


Abstract
The cytochrome P450 3A subfamily plays vital roles in the metabolism of endogenous chemicals and xenobiotics. Understanding the basal expression of CYP3A in humans and pigs is crucial for drug evaluation. In this study, we demonstrated that the basal transcriptional regulation of CYP3A genes in hepatocytes is evolutionarily conserved between humans and pigs. The basal expression of CYP3A genes is transactivated by two cis-acting elements, the CCAAT and GC boxes, located a constant distance apart in the proximal promoter region of six CYP3A genes. Mutation analysis of these two cis-acting elements suggested that they play important roles in mediating basal expression, but to different extents because of the nucleotide variations in the elements. Two transcription factors, nuclear transcription factor Y (NF-Y) and specificity protein 1 (Sp1), directly bind to these cis-acting elements in CYP3A proximal promoters in HepG2 cells and porcine hepatocytes. Furthermore, changing the distance between the NF-Y and Sp1 binding sites resulted in decreases in the promoter activity of CYP3A genes. Conclusively, our results show that human and porcine CYP3A genes are regulated by NF-Y and Sp1 in a coordinated manner, and that the distance between these two cis-acting elements is crucial for constitutive CYP3A expression.
Introduction
Cytochrome P450s belong to a heme monooxygenase superfamily named according to their characteristic absorption at 450 nm (Omura and Sato, 1962). Cytochrome P450 genes are categorized into different families and subfamilies based on the divergent evolution and amino acid sequence homology of this superfamily (Nebert et al., 1991). Cytochrome P450 family members 1–4 play crucial roles in the metabolism of endogenous substances, including sterols, fatty acids, and vitamins, and the biotransformation of exogenous compounds, including drugs, environmental pollutants, and toxins (Isin and Guengerich, 2007; Zanger and Schwab, 2013). In particular, approximately 50%–60% of known clinical drugs can be metabolized by the human CYP 3A4 enzyme (Li et al., 1995; Kumar and Surapaneni, 2001).
CYP3A4 and CYP3A5 are mainly expressed in the liver and gastrointestinal tract (Kivistö et al., 1996; Guengerich, 1999). In the liver and hepatocytes, the basal expression of CYP3A4 is regulated by many transcription factors, such as the binding of CCAAT enhancer binding protein alpha, hepatocyte nuclear factor 3, and pregnane X receptor to the proximal promoter and distal regions of CYP3A4 (Rodríguez-Antona et al., 2003; Bombail et al., 2004; Matsumura et al., 2004). Crosstalk between transcription factors has been extensively studied. It has been reported that the interaction of cell cycle and apoptosis regulator 1 with the constitutive androstane receptor enhances the expression of constitutive androstane receptor–induced genes, such as CYP3A4 (Kanno et al., 2019). The proximal pregnane X receptor, constitutive androstane receptor, nuclear transcription factor (NF-Y), specificity protein 1 (Sp1), and specificity protein 3 cooperatively regulate the basal transcription of CYP3A5 (Iwano et al., 2001; Burk et al., 2004).
Due to their physiologic and anatomic similarity to humans, pigs represent an important animal model for drug evaluations in humans (Liu et al., 2008). Our previous report suggested that porcine CYP3A46 is related to human CYP3A4 based on the high amino acid sequence identity and similar oxidation activity (Jiang et al., 2011). In addition, our previous studies suggested that in the proximal promoter region the GC box transactivates the expression of CYP3A46 and that both the GC and CCAAT boxes determine the basal transactivation of CYP3A22 in porcine primary hepatocytes (Dong et al., 2015; Liu et al., 2016). This regulation of porcine CYP3A22 is similar to the regulation of the basal transactivation of human CYP3A5 (Iwano et al., 2001). This regulatory pattern remains to be addressed in the basal regulation of other CYP3A genes, including porcine CYP3A29, CYP3A46, and human CYP3A4 (Rodríguez-Antona et al., 2003; Tirona et al., 2003).
Sp1 is the first identified promoter-specific factor required for the transcription of the simian vacuolating virus 40 genome (Dynan and Tjian, 1983). Sp1 binds to GC boxes through the three conserved C2H2-type zinc fingers in the C-terminus. NF-Y is a trimeric protein formed by NF-Y subunit alpha (NF-YA), NF-Y subunit beta, and NF-Y subunit gamma, and exhibits sequence-specific binding to CCAAT boxes (Maity and de Crombrugghe, 1998). A genome-wide study indicated that CCAAT boxes are mainly located within a conserved distance of −150 to +50 base pairs (bp) from the transcription start sites in 18% of the 21,000 human promoters (Benner et al., 2013). Structural analysis revealed that NF-Y bound DNA in a histone-like mode (Ceribelli et al., 2008). Coimmunoprecipitation suggested the direct interaction between Sp1 and NF-Y in human cells (Roder et al., 1999; Lim and Chang, 2009). In the human genome, CCAAT and GC boxes are observed to have a high tendency to co-occur in regions −150/+50 bp from transcription start sites (Benner et al., 2013). In addition, NF-Y has genome-wide partnerships with Sp1, and the distances between the cis-acting elements are conversed (Dolfini et al., 2016).
In this study, we aimed to determine whether porcine CYP3A genes have a regulatory mechanism of CYP3A4 transactivation similar to that in human cells. We found that both NF-Y and Sp1 can bind the proximal CCAAT and GC boxes in the promoters of CYP3A29 and CYP3A46 as previously reported for CYP3A22 (Liu et al., 2016). However, in contrast to the regulation of CYP3A22, Sp1 is the main regulator of CYP3A29 and CYP3A46 basal expression, while NF-Y minimally contributes to the basal expression. A base-pair mismatch (ACTGG) in the CCAAT box is considered as a main cause of this discrepancy in the regulation of basal transcription. Then, we found similar cis-acting elements in the proximal promoters of CYP3A4 and CYP3A5. A series of experiments showed that the corresponding proximal CCAAT and GC boxes regulated the expression of CYP3A4 and CYP3A5. This transcription model is similar to the model summarized in porcine CYP3A genes and unveils a conserved transcriptional activity pattern between human and porcine CYP3A isoforms. Changing the distance between the NF-Y and Sp1 binding sites repressed the transactivation activity of the proximal promoters in both human and porcine CYP3A genes, and the decreased promoter activity suggested that NF-Y and Sp1 coordinated the constitutive expression of human and porcine CYP3A genes in a spatially coordinated manner.
Materials and Methods
Ethics Statement.
All related experiments were performed in strict accordance with the regulations of the Administration of Affairs Concerning Experimental Animals of Guangdong Province, China, under approval number 2017-D007. All surgeries were performed under anesthesia, and all efforts were made to minimize suffering.
Animals and Cell Culture.
Hepatocytes were isolated from a 9-day-old Danish Landrace × Yorkshire × Duroc cross-breed male pig purchased from the College of Veterinary Medicine, South China Agricultural University. A modified two-step in situ collagenase perfusion procedure was performed as previously described (Liu et al., 2016). Briefly, hepatocytes were isolated by collagenase digestion. After filtering through a cell strainer, the fresh hepatocytes were centrifuged and resuspended in William’s E medium (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) supplemented with 10% FBS (PAN-Biotech, Aidenbach, Germany), 1% penicillin G/streptomycin (Life Technologies, Inc., Invitrogen, CA), 1X SITE (Sigma), 0.1 μM dexamethasone, and 0.1 μM insulin (Invitrogen). The hepatocytes were seeded on a plate at a density of 1 × 105 cells/cm2 and cultured in a humidified 37°C incubator with 5% CO2. After 4 hours, the medium was removed, and the adherent cells were maintained in William’s E medium supplemented with 10% FBS and 1% penicillin G/streptomycin.
HepG2 cells (American Type Culture Collection, Manassas, VA) and COS-7 cells (American Type Culture Collection) were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% FBS and 1% penicillin G/streptomycin antibiotics (Invitrogen) at 37°C and 5% CO2. Huh-7, QGY-7703, and QSG-7701 cells were kindly provided by Professor Xiaofeng Zhu (State Key Laboratory of Oncology in South China, Sun Yat-Sen University Cancer Center, Guangzhou, China). The HepaRG cell line was purchased from Shanghai Guandao Biologic Engineering Co., Ltd. (Shanghai, China). All of the hepatocytes were cultured in DMEM/F-12 supplemented with 10% FBS and 1% penicillin G/streptomycin antibiotics (Life Technologies, Inc., Invitrogen) at 37°C and 5% CO2.
Plasmid Constructs.
Based on NCBI GenBank (https://www.ncbi.nlm.nih.gov/nuccore/NC_010445.4?from=6734565&to=6777685), we amplified the 5′-flanking regions from −4349 to −1 and the deletion constructs from −707 to −1, −364 to −1, −244 to −1, −157 to −1, −131 to −1, and −120 to −1 (the translational start site was designated +1) of porcine CYP3A29 (GenBank Accession Number: NC_010445.4) by polymerase chain reaction (PCR). Porcine genomic DNA from liver tissue was used as a template. The Mlu I (TaKaRa, Qingdao, China) and XhoI (TaKaRa) digested PCR fragments were inserted upstream of pGL3-Basic (Promega Corp., Madison, WI) to generate the luciferase reporter constructs. The 5′-flanking regions of CYP3A5 (−180 to −1) and CYP3A4 (−181 to −1) were cloned from genomic DNA from HepG2 cells. Then, the DNA fragments were inserted into the Mlu I/XhoI sites of pGL3-Basic (Promega Corp.). To further analyze the transcriptional activation of cis-acting elements, variant luciferase reporter constructs were generated using a QuikChange Site-Directed Mutagenesis Kit (Stratagene, La Jolla, CA). The NF-YA and Sp1 overexpression plasmids were generated as previously reported (Liu et al., 2016). All primer sequences are shown in Supplemental Table 1.
KOD-FX DNA polymerase (Toyobo, Osaka, Japan) was used for the PCR. The plasmids were extracted with a TIANprep Mini Plasmid Kit II (TIANGEN, Beijing, China) and verified by DNA sequencing (Sangon Biotech, Shanghai, China). All corresponding constructs were prepared using an Endo-Free Plasmid Kit (Omega, Norcross, GA) for transfection. The extraction and purification of the plasmids were performed according to the manufacturer’s instructions. T4 DNA ligase was purchased from TaKaRa.
Transient Transfection and Luciferase Activity Detection.
Porcine primary hepatocytes were seeded into 24-well plates and cultured in DMEM to 80% confluence. Opti-MEM (Life Technologies) was used as the transfection reagent. The cells in each well of the 24-well plates were transfected with 0.4 μg of the firefly luciferase reporter construct, 0.2 μg of the Renilla luciferase reporter construct (pRL-TK), and 1.5 μl of Lipofectamine 3000 (Invitrogen). HepG2 cells were seeded and cultured in 24-well plates in DMEM. For the CYP3A4 and CYP3A5 reporter constructs, 0.6 μg of the firefly luciferase reporter plasmids and 60 ng of the Renilla luciferase reporter plasmid (pRL-TK) were incubated with 1.5 μl of Lipofectamine 3000 (Invitrogen). The transfected cells were lysed after 24 hours of incubation, and luciferase activity was measured with a dual luciferase reporter assay system (Promega) according to the manufacturer’s instructions. The promoter transcription strength was represented by the relative luciferase activity, and the firefly luciferase activity for each construct was normalized to the Renilla luciferase activity as a transfection reference. In this quantitative analysis, the fold changes in the activity of each construct were calculated relative to the activity of the empty vector (Basic). Each assay was performed in triplicate, and the obtained results represent those of three independent experiments.
To overexpress NF-YA/Sp1, COS-7 cells were seeded and cultured in 10-cm plates until reaching 80% confluence. Then, the cells were transfected with mixtures containing 6 μg of the NF-YA/Sp1 overexpression vector. After 24 hours of transfection, the cells were harvested, the protein level was determined, and the nuclear fractions were isolated.
For the RNA interference, small interfering RNA (siRNA) targeting Sp1, NF-YA, and the unspecific siRNA control were designed and synthesized by GenePharma (Suzhou, China). In total, 40 pmol of siRNA was transiently transfected into HepG2/porcine primary hepatocytes (a well in 12-well plates) using Lipofectamine 3000 (Invitrogen). After 24 hours of transfection, the cells were lysed and subsequently analyzed by western blotting.
Electrophoretic Mobility Shift Assays.
The nuclear fractions from NF-YA/Sp1 overexpressed COS-7 or HepG2 cells were isolated using a Nuclear Extraction Kit (Beyotime, Haimen, China). Then, 5′-biotin–labeled probes corresponding to putative NF-Y or Sp1 binding sites of porcine CYP3A29 and human CYP3A4 and CYP3A5 were prepared using an electrophoretic mobility shift assay (EMSA) Probe Biotin Labeling Kit (Beyotime). EMSA was performed with an EMSA Kit (Beyotime) according to a previously described method (Yang et al., 2014; Dong et al., 2015). Briefly, 5 μg nuclear extract protein from NF-YA/Sp1 overexpressed COS-7 or HepG2 cells were preincubated in binding buffer for 10 minutes at 20°C. For the specific and nonspecific competition assays, 100-fold unlabeled wild-type (WT)/mutant probes were added. For the supershift assay, 1 μg anti-NF-YA antibody (Santa Cruz Biotechnology, Santa Cruz, CA) or anti-Sp1 (Abcam, Cambridge, MA) was added. Then, 0.1 pmol of the 5′-biotin–labeled double-stranded probe was added in a total volume of 10 μl for further incubation for 20 minutes at 20°C.
RNA Isolation and Real-Time PCR.
Total RNA was isolated from HepG2 and porcine primary hepatocytes using TRIzoL reagent (Invitrogen) and reverse transcribed to cDNA using Moloney murine leukemia virus reverse transcriptase (Promega) as previously described (Jiang et al., 2018). Real-time PCR was performed on a CFX Connect Real-Time System (Bio-Rad, Hercules, CA). Reactions were performed using GoTaq qPCR Master Mix (Promega) in 20 µl volumes according to the manufacturer’s recommendations. The primers used were as follows (where GAPDH denotes glyceraldehyde-3-phosphate dehydrogenase):
Porcine GAPDH forward: 5′-GTCGGTTGTGGACCTGAC-3′;
Porcine GAPDH reverse: 5′-TGGTCGTTGAGGGCAATG-3′;
Porcine CYP3A22 forward: 5′-TACCGTAAGGGCATGTGGCA-3′;
Porcine CYP3A22 reverse: 5′-GGTTGGAGACAGCAATGTTCGT-3′;
Porcine CYP3A29 forward: 5′-CCTGAAATTAACCACGCAAGGGCT-3′;
Porcine CYP3A29 reverse: 5′-TCTGGGATGCAGCTTTCTTGACCA-3′;
Porcine CYP3A46 forward: 5′-CGAATCTCTCAGAATATTCCCAGT-3′;
Porcine CYP3A46 reverse: 5′-GCGCAAAGATTGGCACCATCAT-3′;
Human GAPDH forward: 5′-CCTCAAGATCATCAGCAATGCC-3′;
Human GAPDH reverse: 5′-TGGTCATGAGTCCTTCCACGAT-3′;
Human CYP3A4 forward: 5′-GCCTGGTGCTCCTCTATCTA-3′;
Human CYP3A4 reverse: 5′-GGCTGTTGACCATCATAAAAG-3′;
Human CYP3A5 forward: 5′-TGACCCAAAGTACTGGACAG -3′; and
Human CYP3A5 reverse: 5′-TGAAGAAGTCCTTGCGTGTC -3′.
Western Blotting.
The cells were lysed in radioimmunoprecipitation assay buffer as previously described (Ma et al., 2018). The protein expression levels were determined via western blotting as previously described (Liu et al., 2016). The antibodies used in this work included anti-GAPDH antibody (Santa Cruz), porcine CYP3A antibody (mouse polyclonal antibody, laboratory stock), anti-CYP3A4 antibody (Absin), anti-CYP3A5 (Absin), anti-NF-YA antibody (Santa Cruz), and anti-Sp1 antibody (Abcam).
DNA Affinity Precipitation Assay.
The 5′-biotinylated probes were synthesized by PCR amplification of the CYP3A22/CYP3A29 luciferase reporter plasmids with the following primers: forward: 5′ biotin-AGCAGGTGGAGCACAGGGG-3′ and reverse: 5′ biotin- CTGGAGAGTCACTGGGGCT-3′ for CYP3A22 and forward: 5′ biotin-GTGATGACTCGCTGAGGCT-3′ and reverse: 5′ biotin-CTGCAGGAAGAGCACTGGT-3′ for CYP3A29. The assay was performed by incubating 0.5 µg of biotin-labeled probe with 50 µg of nuclear extract in 400 µl binding buffer containing 20 mM HEPES, pH 7.9, 10% glycerol, 50 mM KCl, 0.2 mM EDTA, 1.5 mM MgCl2, 10 µM ZnCl2, 1 mM dithiothreitol, 0.25% Triton-X100, and 1X protease inhibitor (Biotool, Huston) on ice. After 45 minutes, the DNA/protein complexes were incubated with 40 µl pre-equilibrated streptavidin-coated magnetic beads (from mTRAP Maxi) (Active Motif, Carlsbad, CA) for 1 hour with rotation at 4°C. Then, the magnetic beads were pelleted by placing the tubes in a magnetic separation rack, and 1 ml of binding buffer was mixed with the beads, rotated for 5 minutes, and washed five times to remove nonspecific binding. Finally, the binding proteins were eluted with 2X SDS sample buffer with boiling for 5 minutes. Then, the protein samples were separated by SDS-PAGE and analyzed by western blotting with Sp1 or NF-Y antibody to confirm protein binding to the applied probes.
Chromatin Immunoprecipitation Assay.
Chromatin immunoprecipitation (ChIP) assays were performed according to the recommendations of the SimpleChIP Enzymatic Chromatin IP Kit (Cell Signaling Technology Inc., Danvers, MA). In brief, the porcine primary hepatocytes were cross-linked via the addition of 1% final formaldehyde concentration at room temperature for 10 minutes. The addition of micrococcal nuclease and sonication were used to fragment the cross-linked chromatin to a length of approximately 150–900 bp. In total, 10 µg of digested chromatin was used for each immunoprecipitation. The cross-linked DNA-protein complex was immunoprecipitated with 1 µg of antibodies and protein G magnetic beads (Cell Signaling Technology). The anti-H3 antibody (Cell Signaling Technology) and normal rabbit IgG (Cell Signaling Technology) were added as a positive and negative control, respectively, and chromatin was subjected to immunocomplexes with the anti-NF-YA (Santa Cruz) or anti-Sp1 (Abcam) antibodies. The DNA was eluted, the cross-linking was reversed, and the DNA was purified according to the protocol. Then, the immunoprecipitated DNA was used as a template for real-time PCR with special primers. The primers used for the design of the proximal NF-Y and Sp1 binding sites of the CYP3A29 promoter were forward: 5′-GTCCATGGTGCTTTGCCAACTA-3′ and reverse: 5′- CGCTTTGCCTGCTGCTTTCACT-3′. For CYP3A4 and CYP3A5, real-time PCR was performed with the following primers: forward 5′-CAGTGAGGCTGTTGGATTGT-3′ and reverse 5′-TGTGTGATTCTTTGCCAACTTC-3′ for CYP3A4; and forward 5′-TGGACTCCCCGATAACACTGAT-3′ and reverse 5′- AGCCAAGCTGCTGAAAGATTTA-3′ for CYP3A5.
Statistical Analysis.
All statistical procedures were analyzed using SPSS 16.0 (IBM). All figures were plotted using GraphPad Prism 5.0 (GraphPad Software, San Diego, CA). Statistically significant differences among more than two groups were determined using one- or two-way ANOVA, followed by Dunnett’s multiple comparisons tests. For two groups, independent samples and Student’s t tests (two-tailed) were used. The correlations between two factors were evaluated by Pearson correlation analysis. The data are presented as the mean ± S.D. of at least three independent experiments, and significance is defined as ***P < 0.001, **P < 0.01, and *P < 0.05.
Results
The Conserved Proximal CCAAT and GC Boxes in the Promoters of Porcine and Human CYP3As Affected the mRNA Expression of CYP3A Genes.
To investigate the mechanism underlying the regulation of CYP3A gene basal transcription in both porcine and human hepatocytes, sequence alignment was performed to analyze the conservation of cis-acting elements in the proximal promoters of different CYP3A isoforms. As shown in Fig. 1A, two cis-acting elements, i.e., a CCAAT box and a GC box, were identified in the proximal promoter region of six CYP3A genes in both the human and porcine genome. As indicated, the cis-acting elements in the proximal promoter sequence of porcine CYP3A22 were similar to those in human CYP3A5, and the cis-acting elements in the porcine CYP3A29 and CYP3A46 promoters were similar to those in human CYP3A4 and CYP3A7. In the proximal promoters of human CYP3A4 and CYP3A7 and porcine CYP3A29 and CYP3A46, the consensus ACTGG sequence in the CCAAT box is an inverted CCAAT box with a 1 bp mismatch.

The conserved proximal CCAAT and GC boxes in the promoters of porcine and human CYP3A affected the mRNA expression of CYP3A genes. (A) Sequence alignment showing the transcription factor binding sites in the proximal promoter region of typical human and porcine CYP3A family isoforms. The conserved NF-Y and Sp1 binding sites are underlined. (B) Binding of transcription factors NF-Y and Sp1 to the proximal promoter of porcine CYP3A genes in vitro. A DAPA was performed, and the precipitates were analyzed by western blotting with anti-Sp1 and anti-NF-YA antibodies. The histograms represent the DNA affinity precipitated Sp1/input Sp1 ratio and DNA affinity precipitated NF-YA/input NF-YA ratio (n = 3). “ns” represents no significant difference. (C) Effects of Sp1 and NF-Y knockdown on the mRNA levels of pCYP3A genes in porcine primary hepatocytes. Real-time PCR to determine CYP3A22, CYP3A29, and CYP3A46 expression normalized to GAPDH expression in NC, si-Sp1, and si-NF-YA cells (n = 3). “NC”, “si-Sp1”, and “si-NF-YA” cells represent that the cells were transfected with unspecific siRNA control, siRNA targeting Sp1, and NF-YA, respectively. (D) Effects of Sp1 and NF-Y knockdown on the mRNA level of hCYP3A genes in HepG2 cells. Real-time PCR to determine CYP3A4 and CYP3A5 expression normalized to GAPDH expression in NC, si-Sp1, and si-NF-YA cells (n = 3). Statistical significance was assessed using Student’s t test (two-tailed) (B) and two-way ANOVA, followed by Dunnett’s multiple comparisons tests (C and D). The results are shown as the mean ± S.D. of at least three independent experiments and significance was defined as ***P < 0.001; **P < 0.01; or *P < 0.05.
To demonstrate the binding specificity of transcription factors NF-Y and Sp1 to the proximal promoters of porcine CYP3A genes, a DNA affinity precipitation assay (DAPA) was used to verify the ability of these transcription factors to bind promoter DNA in vitro. As indicated in Fig. 1A, the sequences of the proximal promoters of CYP3A29 and CYP3A46 were nearly identical, suggesting the CYP3A29 probe was representative of both CYP3A29 and CYP3A46. By incubating the nuclear fractions of porcine primary hepatocytes with the 5′-biotinylated promoter oligonucleotide of CYP3A29/CYP3A46 and CYP3A22, we demonstrated via the DAPA that the endogenous transcription factors NF-Y and Sp1 directly bound the labeled promoter DNA of CYP3A22 and CYP3A29/CYP3A46 (Fig. 1B). Relative to CYP3A22 promoter DNA binding, Sp1 was almost equally precipitated by the labeled CYP3A29 probe, but the binding of NF-YA to this probe was reduced. This finding suggests that the GC boxes of these two cis-acting elements likely play a vital role in the basal transactivation of genes in the porcine and human CYP3A families. The ability of Sp1 and NF-Y to regulate porcine CYP3A expression in vivo was tested using siRNA targeting Sp1 and NF-YA, respectively; siRNA targeting the coding region of Sp1 and NF-YA was transfected into porcine primary hepatocytes, HepG2 (human hepatocellular carcinoma) cells, QSG-7701cells (normal hepatocytes), and HepaRG cells (a human hepatocellular carcinoma–derived cell line with high CYP450 expression). Consequently, the mRNA levels of porcine and human CYP3As in cells transfected with Sp1 siRNA were decreased (Fig. 1, C and D; Supplemental Fig. 1, A and B). In addition, the mRNA levels of CYP3A4 and CYP3A5 were increased in cells overexpressing Sp1 (Supplemental Fig. 2). We further analyzed the efficiency in knocking down protein expression, as shown in Supplemental Fig. 3. Taken together, these results indicate that knockdown of Sp1 and NF-YA clearly decreased the mRNA expression levels of porcine and human CYP3A genes.
NF-Y and Sp1 Bind the CCAAT and GC Boxes, Respectively, in the Proximal Core Transactivation Region of CYP3A29/CYP3A46.
We generated a series of 5′ deletion firefly luciferase reporter constructs using the pGL3-Basic vector to determine whether this proximal promoter is a core regulatory region in porcine CYP3A29 and CYP3A46. The promoter sequence was identical to the CYP3A29 genome sequence (Accession No. NC_010445.4). These reporters were individually transfected into porcine primary hepatocytes with the reference plasmid pRL-TK containing the Renilla luciferase gene. In this study, the start codon in the CYP3A29 open reading frame was defined as 0. As shown in Fig. 2A, the promoter region from −244 to −1 had the strongest transactivation activity among all constructs, as follows: 31-fold higher than that of the control (pGL3-Basic transfected group). In contrast to the region from −244 to −1, the region from −4349 to −244 did not strongly contribute to basal transactivation and led to a 1.4-fold decrease (Fig. 2A). This finding suggests that consistent with CYP3A22 and CYP3A46, the proximal DNA region of the CYP3A29/CYP3A46 promoter shows the highest promoter activity. More interestingly, the relative luciferase activity gradually decreased from 31-fold (from −244 to −1), 21-fold (from −157 to −1), and 11-fold (from −131 to −1) to 1.6-fold (from −120 to −1) compared with that in the control (Fig. 2A). These results strongly suggest that the minimal and essential region responsible for the basal transactivation of CYP3A29/CYP3A46 is a 36-bp region from −157 to −120 and that some cis-acting elements are likely located in this region (Fig. 2A). As indicated in the sequence alignment shown in Fig. 1A, these cis-acting elements are likely the binding sites of NF-Y and Sp1.

NF-Y and Sp1 bind the CCAAT and GC boxes, respectively, in the proximal core transactivation region of CYP3A29/CYP3A46. (A) Deletion analysis of the CYP3A29 promoter in porcine primary hepatocytes. Porcine primary hepatocytes were transiently transfected with the luciferase (LUC) reporter plasmid pGL3-Basic, which contained different lengths of the CYP3A29 promoter. The left panel shows the position and length of the constructs. The start codon in the CYP3A29 open reading frame was defined as +1. The Renilla luciferase reporter plasmid pRL-TK was cotransfected into cells as a normalization control. The right panel shows the relative activity of each construct with respect to that of emptypGL3-Basic vector under the same conditions (n = 3). (B) An EMSA was used to confirm the binding of Sp1 to the CYP3A29/CYP3A46 proximal promoter. The nuclear fraction was extracted from Sp1-overexpressing COS-7 cells. The top panel shows the sequences of the CYP3A29 Sp1 and mutant probes. The predicted binding sites and nonbinding mutant sequences are underlined. A 100-fold excess of the unlabeled CYP3A29 Sp1 and mutant probes were added to the reactions in lanes 3 and 4 as competition experiments. The formation of DNA/Sp1 complexes (arrows) was then monitored by 5% native gel electrophoresis. (C) A DAPA was used to further confirm the direct binding of Sp1 to the CYP3A29/CYP3A46 proximal promoter. The DNA/protein complexes were analyzed by western blotting with anti-Sp1 and anti-NF-Y antibodies (n = 3). mut-Sp1 represents the Sp1 with a binding site mutation, and mut-NF-Y represents the NF-Y with a binding site mutation. The histograms show the DNA affinity precipitated Sp1/input Sp1 ratio and DNA affinity precipitated NF-YA/input NF-YA ratio, normalized to that of the WT (CYP3A29). (D) A ChIP assay was performed to quantitatively analyze the binding of NF-YA and Sp1 to the cis-acting elements in the CYP3A29/CYP3A46 proximal promoter in porcine hepatocytes (n = 3). Statistical significance was determined using one-way ANOVA, followed by Dunnett’s tests (A and D) and two-way ANOVA, followed by Dunnett’s tests (C). The data are presented as the mean ± S.D. of at least three independent experiments, and significance was defined as ***P < 0.001.
An EMSA and a DAPA were used to further demonstrate the binding specificity of transcription factors NF-Y and Sp1 to the CYP3A29/CYP3A46 proximal promoter in vitro. As shown in Fig. 2B, two shifted bands formed in the Sp1-overexpressing COS-7 nuclear fractions incubated with the 5′-biotin–labeled CYP3A29/CYP3A46 probe (Fig. 2B, Sp1-WT). The addition of a 100-fold excess of unlabeled WT probe but not the cold mutant probe (Fig. 2B, Sp1-mut) competed out the shifted bands (Fig. 2B, lanes 3 and 4). The shifted bands were slowed by protein specifically binding the GC box in the WT CYP3A29/CYP3A46 probe. The DNA-protein complex was supershifted following incubation with anti-Sp1 antibody, and the density of the upper binding band was markedly reduced (Fig. 2B, lane 5). Thus, Sp1 specifically bound the proximal promoter of CYP3A29/CYP3A46, and the upper shifted band was formed by Sp1 binding. The DAPA was performed using WT oligonucleotides for the CYP3A29/CYP3A46 promoter and mutated oligonucleotides for Sp1 or NF-Y to further confirm the binding specificity of Sp1 and NF-Y to the CYP3A29/CYP3A46 proximal promoter region (Fig. 2C). Moreover, the ChIP assays demonstrated that NF-YA and Sp1 were abundantly recruited to the proximal promoter region from nucleotides −212 to −54 of CYP3A29/CYP3A46 in vivo (Fig. 2D).
Sp1 and NF-Y Directly Transactivate the Transcription of CYP3A29/CYP3A46.
As shown in Fig. 1A, certain proximal promoter regions in CYP3A29 and CYP3A46 have only one mismatch that is unrelated to the two cis-acting elements. We hypothesized that porcine CYP3A genes have a unified mechanism that regulates basal transcription and that the CCAAT boxes in CYP3A29/CYP3A46 and CYP3A22 make various contributions to transcription. To investigate the activation capability of the GC and CCAAT boxes, we used site-directed mutagenesis to mutate or remove these binding sites to disrupt the binding of these transcription factors to the promoter DNA. The mutation and deletion sites are underlined in the top panel of Fig. 3A. Both the mutation and deletion of the Sp1 binding site resulted in a 90% decrease in luciferase activity compared with that driven by the WT promoter. However, relative to the activity driver by the CCAAT box in CYP3A22, the change in luciferase activity was not dramatic, but a decrease was observed when the NF-Y binding site was mutated (Fig. 3A). These data suggest that the proximal GC box dominates the basal transactivation of porcine CYP3A29 and CYP3A46. The difference between CYP3A29/CYP3A46 and CYP3A22 may be due to the single base-pair mutation in the CCAAT box of the CYP3A29/CYP3A46 proximal promoter.
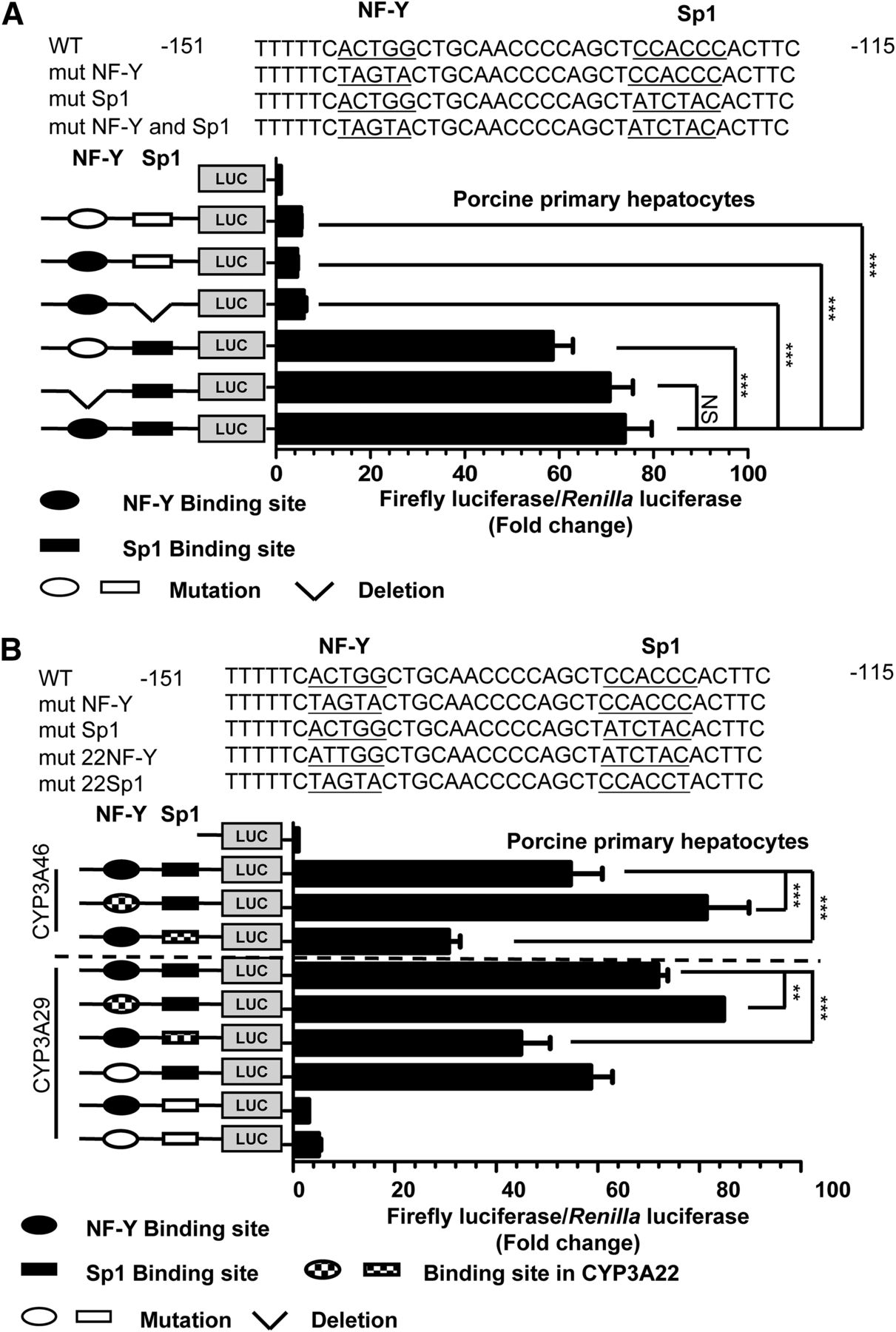
Sp1 (NF-Y) directly transactivates the transcription of CYP3A29 (CYP3A46). (A) Mutation analysis of the CYP3A29/CYP3A46 proximal promoters in porcine primary hepatocytes. The top panel shows the −151 to −115 promoter region; the CCAAT box, GC box, and mutated bases are underlined. The ellipses and rectangles represent the binding sites of transcription factors NF-Y and Sp1, respectively. The black and white shapes represent the WT and mutated transcription factor binding sites, respectively. The right panel shows the relative activity of each construct relative to that of the empty vector pGL3-Basic vector under the same conditions (n = 3). “NS” represents no significant difference. (B) Replacement of the NF-Y and Sp1 binding sites in CYP3A29/CYP3A46 with the sequence in CYP3A22. The sequences of the WT and mutated promoters used in the experiment are shown in the top panel. In the bottom panel, each column represents the fold change in the luciferase activity of each mutated construct relative to that of Basic (the pGL3-Basic plasmid) (n = 3). mut-Sp1 represents the Sp1 binding site mutation, and mut-NF-Y represents the NF-Y binding site mutation. Statistical significance was determined using one-way ANOVA, followed by Dunnett’s tests (A) and two-way ANOVA, followed by Dunnett’s tests (B). The data are presented as the mean ± S.D. of at least three independent experiments, and significance was defined as **P < 0.01, ***P < 0.001.
To explore whether this variation in the NF-Y binding site resulted in different effects on the transcriptional activation of CYP3A22 and CYP3A29, we replaced the NF-Y and Sp1 binding sites for CYP3A29 with the binding site for CYP3A22 in the basal regulatory region of CYP3A29 and CYP3A46. The mutated/exchanged region is underlined in the top panel of Fig. 3B. Mutation of the NF-Y binding site from ACTGG to ATTGG, similar to the cis-acting element in the CYP3A22 proximal promoter, caused a 17% increase in luciferase activity. Unlike the WT promoter, the Sp1 binding site contained a CCACCT sequence (the GC box sequence of CYP3A22) rather than a CCACCC sequence (the GC box sequence of CYP3A29), resulting in a 37% decrease in CYP3A29 promoter activity. Similarly, in the proximal promoter of CYP3A46, the replacement of cis-acting elements from the CYP3A22 proximal promoter resulted in a 45% decrease in activity for the GC box, and a 38% increase in activity for the CCAAT box. As Supplemental Fig. 4 indicates, we replaced the NF-YA and Sp1 binding sites in CYP3A22 with the binding sites in CYP3A29/CYP3A46, and the promoter activity of CYP3A22 decreased for mutated 29NF and increased for mutated 29Sp1. These data indicate that Sp1 and NF-Y play important roles in porcine CYP3A basal expression, and the variations in the cis-acting elements altered their individual contributions to the basal transcription levels.
Both NF-Y and Sp1 Activate CYP3A5 and CYP3A4 Transcription by Binding the Proximal CCAAT and GC Boxes.
The proximal promoter regions of six CYP3A genes were highly conserved between humans and pigs (Fig. 1A). These six CYP3A genes contained an inverted CCAAT box and one proximal GC box. To determine whether these two conserved motifs truly activate human CYP3A5 and CYP3A4, we amplified the proximal 5′ upstream sequence in human CYP3A5 (from −180 to −1) and CYP3A4 (from −181 to −1). These regions were similar to the region from −157 to −1 in CYP3A29, and the genomic sequence was confirmed (Accession No. NC_000007.14). Based on the proximal sequences, we generated a series of reporter constructs with mutations or deletions of these two motifs. These luciferase reporters were transfected into HepG2 cells individually with the reference plasmid pRL-TK. As shown in Fig. 4A, the relative luciferase activity decreased by 17-fold for the constructs with deletion of either the CCAAT or GC box in the CYP3A5 proximal promoter compared with that driven by the constructs containing the region from −180 to −1. For the constructs with mutation of a cis-acting element, the decrease in luciferase activity was similar to the decrease in the deletion constructs. These reductions in luciferase activity when each cis-acting element was dysfunctional demonstrated that both the CCAAT box and the GC box are vital for the basal transactivation of CYP3A5. These results are consistent with a previous report of the molecular mechanism underlying basal CYP3A5 expression (Iwano et al., 2001).

Both NF-Y and Sp1 activate CYP3A5 and CYP3A4 transcription by binding to the proximal CCAAT and GC boxes. (A and B) Mutation analysis of the proximal CYP3A5 and CYP3A4 promoters in HepG2 cells. The top panels in (A and B) show the WT and mutated promoter sequences. The mutated region (underlined) destroyed the CCAAT and GC boxes. The schematic maps on the left show the mutant constructs. The right panel shows the promoter activity relative to that of Basic (the pGL3-Basic plasmid) (n = 3). mut-Sp1 represents the Sp1 binding site mutation, and mut-NF-Y represents the NF-Y binding site mutation. Statistical significance was determined using one-way ANOVA, followed by Dunnett’s tests. The data are presented as the mean ± S.D. of at least three independent experiments, and significance was defined as ***P < 0.001.
Another Sp1 binding site was previously identified in the proximal promoter of CYP3A4, and this site was deduced to be irrelevant to the promoter activity of CYP3A4 (Bombail et al., 2004). However, according to the sequence alignment shown in Fig. 1A, the previously reported Sp1 site is 32 bp upstream of our identified Sp1 binding site. Whether this conserved CCAAT box/GC box is involved in the transactivation of CYP3A4 remains unclear. To elucidate the role of the NF-Y and Sp1 binding sites in the transcriptional activation of human CYP3A4, we performed a dual luciferase assay with the construct containing the promoter region from −181 to −1 and constructs containing binding site mutations. As shown in Fig. 4B, the relative luciferase activity was reduced by 54% and 60% in HepG2 cells with NF-Y binding site deletion and mutation, respectively, compared with that driven by the WT promoter. The deletion and mutation of the Sp1 binding site resulted in luciferase activity decreases of 53% and 77%, respectively. Due to the low expression level of CYP3A4 and CYP3A5 in HepG2 cells, we performed luciferase assays in Huh-7, QGY-7703, QSG-7701, and HepaRG cells (Supplemental Fig. 5), and obtained the same results. These results suggest that the conserved CCAAT and GC boxes are important elements responsible for the proximal transactivation of human CYP3A4.
NF-Y and Sp1 Bind the Proximal CCAAT and GC Boxes, Respectively, of CYP3A5/CYP3A4.
The deletion and mutation constructs demonstrated that the CCAAT and GC boxes in the proximal promoters of CYP3A4 and CYP3A5 were involved in the basal transactivation of CYP3A4 and CYP3A5. To further confirm that NF-Y and Sp1 activate transcription by directly binding the CCAAT and GC boxes, respectively, an EMSA was performed using nuclear protein prepared from HepG2 cells. The sequences of the probes are shown in the top panel of Fig. 5A. As shown in the bottom panel of Fig. 5A (lane 2), a shift band was detected when the HepG2 nuclear fractions were incubated with the 5′-biotin–labeled WT probe (corresponding to the CCAAT box in the proximity of CYP3A5). The protein/DNA complex could be completed by the addition of the 100-fold unlabeled WT probe, but not by the addition of the 100-fold unlabeled binding site mutant probe. This result suggests that the protein/DNA binding complex was formed through specific binding (Fig. 5A, lanes 3 and 4). The addition of the NF-YA antibody resulted in a shift band that was supershifted (Fig. 5A, lane 5). Our results reveal that NF-Y directly binds the CCAAT box to activate the transcription of CYP3A5. Meanwhile, an EMSA was performed to confirm that Sp1 binds the GC box. The sequence of the 5′-biotin–labeled probe corresponded to the proximal GC box of CYP3A5. The labeled probe was incubated with nuclear fractions from HepG2, and two shift bands were detected in the gel (Fig. 5B, lane 2). The addition of the 100-fold unlabeled probe and the unlabeled mutant probe further confirmed that the shift bands were specifically formed by DNA/protein binding (Fig. 5B, lanes 3 and 4). The upper band was abolished when the complex was supershifted by the Sp1 antibody, confirming that the upper band was formed by the specific binding of Sp1 to the GC box. Taken together, NF-Y and Sp1 are directly associated with the two motifs identified in the proximal promoter of CYP3A5 in vitro.
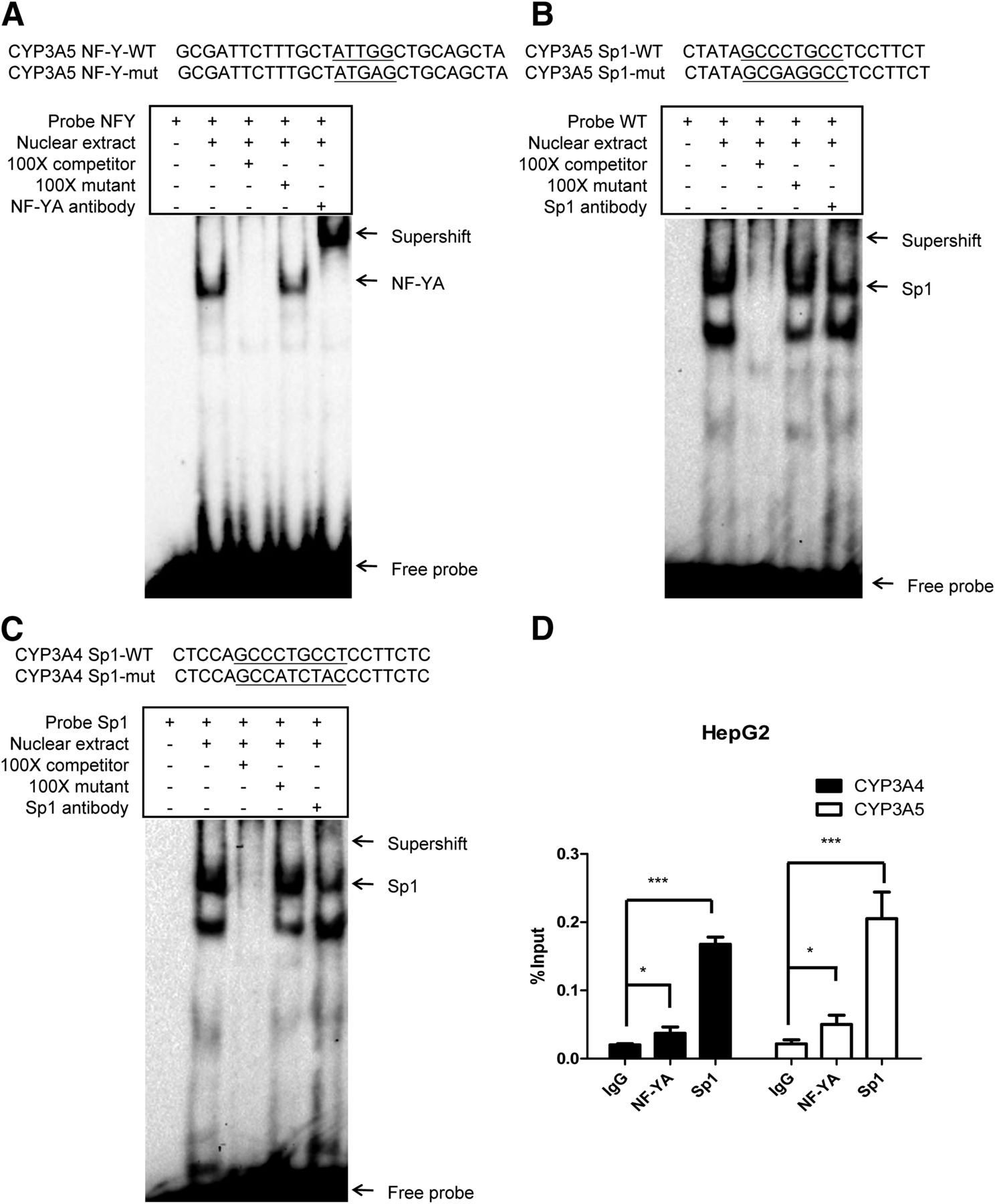
NF-Y and Sp1 bind the proximal CCAAT and GC boxes, respectively, of CYP3A5/CYP3A4. (A and B) EMSA of CYP3A5 NF-Y and Sp1 probes. Probes corresponding to the CYP3A5 proximal CCAAT box (A) and GC box (B) are shown above the gel. Binding sites and mutated sequences are underlined. Probe incubated with nuclear fraction (5 μg) from HepG2 cells. EMSAs were performed as described in Materials and Methods. DNA/protein complexes and the supershift band are indicated with arrows. (C) EMSA of CYP3A4 Sp1 probes. (D) ChIP assays were performed to quantitative analysis of the binding of NF-YA and Sp1 to cis-acting elements in the CYP3A4/CYP3A5 proximal promoter in HepG2 cells. Statistical significance was determined using two-way ANOVA, followed by Dunnett’s multiple comparisons tests. Data are presented as the mean ± S.D. of at least three independent experiments, and significance was defined as *P < 0.05, ***P < 0.001.
To determine whether Sp1 is recruited to the proximal GC box of CYP3A4, an EMSA was performed with nuclear protein from HepG2 cells and a probe corresponding to the GC box of CYP3A4. After incubation, two shift bands of DNA/protein complexes were detected (Fig. 5C, lane 2). The upper band was supershifted against the Sp1 antibody (Fig. 5C, lane 5). As shown in Fig. 5C (lane 3), the shift bands could be competed with the addition of the 100-fold unlabeled WT probe containing the Sp1 binding site. The addition of the 100-fold unlabeled binding site mutant probe did not affect the formation of the shift band. This finding further confirms that Sp1 directly and specifically binds the proximal GC box of CYP3A4 in vitro.
We corroborated the direct binding in vivo through a ChIP assay with anti-NF-YA and anti-Sp1 antibodies. As shown in Fig. 5D, the NF-Y and Sp1 binding is enriched at the CYP3A4 and CYP3A5 proximal promoters. Consistent with our previous results, compared with CYP3A5, the CYP3A4 proximal promoter with the mutated CCAAT box exhibits weaker binding of NF-YA.
Coordinated Regulation of Human and Porcine CYP3A Genes by NF-Y and Sp1.
Notably, we observed that the distance between the two cis-acting elements is constant at 11 bp (approximately one helix of the DNA double helix). To explore whether NF-Y and Sp1 coordinately participate in the transcriptional activation of human CYP3A genes, we generated a series of constructs that alter the distance between the NF-Y and Sp1 binding sites of CYP3A5 and CYP3A4 as 1, 6, and 16 bp without changing the binding motifs that were then transfected into HepG2 cells. As shown in Fig. 6A, the activity of the construct containing the region from −180 to −1 of CYP3A5 was 65-fold higher than that of the empty pGL3-Basic vector. When we deleted the 10 bases (−10) between the NF-Y and Sp1 binding sites, the promoter activity decreased by 11-fold compared with that with the Basic vector. The four-base deletion (−4) and five-base insertion (+5) decreased the activity by 33- and 25-fold, respectively. Combined with the data from the element mutation/deletion analysis by the dual luciferase assay (Fig. 4A), these results indicate that these two factors may play a synergistic role in the transcriptional activation of CYP3A5. As shown in Fig. 6B, the proximal promoter activity of the −10, −4, and +5 constructs were reduced by 75%, 75%, and 54%, respectively, for CYP3A4, and we further confirmed the results in Huh-7, QGY-7703, QSG-7701, and HepaRG cell lines (Supplemental Fig. 6).
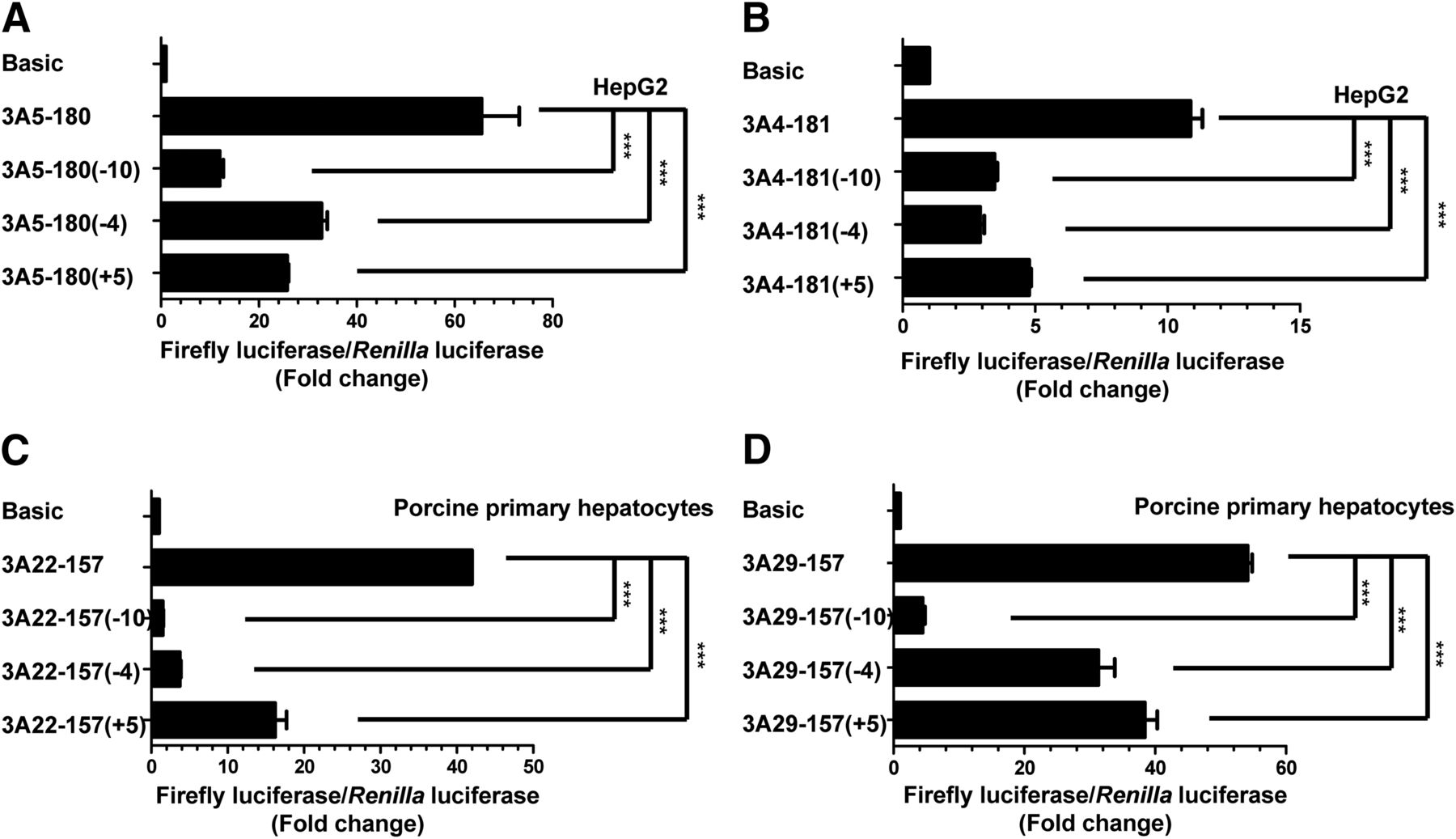
Coordinated regulation of human and porcine CYP3A genes by NF-Y and Sp1. (A and B) Change in distance between the NF-Y and Sp1 binding sites in the CYP3A5 (A) or CYP3A4 (B) promoter region impacts transcript activation. As described in Materials and Methods, a series of constructs with different distances between these two transcription factors were generated and transfected into HepG2 cells. The left panel shows the distance between the NF-Y and Sp1 binding sites before and after the changes. The right panel represents the fold change in luciferase activities relative to pGL3-Basic (Basic) (n = 3). (C and D) Change in the distance between the NF-Y and Sp1 binding sites in the CYP3A22 (C) and CYP3A29 (D) promoter regions impacts transcript activation. Distance-changed constructs were generated as described in Materials and Methods and then transfected into porcine primary hepatocytes. Statistical significance was determined using one-way ANOVA, followed by Dunnett’s multiple comparisons tests. Data are presented as the mean ± S.D. of at least three independent experiments, and significance was defined as ***P < 0.001.
To further investigate whether this coordinated relationship between NF-Y and Sp1 is conserved between the human and porcine CYP3A family, we generated similar distance-changed constructs for CYP3A22 (Fig. 6C) and CYP3A29/CYP3A46 (Fig. 6D) that were then transfected into porcine primary hepatocytes. The effect of the different distances between NF-Y and Sp1 on the promoter activity was tested via a dual luciferase assay. The proximal promoter activity of the −10, −4, and +5 constructs was reduced by 93%, 90%, and 63% for CYP3A22 (Fig. 6C) and 91%, 43%, and 31% for CYP3A29/CYP3A46 (Fig. 6D), respectively. Taken together, without changing the binding sites of NF-Y and Sp1, the spatial distance determined the proximal promoter activities of CYP3A5 and CYP3A22 harboring the canonical CCAAT box and CYP3A29 harboring the mutated CCAAT box, which might weaken the binding of NF-Y, including CYP4A46 and CYP3A4, although to different extents. Overall, these data indicate that Sp1 and NF-Y probably have a conserved synergistic relationship in the transcriptional activity of human and porcine CYP3A genes.
Discussion
The cytochrome P450 family is a key metabolic enzyme superfamily that is responsible for numerous metabolic pathways of endogenous and xenobiotic compounds in plants, animals, and microorganisms (Nelson et al., 1996). The porcine CYP3A subfamily consists of CYP3A22 (also known as CYP3A39), CYP3A29, and CYP3A46 (Sakuma et al., 2004). All three porcine CYP3A isoforms perform the same function as CYP3A4 in the metabolism of endogenous and xenobiotic compounds (Wang et al., 2011; Wu et al., 2016). Indeed, we found that distal hepatocyte nuclear factor 1 and the proximal GC box function in the basal transcriptional regulation of CYP3A46 (Dong et al., 2015), which is similar to the regulation of CYP3A4 (Tirona et al., 2003; Bombail et al., 2004; Matsumura et al., 2004). We recently demonstrated that both the CCAAT box and the GC box in the proximal promoter play important roles in the basal expression of CYP3A22 (Liu et al., 2016), which is similar to the regulation of CYP3A5 in humans (Iwano et al., 2001). These studies indicate that similar to human CYP3A genes, the porcine CYP3A isoforms might be regulated by different mechanisms rather than by the conserved regulation of CYP3A4.
The CYP3A genes are clustered on human chromosome 7 and porcine chromosome 3. Evolutionarily, these genes are potentially regulated by a conserved pattern due to gene duplication (McArthur et al., 2003; Yan and Cai, 2010), but regulation by diversified DNA alterations, including in the promoter region and coding sequence, is possible. To confirm this hypothesis, the alignment of the proximal promoter sequences between the human CYP3A genes, including CYP3A4, CYP3A5, and CYP3A7, and the porcine CYP3A genes, including CYP3A22, CYP3A29, and CYP3A46, were apparently two conserved proximal cis-acting elements (a GC box and a CCAAT box at a constant 11-bp distance). Similarly, the CCAAT box in the proximal promoter regions of CYP3A4 and CYP3A7 contained 1 bp mutation, which is identical to that in porcine CYP3A29 and CYP3A46 (Fig. 1A). These aspects of the proximal promoter sequences support our hypothesis that the porcine CYP3A and human CYP3A genes may share the same core mechanism for the regulation of basal transactivation.
Subsequently, we explored whether these two proximal cis-acting elements play determinant roles in the basal expression of CYP3A29 and CYP3A46 as previously reported in CYP3A22 (Liu et al., 2016). The EMSA and DAPA confirmed that Sp1 directly binds the canonical GC box of CYP3A29/CYP3A46 in vitro (Figs. 1B and 2, B and C). The mutated CCAAT box in the proximal region of the CYP3A29/CYP3A46 promoter recruited the NF-Y complex, but to a much weaker extent, and in contrast to NF-Y, apparently activated the basal expression of CYP3A22. Both direct bindings were shown in the ChIP assay in vivo, and thus Sp1 strongly (while NF-Y weakly) contributes to the basal expression of CYP3A29/CYP3A46 (Fig. 2D). This unequal contribution of the two transcription factors to the basal transactivation of porcine CYP3A genes was further supported by the truncate promoter analysis of CYP3A29 and binding-site mutagenesis analysis of porcine primary hepatocytes (Figs. 2A and 3A). Together with our previous data (Dong et al., 2015; Liu et al., 2016), we conclude that the core transactivation region of porcine CYP3A genes is in the proximal promoter region, and Sp1 plays a significant role in the basal transcription of CYP3A22, CYP3A29, and CYP3A46. However, the function of NF-Y in the basal expression of CYP3A29/CYP3A46 remains to be further elucidated due to its contribution to the basal transactivation of CYP3A22, which is much stronger than that to the transactivation of CYP3A29 and CYP3A46. To further determine whether the weak binding of NF-Y to the mutated CCAAT box has an impact on its basal expression, we replaced the mutated CCAAT box with either a typical CCAAT box from CYP3A22 or another null site without NF-Y binding. The luciferase activity apparently increased following the canonical CCAAT box replacement and decreased with the null site construct. Additionally, the DAPA revealed that the binding capabilities of NF-Y at the CYP3A29/CYP3A46 probe are much weaker than those at the CYP3A22 probe in the proximal promoter region (Fig. 1B). Taken together, NF-Y also regulates porcine CYP3A expression to different extents, because the single base variation of the CCAAT site caused alterations in the binding capacity of NF-Y in the proximal promoter region of CYP3A29, CYP3A46, and CYP3A22.
As previously mentioned, the same cis-acting elements were identified in the proximal promoter regions of three human CYP3A genes. Thus, we were interested to explore the regulatory functions of these two cis-acting elements in human CYP3A genes. As the predominant cytochrome P450 in the human adult liver (Lacroix et al., 1997), CYP3A4 is directly regulated by many transcription factors, and its expression has been extensively studied. However, few studies have investigated the proximal CCAAT and GC boxes, and only one study reported a Sp1 site previously identified in the CYP3A4 promoter region with no functions in transcriptional activation (Bombail et al., 2004). However, this reported Sp1 binding site completely differs from the identified GC box in our study and is 32 bp upstream. Thus, the basal regulation of CYP3A4, especially the importance of its proximal promoter region with these two cis-acting elements, must be reconsidered, and the cis-acting elements must be further elucidated. Regarding the study strategy used to investigate the porcine CYP3A genes, the mutation analysis of the proximal binding sites of the CYP3A5 (Fig. 4A) and CYP3A4 (Fig. 4B) promoters revealed that both NF-Y and Sp1 participated in the basal transcription activity of human CYP3A isoforms. The EMSA and ChIP assay confirmed that both NF-Y and Sp1 directly bind the CYP3A5 and CYP3A4 proximal promoters in vitro and in vivo (Fig. 5, A–D).
The interaction between NF-YA and Sp1 has been shown using a yeast two-hybrid system and glutathione S-transferase pull-down assay (Roder et al., 1999). Sp1 and NF-Y may coordinate in the basal regulation of CYP3A genes in humans and pigs. Interestingly, we found that these two cis-acting elements in the proximal promoter regions of six CYP3A genes maintain a constant distance at 11 bp as a DNA double helix. This coordination was confirmed by a global promoter analysis of the human genome (Benner et al., 2013; Dolfini et al., 2016) as follows: 27% and 18% of 21,000 human promoters have a GC box and a CCAAT box in the region from −150 to +50, and the CCAAT box and GC box co-occur with high tendencies in the same promoter with a co-binding pattern. A previous report showed that the heterotrimeric transcription factor NF-Y consists of a minor groove-binding subunit NF-YA and two histone-fold domains containing NF-Y subunit beta/NF-Y subunit gamma (Romier et al., 2003; Nardini et al., 2013). The C-terminus of Sp1 contains a canonical DNA binding domain composed of major groove-binding C2H2-type zinc fingers (Thiesen and Schröder, 1991). Based on the known DNA binding structure of NF-Y (Protein Data Bank ID: 4AWL) (Nardini et al., 2013) and Sp1 (Protein Data Bank ID: 1AAY) (Elrod-Erickson et al., 1996; Oka et al., 2004), the spatial configuration of NF-Y, the Sp1-DNA binding to promoters with one helix distance, and the direction of the binding could provide an exact space for NF-Y and Sp1 to bind the cis-acting elements. Changing the distance between the NF-Y and Sp1 binding sites repressed the transactivation activities in the proximal promoters of the porcine and human CYP3A genes (Fig. 6, A–D). These results support the hypothesis that the spatial colocalization of two trans-acting factors is required for the basal transcription of CYP3A genes in an evolutionarily conserved manner.
In conclusion, our study suggests that porcine and human CYP3A genes are constitutively expressed under the control of their proximal promoters. The basal expression of these CYP3A genes depends on the recruitment of Sp1 and NF-Y via cis-acting elements in the proximal region. Regarding the CYP3A genes, the CCAAT and GC boxes contribute to the conserved regulation mechanism but to different extents due to a single base DNA variation.
Acknowledgments
We thank Xiaofeng Zhu (State Key Laboratory of Oncology in South China, Sun Yat-Sen University Cancer Center, Guangzhou, China) for kindly provided the Huh-7, QGY-7703, and QSG-7701 cell lines.
Authorship Contributions
Participated in research design: Chen, Jiang, Wen, Deng.
Conducted experiments: Chen, Hu, Ye, Li.
Performed data analysis: Chen, Yuan.
Wrote or contributed to the writing of the manuscript: Chen, Jiang, Wen, Deng.
Footnotes
- Received September 12, 2018.
- Accepted February 3, 2019.
This work was supported by the National Nature Science Foundation of China [Grant 31471234], the Natural Science Foundation of Guangdong Province [Grant 2015A030312005], the Science and Technology Program of Guangzhou [Grants 201804020067 and 201607010177], the Department of Education of Guangdong Province [Grants 2014GKXM020 and 2017KCXTD001], and the Pearl River S&T Nova Program of Guangzhou [Grant 201610010063].
↵
 This article has supplemental material available at molpharm.aspetjournals.org.
This article has supplemental material available at molpharm.aspetjournals.org.

Abbreviations
- bp
- base pair
- ChIP
- chromatin immunoprecipitation
- DAPA
- DNA affinity precipitation assay
- DMEM
- Dulbecco’s modified Eagle’s medium
- EMSA
- electrophoretic mobility shift assay
- GAPDH
- glyceraldehyde-3-phosphate dehydrogenase
- NF-Y
- nuclear transcription factor Y
- NF-YA
- nuclear transcription factor Y subunit alpha
- PCR
- polymerase chain reaction
- siRNA
- small interfering RNA
- Sp1
- specificity protein 1
- WT
- wild type
- Copyright © 2019 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}