Field margins enhance weed seed predation in adjacent fields in early spring
 Sandrine Petit
Sandrine Petit Benjamin Carbonne
Benjamin Carbonne Zoé Etcheverria1
Zoé Etcheverria1  Nathalie Colbach
Nathalie Colbach David Andrew Bohan
David Andrew Bohan- 1Agroécologie, INRAE, Institut Agro, Univ. Bourgogne, Univ. Bourgogne Franche-Comté, Dijon, France
- 2IGEPP, INRAE, Institut Agro, Univ. Rennes, Angers, France
Seed depletion by granivorous organisms can regulate weeds in arable agriculture. Enhancing this regulation can be achieved by adopting farming practices that favour seed predators. Here, we test the hypothesis that the presence of grassy field margins along field edges will increase in-field weed seed predation, in comparison to situations where no grassy field margin is present. Predation cards with Poa annua were exposed in 15 wheat fields in May and June 2018 along 57 transects at distances of 4, 8, 16, 32 meters from the field edge. Cards were either caged (predation by invertebrates) or uncaged predation byall seed predators). We found that in May, the presence of grassy field margins led to higher in-field predation rates at all distances from the field edge, with a very high contribution of invertebrates to seed predation. In June, the presence of grass margin had no impact on in-field seed predation, to which invertebrates and vertebrates contributed equally. This preliminary study provides some support to the hypothesis that grassy field margins augment in-field weed seed predation in early spring. It is plausible that these habitats are emergence sites for invertebrates, with a subsequent high abundance of adults nearby grass margins in early spring, before they disperse more widely across fields and/or switch to alternative prey. These results call for further comparative research on the impact of grass margins on seed predation, seed predators and alternative prey during the whole cropping season.
1 Introduction
In the context of a necessary shift towards pesticide reduction in arable farming, weed management remains a challenging issue. In addition to adopting agronomic practices that limit weed infestations (Deguine et al., 2023), mobilising biotic interactions that exert some control on weeds is a promising avenue (Petit et al., 2018). Among those interactions, weed depletion by seed-eating invertebrates and small mammals is a widespread process (Davis et al., 2011; Kulkarni et al., 2015) and there is increasing empirical evidence that seed consumers significantly regulate weeds in arable fields (Bohan et al., 2011; Blubaugh and Kaplan, 2016; Carbonne et al., 2020). In some instances, seed predators such as field voles (Microtus arvalis) can concomitantly consume crop seeds (Fischer et al., 2018) but we largely ignore how this impacts crop yield (Tschumi et al., 2018). Adopting farming practices that augment the abundance and diversity of seed predators can enhance weed seed predation, e.g. practices of conservation agriculture (Menalled et al., 2007; Petit et al., 2017), organic farming (Diekotter et al., 2016) and crop diversification at the farm scale (Heggenstaller et al., 2006).
The presence of field margins, i.e. linear semi-natural habitats that define the edge of fields (Marshall and Moonen, 2002) can also enhance the diversity and activity of seed-eating organisms. Elements such as grass strips and flower strips around the field can increase the in-field abundance and fitness of seed-eating carabids (Lys and Nentwig, 1992; Labruyere et al., 2016). These habitats indeed provide appropriate and relatively stable conditions during the winter and can increase the survival of some seed predator invertebrates which then recolonize the adjacent crop field during the crop growing season (Wissinger, 1997). Similarly, field margins are stable habitats and key refuges for seed-eating rodents in arable farmland (Broughton et al., 2014) and species such as field voles can colonize adjacent crops from the margins during population increases (Rodriguez-Pastor et al., 2016). Field margins are also a refuge from agricultural disturbances that occur in-field for many vertebrate and invertebrate seed consumers (Landis et al., 2000; Pfiffner and Luka, 2000). Despite the demonstration of a beneficial effect of field margins on predators, their contribution to the natural control of crop pests in the adjacent field is poorly documented and, where it exists, is often highly variable (Holland et al., 2016). In particular, there is, to our knowledge, no study that has directly measured the effects of grassy field margins on in-field weed seed predation. Here we test the hypothesis that in-field weed seed predation will be of higher magnitude in the presence of grassy field margin than when no field margin is present.
2 Methods
2.1 Study area and experimental fields
The study was conducted in spring 2018 in the monitoring site of Fénay, an arable landscape located in the plain south of Dijon in eastern France (47°13’N, 5°03’E). The area is on deep marly soils and exposed to semi-continental climate conditions (annual mean T°=10.7°C, total annual precipitation =744mn). Most fields are cultivated with a 3-year rotation of winter crops (i.e. winter oilseed rape or mustard/winter wheat/winter barley). We selected 15 winter wheat fields that were representative of the mainstream farming management in the study area (Yvoz et al., 2020). All fields were either superficially tilled or ploughed every year of the crop rotation. Pesticide use in 2018 was quite similar across fields. Supplementary Material Table 1 provides a description of the farming management of each field.
2.2 Measuring seed predation
We placed up to four transects per field, perpendicular to each edge of the field and ended up with 25 transects starting from a grassy field margin and 32 starting at the boundary with another arable field, with no spontaneous vegetation. Weed seed predation measures on each transect were conducted at four distances, i.e. at 4, 8, 16 and 32 m from the field border. The maximal distance was derived from previous research showing that the vegetation found at 30m within an arable field is representative of the vegetation found further away from the field margin (Firbank et al., 2003).
We used weed seed cards with fifty Poa annua seeds, a common weed across Europe. Seeds were glued with 3M repositionable glue to ca. 95 x 40 mm card of brown sand paper (grain size 60). At each location along the transect (plot level), we exposed one seed card enclosed in a mesh cage (1 cm² wire mesh) to exclude vertebrates (caged modality) and one seed card with no cage, thus available to all types of seed consumers (uncaged modality). The cards were exposed over 7 days and we conducted two sessions of measures, one at the ‘mid-season’ of cereal growing (early May) and one ‘just prior to harvest’ (early June). Weather conditions during a session were considered similar across the 15 fields (same landscape and same dates) but slightly differed between the two sessions (see Supplementary Material Table 2). Seed predation rates per week were estimated at the plot level using the number of seeds removed from the seed cards. In addition, we estimatedthe contribution of vertebrates to seed predation, i.e. uncaged predation minus caged predation (see Westerman et al., 2003). Such estimation assumes that predation rates by different predators are additive, an assumption that is only valid at a broad level (e.g. field level) to ensure that it is based on a large number of seed cards. Here, we estimated the contribution of vertebrates on four subsets of the data, i.e. measures in May along grass margins, in May in the absence of grass margin, in June along grass margin and in June in the absence of a grass margin.
2.3 Statistical analyses
All analyses and figures were conducted in R version 4.2.1 (Core Team, 2022). We used Generalised Linear Mixed Models (GLMM), with the binomial (link=“logit”) family to assess the effect of the presence/absence of field margin in interaction with the distance to the field border (4, 8, 16 and 32m), the tillage regime (superficial tillage, ploughing) and the sampling session (early May, early June) on the caged and uncaged weed seed predation rates. We thus compared two treatment contrasts: grassy margin (n = 200 obs. i.e. 25 transects * 4 distances * 2 sessions) and no margin (n = 256).
We analysed separately caged and uncaged weed seed predation rates. The random structure of all models includes the transect identity (1-4) nested within the field identity (1-15) to account for the non-independence of points belonging to the same field or transect. We also included an observation-level random effect (1-456) to correct model overdispersion. Model selection was performed based on the AICc and we kept the most parsimonious model with the lowest AICc. We used packages lme4 (procedures ‘glmer’; Bates et al., 2015); MuMIn (‘dredge’; Bartoń, 2022), ggplot2 (Wickham, 2016) and interactions for the figures (Long, 2019) The model residuals were visually checked for normality and homoscedasticity using the package DHARMa (Hartig, 2022). Ther was no multicollinearity between explanatory variables as all Variation Inflation Factors (VIFs) values were below 3. The P-values and model parameters were recovered from the ‘Anova’ procedure (package ‘car’) using a type = “III” analysis-of-variance tables with Wald chi-square. Finally, for significant effects, paired comparisons were performed using the estimated marginal means with the package emmeans (Lenth, 2020).
3 Results
Predation rates were on average 9.3% caged and 15.1% uncaged and were higher in May than in June (Figure 1; Supplementary Material Table 3). The contribution of vertebrates to Poa annua seed predation in May was 20% in the presence of grass margins and 42% in the absence of grass margin. This contribution was higher in June, respectively 47% in the presence of grass margins and 59% in the absence of grass margins.
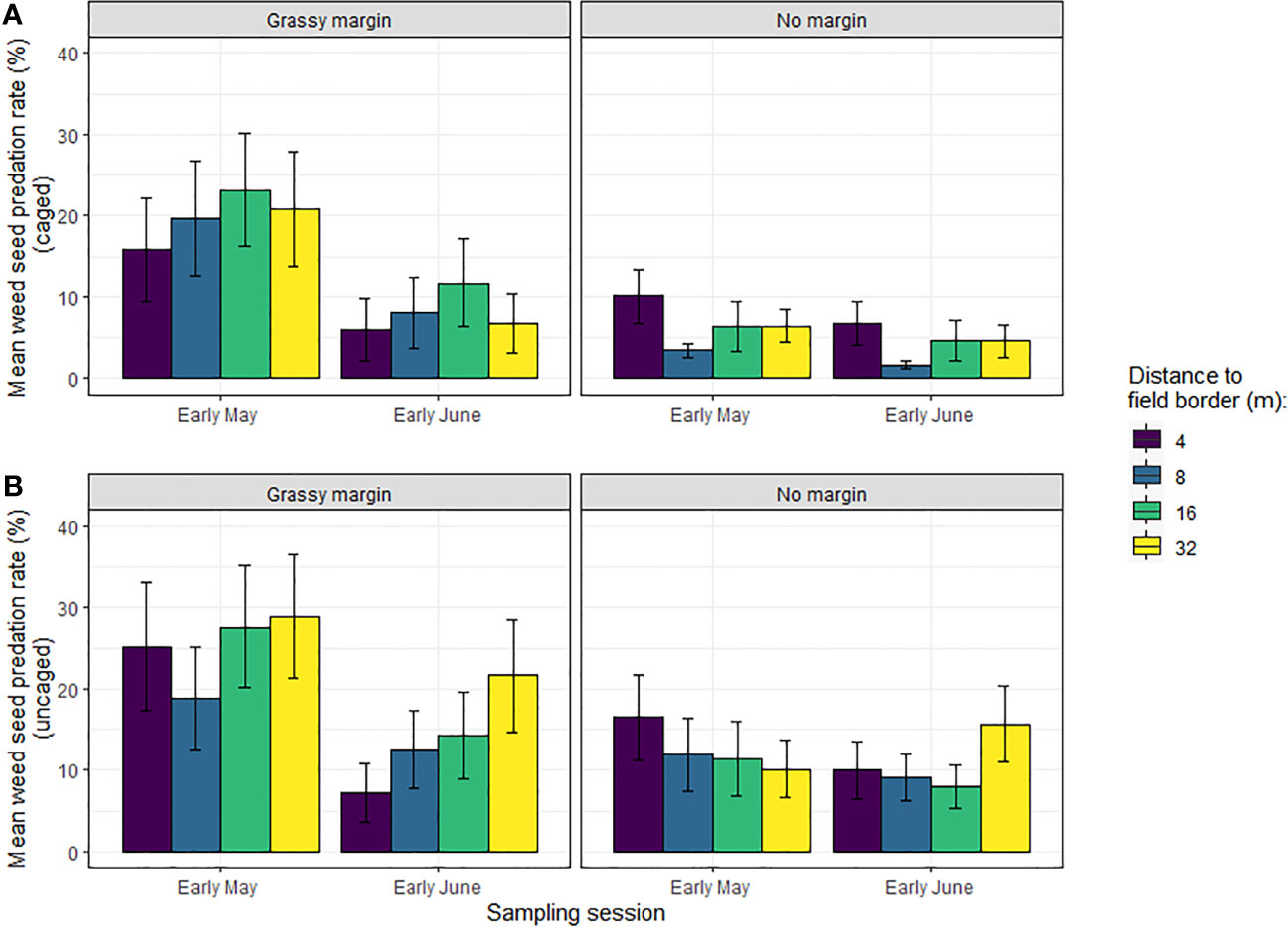
Figure 1 Barplot representation of the mean (+/- SE) of (A) caged and (B) uncaged weed seed predation rates of Poa annua in winter wheat in Burgundy, according to field margin (presence/absence), distance to the field edge and sampling session. Cages were used to exclude vertebrate seed predators.
We found a significant interaction effect between field margin (presence/absence) and sampling session on weed seed predation with cages and without cages (Table 1). There was a consistent positive effect of the presence of a grassy field margin on seed predation rates, whether in caged or uncaged situations. This effect was only significant during the first session in early May (Figure 2; Table 2), however. The distance to the field edge had no impact on seed predation rates. Similarly, we detected no effect of the tillage regime on in-field seed predation rates (Supplementary Material Figure 1).
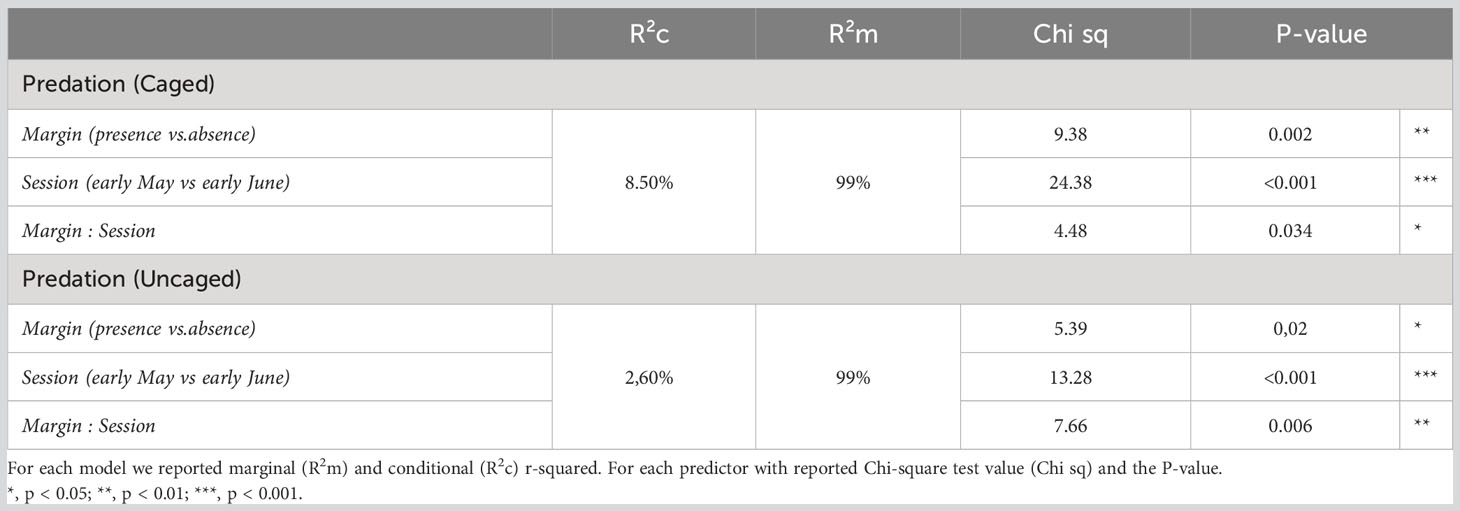
Table 1 Parameter estimates from the two Generalised Linear Mixed Models for weed seed predation of Poa annua caged and uncaged.
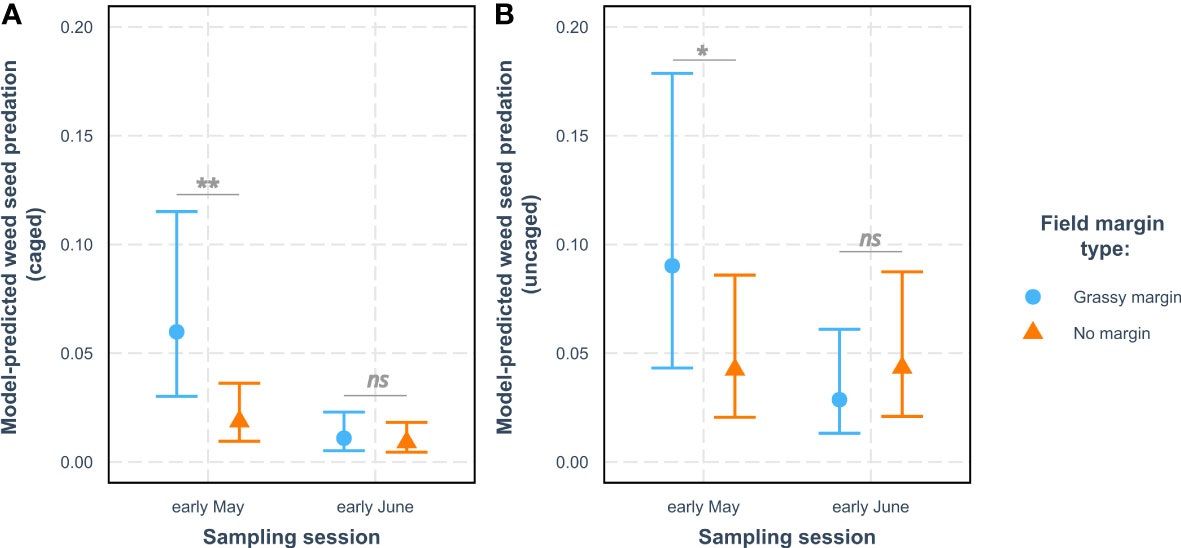
Figure 2 Mean model prediction (+/- SE) for weed seed predation rates of Poa annua in winter wheat in Burgundy (A) caged and (B) uncaged according to field margin type and sampling session. For each sampling session, the level of significance of the differences in predation between grassy margin and no margin is indicated above the horizontal bars (ns, non-significant; *, p < 0.05 and **, p < 0.01 – see Table 2).
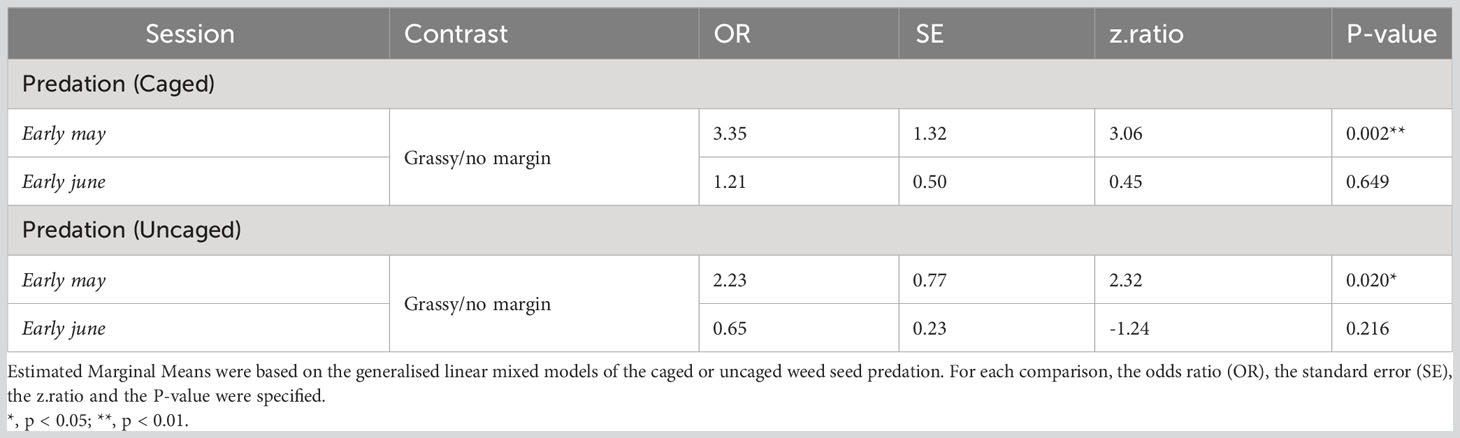
Table 2 Results of the Estimated Marginal Means pairwise comparison of weed seed predation of Poa annua in winter wheat in Burgundy associated to grassy margin versus no margin at each sampling session (early May and early June).
4 Discussion
These first results suggest that the presence of a grass margin leads to an overall increase of weed seed predation of Poa annua seeds in the adjacent cereal field during the wheat growing season, irrespective of the distance to the grass margin, which gives support to our initial hypothesis. This effect was however only detected during the wheat growing season in very early spring (May) and was no longer significant in June, just prior to wheat harvest. We detected no significant effect of the distance to the field edge on weed seed predation rates, which corroborates previous studies (Marino et al., 1997; Tooley et al., 1999; Alignier et al., 2008). Here, the maximum distance between boundary and field centre was 32 meters and it has been shown that many carabid species penetrated into the field up to 60 meters (Holland et al., 1999) whereas small rodents are even more mobile. Therefore, the largest distance in the present experiment may still be in a range of dispersal from the boundary for many species.
Overall, we observed that seed predation along field margins was particularly high in May and was mostly due to invertebrates whereas in the absence of field margins, predation rates were lower and resulted equally from the activity of both invertebrates and vertebrates. It is thus plausible that the high predation rates observed in May along grassy margins results from a positive effect of these elements on the distribution and/or foraging activity of invertebrate seed predators (mostly carabids here, Carbonne et al., 2020). This explanation would be in line with the fact that several carabid species prefer to overwinter in grassy field boundaries (Sotherton, 1985; Saska et al., 2007; Holland et al., 2009) so that the presence of such habitats enhance the populations of those species. This positive effect of grass margins on seed predation was no longer detected in early June and vertebrates and invertebrates equally contributed to seed removal. One possible explanation is that seed-eating invertebrates have spread and redistributed over the whole field in June, as already shown for carabids (Holland et al., 2009). The magnitude and timing of this redistribution depends on the probability of an individual leaving a field margin, which is notably driven by the relative availability of food resources between the grassy habitat and the adjacent crop as well as on carabid hunger level, as shown for the abundant carabid predator P. melanarius (Fournier and Loreau, 2002). It is also possible that the availability of alternative prey increased in June and that carabids have switched to alternative prey such as aphids and collembola at that time, as established in previous studies (Gray et al., 2021). Identifying if one of the above -mentioned processes, or both, are at play would require additional studies to monitor the distribution of seed predators, seed predation and alternative prey over time and over larger areas in each field, for example using a grid sampling design (e.g. see Trichard et al., 2014). Our results also suggest that in the experimental fields we surveyed, the presence of grassy margins did not favour the contribution of small mammals to weed seed predation in the adjacent field during spring. The spillover of small mammals into the adjacent fields is complex as it results from multiple interacting factors, such as the peak phase, the vegetation structure and cover of the grassy field margin and the identity and cover/height of the adjacent crop (Rodriguez-Pastor et al., 2016). Understanding the seasonal variation in the role of grassy margins to vertebrates and their contribution in-field seed predation in our study area would require additional studies.
5 Conclusion
This preliminary study suggests that grassy field margins can enhance weed seed predation in early spring and do not necessarily lead to increased seed predation by small mammals. Further studies are necessary to get a full understanding of the role of grassy field margins on in-field seed predators and alternative prey and how this varies over the course of the cropping season. This is a pre-requisite to assess if the establishment of grassy margins could be an effective way to enhance weed seed predation in arable farming.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
SP, DB, and BC designed the empirical design and BC collected data. BC, ZE, NC, and SP analysed the data. SP led the writing of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by the COPRAA project, funded by the French Office for Biodiversity (OFB) as part of the Ecophyto call on “Global approaches to limit the use of phytopharmaceuticals”. Field data was collected as part of the ERA-NET C-IPM project BioAWARE.
Acknowledgments
We are particularly grateful to Luc Biju-Duval, Chantal Ducourtieux, Claude Sarrasin and Britta Frei for their work in the field.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2023.1228395/full#supplementary-material
Supplementary Figure 1 | Barplot representation of the mean (+/- SE) of the (A) caged and (B) uncaged weed seed predation rates according to field margin type, distance to the field margin and tillage regime.
References
Alignier A., Meiss H., Petit S., Reboud X. (2008). Variation of post-dispersal weed seed predation according to weed species, space and time. J. Plant Dis. Prot. 21, 221–226.
Bartoń K. (2022). MuMIn: Multi-Model Inference. R package version 1.47. CRAN. Available at: https://CRAN.R-project.org/package=MuMIn.
Bates D., Mächler M., Bolker B., Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Software 67 (1), 1–48. doi: 10.18637/jss.v067.i01
Blubaugh C. K., Kaplan I. (2016). Invertebrate seed predators reduce weed emergence following seed rain. Weed Sci. 64, 80–86. doi: 10.1614/WS-D-15-00111.1
Bohan D. A., Boursault A., Brooks D., Petit S. (2011). National-scale regulation of the weed seedbank by carabid predators. J. Appl. Ecol. 48, 388–398. doi: 10.1111/j.1365-2664.2011.02008.x
Broughton R. K., Shore R., Heard M. S., Amy S. R., Meek W. R., Redhead J. W., et al. (2014). Agri-environment scheme enhances small mammal diversity and abundance at the farm-scale. Agric. Ecosyst. Environ. 192, 122–129. doi: 10.1016/j.agee.2014.04.009
Carbonne B., Petit S., Neidel V., Foffova H., Daouti E., Skuhrovec J., et al. (2020). The resilience of weed seedbank regulation by carabid beetles, at continental scales, to alternative prey. Sci. Rep. 10, 19315. doi: 10.1038/s41598-020-76305-w
Core Team R. (2022). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing).
Davis A. S., Daedlow D., Schutte B. J., Westerman P. R. (2011). Temporal scaling of episodic point estimates of seed predation to long-term predation rates. Methods Ecol. Evol. 2, 682–890. doi: 10.1111/j.2041-210X.2011.00119.x
Deguine J. P., Aubertot J. N., Bellon S., Côte F., Lauri P. E., Lescourret F., et al. (2023). Agroecological crop protection for sustainable agriculture. Adv. Agron. 178, 1–59. doi: 10.1016/bs.agron.2022.11.002
Diekotter T., Wamser S., Dörner T., Wolters V., Birkhofer K. (2016). Organic farming affects the potential of a granivorous carabid beetle to control arable weeds at local and landscape scales. Agr For. Entomol 18, 167–173. doi: 10.1111/afe.12150
Firbank L.G., Heard M.S., Woiwod I.P., Hawes C., Haughton A.J., Champion G.T., et al. (2013). An introduction to the farm-scale evaluations of genetically modified herbicide-tolerant crops. J. Appl. Ecol. 40, 2–16. doi: 10.1046/j.1365-2664.2003.00787.x
Fischer C., Gayer C., Kurucz K., Riesch F., Tscharntke T., Batáry P. (2018). Ecosystem services and disservices provided by small rodents in arable fields: Effects of local and landscape management. J. Appl. Ecol. 55, 548–558. doi: 10.1111/1365-2664.13016
Fournier E., Loreau M. (2002). Foraging activity of the carabid beetle Pterostichus melanarius Ill. Field margin habitats. Agric. Ecosyst. Environ. 89, 253–259. doi: 10.1016/S0167-8809(01)00216-X
Gray C., Ma A., McLaughlin O., Petit S., Woodward G., Bohan D. A. (2021). Ecological plasticity governs ecosystem services in multilayer networks. Commun. Biol. 4, 75. doi: 10.1038/s42003-020-01547-3
Hartig F. (2022). DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R package version 0.4.5. Available at: https://CRAN.R-project.org/package=DHARMa.
Heggenstaller A., Menalled F. D., Liebman M., Westerman P. R. (2006). Seasonal patterns in post-dispersal seed predation of Abutilon theophrasti and Setaria faberi in three cropping systems. J. Appl. Ecol. 43, 999–1010. doi: 10.1111/j.1365-2664.2006.01198.x
Holland J. M., Bianchi F. J., Entling M. H., Moonen A.-C., Smith B. M., Jeanneret P. (2016). Structure, function and management of seminatural habitats for conservation biological control: a review of European studies. Pest Manage. Sci. 72, 1638–1651. doi: 10.1002/ps.4318
Holland J. M., Birkett T., Southway S. (2009). Contrasting the farm-scale spatio-temporal dynamics of boundary and field overwintering predatory beetles in arable crops. BioControl 54, 19–33 (). doi: 10.1007/s10526-008-9152-2
Holland J. M., Perry J. N., Winder L. (1999). The within-field spatial and temporal distribution of arthropods in winter wheat. Bull. Entomological Res. 89, 499–513. doi: 10.1017/S0007485399000656
Kulkarni S. S., Dosdall L. M., Willenborg C. J. (2015). The role of ground beetles (Coleoptera: carabidae) in weed seed consumption: A review. Weed Sci. 63, 355–376. doi: 10.1614/WS-D-14-00067.1
Labruyere S., Ricci B., Lubac A., Petit S. (2016). Crop type, crop management and grass margins affect the abundance and the nutritional state of seed eating carabid species in arable landscapes. Agric. Ecosyst. Environ. 231, 183–192. doi: 10.1016/j.agee.2016.06.037
Landis D. A., Wratten S. D., Gurr G. M. (2000). Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol 4, 175–201. doi: 10.1146/annurev.ento.45.1.175
Lenth R. (2020). emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.4.7. Available at: https://cran.r-project.org/package=emmeans.
Long J. A. (2019). interactions: Comprehensive, User-Friendly Toolkit for Probing Interactions. R package version 1.1.0. Available at: https://cran.r-project.org/package=interactions.
Lys J. A., Nentwig W. (1992). Augmentation of beneficial arthropods by strip-management 4. Surface activity, movements and activity density of abundant carabid beetles in a cereal field. Oecologia 92, 373–382. doi: 10.1007/BF00317463
Marino P. C., Gross K. L., Landis D. A. (1997). Weed seed loss due to predation in a Michigan maize fields. Agric. Ecosyst. Environ. 66, 189–196. doi: 10.1016/S0167-8809(97)00091-1
Marshall E. J. P., Moonen A. C. (2002). Field margins in northern Europe: their functions and interactions with agriculture. Agric. Ecosyst. Environ. 89, 5–21. doi: 10.1016/S0167-8809(01)00315-2
Menalled F. D., Smith R. G., Dauer J. T., Fox T. B. (2007). Impact of agricultural management on carabid communities and weed seed predation. Agric. Ecosyst. Environ. 118, 49–54. doi: 10.1016/j.agee.2006.04.011
Petit S., Cordeau S., Chauvel B., Bohan D., Guillemin J.-P., Steinberg C. (2018). Biodiversity-based options for arable weed management. A review. Agron. Sustain. Dev. 38, 48. doi: 10.1007/s13593-018-0525-3
Petit S., Trichard A., Biju-Duval L., McLaughlin O., Bohan D. (2017). Interactions between conservation agricultural practice and landscape composition promote weed seed predation by invertebrates. Agric. Ecosyst. Environ. 240, 45–53. doi: 10.1016/Fj.agee.2017.02.014
Pfiffner L., Luka H. (2000). Overwintering of arthropods in soils of arable fields and adjacent semi-natural habitats. Agric. Ecosyst. Environ. 78, 215–222. doi: 10.1016/S0167-8809(99)00130-9
Rodriguez-Pastor R., Luque-Larena J. J., Lambin X., Mougeot F. (2016). Living on the edge: The role of field margins for common vole (Microtus arvalis) populations in recently colonised Mediterranean farmland. Agric. Ecosyst. Environ. 231, 206–217. doi: 10.1016/j.agee.2016.06.041
Saska P., Vodde M., Heijerman T., Westerman P., van der Werf W. (2007). The significance of a grassy field boundary for the spatial distribution of carabids within two cereal fields. Agric. Ecosyst. Environ. 122, 427–434. doi: 10.1016/j.agee.2007.02.013
Sotherton N. W. (1985). The distribution and abundance of predatory Coleoptera overwintering in field boundaries. Ann. Appl. Biol. 106, 17–21. doi: 10.1111/j.1744-7348.1985.tb03089.x
Tooley J. A., Froud-Williams R. J., Boatman N. D., Holland J. M. (1999). Weed seed predation in arable field margins by carabid beetles. Aspects Appl. Biol. 54, 211–216.
Trichard A., Ricci B., Ducourtieux C., Petit S. (2014). The spatio-temporal distribution of weed seed predation differs between conservation agriculture and conventional tillage. Agric. Ecosyst. Environ. 180, 40–47. doi: 10.1016/Fj.agee.2014.01.031
Tschumi M., Ekroos J., Hjort C., Smith H. G., Birkhofer K. (2018). Predation-mediated ecosystem services and disservices in agricultural landscapes. Ecol. Appl. 28, 2109–2118. doi: 10.1002/eap.1799
Westerman P. R., Hofman A., van der Werf W. (2003). Relative importance of vertebrates and invertebrates in epigeaic weed seed predation in organic cereal fields. Agric. Ecosyst. Environ. 95, 417–425. doi: 10.1016/S0167-8809(02)00224-4
Wickham H. (2016). ggplot2: elegant graphics for data analysis (New York: Springer-Verlag). Available at: https://ggplot2.tidyverse.org, ISBN: ISBN 978-3-319-24277-4.
Wissinger S. A. (1997). Cyclic colonization in predictably ephemeral habitats: a template for biological control in annual crop systems. Biol. Control 10, 4–15. 29. doi: 10.1006/bcon.1997.0543
Keywords: biological control, grass margins, seed predators, tillage regime, weed control
Citation: Petit S, Carbonne B, Etcheverria Z, Colbach N and Bohan DA (2023) Field margins enhance weed seed predation in adjacent fields in early spring. Front. Agron. 5:1228395. doi: 10.3389/fagro.2023.1228395
Received: 24 May 2023; Accepted: 11 August 2023;
Published: 05 September 2023.
Edited by:
Giovanni Bubici, Consiglio Nazionale delle Ricerche (CNR), ItalyReviewed by:
Euro Pannacci, University of Perugia, ItalyRajiv Kumar Srivastava, University of Hohenheim, Germany
Copyright © 2023 Petit, Carbonne, Etcheverria, Colbach and Bohan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sandrine Petit, sandrine.petit-michaut@inrae.fr