Biosurfactants in Plant Protection Against Diseases: Rhamnolipids and Lipopeptides Case Study
 Jérôme Crouzet1†
Jérôme Crouzet1†  Anthony Arguelles-Arias2†
Anthony Arguelles-Arias2†  Sandrine Dhondt-Cordelier1†
Sandrine Dhondt-Cordelier1†  Sylvain Cordelier1†
Sylvain Cordelier1†  Jelena Pršić2 Gregory Hoff2 Florence Mazeyrat-Gourbeyre1
Jelena Pršić2 Gregory Hoff2 Florence Mazeyrat-Gourbeyre1  Fabienne Baillieul1
Fabienne Baillieul1  Christophe Clément1
Christophe Clément1  Marc Ongena2†
Marc Ongena2†  Stéphan Dorey1*†
Stéphan Dorey1*†- 1Unité RIBP EA 4707, SFR Condorcet FR CNRS 3417, University of Reims Champagne-Ardenne, Reims, France
- 2MiPI laboratory, Gembloux Agro-Bio Tech, SFR Condorcet FR CNRS 3417, University of LieÌge, Gembloux, Belgium
Biosurfactants are amphiphilic surface-active molecules that are produced by a variety of microorganisms including fungi and bacteria. Pseudomonas, Burkholderia, and Bacillus species are known to secrete rhamnolipids and lipopeptides that are used in a wide range of industrial applications. Recently, these compounds have been studied in a context of plant-microbe interactions. This mini-review describes the direct antimicrobial activities of these compounds against plant pathogens. We also provide the current knowledge on how rhamnolipids and lipopeptides stimulate the plant immune system leading to plant resistance to phytopathogens. Given their low toxicity, high biodegradability and ecological acceptance, we discuss the possible role of these biosurfactants as alternative strategies to reduce or even replace pesticide use in agriculture.
Introduction
Plant pathogens cause significant agricultural damages ranging from 10 to 40% depending on the crops before or after harvest, resulting in huge economic losses (Savary et al., 2019). Diseases and pests are therefore major problems for sustainable agriculture in the world. Chemical pesticides are largely used to control plant disease worldwide (Popp et al., 2013). However, chemical pesticides can be detrimental to human and environmental health and therefore, development and optimization of alternative strategies to reduce their utilization for crop protection is becoming a necessity. Biocontrol is a promising strategy based on the use of organisms that decrease disease pressure by competing with the pathogen for space and nutrients, by inducing the plant’s natural defense system, and/or by the production of antimicrobial substances (Berg et al., 2017; Bonanomi et al., 2018; Syed Ab Rahman et al., 2018). In addition, natural, ecofriendly and biodegradable compounds isolated from (micro)organisms can also be part of a biocontrol strategy. These compounds can act directly onto the pathogen via antimicrobial properties or by stimulating the plant immune system to prime plant protection against disease (Delaunois et al., 2014; Bardin et al., 2015; Keswani et al., 2019). Molecules from microbial origin stimulating the plant immune system are called invasion patterns or elicitors and are highly diverse both in nature and origins (Vatsa et al., 2010; Delaunois et al., 2014; Burketova et al., 2015; Schellenberger et al., 2019).
The main classes of microbial biosurfactants are represented by mannosylerythritol lipids (MEL), trehalose dimycolate (TDM), trehalolipids, sophorolipids, rhamnolipids and lipopeptides. They are used in detergent and cleaning solutions and display a very wide range of applications such as food industry, soil and water remediation, microbial enhanced oil recovery, biomedical science, cosmetic industry, nanotechnology and agriculture (Naughton et al., 2019; Singh et al., 2019). They have been studied since decades in biomedical sciences, especially for their antimicrobial properties and as modulators of human innate immunity [for extensive reviews see (Singh and Cameotra, 2004; Banat et al., 2010; Vatsa et al., 2010; Naughton et al., 2019; Coelho et al., 2020; Jahan et al., 2020; Sajid et al., 2020)]. Some of these microbial biosurfactants exhibit antimicrobial properties that are effective against a large panel of plant pathogens (Mnif and Ghribi, 2016; Penha et al., 2020). In addition, rhamnolipids and lipopeptides have been shown to stimulate the plant immune system conferring a better resistance to fungal and bacterial pathogens (D’aes et al., 2010; Raaijmakers et al., 2010; Vatsa et al., 2010; Schellenberger et al., 2019). In this review, we will provide current knowledge and recent advances on the role of biosurfactants in plant protection. We will focus on rhamnolipids and lipopeptides since these biosurfactants are the most studied for agricultural applications and are among the most effective and economically profitable for industrial production.
Biosurfactants as Biopesticides
The main interest in the use of biosurfactants as biopesticides in disease management is their environmental-friendly characteristics, high biodegradability and production from renewal energy resources. Rhamnolipids and lipopeptides have been extensively studied in the context of crop protection. Conversely, other biosurfactants such as sophorolipids, MEL and cellobiose lipids have marginally been investigated for their antimicrobial properties toward plant pathogens (Yoshida et al., 2015; Mnif and Ghribi, 2016; Sen et al., 2017; Chen et al., 2020; Penha et al., 2020). Because rhamnolipids and lipopeptides display a good balance between industrial production, efficacy and preservation of the environment, they are very interesting candidates in biocontrol strategies.
Rhamnolipids Are Efficient Bio-Fungicides
Rhamnolipids are glycolipids produced by various bacterial species including some Pseudomonas sp. and Burkholderia sp. (Abdel-Mawgoud et al., 2010). Whereas no direct or robust evidences have been reported for rhamnolipid antibacterial or antiviral activities against plant pathogens, a large number of studies described their antifungal activities on pathogens affecting crops. These activities were mainly targeted to fungi and oomycetes including Botrytis sp., Rhizoctonia sp., Fusarium sp., Alternaria sp., Pythium sp., Phytophthora sp., or Plasmopara sp. species (Table 1). In these different studies, rhamnolipids, mainly originating from P. aeruginosa, were applied either as a mixture or purified molecules. Among all the congeners that are present in the mixtures, purified mono-rhamnolipids (Rha-C10-C10) and di-rhamnolipids (Rha-Rha-C10-C10) generally displayed the strongest activity. Moreover, all these studies demonstrated a canonical antimicrobial effect such as zoospore lysis, spore germination abortion and mycelial growth inhibition (Table 1). Because of their amphiphilic nature, glycolipids should be able to interact directly with plasma membranes (Otzen, 2017). It was thus proposed that the mode of action of rhamnolipids against zoospore-producing plant pathogens could be a direct lysis of zoospores via the intercalation of the glycolipids within plasma membranes which are not protected by a cell wall (Stanghellini and Miller, 1997). Rhamnolipids could also affect mycelial cells resulting in their destabilization or lysis. Rhamnolipid partition into membranes strongly depends on lipid composition (Aranda et al., 2007). It was shown that purified mono and di-rhamnolipids are able to intercalate into phosphatidylcholine and phosphatidylethanolamine bilayers, notably altering their packing (Ortiz et al., 2006; Sánchez et al., 2006, 2009; Abbasi et al., 2012, 2013). These insertions thus produce structural perturbations, which might affect the function of the membranes. These compounds also alter the physicochemical properties of the bilayer and disturb the hydration status of the water/lipid interface. Depending on the lipid composition of the membrane and on their concentration, rhamnolipids are also able to permeabilize membranes (Sánchez et al., 2010) that could result in their lysis. Amphiphilic compounds such as glycolipids form aggregates in solution depending on their concentration. Above their critical micelle concentration (CMC), they will be present in both forms of aggregates and monomers. Although glycolipid biosurfactants are described to be stable over extreme conditions of pH and temperature (Mnif and Ghribi, 2016), these external factors could also influence rhamnolipid CMC, and therefore their aggregation into micelles (Zhong et al., 2015). The change in CMC could in turn affect their efficiency as biopesticide. Depending on their conformation, the glycolipids could potentially reach more or less easily the pathogen membrane (Aslam et al., 2009) to provoke its disruption (Table 1).
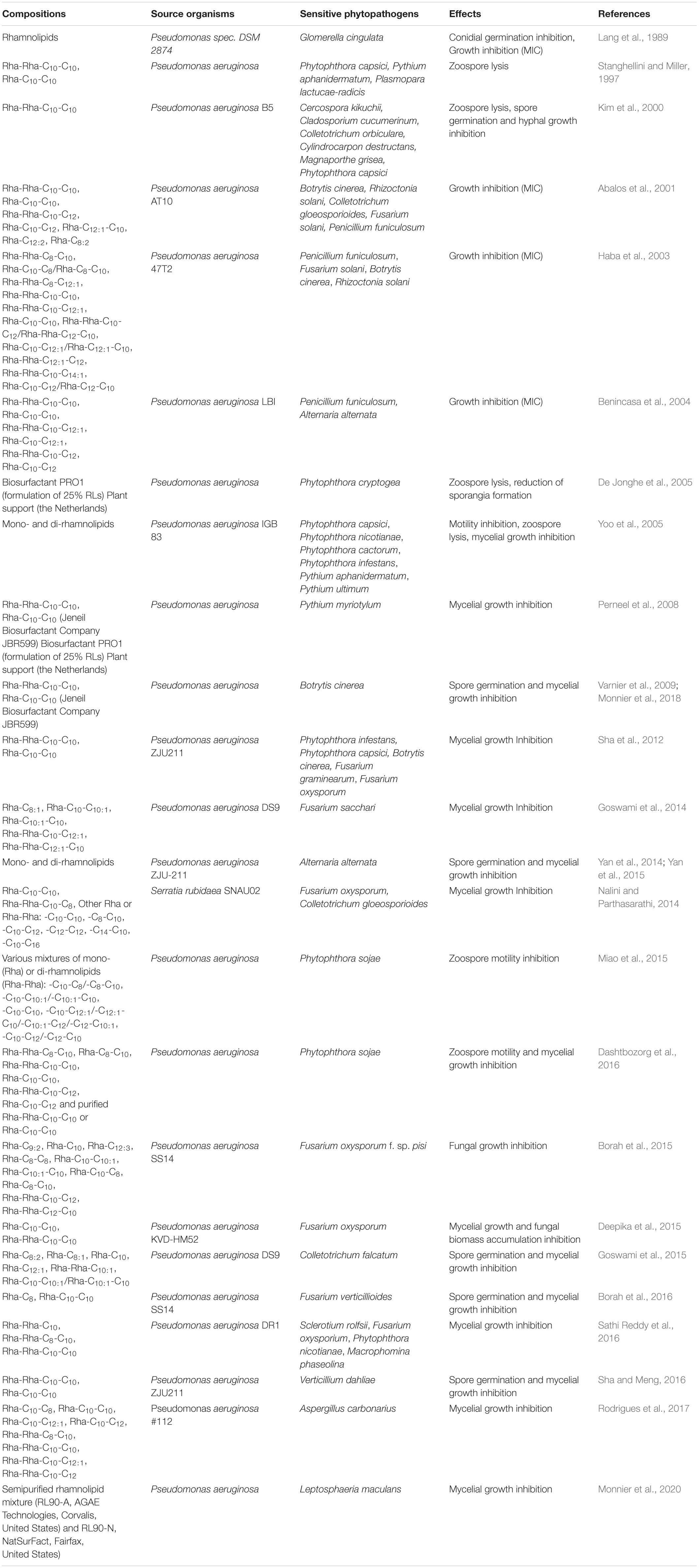
Table 1. Anti-phytopathogenic properties of rhamnolipids.
Lipopeptides as Antimicrobial Agents
Lipopeptide biosurfactants (LPs) are composed of a lipid tail linked to a short linear or cyclic oligopeptide. They are produced by fungi and various bacterial genera mainly in the cyclized form and have received considerable attention for their antimicrobial, cytotoxic, antitumor, immunosuppressant and surfactant properties (Raaijmakers et al., 2010). Cyclic lipopeptides (CLPs) represent a class of biosurfactant widely produced by various bacterial species referred as plant-beneficial bacteria (Ongena and Jacques, 2008; D’aes et al., 2010). Among them, Bacillus and Pseudomonas are exploited as biocontrol agents and are also the best known for the production of a range of structurally distinct and multifunctional CLPs with strong biological activities related to plant protection (D’aes et al., 2010; Raaijmakers et al., 2010). Bacterial CLPs are powerful biosurfactants retaining strong destabilizing activities on biological membranes. Their antimicrobial activity is well documented in a context of biocontrol via direct inhibition of phytopathogens. In vitro-based assays using purified CLPs combined or not with loss of function mutants of natural producers have highlighted the extremely wide range of fungal and oomycete plant pathogens that are affected by bacterial CLPs such as fengycins and iturins (see recent reviews (Caulier et al., 2019; Rabbee et al., 2019) for Bacillus and (Geudens and Martins, 2018; Götze and Stallforth, 2020) for Pseudomonas CLPs, respectively). Many studies indicate that CLP activity is linked to their capacity to compromise the fungal cell membrane stability, resulting in cytoplasm leakage and hyphae death or inhibition of spore germination (Chitarra et al., 2003; Romero et al., 2007; Etchegaray et al., 2008; Pérez-García et al., 2011; Gong et al., 2015; Qian et al., 2016). However, the mechanistic basis of antifungal activity may be more complex and, as for rhamnolipids, the lipid composition of the targeted cell membrane could play an essential role in the microbicidal activity (Grau et al., 1999; Tao et al., 2011; Wise et al., 2014). Like other antimicrobial peptides, CLPs are not only membrane disruptive but can also directly or indirectly act on intracellular targets and alter fungal cell functions (Latoud et al., 1987; Qi et al., 2010).
Antibacterial activity has also been occasionally reported for Bacillus CLPs such as iturin A, bacillomycin and locillomycins toward several plant pathogens of agronomic importance (Zeriouh et al., 2011; Luo et al., 2015; Cao et al., 2018). However, there are globally few convincing evidences for a direct bactericidal effect of Bacillus CLPs and surfactin in particular on phytopathogens or soil-borne bacterial attackers. The precise antibiotic mechanistic of Bacillus CLPs against bacterial phytopathogens remains unclear even if a direct interaction with the cellular membrane of the target is also obvious (Zeriouh et al., 2011; Gao et al., 2017). However, in some instances, the inhibitory effect of some Bacillus CLPs such as surfactin (or related lichenysin and pumilacidin) is not related to a direct effect on target cell viability but rather due to some interference with key developmental processes of the pathogen such as efficient biofilm formation by Pseudomonas syringae and Ralstonia solanacearum (Bais et al., 2004; Chen et al., 2013; Xiu et al., 2017) or inhibition of aerial hyphal development of Streptomyces coelicolor (Straight et al., 2006; Hoefler et al., 2012).
Stimulation of Plant Immunity by Biosurfactants
Plants have developed complex defense mechanisms leading to enhance resistance to phytopathogens. After microbial perception, early signaling events are set up including ion fluxes, reactive oxygen species (ROS) accumulation and phosphorylation cascades (Garcia-Brugger et al., 2006; Bigeard et al., 2015). These early signaling and the activation of an intricate network of phytohormones, such as salicylic acid or jasmonic acid, regulate late defense-related responses (Pieterse et al., 2012) including synthesis of antimicrobial metabolites and cell wall reinforcement. These defense responses collectively allow local plant immunity (Boller and Felix, 2009). In addition, microbial perception triggers systemic responses that are effective against a large panel of microorganisms in the whole plant (Fu and Dong, 2013; Pieterse et al., 2014). Activation of the plant immune system involves invasion patterns (IPs) molecules also known as elicitors which can originate from or be produced by the microbe (Schellenberger et al., 2019).
Rhamnolipids Trigger Local Resistance Against Plant Pathogens
Whereas most studies on glycolipid biosurfactants were focused on their antimicrobial and antifouling activities, it was recently discovered that rhamnolipids may also stimulate plant innate immunity (Vatsa et al., 2010; Figure 1). Interestingly, despite their antimicrobial and mammalian immunomodulatory properties, to our knowledge, no study on sophorolipids, trehalolipids, MELs or cellobiose lipids, described their potential role in the activation of plant defense responses so far. Following plant sensing, rhamnolipids trigger early signaling events like accumulation of ROS in grapevine and Brassica napus (Varnier et al., 2009; Monnier et al., 2018) as well as a calcium influx and a phosphorylation cascade in grapevine (Varnier et al., 2009). Callose deposition, hormone production, defense gene activation and a hypersensitive reaction-like response are also hallmarks of rhamnolipid-triggered immunity in Brassicaceae and grapevine (Varnier et al., 2009; Sanchez et al., 2012; Monnier et al., 2018, 2020). It was demonstrated in Arabidopsis that rhamnolipid-mediated local resistance to Botrytis cinerea, Hyaloperonospora arabidopsidis or P. syringae pv. tomato (Pst) involves different signaling pathways that depend on the type of pathogen (Sanchez et al., 2012). In addition, rhamnolipid potentiate defense responses induced by other elicitors like chitosan. The immune response triggered by rhamnolipids also participates in local resistance against B. cinerea and the hemibiotrophic fungus Leptosphaeria maculans in B. napus (Monnier et al., 2018, 2020). A large range of rhamnolipid concentrations from 0.005 to 1 mg/mL have been used to induce immunity on these various plant species (Varnier et al., 2009; Sanchez et al., 2012; Monnier et al., 2018, 2020). Synthetic biosurfactants derived from rhamnolipid structure are also elicitors. For instance, synthetic rhamnolipid bolaforms, composed of two rhamnoses separated by a fatty acid chain, trigger an immune response in Arabidopsis that varies according to fatty acid chain length (Luzuriaga-Loaiza et al., 2018). In addition, RL harboring carboxylic acid (Ac-RL) and methyl (Alk-RL) induce ROS production in this plant (Nasir et al., 2017).
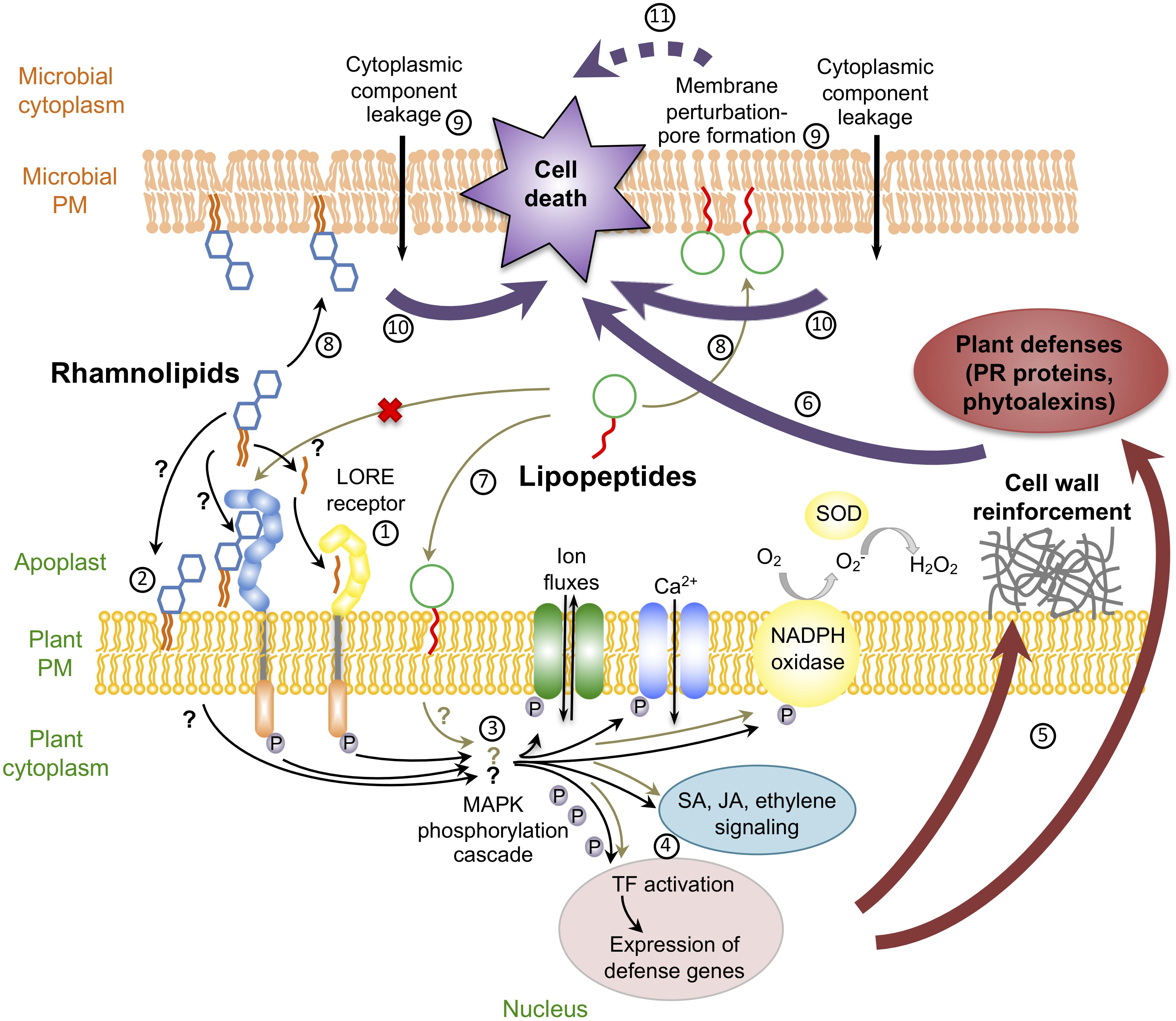
Figure 1. Schematic representation of dual effects of rhamnolipids and lipopeptides: antimicrobial activities and plant defense induction. mc-3-OH-acyl building block of rhamnolipids is perceived by plant through the LORE receptor ①; Rhamnolipid could be sensed through their direct insertion in plasma membrane ②. Recognition of rhamnolipids leads to early signaling events like ion fluxes (Ca2+), reactive oxygen species production (H2O2) and MAPK phosphorylation cascade ③. These early responses trigger defense gene expression, probably through activation of transcription factors (TF) and hormonal signaling pathways ④. This leads to defense mechanisms like cell wall reinforcement and PR protein accumulation ⑤ triggering the resistance to the microbes ⑥. Plant immunity due to lipopeptides does not involve a protein receptor and rely on interaction between lipopeptides and the plant membrane ⑦. Both rhamnolipids and lipopeptides can also have direct antimicrobial effects through direct insertion into the microbial plasma membrane ⑧. These insertions trigger loss of cell morphology leading to pore formation ⑨. The pore formation causes cellular component leakage triggering microbial cell death ⑩. Cell death due to lipopeptides can also be indirectly due to the inhibition or activation of microbial cell functions  .
.
The way by which rhamnolipids are perceived by plant cells still remains unknown. Given their amphiphilic nature, it is postulated that they could interact with plant membrane lipids (Sanchez et al., 2012; Schellenberger et al., 2019). Recently it has been demonstrated that natural rhamnolipids fit into plant lipid-based membrane models and are located near the lipid phosphate group of the phospholipid bilayers, nearby phospholipid glycerol backbones (Monnier et al., 2019). Rhamnolipid insertion inside the lipid bilayer does not strongly affect lipid dynamics but the nature of the phytosterols could influence the effect of the glycolipids on plant plasma membrane destabilization. These subtle changes in lipid dynamics could be linked with plant defense induction (Monnier et al., 2019). Interestingly, whereas no receptor for rhamnolipid perception has been identified so far, the mc-3-OH-acyl building block of rhamnolipids is sensed by the lectin S-domain-1 receptor-like kinase LORE (Kutschera et al., 2019; Figure 1).
For some studies, it is not demonstrated whether rhamnolipid-triggered protection is driven by activation of plant defense responses and/or antimicrobial properties. For instance, treatments of pepper plants with rhamnolipids result in an enhanced protection to Phytophthora blight disease and also prevent the development of Colletotrichum orbiculare infection on leaves of cucumber plants (Kim et al., 2000). Rhamnolipids significantly decrease the incidence of water-borne damping-off disease by Phytophthora sp. and Pythium sp. (Yoo et al., 2005). Similar results were obtained in field trials on chili pepper and tomato (Sharma et al., 2007a, b). Using bacterial mutants, it was demonstrated that phenazine and rhamnolipids interact in the biological control of soil-borne diseases caused by Pythium sp. (Perneel et al., 2008). Syringomycin E and rhamnolipids can also act synergistically to control pathogenic and opportunistic fungi recovered from diseased grape (Takemoto et al., 2010). The control of postharvest phytopathogens on seeds or fruits for a better conservation is often related to antimicrobial activities. But we cannot exclude that protection could also be due to plant defense responses (Borah et al., 2016). When applied alone, rhamnolipids induce antioxidative reactions in cherry tomato fruit, leading to a significant reduction of fungal disease (Yan et al., 2015). When applied in combination with the biocontrol yeast agent Rhodotorula glutinis, a synergistic inhibitory effect on Alternaria alternata infection could be observed in cherry tomato fruit, leading to an efficient protection (Yan et al., 2014). This protection is associated with a higher induction of defense-related enzymes and the accumulation of antimicrobial metabolites.
Lipopeptides as Powerful Inducers of Plant Systemic Resistance
Several studies have reported the involvement of Bacillus CLPs in plant immunity induction on various pathosystems. The potential of fengycin and surfactin CLPs to trigger plant systemic resistance was first shown on bean and tomato plants. When applied as pure compounds at micromolar concentrations, surfactin and to a lower extend fengycin induced significant disease reduction in bean and tomato infected with B. cinerea (Ongena et al., 2007). More recently, a study performed with a large range of natural Bacillus isolates strengthened the role of surfactin as ISR (induced systemic resistance) inducer since strong correlation was observed between defense-inducing activity and the amount of surfactin produced by the different strains (Cawoy et al., 2014). In the same way, B. velezensis FZB42 mutant strains unable to synthesize surfactin are impaired in their ISR to Rhizoctonia solani in lettuce (Chowdhury et al., 2015). Further studies allowed enlarging the ISR elicitor role of surfactin to other plants. For example, purified surfactin was shown to increase resistance against the cucurbit powdery mildew in melon plants (García-Gutiérrez et al., 2013). In the pathosystem citrus fruit/Penicillium digitatum, surfactin stimulates defense responses involved in generating signal molecules for ISR activation (Waewthongrak et al., 2014). This lipopeptide activates a plant innate response effective against Magnaporthe oryzae in perennial ryegrass (Rahman et al., 2015) or Plasmopara viticola in grapevine (Li et al., 2019). It also reduces infection by the rhizomania disease vector Polymyxa betae in sugar beet (Desoignies et al., 2013) or by Colletotrichum gloeosporioides in strawberry leaves (Yamamoto et al., 2015). Finally, a recent study showed that Sclerotium rolfsii disease incidence was strongly reduced in Arachis hypogaea when pretreated with surfactin (Rodríguez et al., 2018). Interestingly, CLPs like surfactin do not globally provoke a strong plant defensive response associated with major genetic reprograming and fitness cost but rather act by priming host defenses to trigger systemic resistance (Ongena et al., 2007; Jourdan et al., 2009; Debois et al., 2015). Induction of plant defenses by CLPs of the iturin group has also been occasionally reported. Iturin A was shown to have a similar role as surfactin in strawberry leaves (Yamamoto et al., 2015) and also acted as an inducer of plant defense gene expression in cotton plants upon Verticillium dahliae attack (Han et al., 2015). Mycosubtilin is the most efficient lipopeptide inducing an immune response in grapevine (Farace et al., 2015). Compared to surfactin, bacillomycin D produced by B. velezensis SQR9 has a comparable efficacy in Arabidopsis ISR elicitation to prevent infection by P. syringae or B. cinerea (Wu et al., 2018). In wheat plants, resistance toward Zymoseptoria tritici, was activated by pure surfactin used at concentrations ranging from 1 to 100 μM upon foliar application (Le Mire et al., 2018). Some CLPs synthesized by Pseudomonas sp. also display consistent ISR-triggering activity. It was first demonstrated that massetolide A produced by Pseudomonas fluorescens strain SS101 retains ISR-eliciting activity in tomato plants for the control of Phytophthora infestans (Tran et al., 2007). Pseudomonas sp. strain CMR12a is a soil isolate retaining high biocontrol potential against R. solani relying mainly on the interplay between two different lipopeptides (sessilin and orfamide) and phenazine for inducing plant immunity (D’aes et al., 2011, 2014). These CLPs were also active at protecting Brassica chinensis against R. solani (Olorunleke et al., 2015). In monocots, such as rice, orfamide and other Pseudomonas CLPs such as WLIP, lokisin and entolysin, successfully induced resistance toward C. miyabeanus or M. oryzae (Ma et al., 2016, 2017; Omoboye et al., 2019).
Up to now, how lipopeptides act and are recognized by plant cells to activate ISR remains unclear. CLPs are in most instances only active in micromolar concentrations, and defenses are more intensively induced at the highest surfactin doses. This suggests that the recognition mechanism at the plant cell surface should be of quite low affinity in contrast to other elicitors (Jourdan et al., 2009). Such a low specificity may be explained by the fact that CLPs like surfactin are not perceived by a protein receptor, but rather involve a process driven by an uncommon pathway based on interaction with the lipid bilayer fraction of plant plasma membranes (Henry et al., 2011). This is supported by some studies revealing that CLP structure plays an important role for the ISR eliciting activity. Both the fatty acid chain length, the cyclic conformation of the molecule and amino acid positions in the peptide chain impact the eliciting potential of surfactin in tobacco cells (Jourdan et al., 2009; Henry et al., 2011). The activation of defense genes in Arabidopsis upon iturin A foliar treatment was also dependent on the structure of the molecule, i.e., cyclization and/or nature of the fatty acid chain (Kawagoe et al., 2015). As for rhamnolipids, the hypothesis is that lipopeptides have the ability to create some disturbance in the plant plasma membrane and could consequently activate a cascade of molecular events leading to the activation of defense mechanisms (Schellenberger et al., 2019; Figure 1).
Conclusion
Biosurfactants, produced by bacteria, yeast, and fungi, are promising molecules for a wide variety of applications due to their potential to be commercially produced at large scales, their low toxicity and high biodegradability. In this mini-review, we provided evidences about the potential of rhamnolipids and lipopeptides for plant protection in a context of sustained agriculture. These molecules have similar dual effects by protecting plants through antimicrobial properties and stimulation of local and/or systemic plant immunity. These singular properties are essential for the efficiency of these biopesticides. Although numerous elicitors are perceived by plasma membrane receptors, recent studies on amphiphilic biosurfactants such as rhamnolipids or lipopeptides suggest that they are sensed by an uncommon way involving lipids in the bilayer of the plant plasma membrane that could explain their singular elicitor activity. To better understand the mechanisms of action of biosurfactants, experiments or trials need to be realized not only on mixture but also on highly purified molecules in the future. Nevertheless, several obstacles to the development of rhamnolipid and lipopeptide applications still remain. Biosurfactant costs, their efficacies in the field and purity of compounds have to be improved to allow their use at a higher degree in crop protection. In this respect, combination of biosurfactants should be considered to increase efficacy in field conditions. Finally, given their interesting properties it is now time to really consider ecofriendly biosurfactants as biocontrol solutions in integrated pest management.
Author Contributions
JC, AA-A, SD-C, SC, MO, and SD participated in the conception, information search and assisted in drafting the manuscript. JP, GH, FM-G, FB, and CC assisted in drafting and correction of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
Research in the MO lab is funded by the EOS (excellence of science, FRS-FNRS) grant no 2018000802.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Support from the MESRI (Ministère de l’Enseignement Supérieur, de la Recherche et de l’Innovation), the Federative Research Structure SFR Condorcet and the Interreg V France-wallonie-vlaanderen program Smartbiocontrol/Bioscreen are gratefully acknowledged. MO thanks the Fonds National de la Recherche Scientifique from Belgium (FRS-FNRS) for his position as Senior Research Associate.
References
Abalos, A., Pinazo, A., Infante, M. R., Casals, M., García, F., and Manresa, A. (2001). Physicochemical and antimicrobial properties of new rhamnolipids produced by Pseudomonas aeruginosa AT10 from soybean oil refinery wastes. Langmuir 17, 1367–1371. doi: 10.1021/la0011735
Abbasi, H., Aranda, F. J., Noghabi, K. A., and Ortiz, A. (2013). A bacterial monorhamnolipid alters the biophysical properties of phosphatidylethanolamine model membranes. Biochim. Biophys. Acta Biomemb. 1828, 2083–2090. doi: 10.1016/j.bbamem.2013.04.024
Abbasi, H., Noghabi, K. A., and Ortiz, A. (2012). Interaction of a bacterial monorhamnolipid secreted by Pseudomonas aeruginosa MA01 with phosphatidylcholine model membranes. Chem. Phys. Lipids 165, 745–752. doi: 10.1016/j.chemphyslip.2012.09.001
Abdel-Mawgoud, A. M., Lepine, F., and Deziel, E. (2010). Rhamnolipids: diversity of structures, microbial origins and roles. Appl. Microbiol. Biotechnol. 86, 1323–1336. doi: 10.1007/s00253-010-2498-2
Aranda, F. J., Espuny, M. J., Marqués, A., Teruel, J. A., and Manresa, Á, and Ortiz, A. (2007). Thermodynamics of the interaction of a dirhamnolipid biosurfactant secreted by Pseudomonas aeruginosa with phospholipid membranes. Langmuir 23, 2700–2705. doi: 10.1021/la061464z
Aslam, S. N., Erbs, G., Morrissey, K. L., Newman, M.-A., Chinchilla, D., Boller, T., et al. (2009). Microbe-associated molecular pattern (MAMP) signatures, synergy, size and charge: influences on perception or mobility and host defence responses. Mol. Plant Pathol. 10, 375–387. doi: 10.1111/j.1364-3703.2009.00537.x
Bais, H. P., Fall, R., and Vivanco, J. M. (2004). Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol. 134, 307–319. doi: 10.1104/pp.103.028712
Banat, I. M., Franzetti, A., Gandolfi, I., Bestetti, G., Martinotti, M. G., Fracchia, L., et al. (2010). Microbial biosurfactants production, applications and future potential. Appl. Microbiol. Biotechnol. 87, 427–444. doi: 10.1007/s00253-010-2589-0
Bardin, M., Ajouz, S., Comby, M., Lopez-Ferber, M., Graillot, B., Siegwart, M., et al. (2015). Is the efficacy of biological control against plant diseases likely to be more durable than that of chemical pesticides? Front. Plant Sci. 6:566. doi: 10.3389/fpls.2015.00566
Benincasa, M., Abalos, A., Oliveira, I., and Manresa, A. (2004). Chemical structure, surface properties and biological activities of the biosurfactant produced by Pseudomonas aeruginosa LBI from soapstock. Antonie Van Leeuwenhoek 85, 1–8. doi: 10.1023/B:ANTO.0000020148.45523.415105181
Berg, G., Koberl, M., Rybakova, D., Muller, H., Grosch, R., and Smalla, K. (2017). Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol. Ecol. 93:fix050. doi: 10.1093/femsec/fix050
Bigeard, J., Colcombet, J., and Hirt, H. (2015). Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 8, 521–539. doi: 10.1016/j.molp.2014.12.022
Boller, T., and Felix, G. (2009). A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 60, 379–406. doi: 10.1146/annurev.arplant.57.032905.105346
Bonanomi, G., Lorito, M., Vinale, F., and Woo, S. L. (2018). Organic amendments, beneficial microbes, and soil microbiota: toward a unified framework for disease suppression. Annu. Rev. Phytopathol. 56, 1–20. doi: 10.1146/annurev-phyto-080615-100046
Borah, S. N., Goswami, D., Lahkar, J., Sarma, H. K., Khan, M. R., and Deka, S. (2015). Rhamnolipid produced by Pseudomonas aeruginosa SS14 causes complete suppression of wilt by Fusarium oxysporum f. sp. pisi in Pisum sativum. Biocontrol 60, 375–385. doi: 10.1007/s10526-014-9645-0
Borah, S. N., Goswami, D., Sarma, H. K., Cameotra, S. S., and Deka, S. (2016). Rhamnolipid biosurfactant against Fusarium verticillioides to control stalk and ear rot disease of maize. Front. Microbiol. 7:1505. doi: 10.3389/fmicb.2016.01505
Burketova, L., Trda, L., Ott, P. G., and Valentova, O. (2015). Bio-based resistance inducers for sustainable plant protection against pathogens. Biotechnol. Adv. 33(6 Pt 2), 994–1004. doi: 10.1016/j.biotechadv.2015.01.004
Cao, Y., Pi, H., Chandrangsu, P., Li, Y., Wang, Y., Zhou, H., et al. (2018). Antagonism of two plant-growth promoting Bacillus velezensis isolates against Ralstonia solanacearum and Fusarium oxysporum. Sci. Rep. 8:4360. doi: 10.1038/s41598-018-22782-z
Caulier, S., Nannan, C., Gillis, A., Licciardi, F., Bragard, C., and Mahillon, J. (2019). Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front. Microbiol. 10:302. doi: 10.3389/fmicb.2019.00302
Cawoy, H., Mariutto, M., Henry, G., Fisher, C., Vasilyeva, N., Thonart, P., et al. (2014). Plant defense stimulation by natural isolates of Bacillus depends on efficient surfactin production. Mol. Plant Microb. Interact. 27, 87–100. doi: 10.1094/mpmi-09-13-0262-r
Chen, J., Liu, X., Fu, S., An, Z., Feng, Y., Wang, R., et al. (2020). Effects of sophorolipids on fungal and oomycete pathogens in relation to pH solubility. J. Appl. Microbiol. 128, 1754–1763. doi: 10.1111/jam.14594
Chen, Y., Yan, F., Chai, Y., Liu, H., Kolter, R., Losick, R., et al. (2013). Biocontrol of tomato wilt disease by Bacillus subtilis isolates from natural environments depends on conserved genes mediating biofilm formation. Environ. Microbiol. 15, 848–864. doi: 10.1111/j.1462-2920.2012.02860.x
Chitarra, G. S., Breeuwer, P., Nout, M. J. R., Van Aelst, A. C., Rombouts, F. M., and Abee, T. (2003). An antifungal compound produced by Bacillus subtilis YM 10-20 inhibits germination of Penicillium roqueforti conidiospores. J. Appl. Microbiol. 94, 159–166. doi: 10.1046/j.1365-2672.2003.01819.x
Chowdhury, S. P., Hartmann, A., Gao, X., and Borriss, R. (2015). Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42 - a review. Front. Microbiol. 6:780. doi: 10.3389/fmicb.2015.00780
Coelho, A. L. S., Feuser, P. E., Carciofi, B. A. M., de Andrade, C. J., and de Oliveira, D. (2020). Mannosylerythritol lipids: antimicrobial and biomedical properties. Appl. Microbiol. Biotechnol. 104, 2297–2318. doi: 10.1007/s00253-020-10354-z
D’aes, J., De Maeyer, K., Pauwelyn, E., and Höfte, M. (2010). Biosurfactants in plant-Pseudomonas interactions and their importance to biocontrol. Environ. Microbiol. Rep. 2, 359–372. doi: 10.1111/j.1758-2229.2009.00104.x
D’aes, J., Hua, G. K., De Maeyer, K., Pannecoucque, J., Forrez, I., Ongena, M., et al. (2011). Biological control of Rhizoctonia root rot on bean by phenazine- and cyclic lipopeptide-producing Pseudomonas CMR12a. Phytopathology 101, 996–1004. doi: 10.1094/phyto-11-10-0315
D’aes, J., Kieu, N. P., Léclère, V., Tokarski, C., Olorunleke, F. E., De Maeyer, K., et al. (2014). To settle or to move? The interplay between two classes of cyclic lipopeptides in the biocontrol strain Pseudomonas CMR12a. Environ. Microbiol. 16, 2282–2300. doi: 10.1111/1462-2920.12462
Dashtbozorg, S. S., Miao, S., and Ju, L.-K. (2016). Rhamnolipids as environmentally friendly biopesticide against plant pathogen Phytophthora sojae. Environ. Prog. Sustain. Energy 35, 169–173. doi: 10.1002/ep.12187
De Jonghe, K., De Dobbelaere, I., Sarrazyn, R., and Höfte, M. (2005). Control of Phytophthora cryptogea in the hydroponic forcing of witloof chicory with the rhamnolipid-based biosurfactant formulation PRO1. Plant Pathol. 54, 219–226.
Debois, D., Fernandez, O., Franzil, L., Jourdan, E., de Brogniez, A., Willems, L., et al. (2015). Plant polysaccharides initiate underground crosstalk with bacilli by inducing synthesis of the immunogenic Lipopeptide surfactin. Environ. Microbiol. Rep. 7, 570–582. doi: 10.1111/1758-2229.12286
Deepika, K. V., Ramu Sridhar, P., and Bramhachari, P. V. (2015). Characterization and antifungal properties of rhamnolipids produced by mangrove sediment bacterium Pseudomonas aeruginosa strain KVD-HM52. Biocatalys. Agric. Biotechnol. 4, 608–615. doi: 10.1016/j.bcab.2015.09.009
Delaunois, B., Farace, G., Jeandet, P., Clement, C., Baillieul, F., Dorey, S., et al. (2014). Elicitors as alternative strategy to pesticides in grapevine? Current knowledge on their mode of action from controlled conditions to vineyard. Environ. Sci. Pollut. Res. Intern. 21, 4837–4846. doi: 10.1007/s11356-013-1841-4
Desoignies, N., Schramme, F., Ongena, M., and Legrève, A. (2013). Systemic resistance induced by Bacillus lipopeptides in Beta vulgaris reduces infection by the rhizomania disease vector Polymyxa betae. Mol. Plant Pathol. 14, 416–421. doi: 10.1111/mpp.12008
Etchegaray, A., de Castro Bueno, C., de Melo, I. S., Tsai, S. M., de Fátima Fiore, M., Silva-Stenico, M. E., et al. (2008). Effect of a highly concentrated lipopeptide extract of Bacillus subtilis on fungal and bacterial cells. Archiv. Microbiol. 190, 611–622. doi: 10.1007/s00203-008-0409-z
Farace, G., Fernandez, O., Jacquens, L., Coutte, F., Krier, F., Jacques, P., et al. (2015). Cyclic lipopeptides from Bacillus subtilis activate distinct patterns of defence responses in grapevine. Mol. Plant Pathol. 16, 177–187. doi: 10.1111/mpp.12170
Fu, Z. Q., and Dong, X. (2013). Systemic acquired resistance: turning local infection into global defense. Annu. Rev. Plant Biol. 64, 839–863. doi: 10.1146/annurev-arplant-042811-105606
Gao, L., Han, J., Liu, H., Qu, X., Lu, Z., and Bie, X. (2017). Plipastatin and surfactin coproduction by Bacillus subtilis pB2-L and their effects on microorganisms. Antonie Van Leeuwenhoek 110, 1007–1018. doi: 10.1007/s10482-017-0874-y
Garcia-Brugger, A., Lamotte, O., Vandelle, E., Bourque, S., Lecourieux, D., Poinssot, B., et al. (2006). Early signaling events induced by elicitors of plant defenses. Mol. Plant Microb. Interact. 19, 711–724.
García-Gutiérrez, L., Zeriouh, H., Romero, D., Cubero, J., de Vicente, A., and Pérez-García, A. (2013). The antagonistic strain Bacillus subtilis UMAF6639 also confers protection to melon plants against cucurbit powdery mildew by activation of jasmonate- and salicylic acid-dependent defence responses. Microb. Biotechnol. 6, 264–274. doi: 10.1111/1751-7915.12028
Geudens, N., and Martins, J. C. (2018). Cyclic lipodepsipeptides from Pseudomonas spp. - biological swiss-army knives. Front. Microbiol. 9:1867. doi: 10.3389/fmicb.2018.01867
Gong, A.-D., Li, H.-P., Yuan, Q.-S., Song, X.-S., Yao, W., He, W.-J., et al. (2015). Antagonistic mechanism of iturin A and plipastatin A from Bacillus amyloliquefaciens S76-3 from wheat spikes against Fusarium graminearum. PLoS One 10:e0116871. doi: 10.1371/journal.pone.0116871
Goswami, D., Borah, S. N., Lahkar, J., Handique, P. J., and Deka, S. (2015). Antifungal properties of rhamnolipid produced by Pseudomonas aeruginosa DS9 against Colletotrichum falcatum. J. Basic Microbiol. 55, 1265–1274. doi: 10.1002/jobm.201500220
Goswami, D., Handique, P. J., and Deka, S. (2014). Rhamnolipid biosurfactant against Fusarium sacchari–the causal organism of pokkah boeng disease of sugarcane. J. Basic Microbiol. 54, 548–557. doi: 10.1002/jobm.201200801
Götze, S., and Stallforth, P. (2020). Structure, properties, and biological functions of nonribosomal lipopeptides from pseudomonads. Nat. Product Rep. 37, 29–54. doi: 10.1039/c9np00022d
Grau, A., Gómez Fernández, J. C., Peypoux, F., and Ortiz, A. (1999). A study on the interactions of surfactin with phospholipid vesicles. Biochim. Biophys. Acta Biomemb. 1418, 307–319. doi: 10.1016/S0005-2736(99)00039-5
Haba, E., Pinazo, A., Jauregui, O., Espuny, M. J., Infante, M. R., and Manresa, A. (2003). Physiochemical characterization and antimicrobial properties of rhamnolipids produced by Pseudomonas aeruginosa 47T2 NCBIM 40044. Biotechnol. Bioeng. 81, 316–322.
Han, Q., Wu, F., Wang, X., Qi, H., Shi, L., Ren, A., et al. (2015). The bacterial lipopeptide iturins induce Verticillium dahliae cell death by affecting fungal signalling pathways and mediate plant defence responses involved in pathogen-associated molecular pattern-triggered immunity. Environ. Microbiol. 17, 1166–1188. doi: 10.1111/1462-2920.12538
Henry, G., Deleu, M., Jourdan, E., Thonart, P., and Ongena, M. (2011). The bacterial lipopeptide surfactin targets the lipid fraction of the plant plasma membrane to trigger immune-related defence responses. Cell. Microbiol. 13, 1824–1837. doi: 10.1111/j.1462-5822.2011.01664.x
Hoefler, B. C., Gorzelnik, K. V., Yang, J. Y., Hendricks, N., Dorrestein, P. C., and Straight, P. D. (2012). Enzymatic resistance to the lipopeptide surfactin as identified through imaging mass spectrometry of bacterial competition. Proc. Natl. Acad. Sci. U.S.A. 109, 13082–13087. doi: 10.1073/pnas.1205586109
Jahan, R., Bodratti, A. M., Tsianou, M., and Alexandridis, P. (2020). Biosurfactants, natural alternatives to synthetic surfactants: physicochemical properties and applications. Adv. Coll. Interf. Sci. 275:102061. doi: 10.1016/j.cis.2019.102061
Jourdan, E., Henry, G., Duby, F., Dommes, J., Barthelemy, J. P., Thonart, P., et al. (2009). Insights into the defense-related events occurring in plant cells following perception of surfactin-type lipopeptide from Bacillus subtilis. Mol. Plant Microb. Interact. 22, 456–468. doi: 10.1094/MPMI-22-4-0456
Kawagoe, Y., Shiraishi, S., Kondo, H., Yamamoto, S., Aoki, Y., and Suzuki, S. (2015). Cyclic lipopeptide iturin A structure-dependently induces defense response in Arabidopsis plants by activating SA and JA signaling pathways. Biochem. Biophys. Res. Commun. 460, 1015–1020. doi: 10.1016/j.bbrc.2015.03.143
Keswani, C., Singh, H. B., Hermosa, R., Garcia-Estrada, C., Caradus, J., He, Y. W., et al. (2019). Antimicrobial secondary metabolites from agriculturally important fungi as next biocontrol agents. Appl. Microbiol. Biotechnol. 103, 9287–9303. doi: 10.1007/s00253-019-10209-2
Kim, B. S., Lee, J. Y., and Hwang, B. K. (2000). In vivo control and in vitro antifungal activity of rhamnolipid B, a glycolipid antibiotic, against Phytophthora capsici and Colletotrichum orbiculare. Pest Manag. Sci. 56, 1029–1035.
Kutschera, A., Dawid, C., Gisch, N., Schmid, C., Raasch, L., Gerster, T., et al. (2019). Bacterial medium-chain 3-hydroxy fatty acid metabolites trigger immunity in Arabidopsis plants. Science 364, 178–181. doi: 10.1126/science.aau1279
Lang, S., Katsiwela, E., and Wagner, F. (1989). Antimicrobial effects of biosurfactants. Lipid Fett 91, 363–366. doi: 10.1002/lipi.19890910908
Latoud, C., Peypoux, F., and Michel, G. (1987). Action of iturin A, an antifungal antibiotic from Bacillus subtilis, on the yeast Saccharomyces cerevisiae: modifications of membrane permeability and lipid composition. J. Antibiot. 40, 1588–1595. doi: 10.7164/antibiotics.40.1588
Le Mire, G., Siah, A., Brisset, M.-N., Gaucher, M., Deleu, M., and Jijakli, M. H. (2018). Surfactin Protects Wheat against Zymoseptoria tritici and activates both salicylic acid- and jasmonic acid-dependent defense responses. Agriculture 8:11.
Li, Y., Héloir, M.-C., Zhang, X., Geissler, M., Trouvelot, S., Jacquens, L., et al. (2019). Surfactin and fengycin contribute to the protection of a Bacillus subtilis strain against grape downy mildew by both direct effect and defence stimulation. Mol. Plant Pathol. 20, 1037–1050. doi: 10.1111/mpp.12809
Luo, C., Zhou, H., Zou, J., Wang, X., Zhang, R., Xiang, Y., et al. (2015). Bacillomycin L and surfactin contribute synergistically to the phenotypic features of Bacillus subtilis 916 and the biocontrol of rice sheath blight induced by Rhizoctonia solani. Appl. Microbiol. Biotechnol. 99, 1897–1910. doi: 10.1007/s00253-014-6195-4
Luzuriaga-Loaiza, P., Schellenberger, R., De Gaetano, Y., Obounou Akong, F., Villaume, S., Crouzet, J., et al. (2018). Synthetic Rhamnolipid bolaforms trigger an innate immune response in Arabidopsis thaliana. Sci. Rep. 8:8534.
Ma, Z., Hua, G. K. H., Ongena, M., and Hofte, M. (2016). Role of phenazines and cyclic lipopeptides produced by pseudomonas sp. CMR12a in induced systemic resistance on rice and bean. Environ. Microbiol. Rep. 8, 896–904. doi: 10.1111/1758-2229.12454
Ma, Z., Ongena, M., and Hofte, M. (2017). The cyclic lipopeptide orfamide induces systemic resistance in rice to Cochliobolus miyabeanus but not to Magnaporthe oryzae. Plant Cell Rep. 36, 1731–1746. doi: 10.1007/s00299-017-2187-z
Miao, S., Dashtbozorg, S. S., Callow, N. V., and Ju, L.-K. (2015). Rhamnolipids as platform molecules for production of potential anti-zoospore agrochemicals. J. Agric. Food Chem. 63, 3367–3376. doi: 10.1021/acs.jafc.5b00033
Mnif, I., and Ghribi, D. (2016). Glycolipid biosurfactants: main properties and potential applications in agriculture and food industry. J. Sci. Food Agric. 96, 4310–4320. doi: 10.1002/jsfa.7759
Monnier, N., Cordier, M., Dahi, A., Santoni, V., Guenin, S., Clement, C., et al. (2020). Semi-purified rhamnolipid mixes protect Brassica napus against Leptosphaeria maculans early infections. Phytopathology 110, 834–842. doi: 10.1094/phyto-07-19-0275-r
Monnier, N., Furlan, A., Botcazon, C., Dahi, A., Mongelard, G., Cordelier, S., et al. (2018). Rhamnolipids from Pseudomonas aeruginosa are elicitors triggering Brassica napus protection against Botrytis cinerea without physiological disorders. Front. Plant Sci. 9:1170. doi: 10.3389/fpls.2018.01170
Monnier, N., Furlan, A. L., Buchoux, S., Deleu, M., Dauchez, M., Rippa, S., et al. (2019). Exploring the dual interaction of natural rhamnolipids with plant and fungal biomimetic plasma membranes through biophysical studies. Intern. J. Mol. Sci. 20:1009. doi: 10.3390/ijms20051009
Nalini, S., and Parthasarathi, R. (2014). Production and characterization of rhamnolipids produced by Serratia rubidaea SNAU02 under solid-state fermentation and its application as biocontrol agent. Bioresour. Technol. 173, 231–238. doi: 10.1016/j.biortech.2014.09.051
Nasir, M. N., Lins, L., Crowet, J. M., Ongena, M., Dorey, S., Dhondt-Cordelier, S., et al. (2017). Differential interaction of synthetic glycolipids with biomimetic plasma membrane lipids correlates with the plant biological response. Langmuir 33, 9979–9987. doi: 10.1021/acs.langmuir.7b01264
Naughton, P. J., Marchant, R., Naughton, V., and Banat, I. M. (2019). Microbial biosurfactants: current trends and applications in agricultural and biomedical industries. J. Appl. Microbiol. 127, 12–28. doi: 10.1111/jam.14243
Olorunleke, F. E., Hua, G. K. H., Kieu, N. P., Ma, Z., and Höfte, M. (2015). Interplay between orfamides, sessilins and phenazines in the control of Rhizoctonia diseases by Pseudomonas sp. CMR12a. Environ. Microbiol. Rep. 7, 774–781. doi: 10.1111/1758-2229.12310
Omoboye, O. O., Oni, F. E., Batool, H., Yimer, H. Z., De Mot, R., and Höfte, M. (2019). Pseudomonas cyclic lipopeptides suppress the rice blast fungus magnaporthe oryzae by induced resistance and direct antagonism. Front. Plant Sci. 10:901. doi: 10.3389/fpls.2019.00901
Ongena, M., and Jacques, P. (2008). Bacillus lipopeptides: versatile weapons for plant disease biocontrol. Trends Microbiol. 16, 115–125.
Ongena, M., Jourdan, E., Adam, A., Paquot, M., Brans, A., Joris, B., et al. (2007). Surfactin and fengycin lipopeptides of Bacillus subtilis as elicitors of induced systemic resistance in plants. Environ. Microbiol. 9, 1084–1090. doi: 10.1111/j.1462-2920.2006.01202.x
Ortiz, A., Teruel, J. A., Espuny, M. J., Marqués, A., Manresa, Á, and Aranda, F. J. (2006). Effects of dirhamnolipid on the structural properties of phosphatidylcholine membranes. Intern. J. Pharm. 325, 99–107. doi: 10.1016/j.ijpharm.2006.06.028
Otzen, D. E. (2017). Biosurfactants and surfactants interacting with membranes and proteins: same but different? Biochim. Biophys. Acta Biomemb. 1859, 639–649. doi: 10.1016/j.bbamem.2016.09.024
Penha, R. O., Vandenberghe, L. P. S., Faulds, C., Soccol, V. T., and Soccol, C. R. (2020). Bacillus lipopeptides as powerful pest control agents for a more sustainable and healthy agriculture: recent studies and innovations. Planta 251:70. doi: 10.1007/s00425-020-03357-7
Pérez-García, A., Romero, D., and de Vicente, A. (2011). Plant protection and growth stimulation by microorganisms: biotechnological applications of Bacilli in agriculture. Curr. Opin. Biotechnol. 22, 187–193. doi: 10.1016/j.copbio.2010.12.003
Perneel, M., D’Hondt, L., De Maeyer, K., Adiobo, A., Rabaey, K., and Hofte, M. (2008). Phenazines and biosurfactants interact in the biological control of soil-borne diseases caused by Pythium spp. Environ. Microbiol. 10, 778–788.
Pieterse, C. M., Van der Does, D., Zamioudis, C., Leon-Reyes, A., and Van Wees, S. C. (2012). Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 28, 489–521. doi: 10.1146/annurev-cellbio-092910-154055
Pieterse, C. M. J., Zamioudis, C., Berendsen, R. L., Weller, D. M., Wees, S. C. M. V., and Bakker, P. A. H. M. (2014). Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 52, 347–375. doi: 10.1146/annurev-phyto-082712-102340
Popp, J., Petö, K., and Nagy, J. (2013). Pesticide productivity and food security. A review. Agron. Sustain. Dev. 33, 243–255. doi: 10.1007/s13593-012-0105-x
Qi, G., Zhu, F., Du, P., Yang, X., Qiu, D., Yu, Z., et al. (2010). Lipopeptide induces apoptosis in fungal cells by a mitochondria-dependent pathway. Peptides 31, 1978–1986. doi: 10.1016/j.peptides.2010.08.003
Qian, S., Lu, H., Sun, J., Zhang, C., Zhao, H., Lu, F., et al. (2016). Antifungal activity mode of Aspergillus ochraceus by bacillomycin D and its inhibition of ochratoxin A (OTA) production in food samples. Food Control 60, 281–288. doi: 10.1016/j.foodcont.2015.08.006
Raaijmakers, J. M., De Bruijn, I., Nybroe, O., and Ongena, M. (2010). Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol. Rev. 34, 1037–1062. doi: 10.1111/j.1574-6976.2010.00221.x
Rabbee, M. F., Ali, M. S., Choi, J., Hwang, B. S., Jeong, S. C., and Baek, K. H. (2019). Bacillus velezensis: a valuable member of bioactive molecules within plant microbiomes. Molecules 24:46. doi: 10.3390/molecules24061046
Rahman, A., Uddin, W., and Wenner, N. G. (2015). Induced systemic resistance responses in perennial ryegrass against Magnaporthe oryzae elicited by semi-purified surfactin lipopeptides and live cells of Bacillus amyloliquefaciens. Mol. Plant Pathol. 16, 546–558. doi: 10.1111/mpp.12209
Rodrigues, A. I., Gudina, E. J., Teixeira, J. A., and Rodrigues, L. R. (2017). Sodium chloride effect on the aggregation behaviour of rhamnolipids and their antifungal activity. Sci. Rep. 7:12907. doi: 10.1038/s41598-017-13424-x
Rodríguez, J., Tonelli, M. L., Figueredo, M. S., Ibáñez, F., and Fabra, A. (2018). The lipopeptide surfactin triggers induced systemic resistance and priming state responses in Arachis hypogaea L. Eur. J. Plant Pathol. 152, 845–851. doi: 10.1007/s10658-018-1524-6
Romero, D., de Vicente, A., Rakotoaly, R. H., Dufour, S. E., Veening, J.-W., Arrebola, E., et al. (2007). The Iturin and Fengycin families of lipopeptides are key factors in antagonism of Bacillus subtilis toward Podosphaera fusca. Mol. Plant Microb. Interact. 20, 430–440. doi: 10.1094/mpmi-20-4-0430
Sajid, M., Ahmad Khan, M. S., Singh Cameotra, S., and Safar Al-Thubiani, A. (2020). Biosurfactants: potential applications as immunomodulator drugs. Immunol. Lett. 223, 71–77. doi: 10.1016/j.imlet.2020.04.003
Sanchez, L., Courteaux, B., Hubert, J., Kauffmann, S., Renault, J. H., Clément, C., et al. (2012). Rhamnolipids elicit defense responses and induce disease resistance against biotrophic, hemibiotrophic, and necrotrophic pathogens that require different signaling pathways in Arabidopsis and highlight a central role for salicylic acid. Plant Physiol. 160, 1630–1641. doi: 10.1104/pp.112.201913
Sánchez, M., Aranda, F. J., Teruel, J. A., Espuny, M. J., Marqués, A., Manresa, Á, et al. (2010). Permeabilization of biological and artificial membranes by a bacterial dirhamnolipid produced by Pseudomonas aeruginosa. J. Coll. Interf. Sci. 341, 240–247. doi: 10.1016/j.jcis.2009.09.042
Sánchez, M., Aranda, F. J., Teruel, J. A., and Ortiz, A. (2009). Interaction of a bacterial dirhamnolipid with phosphatidylcholine membranes: a biophysical study. Chemi. Phys. Lipids 161, 51–55. doi: 10.1016/j.chemphyslip.2009.06.145
Sánchez, M., Teruel, J. A., Espuny, M. J., Marqués, A., Aranda, F. J., Manresa, Á, et al. (2006). Modulation of the physical properties of dielaidoylphosphatidylethanolamine membranes by a dirhamnolipid biosurfactant produced by Pseudomonas aeruginosa. Chem. Phys. Lipids 142, 118–127. doi: 10.1016/j.chemphyslip.2006.04.001
Sathi Reddy, K., Yahya Khan, M., Archana, K., Gopal Reddy, M., and Hameeda, B. (2016). Utilization of mango kernel oil for the rhamnolipid production by Pseudomonas aeruginosa DR1 towards its application as biocontrol agent. Bioresour. Technol. 221, 291–299. doi: 10.1016/j.biortech.2016.09.041
Savary, S., Willocquet, L., Pethybridge, S. J., Esker, P., McRoberts, N., and Nelson, A. (2019). The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 3, 430–439. doi: 10.1038/s41559-018-0793-y
Schellenberger, R., Touchard, M., Clement, C., Baillieul, F., Cordelier, S., Crouzet, J., et al. (2019). Apoplastic invasion patterns triggering plant immunity: plasma membrane sensing at the frontline. Mol. Plant Pathol. 20, 1602–1616. doi: 10.1111/mpp.12857
Sen, S., Borah, S. N., Bora, A., and Deka, S. (2017). Production, characterization, and antifungal activity of a biosurfactant produced by Rhodotorula babjevae YS3. Microb. Cell Factor. 16:95. doi: 10.1186/s12934-017-0711-z
Sha, R., Jiang, L., Meng, Q., Zhang, G., and Song, Z. (2012). Producing cell-free culture broth of rhamnolipids as a cost-effective fungicide against plant pathogens. J. Basic Microbiol. 52, 458–466. doi: 10.1002/jobm.201100295
Sha, R., and Meng, Q. (2016). Antifungal activity of rhamnolipids against dimorphic fungi. J. Gen. Appl. Microbiol. 62, 233–239. doi: 10.2323/jgam.2016.04.004
Sharma, A., Jansen, R., Nimtz, M., Johri, B. N., and Wray, V. (2007a). Rhamnolipids from the rhizosphere bacterium Pseudomonas sp. GRP(3) that reduces damping-off disease in Chilli and tomato nurseries. J. Nat. Products 70, 941–947. doi: 10.1021/np0700016
Sharma, A., Wray, V., and Johri, B. N. (2007b). Rhizosphere Pseudomonas sp. strains reduce occurrence of pre- and post-emergence damping-off in chile and tomato in Central Himalayan region. Archiv. Microbiol. 187, 321–335. doi: 10.1007/s00203-006-0197-2
Singh, P., and Cameotra, S. S. (2004). Potential applications of microbial surfactants in biomedical sciences. Trends Biotechnol. 22, 142–146. doi: 10.1016/j.tibtech.2004.01.010
Singh, P., Patil, Y., and Rale, V. (2019). Biosurfactant production: emerging trends and promising strategies. J. Appl. Microbiol. 126, 2–13. doi: 10.1111/jam.14057
Stanghellini, M. E., and Miller, R. M. (1997). Biosurfactants : their identity and potential efficacy in the biological control of zoosporic plant pathogen. Plant Dis. 81, 4–12.
Straight, P. D., Willey, J. M., and Kolter, R. (2006). Interactions between Streptomyces coelicolor and Bacillus subtilis: role of surfactants in raising aerial structures. J. Bacteriol. 188, 4918–4925. doi: 10.1128/jb.00162-06
Syed Ab Rahman, S. F., Singh, E., Pieterse, C. M. J., and Schenk, P. M. (2018). Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 267, 102–111. doi: 10.1016/j.plantsci.2017.11.012
Takemoto, J., Bensaci, M., De Lucca, A., Cleveland, T., Gandhi, N., and Skebba, V. P. (2010). Inhibition of fungi from diseased grapeby syringomycin E-rhamnolipid mixture. Am. J. Enol. Viticu. 61, 120–124.
Tao, Y., Bie, X.-M., Lv, F.-X., Zhao, H.-Z., and Lu, Z.-X. (2011). Antifungal activity and mechanism of fengycin in the presence and absence of commercial surfactin against Rhizopus stolonifer. J. Microbiol. 49, 146–150. doi: 10.1007/s12275-011-0171-9
Tran, H., Ficke, A., Asiimwe, T., Hofte, M., and Raaijmakers, J. M. (2007). Role of the cyclic lipopeptide massetolide A in biological control of Phytophthora infestans and in colonization of tomato plants by Pseudomonas fluorescens. New Phytol. 175, 731–742.
Varnier, A. L., Sanchez, L., Vatsa, P., Boudesocque, L., Garcia-Brugger, A., Rabenoelina, F., et al. (2009). Bacterial rhamnolipids are novel MAMPs conferring resistance to Botrytis cinerea in grapevine. Plant Cell Environ. 32, 178–193. doi: 10.1111/j.1365-3040.2008.01911.x
Vatsa, P., Sanchez, L., Clément, C., Baillieul, F., and Dorey, S. (2010). Rhamnolipid biosurfactants as new players in animal and plant defense against microbes. Intern. J. Mol. Sci. 11, 5095–5108.
Waewthongrak, W., Leelasuphakul, W., and McCollum, G. (2014). Cyclic LIPopeptides from Bacillus subtilis ABS-S14 elicit defense-related gene expression in citrus fruit. PLoS One 9:e109386. doi: 10.1371/journal.pone.0109386
Wise, C., Falardeau, J., Hagberg, I., and Avis, T. J. (2014). Cellular lipid composition affects sensitivity of plant pathogens to fengycin, an antifungal compound produced by Bacillus subtilis strain CU12. Phytopathology 104, 1036–1041. doi: 10.1094/phyto-12-13-0336-r
Wu, L., Huang, Z., Li, X., Ma, L., Gu, Q., Wu, H., et al. (2018). Stomatal closure and SA-, JA/ET-signaling pathways are essential for Bacillus amyloliquefaciens FZB42 to restrict leaf disease caused by Phytophthora nicotianae in Nicotiana benthamiana. Front. Microbiol. 9:847. doi: 10.3389/fmicb.2018.00847
Xiu, P., Liu, R., Zhang, D., and Sun, C. (2017). Pumilacidin-like lipopeptides derived from marine bacterium Bacillus sp. strain 176 suppress the motility of Vibrio alginolyticus. Appl. Environ. Microbiol. 83:e00450-17. doi: 10.1128/aem.00450-17
Yamamoto, S., Shiraishi, S., and Suzuki, S. (2015). Are cyclic lipopeptides produced by Bacillus amyloliquefaciens S13-3 responsible for the plant defence response in strawberry against Colletotrichum gloeosporioides? Lett. Appl. Microbiol. 60, 379–386. doi: 10.1111/lam.12382
Yan, F., Xu, S., Chen, Y., and Zheng, X. (2014). Effect of rhamnolipids on Rhodotorula glutinis biocontrol of Alternaria alternata infection in cherry tomato fruit. Postharvest Biol. Technol. 97, 32–35. doi: 10.1016/j.postharvbio.2014.05.017
Yan, F., Xu, S., Guo, J., Chen, Q., Meng, Q., and Zheng, X. (2015). Biocontrol of post-harvest Alternaria alternata decay of cherry tomatoes with rhamnolipids and possible mechanisms of action. J. Sci. Food Agric. 95, 1469–1474. doi: 10.1002/jsfa.6845
Yoo, D. S., Lee, B. S., and Kim, E. K. (2005). Characteristics of microbial biosurfactant as an antifungal agent against plant pathogenic fungus. J. Microbiol. Biotechnol. 15, 1164–1169.
Yoshida, S., Koitabashi, M., Nakamura, J., Fukuoka, T., Sakai, H., Abe, M., et al. (2015). Effects of biosurfactants, mannosylerythritol lipids, on the hydrophobicity of solid surfaces and infection behaviours of plant pathogenic fungi. J. Appl. Microbiol. 119, 215–224. doi: 10.1111/jam.12832
Zeriouh, H., Romero, D., García-Gutiérrez, L., Cazorla, F. M., de Vicente, A., and Pérez-García, A. (2011). The Iturin-like lipopeptides are essential components in the biological control arsenal of Bacillus subtilis against bacterial diseases of cucurbits. Mol. Plant Microb. Interact. 24, 1540–1552. doi: 10.1094/mpmi-06-11-0162
Keywords: rhamnolipids, lipopeptides, antimicrobial, plant immunity, elicitor, plant pathogen
Citation: Crouzet J, Arguelles-Arias A, Dhondt-Cordelier S, Cordelier S, Pršić J, Hoff G, Mazeyrat-Gourbeyre F, Baillieul F, Clément C, Ongena M and Dorey S (2020) Biosurfactants in Plant Protection Against Diseases: Rhamnolipids and Lipopeptides Case Study. Front. Bioeng. Biotechnol. 8:1014. doi: 10.3389/fbioe.2020.01014
Received: 24 May 2020; Accepted: 03 August 2020;
Published: 08 September 2020.
Edited by:
Gloria Soberón-Chávez, National Autonomous University of Mexico, MexicoReviewed by:
Jonas Contiero, São Paulo State University, Campus Rio Claro, BrazilVivek Sharma, Chandigarh University, India
Copyright © 2020 Crouzet, Arguelles-Arias, Dhondt-Cordelier, Cordelier, Pršić, Hoff, Mazeyrat-Gourbeyre, Baillieul, Clément, Ongena and Dorey. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stéphan Dorey, stephan.dorey@univ-reims.fr
†These authors have contributed equally to this work