
More Information
Submitted: November 04, 2022 | Approved: November 09, 2022 | Published: November 10, 2022
How to cite this article: Aarrouf J, Hdech DB, Diot A, Bornard I, Félicie L, et al. Flashes of UV-C light are perceived by UVR8, the photoreceptor of UV-B light. J Plant Sci Phytopathol. 2022; 6: 151-153.
DOI: 10.29328/journal.jpsp.1001089
Copyright License: © 2022 Aarrouf J, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.

Flashes of UV-C light are perceived by UVR8, the photoreceptor of UV-B light
Jawad Aarrouf1*, Douae Ben Hdech1, Alice Diot1, Isabelle Bornard2, Lauri Félicie1 and Laurent Urban1*
1UPRI/ERIT PSII, Avignon University, France
2Plant Pathology Unit, INRAE Avignon, France
*Address for Correspondence: Jawad Aarrouf, UPRI/ERIT PSII, Avignon University, France, Email: jawad.aarrouf@univ-avignon.fr
Laurent Urban, UPRI/ERIT PSII, Avignon University, France, Email: laurent.urban@univ-avignon.fr
Light is an important regulator of plant morphogenesis and plant-pathogen interactions via specific photoreceptors and signaling pathways. Besides visible light, other electromagnetic radiations may play roles, notably ultraviolet (UV) light. The UV part of the electromagnetic spectrum includes UV-A (315 nm - 400 nm), UV-B (280 nm - 315 nm) and UV-C radiations (200 nm - 280 nm). UV-B and UV-C have been reported to increase plant resistance to plant pathogens after the UV perception and signaling stages. The perception of UV-B radiation is achieved by the dimer protein UVR8 (UV RESISTANCE LOCUS 8). Even though the action spectrum of this photoreceptor overlaps in the UV-C domain, it has never been formally demonstrated that UVR8 could also act as a photoreceptor of UV-C light. We provide here original observations showing that UVR8 can indeed perceive UV-C light provided that the latter is in the form of flashes (1s) and not continuous illuminations (the 60s). Our observations also show that the response of UVR8 to flashes of UV-C light is dose-dependent. They could explain why flashes of UV-C light are more effective for stimulating plant defenses than continuous illuminations for the same amount of energy delivered to plants (J/m2). Eventually, our observations support ongoing trials that aim at using UV-C light as an environmental-friendly plant resistance inducer in field conditions.
In addition to visible light, ultraviolet light (UV) has specific properties that are exploited in agriculture and the food industry. There is notably growing evidence that UV-B and UV-C light can be used to stimulate plant defenses [1-6]. Surprisingly, Aarrouf and Urban [7] found both in lab and greenhouse conditions that flashes of UV-C light of less than 2s stimulate plant defenses against fungal diseases substantially better than conventional (the 60s) exposures, for the same amount of cumulated energy. Moreover, Ledermann, et al. [8] observed that UV-C light flashes can be used to boost plant defenses against Erysiphe necator under vineyard conditions and, therefore, help reduce fungicide use. However, further research is needed to improve our understanding of the mechanisms of perception and defense stimulation by UV-C light flashes. One objective is to identify markers of UV-C light perception that could be used to maximize the effectiveness of UV-C light treatments in the field. UVR8, the photoreceptor of UV-B light, is a good candidate since its action spectrum overlaps in the UV-C domain [9]. Absorption of UV-B light by tryptophane dissociates the UVR8 dimer into monomers that migrate into the nucleus, initiating signal transduction through interaction with CONSTITUTIVELY PHOTOMORPHOGENIC1 (COP1). It is believed that this interaction orchestrates the expression of more than 100 genes [10]. Here we present preliminary observations made by using confocal microscopy, showing that UV-C light, notably under the form of flashes, is perceived by UVR8.
The cellular localization of UVR8 was studied by confocal microscopic, using transgenic plants of Arabidopsis thaliana UVR8pro: GFP-UVR8 expressing the protein fused to the Green Fluorescent Protein (GFP) [10]. The DAPI (4',6-diamidino-2-phenylindole, dihydrochloride) staining technique was used to visualize DNA in cells [10,11]. Plants were grown for 5 weeks in a phytotron at constant conditions (daytime fluence of 50 µmol photons/m2/s, photoperiod of 12h/12h, day/night temperatures of 21 °C/20 °C). After a night in the dark, the plants were treated with UV-B and UV-C light, the former under the form of either flash (1s) or continuous exposures (the 60s). Observations were made using a confocal microscope (Leica TCS SP8, Leica Microsystems GmbH, Wetzlar, Germany). Leaf samples of 1 cm2 were cut with a scalpel and then placed between the slide and coverslip in a drop of DAPI. After 5 minutes, cells of the upper epidermis were observed. UV-B fluorescent tubes (Viber, model T-15M OC, https://www.vilber.com/uv-tubes) and UV-C fluorescent tubes Philips, model TUV PL-L 95W HO/4P) were used for light treatments. The emission spectra of the sources were designed with the Iliad software (Spectroradiometer Hera compact, Admesy b.v., Ittervoort, The Netherlands) (Figure 1).
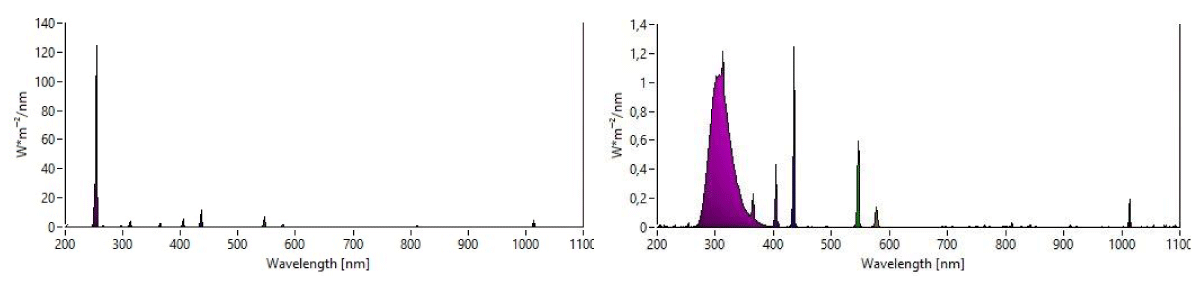
Figure 1: Emission spectraof UV-C (Peak wavelength: 254 nm) and UV-B (Peak wavelength: 312 nm) fluorescent tube.
The two doses of flashes of UV-C light (200 and 400 J/m2) were obtained by modifying the distance of lamps to plants.
For plants treated with UV-B light, the fluorescence of GFP in the nucleus of epidermal cells can be observed (Figure 2).
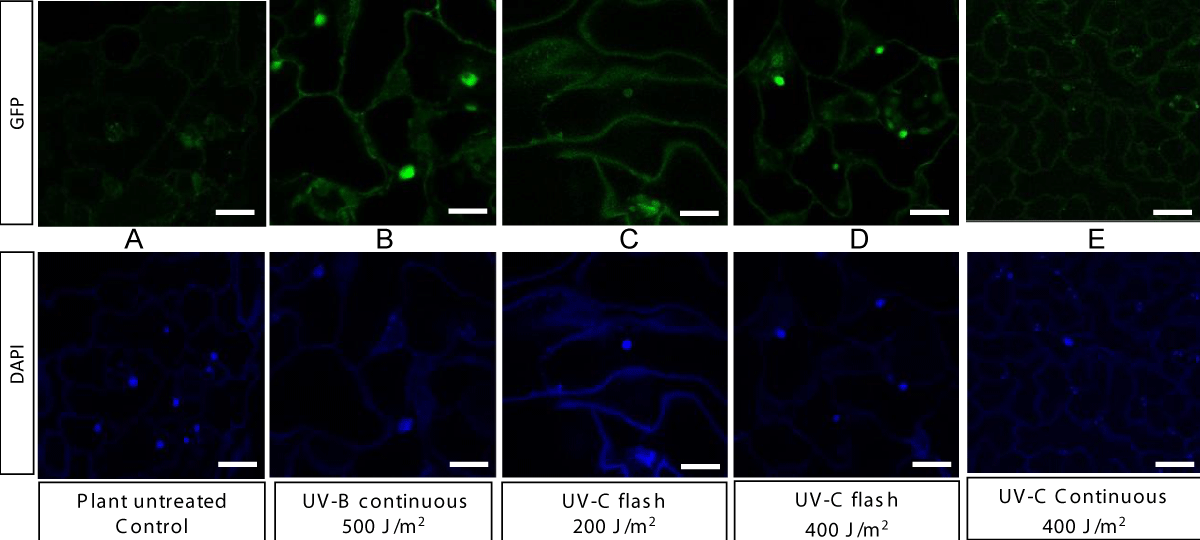
Figure 2: Confocal images of DAPI (excitation of 405 nm and emission range o f412-483 nm) and GFP (excitation of 488 nm and emission range of 498–5420 nm) fluorescence in leaf epidermal tissue of 35-day old UVR8proGFP-UVR8 transgenic Arabidopsis untreated (A) or exposed to UV-B continuous with 500 J/m2 (B); to UV-C flash with 200 J/m2 (C), to UV-C flash with 400 J/m2 (D), to UV-C continuous with 400J/m2 (E). Bar = 20 μm.
The colocalization of GFP and DAPI in confocal images confirms the accumulation of the UVR8 protein in the nucleus after plant treatment. Plants treated with UV-C flash at 200 J/m2 also shows a slight GFP fluorescence in the nucleus, which becomes much more marked with the 400 J/m2 doses (Figure 2). Kaiserli and Jenkins [10,11] similarly found that doses of UV-B light matter for the response of UVR8. Here we provide evidence for the first time that the UVR8 is a UV-C light receptor. The absence of response with the 400 J/m2 doses under the form of continuous illumination, suggests moreover that flashes are essential for penetration and effect and explains why flashes have superior effects than continuous exposures for the same amount of cumulated energy [7].
Our observations give support to the idea that UV-C light needs to be delivered in the form of flashes for efficient perception by UVR8. Further research will need to be conducted with the objective of understanding the role of UVR8 in the stimulation of the signaling and metabolic pathways involved in plant defense. Besides, it cannot be out ruled that other mechanisms of light perception are involved in the effects of flashes of UV-C light. At any rate, flashes of UV-C light emerge as a promising technique of plant treatment, well-suited for field use since light flashes can be supplied by lamps attached to tractors moving at a normal speed between rows and they moreover do not leave any residues on crops.
This study was conducted with the financial support of the French Ministry of Agriculture and Food as part of the CASDAR project OïdUV. The confocal microscope was funded by the European Regional Development Fund, the French government, the Sud Provence-Alpes-Côte d’Azur Region, the Departmental Council of Vaucluse and the Urban Community of Avignon. Transgenic seeds of A. thaliana were donated by Gareth Jenkins and Eirini Kaiserli (University of Glasgow, UK).
- Charles MT, Mercier J, Makhlouf J, and Arul J. Physiological basis of UV-C-induced resistance to Botrytis cinerea in tomato fruit: I. Role of pre- and post-challenge accumulation of the phytoalexin-rishitin. Postharvest Biol. Technol. 2008; 47: 10-20.
- Ouhibi C, Attia H, Nicot P, Lecompte F, Vidal V, Lachâal M, Urban L, and Aarrouf J. Effects of nitrogen supply and of UV-C irradiation on the susceptibility of Lactuca sativa L to Botrytis cinerea and Sclerotinia minor. Plant Soil. 2015; 393: 35-46.
- Urban L, Charles F, de Miranda MRA, Aarrouf J. Understanding the physiological effects of UV-C light and exploiting its agronomic potential before and after harvest. Plant Physiol Biochem. 2016 Aug;105:1-11. doi: 10.1016/j.plaphy.2016.04.004. Epub 2016 Apr 4. PMID: 27064192.
- Forges M, Vàsquez H, Charles F, Sari, DC, Urban L, Lizzi Y, Bardin M, Aarrouf J. Impact of UV-C radiation on the sensitivity of three strawberry plant cultivars [Fragaria x ananassa) against Botrytis cinerea. Sci. Hortic. 2018; 240: 603-613.
- Vàsquez H, Ouhibi C, Forges M, Lizzi Y , Urban L, Aarrouf J. Hormetic doses of UV-C light decrease the susceptibility of tomato plants to Botrytis cinerea infection. J. Phytopathol. 2020; 168: 524-532.
- Vanhaelewyn L, Van Der Straeten D, De Coninck B, Vandenbussche F. Ultraviolet Radiation From a Plant Perspective: The Plant-Microorganism Context. Front Plant Sci. 2020 Dec 15;11:597642. doi: 10.3389/fpls.2020.597642. PMID: 33384704; PMCID: PMC7769811.
- Aarrouf J, Urban L. Flashes of UV-C light: An innovative method for stimulating plant defences. PLoS One. 2020 Jul 9;15[7):e0235918. doi: 10.1371/journal.pone.0235918. PMID: 32645090; PMCID: PMC7347194.
- Ledermann L, Daouda S, Gouttesoulard C, Aarrouf J, Urban L. Flashes of UV-C Light Stimulate Defenses of Vitis vinifera L. 'Chardonnay' Against Erysiphe necator in Greenhouse and Vineyard Conditions. Plant Dis. 2021 Aug;105[8):2106-2113. doi: 10.1094/PDIS-10-20-2229-RE. Epub 2021 Sep 15. PMID: 33393363.
- Jiang L, Wang Y, Olof Björn L, He JX, Li S. Sensing of UV-B radiation by plants. Plant Signal Behav. 2012 Aug;7[8):999-1003. doi: 10.4161/psb.20815. Epub 2012 Jul 3. PMID: 22751358; PMCID: PMC3474703.
- Kaiserli E, Jenkins GI. UV-B promotes rapid nuclear translocation of the Arabidopsis UV-B specific signaling component UVR8 and activates its function in the nucleus. Plant Cell. 2007 Aug;19[8):2662-73. doi: 10.1105/tpc.107.053330. Epub 2007 Aug 24. PMID: 17720867; PMCID: PMC2002606.
- Kapuscinski J. DAPI: a DNA-specific fluorescent probe. Biotech Histochem. 1995 Sep;70[5):220-33. doi: 10.3109/10520299509108199. PMID: 8580206.