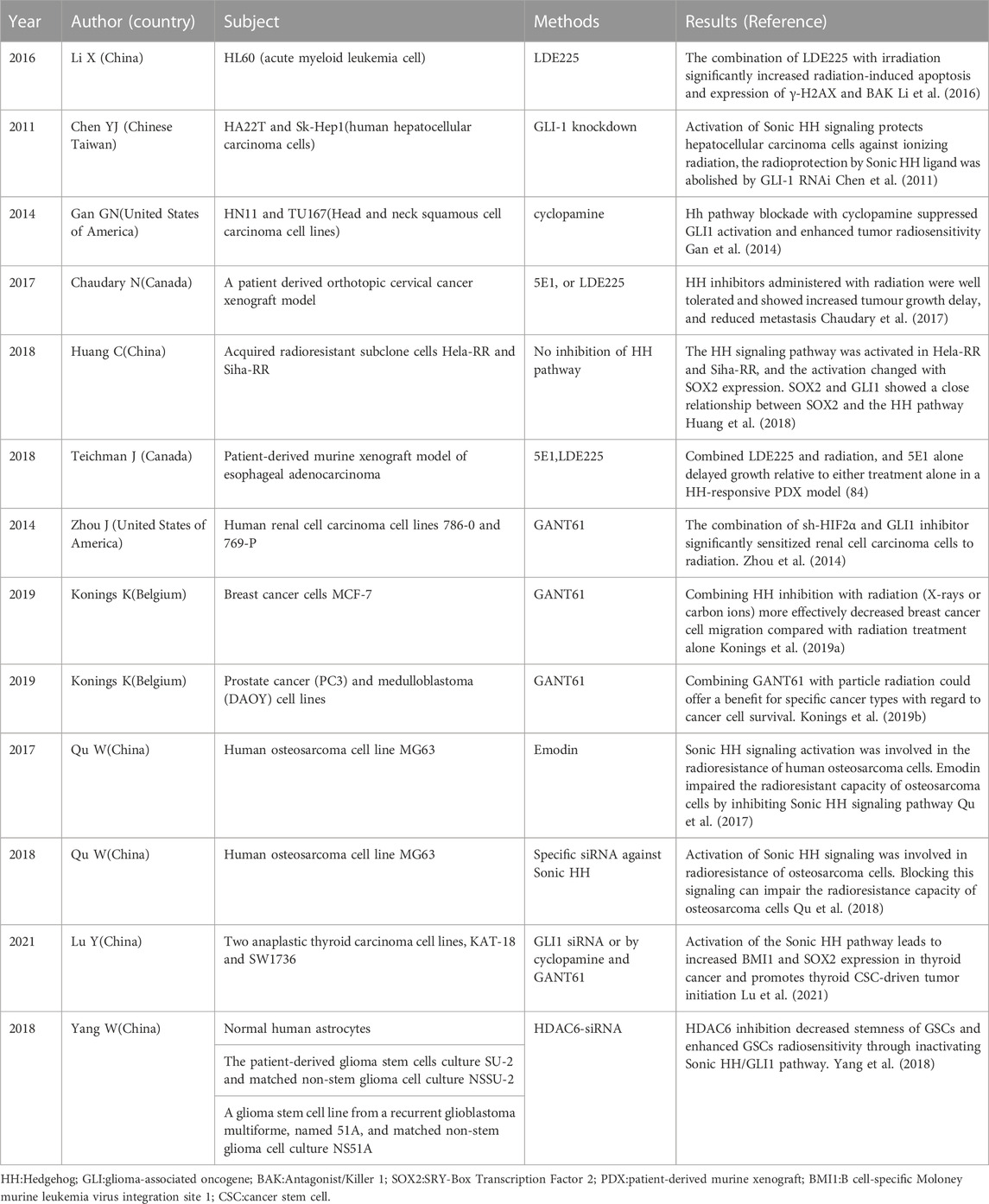
TABLE 1. Summary of HH/GLI signaling and radioresistance.
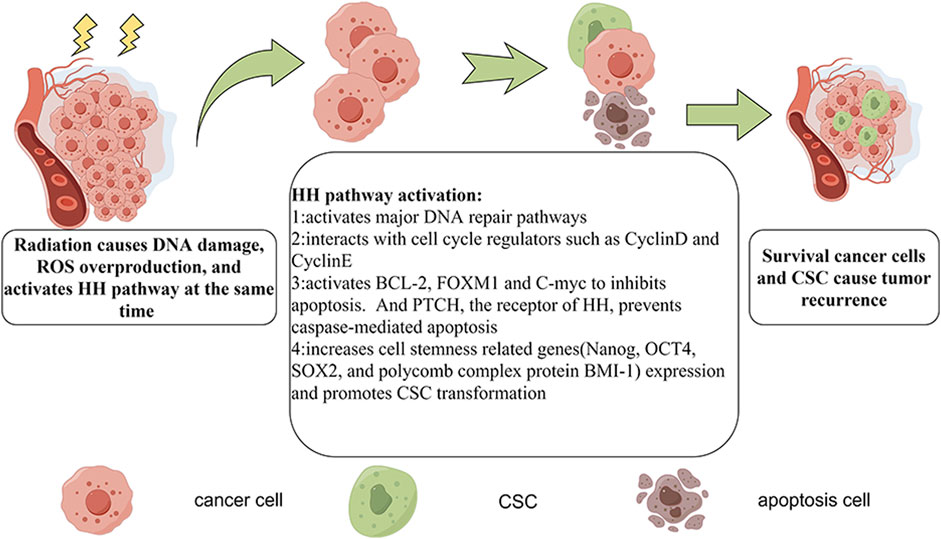
FIGURE 3. The mechanisms of HH pathway-mediated radioresistance. Radiotherapy triggers DNA damage and ROS overproduction, which are lethal to cancer cells. However, the HH pathway is also activated. GLI activates major DNA repair pathways, interacts with cell cycle regulators (e.g., Cyclin D and Cyclin E), activates BCL-2, FOXM1, and C-myc (thereby inhibiting apoptosis), upregulates cell stemness-related genes (e.g., Nanog, OCT4, SOX2, and BMI-1), and promotes CSC transformation. The inactivation of Patched, the receptor of HH, prevents caspase-mediated apoptosis. In this way, some cancer cells survive, whereas others become CSCs, which may lead to cancer recurrence.
3.1 The regulation of DNA repair, the cell cycle, and apoptosis by HH/GLI signaling
The combination of LDE225 (an HH signaling inhibitor) with irradiation is reported to significantly increase radiation-induced apoptosis, as well as the expression of γ-H2AX and B-cell lymphoma 2 (BCL2) antagonist/Killer 1 (BAK), in refractory acute myeloid leukemia (Li et al., 2016). These findings suggest that inhibiting the HH pathway can enhance radiosensitivity in acute myeloid leukemia cells by regulating DNA damage and apoptosis (Li et al., 2016). GLI1 inhibits the repair of DNA mismatch and DSBs by regulating MutL homolog 1 (MLH1) and ataxia telangiectasia-mutated protein kinase (ATR)/checkpoint kinase 1 (CHK1) signaling, respectively. GLI1 also activates nucleotide excision repair and DSB repair by regulating c-JUN and BH3 domain-only death agonist protein (BID)–ATR/CHK1 pathways, respectively. GLI1 response to DNA damage and its effect on subsequent DNA repair may depend on the extent of DNA damage and the characteristics of the specific cell line (Palle et al., 2015). GLI2 overexpression revealed that human keratinocytes develop resistance to ultraviolet B-induced exposure. However, BCL-2 inhibition restored natural genomic instability and DNA damage-induced apoptosis following ultraviolet B radiation. Therefore, abnormal GLI2 expression may significantly impact genomic stability in human epithelial cells and promote the survival of the descendant cells that contain genetic changes by disrupting cell cycle proteins and impairing apoptosis (Pantazi et al., 2014). The HH ligand is reported to protect against radiation in the human HCC cell lines, HA22T and Sk-Hep1. Treating HA22T cells with the HH ligand upregulated HH, PTCH1, and GLI1, and caused the nuclear translocation of GLI1, indicating HH signaling activation (Chen et al., 2011). The radioprotective effect of the HH ligand was partially reduced by an anti-HH antibody and eliminated by GLI1 RNA interference, implying that HH signaling has a key role in radioresistance. Furthermore, the HH ligand inhibits the repair of DNA double-strand breaks by suppressing RT-induced phosphorylation of checkpoint kinase 1 (Chen et al., 2011). Previous studies have reported that the components of the HH pathway are elevated during the transformation of normal epithelial cells into squamous cell carcinoma, implicating this pathway in the development and progression of squamous cell carcinomas. Moreover, radiotherapy elevates GLI1 expression at the intersection of the tumor and stroma in head and neck squamous cell carcinoma, which contributes to the development of stroma-mediated resistance. HH pathway inhibition using cyclopamine has been shown to suppress GLI1 activation and to significantly enhance tumor radiosensitivity (Gan et al., 2014). Studies have shown that HH signaling mediates the proliferation and invasion of cervical cancer cells since GLI3 inhibition using small interfering RNAs reduces their survival. Multiple studies have revealed that HH signaling promotes cervical cancer radioresistance, highlighting the inhibition of this pathway as a groundbreaking therapeutic approach. The combination of HH inhibitors with conventional treatment protocols might improve therapeutic outcomes (Liu and Wang, 2019). Another study demonstrated that elevated GLI1 expression in head and neck squamous cell carcinoma is enhanced by RT, leading to therapy resistance (Chaudary et al., 2017; Zhang et al., 2021). By blocking the HH pathway using cyclopamine, we observed a decrease in GLI1 activation and an increase in tumor sensitivity to RT. Additionally, the mTOR/S6K1 pathway was found to mediate radiotherapy-induced GLI1 expression. Another study found that high SRY-box transcription factor 2 (SOX2) expression causes radioresistance in cervical cancer, indicating that SOX2 is closely associated with changes in irradiation-induced survival, proliferation, apoptosis, and cell cycle (Huang et al., 2018). Moreover, the relationship between SOX2 and the HH pathway was confirmed via immunohistochemical staining for SOX2 and GLI1. Another study involving a patient-derived murine xenograft (PDX) model of esophageal adenocarcinoma found that two out of three PDX models exhibited a continuous increase in HH gene expression after radiation. An HH-responsive PDX model revealed that when compared with either treatment alone, the combination of LDE225 and radiation markedly delayed tumor growth (Huang et al., 2018; Teichman et al., 2018). GLI1 expression is significantly upregulated in esophageal cancer cell lines. Notably, GLI1 overexpression in the parental cell line significantly decreased their radiosensitivity, while its knockdown restored radiosensitivity in the radioresistant cell line. These findings suggest that GLI1 plays a vital role in the development of esophageal cancer radioresistance (Huang et al., 2018). In hypoxic conditions, renal cell carcinoma (RCC) cells have HIF2a-mediated, elevated HH–GLI1 activity. Hypoxia-induced GLI1 activation occurs through SMO-independent pathways and can be inhibited by PI3K or MEK inhibitors. Notably, in normoxic conditions, the HH–GLI1 pathway upregulates HIF2a expression. A clear positive correlation has been observed between HIF2a and GLI1 expression in RCC patients (Zhou et al., 2014). The simultaneous use of sh-HIF2a and a GLI1 inhibitor substantially increased the sensitivity of RCC cells to ionizing radiation (IR). Although HH targeting did not sensitize breast cancer cells to any form of radiation, the co-administration of GANT61 with X-rays or carbon ions suppressed MCF-7 cell migration more significantly than either form of radiation alone (Konings et al., 2019a; Konings et al., 2019b). When compared with irradiated MG63 (an osteosarcoma cell line) cells, MG63R cells exhibited a greater rate of surviving colonies, greater cell viability, and lower levels of apoptosis. The expression levels of HH, BCL2, and GLI, were also markedly elevated in MG63R cells (Qu et al., 2017; Qu et al., 2018). Nevertheless, pretreatment with emodin caused a dose-dependent decrease in cell viability and survival colony formation, and increased apoptosis in irradiated MG63R cells. Additionally, pretreatment with emodin suppressed the expression of HH and BCL2, inhibited GLI1 nuclear translocation, and elevated C-caspase-3 expression in irradiated MG63R cells in a dose-dependent manner (Qu et al., 2017).
3.2 CSC and HH/GLI signaling
Exposure to IR is reported to trigger the CSC phenotype in several cancers, including melanoma, breast, lung, and prostate cancers (Lee et al., 2017). IR- or chemotherapy-induced genotoxic stress can trigger a CSC-like phenotype by intensifying ROS production. It is reported that IR can reprogram differentiated cancer cells into CSCs (Lee et al., 2017). In patients with prostate cancer, radiotherapy elevates the population of CD44+ cells, which exhibit CSC characteristics (Tsao et al., 2019). Furthermore, IR induces the re-expression of specific stem cell regulators, such as SOX2, OCT4, Nanog, and KLF4, which promotes stemness in cancer cells. HH signaling is a critical regulator of various cellular processes, including proliferation, motility, adhesion, and cell fate, as well as the maintenance of stem cells, progenitor cells, and self-renewal (Jeng et al., 2020). Some of the genes that are upregulated by HH signaling are associated with stemness, such as Nanog, OCT4, SOX2, and the polycomb complex protein, BMI-1 (also known as polycomb group RING finger protein 4 [PCGF4] or RING finger protein 51 [RNF51]), which implicates HH signaling in the determination of the cancer stem cell phenotype (Tsao et al., 2019). Furthermore, studies have revealed that HH signaling not only contributes to cancer growth and maintenance but also to cancer drug resistance, which promotes a more aggressive phenotype. In medulloblastoma, PTCH germline mutations or silencing through methylation can impair its tumor suppressor effects. This signaling can be counteracted by the PTCH–SMO inhibitor, cyclopamine, resulting in reduced proliferation and increased differentiation (Takebe et al., 2015). In pancreatic cancer, HH and mTOR signaling may be crucial for CSCs self-renewal. However, targeting these pathways with cyclopamine and paramycine alone does not eliminate pancreatic CSCs, and CSC elimination was only observed upon co-treatment with gemcitabine, a standard chemotherapy agent. This suggests that combining targeted therapy and standard chemotherapy may be effective at eliminating CSCs (Chiorean and Coveler, 2015). HH signaling can also be inhibited using forskolin, which activates protein kinase A (PKA). The stimulation of adenylyl cyclase increases cellular cAMP levels (an indicator of cellular energy depletion) and triggers apoptosis. Conversely, some agents, such as the co-conjugate of chondroitin-6-sulfate and dermatan sulfate, can amplify HH signaling activation and increase IHH expression (Lu et al., 2021). In addition, HDAC6 is upregulated in GSCs when compared with non-stem tumor cells. Inhibiting HDAC6 suppressed the expression and activity of GLI1, PTCH1, and PTCH2 (which are components of the HH pathway) in GSCs (Yang et al., 2018). HDAC6 inhibition suppresses cell proliferation while promoting differentiation and apoptosis in GSCs via the inactivation of the HH–GLI1 signaling pathway. Additionally, HDAC6 inhibition suppresses the DNA damage repair capacity of GSCs by degrading CHK1. These effects increase radiosensitivity (Yang et al., 2018).
3.3 HH/GLI signaling inhibition
Based on mounting evidence that HH inhibitors may be effective against cancer, several multicenter clinical trials have recently assessed the efficacy and safety of vismodegib, sonidegib, taladegib, and patidegib (Kim et al., 2010a; Kim et al., 2010b; Ohashi et al., 2012; Jin et al., 2017; Peer et al., 2019). These clinical trials have revealed a promising objective response rate for locally advanced and metastatic BCC, as well as newly diagnosed or relapsed/refractory myeloid malignancies (Kim et al., 2013; Infante et al., 2015). However, these trials did not assess radiosensitivity and more than 20% of the patients discontinued treatment because of adverse events (AE), such as muscle spasms, alopecia, and dysgeusia (Jin et al., 2017; Peer et al., 2019). In clinical settings, increasing radiosensitivity via HH inhibition was more likely to be accompanied by these AEs. Treatment breaks were introduced to manage AEs without response rate reduction (104). Moreover, the AEs associated with HH inhibitors are attributable to the importance of HH signaling in normal cells. Hence, selective HH agonists, which may help relieve AEs, have been developed in mouse models (105).
Resistance to HH inhibitors also limits their clinical translation. SMO mutations are common in patients with resistance to HH inhibition (106). Based on the evidence as summarized in Table 1, SMO and GLI inhibitors have been developed to increase tumor radiosensitivity. Several SMO inhibitors, such as LEQ-506, TAK-441, itraconazole, and taladegib were discovered as SMO antagonists that enhance cancer treatment efficacy (74,107-109). Resistance to SMO inhibitors can be caused by increased GLI activity through molecular interactions, posttranslational modifications, and non-canonical HH signaling (110). Because of the crucial role of GLI transcription factors in facilitating the oncogenic effects of HH signaling, the possibility of therapeutically targeting GLI proteins in HH signaling-driven cancers is promising but challenging. The GLI antagonist, GANT61, inhibits GLI proteins by impairing their DNA binding capability. However, its limited pharmacological potential has discouraged clinical investigations. ATO, a GLI antagonist, blocks GLI protein function (111) and when combined with the SMO inhibitor, itraconazole, it effectively overcomes the resistance to SMO inhibition seen in models of medulloblastoma and BCC (112). Glabrescione B, which is a direct GLI inhibitor, is an isoflavone that is naturally found in the seeds of Derris glabrescens. By binding to the zinc finger domain of GLI1, it inhibits the interaction between GLI1 and DNA (113). Clinical trials involving HH inhibition in combination with other inhibitors, such as PI3K or programmed death-1 (PD-1) inhibitors, have also been conducted to overcome the resistance (114,115). An exploratory study revealed that a significant proportion of patients with SMO mutations might benefit from a combination of immunotherapy that accounts for the mutational burden (116). Another study reported that the combined use of vismodegib and an anti-PD-1 antibody synergistically reduced mouse liver tumors. This effect was achieved through the transformation of M2 tumor-associated macrophages into M1 macrophages and enhanced CD8+ T-cell migration into the TME (117). Thus, combination therapy may be effective at overcoming resistance to HH inhibitors.
4 Future perspectives
Aberrant HH signaling activation, which has been observed in multiple cancers, can influence cancer development. Many studies have shown that HH signaling significantly contributes to the development of radioresistance in various cancers and have highlighted it as a promising therapeutic target. Combination therapy using HH inhibitors (including GLI inhibitors) and conventional therapies may enhance cancer treatment efficacy. However, because the clinical use of HH signaling inhibitors is associated with toxic side effects and drug resistance, further studies are needed to develop more efficient therapeutic approaches.
Author contributions
LZ: Data curation, Funding acquisition, Investigation, Software, Supervision, Writing–original draft. YZ: Methodology, Resources, Visualization, Writing–original draft. KL: Project administration, Supervision, Validation, Writing–review and editing. SX: Conceptualization, Funding acquisition, Methodology, Writing–review and editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was supported by the Natural Science Funding of Jilin Province (YDZJ202201ZYTS109,YDZJ202301ZYTS458); and the Interdiscipline Subject Foundation of the First Hospital of Jilin University (04033990001).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abad, E., Graifer, D., and Lyakhovich, A. (2020). DNA damage response and resistance of cancer stem cells. Cancer Lett. 474, 106–117. doi:10.1016/j.canlet.2020.01.008
Alexandrou, A. T., and Li, J. J. (2014). Cell cycle regulators guide mitochondrial activity in radiation-induced adaptive response. Antioxidants redox Signal. 20, 1463–1480. doi:10.1089/ars.2013.5684
Babaei, G., Aziz, S. G., and Jaghi, N. Z. Z. (2021). EMT, cancer stem cells and autophagy; the three main axes of metastasis. Biomed. Pharmacother. = Biomedecine Pharmacother. 133, 110909. doi:10.1016/j.biopha.2020.110909
Bakshi, A., Chaudhary, S. C., Rana, M., Elmets, C. A., and Athar, M. (2017). Basal cell carcinoma pathogenesis and therapy involving hedgehog signaling and beyond. Mol. Carcinog. 56, 2543–2557. doi:10.1002/mc.22690
Barker, H. E., Paget, J. T., Khan, A. A., and Harrington, K. J. (2015). The tumour microenvironment after radiotherapy: mechanisms of resistance and recurrence. Nat. Rev. Cancer 15, 409–425. doi:10.1038/nrc3958
Biswas, P., Dey, D., Biswas, P. K., Rahaman, T. I., Saha, S., Parvez, A., et al. (2022). A comprehensive analysis and anti-cancer activities of quercetin in ROS-mediated cancer and cancer stem cells. Int. J. Mol. Sci. 23, 11746. doi:10.3390/ijms231911746
Briscoe, J., and Thérond, P. P. (2013). The mechanisms of Hedgehog signalling and its roles in development and disease. Nat. Rev. Mol. Cell. Biol. 14, 416–429. doi:10.1038/nrm3598
Caglar, H. O., and Biray Avci, C. (2020). Alterations of cell cycle genes in cancer: unmasking the role of cancer stem cells. Mol. Biol. Rep. 47, 3065–3076. doi:10.1007/s11033-020-05341-6
Caja, L., Dituri, F., Mancarella, S., Caballero-Diaz, D., Moustakas, A., Giannelli, G., et al. (2018). TGF-Β and the tissue microenvironment: relevance in fibrosis and cancer. Int. J. Mol. Sci. 19, 1294. doi:10.3390/ijms19051294
Canettieri, G., Di Marcotullio, L., Greco, A., Coni, S., Antonucci, L., Infante, P., et al. (2010). Histone deacetylase and Cullin3-REN(KCTD11) ubiquitin ligase interplay regulates Hedgehog signalling through Gli acetylation. Nat. Cell. Biol. 12, 132–142. doi:10.1038/ncb2013
Chaudary, N., Pintilie, M., Hedley, D., Hill, R. P., Milosevic, M., and Mackay, H. (2017). Hedgehog inhibition enhances efficacy of radiation and cisplatin in orthotopic cervical cancer xenografts. Br. J. cancer 116, 50–57. doi:10.1038/bjc.2016.383
Chen, X., Fu, E., Lou, H., Mao, X., Yan, B., Tong, F., et al. (2019). IL-6 induced M1 type macrophage polarization increases radiosensitivity in HPV positive head and neck cancer. Cancer Lett. 456, 69–79. doi:10.1016/j.canlet.2019.04.032
Chen, Y. J., Lin, C. P., Hsu, M. L., Shieh, H. R., Chao, N. K., and Chao, K. S. (2011). Sonic hedgehog signaling protects human hepatocellular carcinoma cells against ionizing radiation in an autocrine manner. Int. J. Radiat. Oncol. Biol. Phys. 80, 851–859. doi:10.1016/j.ijrobp.2011.01.003
Chiorean, E. G., and Coveler, A. L. (2015). Pancreatic cancer: optimizing treatment options, new, and emerging targeted therapies. Drug Des. Dev. Ther. 9, 3529–3545. doi:10.2147/dddt.S60328
Citrin, D. E. (2017). Recent developments in radiotherapy. N. Engl. J. Med. 377, 1065–1075. doi:10.1056/NEJMra1608986
Clark, V. E., Erson-Omay, E. Z., Serin, A., Yin, J., Cotney, J., Ozduman, K., et al. (2013). Genomic analysis of non-NF2 meningiomas reveals mutations in TRAF7, KLF4, AKT1, and SMO. Sci. (New York, N.Y.) 339, 1077–1080. doi:10.1126/science.1233009
Dewhirst, M. W., and Chi, J. T. (2013). Understanding the tumor microenvironment and radioresistance by combining functional imaging with global gene expression. Seminars Radiat. Oncol. 23, 296–305. doi:10.1016/j.semradonc.2013.05.004
Digomann, D., Linge, A., and Dubrovska, A. (2019). SLC3A2/CD98hc, autophagy and tumor radioresistance: a link confirmed. Autophagy 15, 1850–1851. doi:10.1080/15548627.2019.1639302
Efstathiou, E., Karlou, M., Wen, S., Hoang, A., Pettaway, C. A., Pisters, L. L., et al. (2013). Integrated Hedgehog signaling is induced following castration in human and murine prostate cancers. Prostate 73, 153–161. doi:10.1002/pros.22550
Fattahi, S., Pilehchian Langroudi, M., and Akhavan-Niaki, H. (2018). Hedgehog signaling pathway: epigenetic regulation and role in disease and cancer development. J. Cell. physiology 233, 5726–5735. doi:10.1002/jcp.26506
Fischer, K., Hoffmann, P., Voelkl, S., Meidenbauer, N., Ammer, J., Edinger, M., et al. (2007). Inhibitory effect of tumor cell-derived lactic acid on human T cells. Blood 109, 3812–3819. doi:10.1182/blood-2006-07-035972
Gan, G. N., Eagles, J., Keysar, S. B., Wang, G., Glogowska, M. J., Altunbas, C., et al. (2014). Hedgehog signaling drives radioresistance and stroma-driven tumor repopulation in head and neck squamous cancers. Cancer Res. 74, 7024–7036. doi:10.1158/0008-5472.Can-14-1346
Gong, L., Zhang, Y., Liu, C., Zhang, M., and Han, S. (2021). Application of radiosensitizers in cancer radiotherapy. Int. J. nanomedicine 16, 1083–1102. doi:10.2147/ijn.S290438
Hoque, S., Dhar, R., Kar, R., Mukherjee, S., Mukherjee, D., Mukerjee, N., et al. (2023). Cancer stem cells (CSCs): key player of radiotherapy resistance and its clinical significance. Biomarkers Biochem. Indic. Expo. response, susceptibility Chem. 28, 139–151. doi:10.1080/1354750x.2022.2157875
Huang, C., Lu, H., Li, J., Xie, X., Fan, L., Wang, D., et al. (2018). SOX2 regulates radioresistance in cervical cancer via the hedgehog signaling pathway. Gynecol. Oncol. 151, 533–541. doi:10.1016/j.ygyno.2018.10.005
Huang, Y., Nahar, S., Nakagawa, A., Fernandez-Barrena, M. G., Mertz, J. A., Bryant, B. M., et al. (2016). Regulation of GLI underlies a role for BET bromodomains in pancreatic cancer growth and the tumor microenvironment. Clin. cancer Res. 22, 4259–4270. doi:10.1158/1078-0432.Ccr-15-2068
Huang, Y. H., Yang, P. M., Chuah, Q. Y., Lee, Y. J., Hsieh, Y. F., Peng, C. W., et al. (2014). Autophagy promotes radiation-induced senescence but inhibits bystander effects in human breast cancer cells. Autophagy 10, 1212–1228. doi:10.4161/auto.28772
Infante, P., Mori, M., Alfonsi, R., Ghirga, F., Aiello, F., Toscano, S., et al. (2015). Gli1/DNA interaction is a druggable target for Hedgehog-dependent tumors. EMBO J. 34, 200–217. doi:10.15252/embj.201489213
Ishikawa, K., Ishii, H., and Saito, T. (2006). DNA damage-dependent cell cycle checkpoints and genomic stability. DNA Cell. Biol. 25, 406–411. doi:10.1089/dna.2006.25.406
Jarosz-Biej, M., Smolarczyk, R., Cichoń, T., and Kułach, N. (2019). Tumor microenvironment as A "game changer" in cancer radiotherapy. Int. J. Mol. Sci. 20, 3212. doi:10.3390/ijms20133212
Jeng, K. S., Chang, C. F., and Lin, S. S. (2020). Sonic hedgehog signaling in organogenesis, tumors, and tumor microenvironments. Int. J. Mol. Sci. 21, 758. doi:10.3390/ijms21030758
Jiang, J. (2022). Hedgehog signaling mechanism and role in cancer. Semin. Cancer Biol. 85, 107–122. doi:10.1016/j.semcancer.2021.04.003
Jiang, J., and Hui, C. C. (2008). Hedgehog signaling in development and cancer. Dev. Cell. 15, 801–812. doi:10.1016/j.devcel.2008.11.010
Jiao, Y., Cao, F., and Liu, H. (2022). Radiation-induced cell death and its mechanisms. Health Phys. 123, 376–386. doi:10.1097/hp.0000000000001601
Jin, G., Sivaraman, A., and Lee, K. (2017). Development of taladegib as a sonic hedgehog signaling pathway inhibitor. Archives pharmacal Res. 40, 1390–1393. doi:10.1007/s12272-017-0987-x
Khosravi, N., Mokhtarzadeh, A., Baghbanzadeh, A., Hajiasgharzadeh, K., Shahgoli, V. K., Hemmat, N., et al. (2020). Immune checkpoints in tumor microenvironment and their relevance to the development of cancer stem cells. Life Sci. 256, 118005. doi:10.1016/j.lfs.2020.118005
Kim, J., Aftab, B. T., Tang, J. Y., Kim, D., Lee, A. H., Rezaee, M., et al. (2013). Itraconazole and arsenic trioxide inhibit Hedgehog pathway activation and tumor growth associated with acquired resistance to smoothened antagonists. Cancer Cell. 23, 23–34. doi:10.1016/j.ccr.2012.11.017
Kim, J., Lee, J. J., Kim, J., Gardner, D., and Beachy, P. A. (2010b). Arsenic antagonizes the Hedgehog pathway by preventing ciliary accumulation and reducing stability of the Gli2 transcriptional effector. Proc. Natl. Acad. Sci. U. S. A. 107, 13432–13437. doi:10.1073/pnas.1006822107
Kim, J., Tang, J. Y., Gong, R., Kim, J., Lee, J. J., Clemons, K. V., et al. (2010a). Itraconazole, a commonly used antifungal that inhibits Hedgehog pathway activity and cancer growth. Cancer Cell. 17, 388–399. doi:10.1016/j.ccr.2010.02.027
Kim, J. K., Jeon, H. Y., and Kim, H. (2015). The molecular mechanisms underlying the therapeutic resistance of cancer stem cells. Archives pharmacal Res. 38, 389–401. doi:10.1007/s12272-014-0531-1
Koch, U., Krause, M., and Baumann, M. (2010). Cancer stem cells at the crossroads of current cancer therapy failures--radiation oncology perspective. Semin. Cancer Biol. 20, 116–124. doi:10.1016/j.semcancer.2010.02.003
Konings, K., Belmans, N., Vermeesen, R., Baselet, B., Lamers, G., Janssen, A., et al. (2019a). Targeting the Hedgehog pathway in combination with X-ray or carbon ion radiation decreases migration of MCF-7 breast cancer cells. Int. J. Oncol. 55, 1339–1348. doi:10.3892/ijo.2019.4901
Konings, K., Vandevoorde, C., Belmans, N., Vermeesen, R., Baselet, B., Walleghem, M. V., et al. (2019b). The combination of particle irradiation with the hedgehog inhibitor GANT61 differently modulates the radiosensitivity and migration of cancer cells compared to X-ray irradiation. Front. Oncol. 9, 391. doi:10.3389/fonc.2019.00391
Krause, M., Dubrovska, A., Linge, A., and Baumann, M. (2017). Cancer stem cells: radioresistance, prediction of radiotherapy outcome and specific targets for combined treatments. Adv. drug Deliv. Rev. 109, 63–73. doi:10.1016/j.addr.2016.02.002
Kuwahara, Y., Tomita, K., Urushihara, Y., Sato, T., Kurimasa, A., and Fukumoto, M. (2018). Association between radiation-induced cell death and clinically relevant radioresistance. Histochem. Cell. Biol. 150, 649–659. doi:10.1007/s00418-018-1728-z
Lauth, M., and Toftgård, R. (2007). Non-canonical activation of GLI transcription factors: implications for targeted anti-cancer therapy. Cell. cycleGeorget. Tex.) 6, 2458–2463. doi:10.4161/cc.6.20.4808
Lee, S. Y., Jeong, E. K., Ju, M. K., Jeon, H. M., Kim, M. Y., Kim, C. H., et al. (2017). Induction of metastasis, cancer stem cell phenotype, and oncogenic metabolism in cancer cells by ionizing radiation. Mol. cancer 16, 10. doi:10.1186/s12943-016-0577-4
Li, C. Y. (2023). Non-canonical roles of apoptotic and DNA double-strand break repair factors in mediating cellular response to ionizing radiation. Int. J. Radiat. Biol. 99, 915–924. doi:10.1080/09553002.2021.1948139
Li, X., Chen, F., Zhu, Q., Ding, B., Zhong, Q., Huang, K., et al. (2016). Gli-1/PI3K/AKT/NF-kB pathway mediates resistance to radiation and is a target for reversion of responses in refractory acute myeloid leukemia cells. Oncotarget 7, 33004–33015. doi:10.18632/oncotarget.8844
Liem, K. F., He, M., Ocbina, P. J., and Anderson, K. V. (2009). Mouse Kif7/Costal2 is a cilia-associated protein that regulates Sonic hedgehog signaling. Proc. Natl. Acad. Sci. U. S. A. 106, 13377–13382. doi:10.1073/pnas.0906944106
Liu, C., and Wang, R. (2019). The roles of hedgehog signaling pathway in radioresistance of cervical cancer. Dose-Response 17, 1559325819885293. doi:10.1177/1559325819885293
Logan, C. M., Giordano, A., Puca, A., and Cassone, M. (2007). Prostaglandin E2: at the crossroads between stem cell development, inflammation and cancer. Cancer Biol. Ther. 6, 1517–1520. doi:10.4161/cbt.6.10.4750
Long, J., Li, B., Rodriguez-Blanco, J., Pastori, C., Volmar, C. H., Wahlestedt, C., et al. (2014). The BET bromodomain inhibitor I-BET151 acts downstream of smoothened protein to abrogate the growth of hedgehog protein-driven cancers. J. Biol. Chem. 289, 35494–35502. doi:10.1074/jbc.M114.595348
Lu, Y., Zhu, Y., Deng, S., Chen, Y., Li, W., Sun, J., et al. (2021). Targeting the sonic hedgehog pathway to suppress the expression of the cancer stem cell (CSC)-Related transcription factors and CSC-driven thyroid tumor growth. Cancers (Basel) 13, 418. doi:10.3390/cancers13030418
Matise, M. P., and Joyner, A. L. (1999). Gli genes in development and cancer. Oncogene 18, 7852–7859. doi:10.1038/sj.onc.1203243
Matissek, S. J., and Elsawa, S. F. (2020). GLI3: a mediator of genetic diseases, development and cancer. Cell. Commun. Signal. 18, 54. doi:10.1186/s12964-020-00540-x
Meng, Y., Beckett, M. A., Liang, H., Mauceri, H. J., van Rooijen, N., Cohen, K. S., et al. (2010). Blockade of tumor necrosis factor alpha signaling in tumor-associated macrophages as a radiosensitizing strategy. Cancer Res. 70, 1534–1543. doi:10.1158/0008-5472.Can-09-2995
Niewiadomski, P., Niedziółka, S. M., Markiewicz, Ł., Uśpieński, T., Baran, B., and Chojnowska, K. (2019). Gli proteins: regulation in development and cancer. Cells 8, 8. doi:10.3390/cells8020147
Nikjoo, H., Emfietzoglou, D., Liamsuwan, T., Taleei, R., Liljequist, D., and Uehara, S. (2016). Radiation track, DNA damage and response-a review. Rep. Prog. Phys. Phys. Soc. (Great Britain) 79, 116601. doi:10.1088/0034-4885/79/11/116601
Ogawa, K., Yoshioka, Y., Isohashi, F., Seo, Y., Yoshida, K., and Yamazaki, H. (2013). Radiotherapy targeting cancer stem cells: current views and future perspectives. Anticancer Res. 33, 747–754.
Ohashi, T., Oguro, Y., Tanaka, T., Shiokawa, Z., Tanaka, Y., Shibata, S., et al. (2012). Discovery of the investigational drug TAK-441, a pyrrolo[3,2-c]pyridine derivative, as a highly potent and orally active hedgehog signaling inhibitor: modification of the core skeleton for improved solubility. Bioorg. Med. Chem. 20, 5507–5517. doi:10.1016/j.bmc.2012.07.034
Olivares-Urbano, M. A., Griñán-Lisón, C., Marchal, J. A., Núñez, M. I., and Radioresistance, C. S. C. (2020). CSC radioresistance: a therapeutic challenge to improve radiotherapy effectiveness in cancer. Cells 9, 1651. doi:10.3390/cells9071651
Pal, K., Hwang, S. H., Somatilaka, B., Badgandi, H., Jackson, P. K., DeFea, K., et al. (2016). Smoothened determines β-arrestin-mediated removal of the G protein-coupled receptor Gpr161 from the primary cilium. J. Cell. Biol. 212, 861–875. doi:10.1083/jcb.201506132
Palle, K., Mani, C., Tripathi, K., and Athar, M. (2015). Aberrant GLI1 activation in DNA damage response, carcinogenesis and chemoresistance. Cancers (Basel) 7, 2330–2351. doi:10.3390/cancers7040894
Pantazi, E., Gemenetzidis, E., Trigiante, G., Warnes, G., Shan, L., Mao, X., et al. (2014). GLI2 induces genomic instability in human keratinocytes by inhibiting apoptosis. Cell. death Dis. 5, e1028. doi:10.1038/cddis.2013.535
Patel, N. H., Bloukh, S., Alwohosh, E., Alhesa, A., Saleh, T., and Gewirtz, D. A. (2021). Autophagy and senescence in cancer therapy. Adv. cancer Res. 150, 1–74. doi:10.1016/bs.acr.2021.01.002
Patel, N. H., Sohal, S. S., Manjili, M. H., Harrell, J. C., and Gewirtz, D. A. (2020). The roles of autophagy and senescence in the tumor cell response to radiation. Radiat. Res. 194, 103–115. doi:10.1667/rade-20-00009
Pawlik, T. M., and Keyomarsi, K. (2004). Role of cell cycle in mediating sensitivity to radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 59, 928–942. doi:10.1016/j.ijrobp.2004.03.005
Peer, E., Tesanovic, S., and Aberger, F. (2019). Next-generation hedgehog/GLI pathway inhibitors for cancer therapy. Cancers (Basel) 11, 538. doi:10.3390/cancers11040538
Qu, W., Li, D., Wang, Y., Wu, Q., and Hao, D. (2018). Activation of sonic hedgehog signaling is associated with human osteosarcoma cells radioresistance characterized by increased proliferation, migration, and invasion. Med. Sci. Monit. 24, 3764–3771. doi:10.12659/msm.908278
Qu, W., Wang, Y., Wu, Q., Hao, D., and Li, D. (2017). Emodin impairs radioresistance of human osteosarcoma cells by suppressing sonic hedgehog signaling. Med. Sci. Monit. 23, 5767–5773. doi:10.12659/msm.907453
Ruiz i Altaba, A. (1999). Gli proteins and Hedgehog signaling: development and cancer. Trends Genet. 15, 418–425. doi:10.1016/s0168-9525(99)01840-5
Ruiz i Altaba, A., Sánchez, P., and Dahmane, N. (2002). Gli and hedgehog in cancer: tumours, embryos and stem cells. Nat. Rev. Cancer 2, 361–372. doi:10.1038/nrc796
Rycaj, K., and Tang, D. G. (2014). Cancer stem cells and radioresistance. Int. J. Radiat. Biol. 90, 615–621. doi:10.3109/09553002.2014.892227
Sannigrahi, M. K., Singh, V., Sharma, R., Panda, N. K., and Khullar, M. (2015). Role of autophagy in head and neck cancer and therapeutic resistance. Oral Dis. 21, 283–291. doi:10.1111/odi.12254
Santivasi, W. L., and Xia, F. (2014). Ionizing radiation-induced DNA damage, response, and repair. Antioxidants redox Signal. 21, 251–259. doi:10.1089/ars.2013.5668
Schaue, D., and McBride, W. H. (2015). Opportunities and challenges of radiotherapy for treating cancer. Nat. Rev. Clin. Oncol. 12, 527–540. doi:10.1038/nrclinonc.2015.120
Schulz, A., Meyer, F., Dubrovska, A., and Borgmann, K. (2019). Cancer stem cells and radioresistance: DNA repair and beyond. Cancers (Basel) 11, 862. doi:10.3390/cancers11060862
Shibata, A., and Jeggo, P. A. (2020). Roles for 53BP1 in the repair of radiation-induced DNA double strand breaks. DNA repair 93, 102915. doi:10.1016/j.dnarep.2020.102915
Sinha, R. P., and Häder, D. P. (2002). UV-induced DNA damage and repair: a review. Photochem. photobiological Sci. 1, 225–236. doi:10.1039/b201230h
Skoda, A. M., Simovic, D., Karin, V., Kardum, V., Vranic, S., and Serman, L. (2018). The role of the Hedgehog signaling pathway in cancer: a comprehensive review. Bosnian J. basic Med. Sci. 18, 8–20. doi:10.17305/bjbms.2018.2756
Srinivas, U. S., Tan, B. W. Q., Vellayappan, B. A., and Jeyasekharan, A. D. (2019). ROS and the DNA damage response in cancer. Redox Biol. 25, 101084. doi:10.1016/j.redox.2018.101084
Steele, N. G., Biffi, G., Kemp, S. B., Zhang, Y., Drouillard, D., Syu, L., et al. (2021). Inhibition of hedgehog signaling alters fibroblast composition in pancreatic cancer. Clin. cancer Res. 27, 2023–2037. doi:10.1158/1078-0432.Ccr-20-3715
Suchors, C., and Kim, J. (2022). Canonical hedgehog pathway and noncanonical GLI transcription factor activation in cancer. Cells 11, 2523. doi:10.3390/cells11162523
Suwa, T., Kobayashi, M., Nam, J. M., and Harada, H. (2021). Tumor microenvironment and radioresistance. Exp. Mol. Med. 53, 1029–1035. doi:10.1038/s12276-021-00640-9
Taeb, S., Ashrafizadeh, M., Zarrabi, A., Rezapoor, S., Musa, A. E., Farhood, B., et al. (2022). Role of tumor microenvironment in cancer stem cells resistance to radiotherapy. Curr. cancer drug targets 22, 18–30. doi:10.2174/1568009622666211224154952
Takebe, N., Miele, L., Harris, P. J., Jeong, W., Bando, H., Kahn, M., et al. (2015). Targeting Notch, Hedgehog, and Wnt pathways in cancer stem cells: clinical update. Nat. Rev. Clin. Oncol. 12, 445–464. doi:10.1038/nrclinonc.2015.61
Taylor, M. A., Das, B. C., and Ray, S. K. (2018). Targeting autophagy for combating chemoresistance and radioresistance in glioblastoma. Apoptosis Int. J. Program. Cell. death 23, 563–575. doi:10.1007/s10495-018-1480-9
Teichman, J., Dodbiba, L., Thai, H., Fleet, A., Morey, T., Liu, L., et al. (2018). Hedgehog inhibition mediates radiation sensitivity in mouse xenograft models of human esophageal adenocarcinoma. PloS one 13, e0194809. doi:10.1371/journal.pone.0194809
Tsao, T., Beretov, J., Ni, J., Bai, X., Bucci, J., Graham, P., et al. (2019). Cancer stem cells in prostate cancer radioresistance. Cancer Lett. 465, 94–104. doi:10.1016/j.canlet.2019.08.020
Vattai, A., Kremer, N., Meister, S., Beyer, S., Keilmann, L., Buschmann, C., et al. (2023). Increase of the T-reg-recruiting chemokine CCL22 expression in a progressive course of cervical dysplasia. J. cancer Res. Clin. Oncol. 149, 6613–6623. doi:10.1007/s00432-023-04638-w
Wang, Z., Tang, Y., Tan, Y., Wei, Q., and Yu, W. (2019). Cancer-associated fibroblasts in radiotherapy: challenges and new opportunities. Cell. Commun. Signal. 17, 47. doi:10.1186/s12964-019-0362-2
Wilson, G. D. (2004). Radiation and the cell cycle, revisited. Cancer metastasis Rev. 23, 209–225. doi:10.1023/B:CANC.0000031762.91306.b4
Wu, T., and Dai, Y. (2017). Tumor microenvironment and therapeutic response. Cancer Lett. 387, 61–68. doi:10.1016/j.canlet.2016.01.043
Xiao, Y., and Yu, D. (2021). Tumor microenvironment as a therapeutic target in cancer. Pharmacol. Ther. 221, 107753. doi:10.1016/j.pharmthera.2020.107753
Xu, L., Xiong, H., Shi, W., Zhou, F., Zhang, M., Hu, G., et al. (2019). Differential expression of sonic hedgehog in lung adenocarcinoma and lung squamous cell carcinoma. Neoplasma 66, 839–846. doi:10.4149/neo_2018_181228N1002
Yang, W., Liu, Y., Gao, R., Yu, H., and Sun, T. (2018). HDAC6 inhibition induces glioma stem cells differentiation and enhances cellular radiation sensitivity through the SHH/Gli1 signaling pathway. Cancer Lett. 415, 164–176. doi:10.1016/j.canlet.2017.12.005
Yang, Y., Yang, Y., Yang, X., Zhu, H., Guo, Q., Chen, X., et al. (2015). Autophagy and its function in radiosensitivity. Tumour Biol. 36, 4079–4087. doi:10.1007/s13277-015-3496-x
Zhang, H., Si, J., Yue, J., and Ma, S. (2021). The mechanisms and reversal strategies of tumor radioresistance in esophageal squamous cell carcinoma. J. cancer Res. Clin. Oncol. 147, 1275–1286. doi:10.1007/s00432-020-03493-3
Zhao, H., Chen, W., Zhu, Y., and Lou, J. (2021). Hypoxia promotes pancreatic cancer cell migration, invasion, and epithelial-mesenchymal transition via modulating the FOXO3a/DUSP6/ERK axis. J. Gastrointest. Oncol. 12, 1691–1703. doi:10.21037/jgo-21-359
Zhou, J., Wu, K., Gao, D., Zhu, G., Wu, D., Wang, X., et al. (2014). Reciprocal regulation of hypoxia-inducible factor 2α and GLI1 expression associated with the radioresistance of renal cell carcinoma. Int. J. Radiat. Oncol. Biol. Phys. 90, 942–951. doi:10.1016/j.ijrobp.2014.06.065
Keywords: GLI, cancer, radiotherapy, radioresistance, cancer stem cell, tumor microenvironment
Citation: Zhang L, Zhang Y, Li K and Xue S (2023) Hedgehog signaling and the glioma-associated oncogene in cancer radioresistance. Front. Cell Dev. Biol. 11:1257173. doi: 10.3389/fcell.2023.1257173
Received: 12 July 2023; Accepted: 31 October 2023;
Published: 13 November 2023.
Edited by:
Nan-Shan Chang, China Medical University, TaiwanReviewed by:
Lucia Longhitano, University of Catania, ItalyMaja Sabol, Ruđer Bošković Institute, Croatia
Copyright © 2023 Zhang, Zhang, Li and Xue. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shuai Xue, xueshuai@jlu.edu.cn