
Abstract
Deciduous shrub growth has increased across the Arctic simultaneously with recent climate warming trends. The reduction in albedo associated with shrub-induced 'greening' of the tundra is predicted to cause significant positive feedbacks to regional warming. Enhanced soil fertility arising from climate change is expected to be the primary mechanism driving shrub responses, yet our overall understanding of the relative importance of soil nitrogen (N) and phosphorus (P) availability and the significance of other ecological drivers is constrained by experiments with varying treatments, sites, and durations. We investigated dwarf birch apical stem growth responses to a wide range of ecological factors (enhanced summer temperatures, deepened snow, caribou exclusion, factorial high level nitrogen and phosphorus additions, and low level nitrogen additions) after six years of experimental manipulations in birch hummock tundra. As expected, birch apical stem growth was more strongly enhanced by the substantial increases in nutrient supply than by our changes in any of the other ecological factors. The factorial additions revealed that P availability was at least as important as that of N, and our low N additions demonstrated that growth was unresponsive to moderate increases in soil nitrogen alone. Experimental warming increased apical stem growth 2.5-fold—considerably more than in past studies—probably due to the relatively strong effect of our greenhouses on soil temperature. Together, these results have important implications for our understanding of the biogeochemical functioning of mesic tundra ecosystems as well as predicting their vegetation responses to climate change.
Export citation and abstract BibTeX RIS
Content from this work may be used under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Current rapid warming in the Arctic is leading to substantial changes in vegetation (ACIA 2005). Increasing deciduous shrub growth in response to warmer mean annual temperatures has been reported from remote sensing, dendrochronology, plot-based monitoring, and experimental warming studies (Chapin et al 1995, Goetz et al 2005, Tape et al 2006, Walker et al 2006, Forbes et al 2009, Jia et al 2009, Beck and Goetz 2011, Ropars and Boudreau 2012, Tremblay et al 2012). This increased shrub density and expansion across the landscape is projected to reduce albedo (Sturm et al 2005, Loranty et al 2011), affect permafrost degradation (Blok et al 2010, Bonfils et al 2012, Lawrence and Swenson 2012), alter nutrient cycling and ecosystem carbon balance (Weintraub and Schimel 2005, Natali et al 2011), and may result in a positive feedback to climate warming (Chapin et al 2005, Swann et al 2010, Bonfils et al 2012).
Mean annual temperatures across the Arctic are projected to increase by 5–7 °C by 2100—two times the projected global mean temperature increase (ACIA 2005)—yet it is expected that the indirect effects of higher temperatures, particularly enhanced nutrient availability (Nadelhoffer et al 1991, Chapin et al 1995), will have the greatest impacts on shrub productivity (Sturm et al 2001, Weintraub and Schimel 2005). Experimental manipulations conducted over the past 20 years in arctic tundra have led to the following general conclusions: deciduous shrub growth substantially increases with chronic high level N + P fertilization (Chapin et al 1995, Jonasson et al 1999, Shaver and Jonasson 1999, Bret-Harte et al 2001, 2002, van Wijk et al 2004); is either unaffected or increases modestly with experimental warming (Chapin et al 1995, van Wijk et al 2004, Walker et al 2006, Hudson and Henry 2010); is unaffected or increases with deeper snow (Wahren et al 2005, Vankoughnett 2010) and is either unaffected or decreases with northern ungulate herbivory (Gough et al 2008, Post and Pedersen 2008, Olofsson et al 2009, Van Bogaert et al 2009).
Determination of the relative importance of each of these ecological factors is a prerequisite to comprehending and predicting the complex interactions and feedbacks that will ultimately drive vegetation change in a warming Arctic. Several meta-analyses have integrated the large body of data from previous experimental manipulations (Dormann and Woodin 2002, van Wijk et al 2004), but the conclusions are confounded by inherent differences among sites, differences in treatment durations, and differences in fertilizer application levels across the studies (see van Wijk et al 2004 for a thorough treatment list). Furthermore, although chronic high level N + P addition studies have been common, the relative importance of each nutrient has rarely been investigated. For example, there are only three such studies in mesic tundra vegetation, all of which were of short duration (Haag 1974, Shaver and Chapin 1980, Baddeley et al 1994). These past factorial nutrient addition studies in shrub-dominated mesic tundra systems report N limitation or N and P co-limitation of plant growth after just 1–2 growing seasons of manipulations (Haag 1974, Shaver and Chapin 1980, Baddeley et al 1994), supporting the widespread focus on N as the key growth limiting nutrient in mesic tundra (e.g. Mack et al 2004, Weintraub and Schimel 2005), even though it is well known that longer-term vegetation responses are often quite different from short-term responses (Chapin et al 1995, Chapin and Shaver 1996).
Investigating the influence of N and P separately is important because there are fundamental differences in the biogeochemical cycling of each element, such as the much lower mobility of P in soils and its lack of a significant gaseous component (Schlesinger 1997), which may have critical implications for our understanding of how arctic terrestrial ecosystems function and how they will respond to climate change. Additionally, there is a great need for low level nutrient addition studies, since the high level fertilizer applications remove nutrient limitation as intended but may not be appropriate to understanding possible vegetation responses to climate change since the increases in soil fertility with warming are likely to be relatively slow and subtle (Hobbie et al 2002). To effectively assess the relative importance of these ecological parameters to arctic shrub growth, it is critical that the site and growing seasons be held constant, and that increased resolution of fertilizer treatments be tested.
Here we assess growth responses of Betula glandulosa Michx.—a common deciduous shrub throughout the North American low Arctic (Porslid and Cody 1980)—to a variety of ecological factors using manipulations of equal duration (6 years), in the same vegetation type (mesic birch hummock tundra), and at the same location. We investigated the relative importance of experimental increases in summer temperature, availability of nitrogen (two levels) and phosphorus (factorial N × P additions), snow depth, and caribou exclusion on the production of new apical stems, ramets, and inflorescences in dwarf birch. We focused on new apical stem production since it is highly sensitive to changing conditions in the short term (Chapin and Shaver 1996, Bret-Harte et al 2002), and is a long-term investment component of annual production that we could measure non-destructively in substantially larger sampling areas (9.0 m2) than previous field-based studies (0.04 m2; Chapin et al 1995, Bret-Harte et al 2002). This increased sampling area allows for more robust conclusions given the spatial heterogeneity of shrubs on the tundra landscape. We address the following hypotheses: (1) birch shrub apical stem growth is more responsive to increases in soil nutrient availability than to changes in growing season temperature, snow depth, or caribou herbivory; (2) soil nitrogen is more important than phosphorus in limiting birch apical stem growth; and (3) moderate increases in nitrogen availability enhance birch apical stem growth.
2. Methods
2.1. Study site and treatments
This study was conducted at the Tundra Ecosystem Research Station at Daring Lake, Northwest Territories, Canada (64°52'N, 111°34'W). Temperatures range from a daily mean of − 40 °C in January to 20 °C in July, with approximately 120 days with mean daily temperatures above 0 °C (1996–2010 records; Reid, unpublished data). Mean annual rainfall is 152 mm ± 15 mm and annual snow accumulation is 0.37 m ± 0.17 m in the vegetation type of this study (Buckeridge and Grogan 2008). The research site lies in the centre of the Bathurst caribou (Rangifer tarandus groenlandicus) herd's summer range (Adamczewski et al 2009).
All of our experimental plots were located in mesic birch hummock tundra, in which Betula glandulosa Michx., Rhododendron subarcticum (Harmaja) [formerly Ledum decumbens (Ait.)], Vaccinium vitis-idaea L., Vaccinium uliginosum L., and Eriophorum vaginatum L. are the dominant vascular plants (see Nobrega and Grogan 2008 for further detail). The following four categories of experimental manipulations were all established in the summer of 2004 in four clusters on flat patches of similar birch hummock tundra vegetation within the same 1.6 km long valley (see appendix A for map): (1) nutrient addition plots (5 m × 7 m; n = 5 per treatment) were randomly allocated to one of the following treatments: low N (1 g N m−2 yr−1), high N (10 g N m−2 yr−1), high P (5 g P m−2 yr−1), and high N + P (10 g N m−2 yr−1 and 5 g P m−2 yr−1). The high N and P application levels match those of the comparable acidic tussock tundra manipulations near Toolik Lake, Alaska, and the heath manipulations at Abisko, Sweden. Regular agricultural grade granular ammonium nitrate (NH4NO3) and treble superphosphate (45% P2O5) were applied once yearly during a dry phase in June–July. (2) A warming treatment (n = 5) was established using A-frame greenhouses (1.8 m × 4.7 m; n = 5) covered with heavy polyethylene film (150 μm) that typically reduces photosynthetically active radiation (PAR) by 32% (Chapin et al 1995). The film was put up in early–mid June and taken down in late August–early September each year from 2004 onwards. Small vent holes (20 cm tall) in the apex of each end of the greenhouses prevented extreme temperatures and relative humidity. The greenhouses raised mean summer air temperatures by 2.5 °C (range 0–13.8 °C) and soil temperatures by 2.5 °C and 2.3 °C at 2 cm and 10 cm depth, respectively. (3) Snowfence plots (n = 5) were established 30–60 m apart and oriented perpendicular to the prevailing wind. Snowfences were constructed of heavy duty diamond mesh 15.0 m long and 1.2 m tall and increased snow accumulation 3-fold, from around 0.30 m to 1.0 m (Buckeridge and Grogan 2008). (4) Caribou exclosure plots (n = 5; 19.8 m × 19.8 m) were constructed of metal range fencing (1.2 m tall; aperture 0.15 × 0.20 m) to exclude ungulates but readily permit entry of small herbivores. Although each manipulation had its own set of associated control plots (total = 20), for the purposes of this comparative study, we randomly selected a total of five control plots with at least one from each manipulation.
2.2. Shoot growth measurements
A 3 m × 3 m area was randomly selected within each of the experimental manipulation plots (40 in total), and the lengths of all Betula glandulosa new apical shoots were recorded between 24 July and 7 August 2010. Although this sampling period corresponded with phenological peak plant biomass at the site, plots were measured sequentially through each of the treatments, i.e. control 1, greenhouse 1, high P 1, exclosed 1, high N 1, snowfence 1, low N 1, high N + P 1, followed by the second replicate of each treatment and so on, to prevent a bias that could have resulted if we had sampled all replicates within a treatment before proceeding to the next treatment. Since each sequence of eight plots across the treatments was measured within 2–4 days, we assume that the actual order of measurement of the treatments did not affect our results.
B. glandulosa grows clonally, with low-lying branches becoming covered by moss and producing adventitious roots, and therefore it is difficult to decisively delineate genetically distinct individuals. Considerable care was taken to investigate the connections between branches or the direction of branches under the moss or soil surface, so as to identify individual ramets with high confidence. The number of ramets and structural branches (separate shoots coming up from the moss layer) as well as the current year's inflorescences were also recorded. Four new apical shoots (stem plus leaves) were harvested from random birch plants within each plot, oven-dried, leaves removed, and then stems weighed in order to derive a length to biomass linear relationship for conversion of the field length data (biomass = 0.0075∗length − 0.0013; R2 = 0.8650, p < 0.0001, n = 160, stem biomass in g dry weight, length in cm; see figure B.1). A single equation for length:biomass was used since there were no significant differences in the slopes of the length:biomass relationships between the individual treatments.
2.3. Statistical analyses
In order to test our hypotheses, we ran the following series of ANOVAs on all measured variables: (1) one-way ANOVAs on the primary experimental manipulations (control, warming, snowfence, exclosed, nutrient addition (high N + P)) to identify the primary ecological limitations; (2) two-way ANOVAs on the control, high N, high P, high N + P data to discern the relative importance of each nutrient; (3) one-way ANOVAs on the control, low N and high N data to explore the effect of varying fertilizer levels. Prior to statistical analysis, data were log-transformed if necessary to achieve normality. Post hoc Tukey HSD tests were used to investigate multiple comparisons within each ANOVA. All statistically significant effects (P < 0.05) are reported in the text, and all analyses were performed with JMP 9 (SAS 2010).
3. Results
Of the four primary ecological factors that were manipulated (warming, snowfence, exclosure, high N + P addition), Betula glandulosa new apical stem biomass per unit ground area responded most strongly to the nutrient additions (F4,20 = 11.8, p < 0.001; figure 1). Chronic high level additions of N + P for the previous six years enhanced birch apical stem growth 9-fold relative to the control plots, while the greenhouse warming treatment tended to enhance growth 2.5-fold, but there were no significant effects of deepened snow or caribou exclusion on area-based stem growth. The factorial nutrient additions demonstrated that B. glandulosa apical stem growth was co-limited by N and P (N effect: F3,16 = 6.4, p = 0.022; P effect: F3,16 = 14.2, p = 0.002; figure 1), and the absence of a significant N × P interaction indicates that the effects of the individual nutrients were simply additive. Finally, although the chronic high level N additions enhanced growth 2.5-fold (F2,12 = 12.3, p = 0.001), the low level N additions (1/10th of the above) had no significant impact on growth (figure 1).
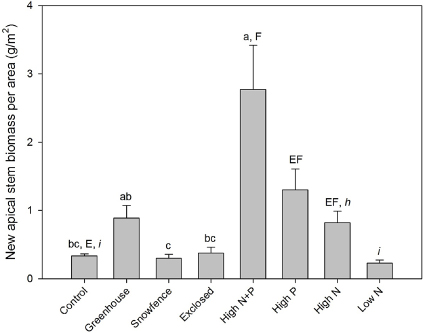
Figure 1. Betula glandulosa new apical stem biomass per unit ground area. Error bars represent standard errors (n = 5). Statistically significant differences (P < 0.05) in the post hoc Tukey tests for each ANOVA are indicated by differences within the following groups of superscript letters: primary manipulations (a–c); N and P factorial additions (E and F); varying levels of N addition (h and i).
Download figure:
Standard imageThe observed increases in Betula new apical stem growth per unit ground area were due principally to enhanced numbers of new shoots (figures 1 and 2(a)). For example, the high level N and P fertilization treatments significantly stimulated initiation of new shoots per unit area (N effect: F3,16 = 4.5, p = 0.049; P effect: F3,16 = 7.4, p = 0.015) resulting in 2-fold increases in shoot number in the separate high N and P treatments compared to that of the control, and a 5-fold increase in the high N + P combined treatment (i.e. an additive effect as above, with no significant interaction). Although high levels of N addition increased the number of new shoots, low N addition levels had no impact (figure 2(a)).
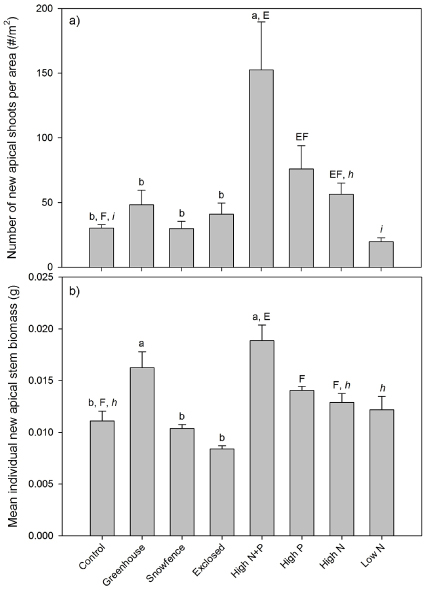
Figure 2. Betula glandulosa number of new apical shoots per unit ground area (a), and mean individual new apical stem biomass (b). Error bars represent standard errors (n = 5). Statistically significant differences (P < 0.05) in the post hoc Tukey tests for each ANOVA are indicated by differences within the following groups of superscript letters: primary manipulations (a–c); N and P factorial additions (E and F); varying levels of N addition (h and i).
Download figure:
Standard imageThe treatments also significantly affected the mean biomass of each new apical stem but to a much lesser extent than their impacts on shoot number (F4,20 = 22.47, p < 0.001; figure 2(b)). Chronic high levels of N + P additions increased mean new shoot biomass 1.6-fold, and the factorial nutrient additions indicated that this response was driven by N and P co-limitation (N effect: F3,16 = 10.90, p = 0.005; P effect: F3,16 = 22.10, p < 0.001), with no significant response to either nutrient alone. Lastly, greenhouse warming increased the mean individual new apical shoot biomass 1.4-fold (figure 2(b)).
The density of ramets (2.0 ±0.5 ramets per m2) and structural branches (10.5 ±0.8 branches per m2) did not differ across the treatments, indicating that all the birch new apical stem growth responses reported above occurred as a result of changes in growth on existing shoots rather than through new ramet or structural branch production.
Inflorescence production was also substantially enhanced in the high N + P addition plots (F4,20 = 5.13, p = 0.005; figure 3). While the factorial additions demonstrated that both nutrients affected inflorescence production (N effect: F3,16 = 4.82, p = 0.043; P effect: F3,16 = 3.38, p = 0.085), the magnitudes of these effects were minimal compared to the substantial increases that occurred when both nutrients were added (N × P interaction: F3,16 = 7.13, p = 0.017).
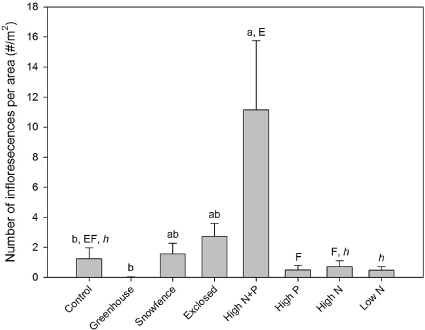
Figure 3. Betula glandulosa inflorescences per unit ground area. Error bars represent standard errors (n = 5). Statistically significant differences (P < 0.05) in the post hoc Tukey tests for each ANOVA are indicated by differences within the following groups of superscript letters: primary manipulations (a–c); N and P factorial additions (E and F); varying levels of N addition (h and i).
Download figure:
Standard image4. Discussion
4.1. Co-limitation of new apical stem growth by N and P, with strong effects of P
The factorial nutrient additions clearly demonstrate that Betula glandulosa apical shoot productivity at our site was co-limited by N and P. The substantial increases in growth we documented with high N + P additions are similar to those observed in many past studies of the effects of fertilization on tundra plants (Chapin et al 1995, Press et al 1998, Shaver and Jonasson 1999, Bret-Harte et al 2002, van Wijk et al 2004). Comparison across the primary ecological manipulations at our site indicates that birch apical stem growth was more responsive to the substantial increases in soil nutrient availability than to our manipulations of growing season temperature, snow depth, or caribou exclusion. Thus, hypothesis 1 is supported, at least for the high level of fertilization tested.
As the first study of factorial annually repeated high N × P additions over more than two years in arctic tundra, our data allowed us to investigate the influence of each element separately and across many growing seasons. The results clearly indicate that the availability of P was at least as important as N in limiting birch apical stem growth in mesic arctic tundra, refuting hypothesis 2. These results contribute to the increasing body of research (see Giesler et al 2012 and citations therein) challenging the widely held perspective amongst arctic terrestrial ecologists that plant growth in mesic tundra ecosystems is primarily N-limited. That perspective seems to be based largely on Shaver and Chapin's (1980) factorial nutrient addition study, in which the results may be a function of the experiment's short duration. The authors concluded that growth of a range of tundra species was 'stimulated most strongly by (high) N and/or N + P fertilization' based on a single set of fertilizer additions in early summer of one year, and harvests in the late spring and late summer of the following year. High N and N + P treatments had significant effects on new shoot growth of a range of species in both harvests, but the magnitudes of the N × P interaction effect were substantially larger in the final harvest, and most importantly, the effect of added P alone only became statistically significant at the final harvest (see table 6 in Shaver and Chapin 1980). The stronger and earlier effect of N compared to P has been widely interpreted as evidence that tundra plant growth is primarily limited by N availability—Shaver and Chapin (1980) has been highly cited in this context (292 citations; ISI Web of Science 30 May 2012). Two other factorial N + P studies found that growth in separate upland dwarf shrub communities was primarily N-limited (Haag 1974, Baddeley et al 1994), but these studies were also of short duration, having been conducted within a single growing season and across one growing season, respectively. Phosphorus is relatively insoluble compared to N, especially in acidic soils, and addition over a single year may not have resulted in sufficiently enhanced plant accessible pools to overcome incipient limitation. As suggested by the emergence of a significant interactive P effect only in Shaver and Chapin's (1980) final harvest, our repeated P additions over six years indicate that shoot growth of birch plants is at least limited by the availabilities of both N and P.
In contrast to mesic tundra ecosystems, P limitation or N and P co-limitation of plant growth has been well documented in wet sedge tundra ecosystems (Haag 1974, Shaver and Chapin 1995, Shaver et al 1998), mesic meadow (Giesler et al 2012), and dry heath (Gough et al 2002) on the basis of single as well as repeated N × P factorial additions, or by inference from plant tissue N:P ratios. These studies have led to the overall conclusion in a global meta-analysis that, similar to other terrestrial ecosystems, tundra productivity is co-limited by N and P, and yet more responsive to added N than to added P (Elser et al 2007). However, the authors do caution that there were only seven studies available for arctic tundra, most of which were located on inundated wet sedge ecosystems, and only two of which had factorial nutrient additions. Our study helps address the almost complete lack of data for mesic ecosystems, challenges the relative importance of N and P in terms of the magnitudes of plant responses, and lends strong support to the conclusion that N:P stoichiometry is a fundamental control on the functioning of all ecosystems (Elser et al 2007).
Further investigations are required to determine whether the P limitation of birch growth reported here extends to other species and other sites. The overall plant community response to factorial additions of N and P at our site is currently being analysed. In terms of inter-site variability, the soils at our site are young, having developed since the last glaciation, suggesting that soil P availability may be higher than on older soils such as Alaskan acidic tussock tundra. Nevertheless, the soil's acidity at our site is similar to the latter and many other mesic tundra locations (pH 4.3; Chu and Grogan 2010), indicating that P availability is likely to be commonly limiting due to its occlusion in iron- and aluminum-based compounds (Schlesinger 1997, Vitousek et al 2010). Furthermore, soil solution phosphate and microbial P pool sizes in our birch hummock ecosystem (Buckeridge and Grogan 2008, Chu and Grogan 2010) are very similar to values reported for heath tundra in north Sweden (Jonasson et al 1999) and higher than values for acidic tussock tundra in Alaska (Giblin et al 1991), together suggesting that P limitation of shrub growth may be quite common across the low Arctic.
4.2. Threshold for effect of enhanced nitrogen availability
The lack of a response in Betula new apical stem growth to low level N additions refutes hypothesis 3 and indicates that there is a nitrogen availability threshold that must be crossed before apical shoot growth is affected. Previous experiments on varying levels of N addition in mesic arctic tundra found either no change in total shrub biomass from 0 to 5 to 25 g N m−2 added (Salix arctica; Henry et al 1986) or a linear increase in shrub leaf biomass from 1 to 5 g N m−2 yr−1 added (Salix polaris; Baddeley et al 1994), following one and two years of fertilization, respectively. Although Betula tends to respond more dramatically than Salix to nutrient additions (Chapin et al 1995), the lack of any Betula apical stem growth impacts after our six years of low level N addition indicates that Betula may be relatively unresponsive to small but nevertheless realistic changes in soil nitrogen availability as the climate warms. While warming is predicted to increase soil fertility in the Arctic (Nadelhoffer et al 1991, Chapin et al 1995), the magnitude of that increase is highly speculative (Hobbie et al 2002). Even high projections for increases in soil fertility with warming (e.g. Mack et al 2004) would happen gradually and may elicit much different plant responses than from the instantaneously high soil fertility generated by chronic high level nutrient addition manipulations (Chapin et al 1995). In this context, there is no doubt that the levels of nutrient amendment tested with the high N and P treatments are well beyond what will probably be observed with climate warming. However, the factorial experiment is important to our understanding of the relative importance of these two nutrients to tundra plant growth. Recent reviews indicate that interpreting the mechanisms underlying co-limitation by N and P is complex (e.g. Ågren et al 2012), but together our results suggest that the absence of a shoot response with low N addition may have been because growth was as limited by the availability of P as of N, while the response to high N may be a consequence of N-enhanced resources becoming available (e.g. particular enzymes) that promoted P acquisition.
4.3. Substantial impacts of experimental warming relative to past studies
The 2.5-fold increase in B. glandulosa new apical stem growth per unit area with greenhouse warming was a substantially stronger experimental warming response than the ∼1.5-fold increase in apical growth or total biomass that has been documented in many past studies (Chapin et al 1995, Jonasson et al 1999, Bret-Harte et al 2001, Dormann and Woodin 2002). Although the increase in air temperatures in our greenhouses was similar to or smaller than past greenhouse studies (Chapin et al 1995, Jonasson et al 1999), we had a substantially stronger soil warming impact. Mean temperatures were raised by around 2.5° over the 0–10 cm soil depth interval where most of the root biomass occurs (Churchland et al 2010), presumably resulting in increased nutrient mineralization (Nadelhoffer et al 1991, Jonasson et al 1999), and enhanced availability to the plants. Together, this pattern of results is consistent with a strong indirect effect of warming on tundra plant growth that is mediated through enhanced soil nutrient availability.
Nonetheless the lack of response to the low N addition indicates that this substantial increase in growth in the greenhouses cannot be explained by moderate increased in soil nitrogen alone. While our factorial experiment indicates that increases in soil phosphorus may be equally as important, it is likely that the greenhouse response is due to a combination of warming-induced increases in soil N and P, as well as direct effects of warming on rates of photosynthesis. The relative importance of increased photosynthesis and temperature-enhanced soil fertility in arctic tundra remain unclear, in part due to the lack of experiments specifically aimed at warming either soils or plant shoots, but also due to high variation in responses between species and systems. For example, experimental warming has increased chlorophyll content in Cassiope tetragona but not in Empetrum hermaphroditum (Michelsen et al 1996), and increased photosynthesis per unit leaf area in Dryas octopetala (Wookey et al 1995) but not in Polygonum viviparum (Wookey et al 1994). At the landscape-scale, responses of gross ecosystem photosynthesis (GEP) and ecosystem respiration (ER) to experimental warming have been variable across microtopography (Grogan and Chapin 2000), sites and soil moisture gradients, with a tendency for increases in both GEP and ER in dry systems and decreases in GEP with no change in ER in moist or wet systems (Welker et al 2004, Oberbauer et al 2007). The impact of experimental warming on soil fertility is similarly variable between systems. For example, at Toolik Lake, Alaska, greenhouses increased extractable ammonium (NH4) in organic soils but had no effect on nitrate or phosphate (Chapin et al 1995), while at Abisko, Sweden, soil inorganic P was increased at a heath site but not at a higher elevation fellfield site, and NH4–N was increased at the fellfield site but not at the heath site (Jonasson et al 1999). Detailed biogeochemical analyses of the soils and plant community in our plots are currently underway to identify the impact of experimental warming at our site on soil nutrient pools, so that we can better determine the likely importance of warming-induced increases in nutrient availability as compared to direct effects of warming on plant photosynthesis.
4.4. No effect of snow depth or caribou exclusion on apical stem growth
The lack of a snowfence effect on the production of new apical shoots contrasts with the predictions of the snow–shrub feedback hypothesis that increased snow depth beneath the shrubs due to the trapping of blowing snow would lead to increased winter soil temperatures and enhanced nutrient cycling, enabling increased shrub growth (Sturm et al 2001). Our snowfence treatment consistently raises peak snow depth from around 0.3 to 1 m and substantially reduces winter soil cooling (Buckeridge and Grogan 2008). This manipulation increased late winter soil microbial nitrogen (Buckeridge and Grogan 2008) and led to very large late thaw pulses of ammonium, phosphate and dissolved organic nitrogen and phosphorus (Buckeridge and Grogan 2010), but did not affect isotopic nitrogen acquisition by any plant species two years after 15N addition (Vankoughnett 2010). The birch apical shoot data reported here reflect six years of snowfence treatment, indicating that if a snow depth related feedback between soil nutrient availability and Betula growth occurs, it must be a longer-term process.
Although we observed no impacts of caribou exclusion on shrub apical stem growth, we found that birch leaf biomass was 50% lower in the control plots than exclosed plots over the same time period (Zamin and Grogan, unpublished data) indicating that some herbivory was occurring. Caribou herbivory impacts are largely a function of population size, which affects range use and browsing intensity, and although caribou populations fluctuate naturally (Zalatan et al 2006), many herds, including the Bathurst herd in our region, have recently been in severe decline (Adamczewski et al 2009, Vors and Boyce 2009). Over the study period, the Bathurst herd population varied from 28% of its previous population maximum (2004) to 7% of its previous size and its lowest size on record (2009; Adamczewski et al 2009); corresponding to a population density of around 0.25–1.3 animals km−2. Consequently, any caribou browsing at our site over the study period was clearly low intensity. Whether caribou browsing will become a more important control on Betula glandulosa productivity depends on how the populations cope with ongoing changes in food quality and accessibility, insect harassment, extreme weather events, industrial development and hunting (Post and Forchhammer 2008, Adamczewski et al 2009, Vors and Boyce 2009).
4.5. Mechanisms for shrub expansion
The greenhouse and nutrient addition results indicate that the dominant mechanisms for increased birch shrub growth in early responses to warming may be enhanced meristem initiation on existing shoots (i.e. conversion of 'short shoots' to 'long shoots') and enhanced growth of each new shoot rather than the production of new individuals from seed or of new structural branches from the belowground stems. Investment in vegetative growth appears to take priority over investment in reproduction, the latter of which is only initiated once both N and P limitations have been removed. This trade-off between investment in apical stem growth and inflorescence production, specifically observed in our warming treatment, could reduce the possibility for dispersal and adaptation, and affect bird populations (e.g. redpolls; Carduelis flammea L. and Carduelis hornemanni Holboll) that depend heavily on this food source (Brooks 1968). Nonetheless, at Toolik Lake a trade-off was observed between stem length and number, where fertilization increased the length of individual stems in Betula nana for several years, after which stem lengths declined to pre-treatment levels, coincident with an increase in the number of new stems (e.g. increased branching; Bret-Harte et al 2002). Meanwhile at our site, high N + P fertilization led to an increase in both the number of new shoots and the biomass of individual new apical stems of B. glandulosa, indicating that this species is capable of increasing both growth parameters when conditions allow.
Increases in shrub vertical growth in the Arctic may be limited by the depth of the snow pack, as snow provides protection from winter desiccation and wind abrasion (Sturm et al 2001). However, since Betula is at a low density throughout our site (2.0± 0.5 individuals per m2) and is the tallest plant in the community, there is ample room for horizontal expansion. Repeat measurements of birch shrub dimensions in unmanipulated plots in a variety of topographical locations at our site over the past five years indicate no change in height, but small increases (∼14%) in lateral extension (Grogan, unpublished data).
Our results interpreted in the context of understanding potential shrub growth responses to climate change indicate that warming, including its direct and indirect effects, can significantly enhance Betula shoot growth. Our greenhouse warming treatment increased dwarf birch new apical stem growth by a factor of 2.5, while the lack of response to the low N addition treatment indicates that moderate increases in N alone that might be expected under climate warming are unlikely to explain shrub growth responses. The absence of a low N addition response may be consistent with the factorial high N and P experiment results which clearly indicate that birch shoot growth is co-limited by the supply of nitrogen and phosphorus. We conclude that birch shrub expansion at our site will be driven primarily by enhanced growing season temperatures, and specifically by a combination of increased photosynthesis and warming-enhanced supply of nitrogen and phosphorus. There is then a need to determine the extent of N–P co-limitation of tundra plant growth by investigations at other mesic sites over at least a 5 yr period. Finally, our results point to the need to improve our understanding of how warming and associated aspects of climate change will affect photosynthesis as well as the biogeochemical cycling of these two very different nutrients if we are to develop more accurate predictions of vegetation change across the Arctic.
Acknowledgments
We thank Sarah Farrow and Linda Cameron for field and lab assistance, Mike Treberg, Robbie Hember, Peter Lafleur, and Greg Henry for support in establishing the experimental manipulations, Steve Matthews (GNWT), Chandra Venables (GNWT) and the Aurora Research Institute for logistics, and NSERC (TZ and PG), NSF (PG), DIAND (TZ) and the Ontario Provincial Government (TZ and PG) for funding.
Appendix A:
Figure A.1. Map of plots used in this study; Daring Lake Tundra Ecosystem Research Station, NWT, Canada.
Download figure:
Standard imageAppendix B:
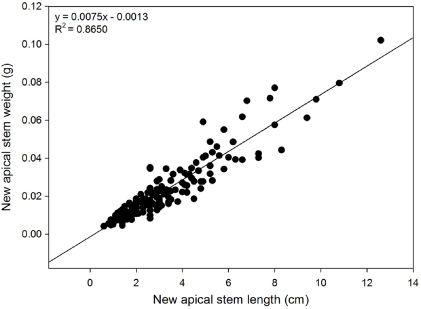
Figure B.1. Allometric relationship between length and biomass of Betula glandulosa new apical stems.
Download figure:
Standard image