
Abstract
The agronomical relevant tomato-Pseudomonas syringae pv. tomato pathosystem is widely used to explore and understand the underlying mechanisms of the plant immune response. Transcript abundance estimation, mainly through reverse transcription-quantitative PCR (RT-qPCR), is a common approach employed to investigate the possible role of a candidate gene in certain biological process under study. The accuracy of this technique relies heavily on the selection of adequate reference genes. Initially, genes derived from other techniques (such as Northern blots) were used as reference genes in RT-qPCR experiments, but recent studies in different systems suggest that many of these genes are not stably expressed. The development of high throughput transcriptomic techniques, such as RNA-seq, provides an opportunity for the identification of transcriptionally stable genes that can be adopted as novel and robust reference genes. Here we take advantage of a large set of RNA-seq data originating from tomato leaves infiltrated with different immunity inducers and bacterial strains. We assessed and validated 9 genes that are much more stable than two traditional reference genes. Specifically, ARD2 and VIN3 were the most stably expressed genes and consequently we propose they be adopted for RT-qPCR experiments involving this pathosystem.
Similar content being viewed by others


Introduction
Gene expression quantification is an important and widely used technique that allows analyzing the state of different cellular processes in specific conditions. Nowadays, reverse transcription-quantitative PCR (RT-qPCR) is the tool most frequently used to determine the mRNA levels in different biological systems1. Because of its sensitivity, accuracy and rapid execution, it is considered the most important mid-throughput gene expression analysis technology2. However, there are several critical steps during the execution of RT-qPCR experiments that affect the accuracy and interpretation of the results, such us the quality of the mRNA, amplification efficiency and the choice of reliable internal controls referred to as reference genes2,3.
Normalization of the sample expression with reference genes is used to account for the technical variation produced during the processing of the samples. A reference gene is an internal control that should have minimal or no variation of its expression in the analyzed conditions2,4. Traditionally, few reference genes are used in plants such as beta-tubulin-4 (TUB4), glyceraldehyde-3-phosphate dehydrogenase (GADPH), 18S ribosomal RNA (18S RNA), polyubiquitin (UBQ), actin (ACT), elongation factor 1 alpha (EF1α)4. Because of their relatively high expression levels in all kinds of cells or tissues, these genes were initially selected as reference genes for qualitative (Northern blot) and semi-quantitative (RT-PCR) approaches and have been widely adopted for RT-qPCR experiments5. However, several recent studies indicate that these traditional reference genes are not very stably expressed in different experimental conditions and have encouraged the systematic selection and validation of better RT-qPCR reference genes previous to performing expression level measurements2,5,6,7,8.
Plants detect and react to pathogens using a two-layer defense mechanism. Pattern-triggered immunity (PTI) is activated after the detection of microbe/pathogen-associated molecular patterns (MAMPs or PAMPs) by membrane receptors called pattern-recognition receptors (PRRs)9,10,11. Some bacterial pathogens use a type III secretion system to introduce effector (virulence) proteins into the cell cytoplasm to undermine PTI12. During evolution, some plants developed the ability to recognize the activity of these effectors and activate a second layer of immunity named effector-triggered immunity (ETI)12,13,14. Large changes in gene expression occur during the development of both immune responses15,16,17,18.
The interaction between tomato (Solanum lycopersicum) and the causal agent of bacterial speck disease, Pseudomonas syringae pv. tomato (Pst), is considered a model for the study of molecular mechanisms leading to plant defense responses. The perception of bacterial flagellin by tomato has been well characterized. This protein contains two MAMPs that are detected by tomato: flg22 and flgII-28, recognized by FLS2 and FLS3 receptors, respectively19,20,21,22. It has been reported that the primary PTI elicitors from Pst in tomato are the flagellin-derived MAMPs and this perception results in extensive transcriptional changes17. Around 30 effectors are introduced into plant cells by Pst strain DC300023. Among them, AvrPto and AvrPtoB are early-acting effectors that suppress PTI by interfering with PRR-mediated signaling and thereby promote bacterial virulence17,24,25. In some tomato lines, members of the Pto kinase family detect and interact with AvrPto and AvrPtoB effectors and jointly with the nucleotide binding-leucine rich repeat (NB-LRR) protein Prf activate ETI26,27,28,29. Changes in tomato gene expression that occur during Pto/Prf-mediated ETI were previously studied18,30.
Recently, several tomato genes have been evaluated and identified as the most suitable RT-qPCR reference genes in different experimental conditions. For example, there are now reference genes available for tomato fruit development31, tomato seeds under different conditions32 and MicroTom-Rg1 genotype fruit33. Similar studies have been conducted in tomatoes under abiotic stresses34 and biotic interactions, such as host responses to viruses35,36,37 and Xanthomonas campestris pv. vesicatoria (Xcv)38. Theses studies support the idea that there is not a single reference gene that can be used for a given species, and consequently these need to be evaluated and selected for each particular expression study.
In most of these tomato publications, the selection of the candidate reference genes was based on genes previously used for the same species in different experimental conditions or for other phylogenetically related plants. Alternatively, in some cases, the authors used transcriptional expression data generated by microarray analysis to identify novel and more stably expressed genes when compared to traditionally employed reference genes38. In the past years, RNA-seq has emerged as a powerful high-throughput technology used for transcriptome analysis in different organisms and treatments39,40,41,42,43. In spite of being a technique used for many years, RNA-seq data has been used in the plant research field for the selection and validation of new and more robust RT-qPCR reference genes only in grape, soybean and Lycoris44,45,46.
Previously, we used an RNA-seq approach for the analysis of PTI activation in tomato and the subsequent inhibition of this response by Pst effectors AvrPto and AvrPtoB17. Additionally, we identified genes that are differentially expressed specifically during activation of PTI or ETI in tomato18. Here, we have taken advantage of the large set of RNA-seq data mentioned above in addition to newly generated data that complements the publicly available set, for the selection of 9 candidate genes with the lowest variation within a total of 37 different treatments/time-points and their biological replicates. We then performed RT-qPCR experiments using tomato leaf tissue infiltrated with different Pseudomonas species and mutants to study their behavior upon activation of plant defense. Validation of these genes was performed using three different tools (geNorm, NormFinder and Bestkeeper), and compared with two traditional housekeeping genes and the most stably expressed gene identified during the analysis of tomato infection with Xcv38. Our results identified a set of novel reference genes that are transcriptionally more stable than the traditional ones and consequently we propose their use in experiments involving tomato-Pseudomonas pathosystem.
Results
Selection of tomato genes with stable expression using RNA-seq data
In order to identify genes whose expression has a low variation across different treatments, we took advantage of the RNA-seq expression data previously published17,18 and newly generated additional data. Treatments are described in Supplementary Table S1 and include different bacterial strains and MAMP infiltrations, along with mock treatments and untreated leaf tissue at different time-points. In all these experiments, the leaf tissue was collected at 30 min, 4 and 6 h after infiltration (hai) to investigate early changes in host gene expression (Supplementary Table S1).
In order to rank the predicted 34,725 tomato genes (ITAG 2.447) based on their transcript level stability, we calculated the variation coefficient (VC) using all the RPKMs (reads per kilobase of transcript per million mapped reads) determined in each experimental condition (37 different conditions, Supplementary Table S2) using the biological replicate information individually. This accounted for 110 total values. The lower the VC is, the more stable the expression of the gene is across all the conditions. In this way we selected 9 genes with the lowest VC for analysis, none of which had been used previously as tomato reference genes in RT-qPCR assays (Supplementary Table S2). For this set of genes, VC ranged from 12.2% to 14.4%. We also selected a gene named PHD (Solyc06g051420, VC 31.5%) previously identified and validated reference gene in tomato-Xanthomonas campestris pv. vesicatoria (Xcv) pathosystem38 and two traditional plant reference genes EF1α (Solyc06g005060, VC 41.6%) and GADPH (Solyc04g009030, VC 52.9%). Gene expression variability across all the RNA-seq treatments of the selected genes was globally analyzed in a log2(RPKM) box plot graph (Supplementary Fig. S1). Relative box and whisker sizes indicated low gene expression stability of PHD, GAPDH and EF1α.
Expression profiles of candidate reference genes showed good amplification efficiencies and primer specificities
We performed RT-qPCR using cDNA dilutions (1:5, 1:10, 1:100, 1:1000). Amplification efficiency E was measured as 10−1/slope and expressed in percentage (Supplementary Table S3). All the primers designed in this work showed high amplification E values ranging from 89% to 117%. Another important aspect to be evaluated is the specificity of the amplification. To achieve this, we performed melting curves for all the pair of primers used and in all cases observed a single peak corresponding to a single amplification product (Supplementary Fig. S2).
Cycle amplification values (Cq) indicate a wide range of expression levels among the selected tomato reference genes
We designed an experiment aimed at evaluating the performance of our set of genes under different plant immune responses. Therefore, in order to activate PTI we infiltrated tomato Rio Grande (RG)-PtoR leaves with Pseudomonas fluorescens 55 (Pf)48 and 10 mM MgCl2 as a mock treatment. Also, we infiltrated the RG-PtoR tomato leaves with Pst DC300049 to activate PTI and ETI and the double mutant Pst DC3000 ΔavrPto ΔavrPtoB50 to induce the development of bacterial speck disease (Table 1). We collected leaf tissue from 3 biological replicates at early time points, 6 and 12 hai, and monitored the development of symptoms in these plants at later time points to confirm the activation of the expected plant responses.
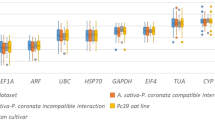
Average Cq values, ranged from 14.9 (EF1α) to 26.6 (VIN3) (Fig. 1) indicating most of the genes (except for EF1α) have a Cq value that is within the recommended values for a RT-qPCR reference gene (higher than 15 and lower than 30)4. Moreover, GADPH expression levels were the most variable, with minimum and maximum values of 17.1 and 22.5, respectively. This represents a difference of 5.4 Cq between them. Importantly, this difference (max Cq - min Cq) was much smaller, ranging between 1.4 and 1.8, for the genes identified in this work.

Box and whisker plot graph showing Cq values of each selected gene in all the samples analyzed (n = 24). Black lines and boxes represent the medians and 25/75 percentiles, respectively. Whisker caps represent the minimum and maximum values. Ο, indicates outliers.
Different algorithms indicate ARD2 and VIN3 are the most stable reference genes
To determine which of the selected genes had the most stable expression levels in our system, we analyzed RT-qPCR data with three different tools to estimate gene expression stability. We first determined the average expression stability value M using geNorm software51. This program calculates the pairwise variation of each reference gene with all other genes analyzed under the same experimental conditions. In this way, the lower the M value, the more stable the gene is. All the analyzed genes, presented M values lower than the usually proposed cutoff value of M ≤ 0.5. The highest variability was observed for GADPH (M = 0.204), EF1α (M = 0.178) and PHD (M = 0.162) (Fig. 2A). The algorithm also selects an optimal pair of reference genes and in our case the most stable ones were ARD2 and VIN3 with an M value of 0.092.

(A) Tomato reference genes were ranked based on expression stability calculated by geNorm. M values represent the average pairwise variation of the gene compared with all other control genes. (B) Pairwise variation (Vn/Vn + 1) for determination of the optimal number of reference genes. The pairwise variation was calculated considering all the samples together (Total), mock and Pf inoculations (PTI), Pst DC3000 and Pst DC3000 ΔavrPto ΔavrPotB (ETI), samples taken at 6 hpi (6 h) or samples taken at 12 hpi (12 h).
We determined the pairwise variation (V) of a normalization factor (NF) calculated by introducing reference genes one by one, starting from the two least variable until the whole set was included. With this approach, the optimal number of reference genes to be used can be estimated. We analyzed our data as a whole, only including PTI activation (Pf 55 and mock), only including ETI activation (Pst DC3000 and Pst DC3000 ΔavrPto ΔavrPtoB), only including 4 hai or only including 12 hai (Fig. 2B). Regardless of the plant response activation or time-point, V2/3 value is considerably smaller than the proposed cut-off (<0.1551), suggesting that using only the two most stable reference genes (ARD2 and VIN3) is sufficient for normalization. The addition of GAPDH (V11/12), the least stable gene, resulted in a particularly large increase of the variation parameter V when analyzing subsets that include ETI induction (Fig. 2B). To look into this phenomenon, we analyzed the individual Cq values for each gene in all conditions (Supplementary Fig. S3). We observed a clear Cq value increase of GADPH when ETI is activated (infiltration with Pst DC3000) at 6 and 12 hai, suggesting down regulation of the corresponding transcript. This result indicates GADPH is not a suitable reference for experiments involving ETI activation.
To further investigate the gene expression stability of the selected genes in our experiments, we analyzed our data with NormFinder52. This algorithm also calculates an M index, but taking into account the intragroup (within each sample/treatment) and the intergroup variation (within different groups of samples/treatments). This analysis revealed similar results than geNorm (Fig. 3). The most suitable reference genes derived from NormFinder analysis were VIN3, ARD2 and KLC with M values of 0.013, 0.016 and 0.019, respectively. On the other hand, GADPH, EF1α, and PHD were among the least stable genes.

Tomato reference genes were ranked based on expression stability calculated by NormFinder. The analysis was performed using expression data from all biological replicates and treatments (n = 24).
The other tool we used to study candidate gene stability, BestKeeper, allows the analysis of up to 10 reference genes53. For this reason, we included in the analysis the top 7 most stable candidate genes based on NormFinder analysis, PHD and the 2 classical reference genes. This tool performs the analysis in two steps. First, it estimates different statistical parameters that allow determining if a gene has an acceptable overall variation to be considered a reference gene (SD [±Cq] < 1 and SD [±x-fold] < 2). All the studied genes, except GADPH, passed this filter (Table 2). Then, a matrix of pairwise comparisons and coefficient of correlation (r) calculation are performed to obtain a BestKeeper index. The r value obtained from the comparison of each gene with this index allows establishing a ranking of reference gene suitability. Higher gene expression stability is associated to r values closer to 1. Our results indicate that APX, followed by ARD2 and VIN3 posses the higher correlation coefficients being the most stable genes of the 10 analyzed (Table 2). Again, EF1α and PHD were ranked as among the least stable genes with r values of 0.061 and 0.125, respectively.
Although the results obtained in this study were largely consistent when comparing the outputs of the statistical programs used, a few discrepancies were observed. It has been proposed that geNorm, NormFinder and BestKeeper tools tend to generate distinct ranking orders of reference genes because they are based on different algorithms35. Therefore, it is recommended to consider them as complementary statistical methods and analyze results globally. Thus, we calculated the arithmetical mean of the ranking value obtained for each gene using all three algorithms35. As expected, ARD2 and VIN3 were rated as the most stable with a mean value of 1.67 (Table 3).
Validation of the selected genes confirmed their suitability as reference genes
To validate the selection of reference genes, we measured the expression of a PTI- and an ETI-specific gene that were previously reported18. As recommended51, we estimated the relative expression using the normalization factor NF calculated as the geometric mean of Cq values obtained for ARD2 and VIN3, the most stable reference genes. Alternatively, we selected the worst condition, which is using the least stable gene (GADPH) as the only reference.
In the case of the PTI-specific marker (Solyc02g069960), we saw the expected increase of gene expression in the samples infiltrated with P. fluorescens 55 (Fig. 4A) at both 6 and 12 hai, regardless of the reference gene used. Although the trend is the same, normalization with an unsuitable reference gene such as GADPH, not only increased gene expression levels, but also resulted in larger standard deviation values.

RT-qPCR of (A) PTI-reporter gene (Solyc02g069960) at two time points (6 and 12 hai) with mock (10 mM MgCl2) or 108 cfu/ml of Pseudomonas fluorescens 55 (Pf) and (B) ETI-reporter gene (Solyc09g092500) at two time points (6 and 12 hai) with 5 × 106 cfu/ml of Pseudomonas syringae pv. tomato DC3000 (Pst DC3000) and Pst DC3000 ΔavrPto ΔavrPtoB (ΔΔ) strains. In both cases, the geometric mean of the two best (ARD2/VIN3) or the worst (GADPH) reference genes were used for normalization of the data. Bars represent the mean of three biological replicates and three technical replicates with their corresponding standard deviation. ** or * indicate significant differences using Student t-test with p-values < 0.01 or <0.05, respectively.
As anticipated, we observed an increase in ETI marker gene (Solyc09g092500) expression in RG-PtoR tomato leaves infiltrated with Pst DC3000 compared with Pst DC3000 ΔavrPto ΔavrPtoB (Fig. 4B). Nevertheless, the expression pattern was quite different if the data was analyzed using ARD2/VIN3 or GAPDH as reference genes (Fig. 4B). As observed for the PTI reporter gene, the activation of the ETI marker gene was over-estimated. Additionally, the gene expression reduction between 6 and 12 h previously reported18, could not be observed when using GAPDH normalization. Again, the combined use of ARD2/VIN3 leads to a drastic reduction in standard deviation values.
To further investigate the influence of using a non-stably expressed gene as reference in RT-qPCR experiments, we analyzed ARD2 expression using VIN3 or GADPH as reference gene (Fig. 5). In this analysis, we show that ARD2 is expressed with remarkably small variation across the different experimental conditions when using VIN3 as a reference. However, when we normalized the data with GADPH the same gene falsely increased its expression upon plant defense activation. This was more evident in the case of the ETI-inducing treatment (Pst DC3000), which can be explained by the noticeable down-regulation of GADPH gene expression we observed in the samples infiltrated with Pst DC3000 (Supplementary Fig. S3). Therefore, our results support the importance of the selection and validation of accurate reference genes RT-qPCR to avoid misinterpretation of the expression data.

RT-qPCR of ARD2 expression in tomato leaves infiltrated with mock (10 mM MgCl2), 108 cfu/ml of Pseudomonas fluorescens 55 (Pf), 5 × 106 cfu/ml of Pseudomonas syringae pv. tomato (Pst) DC3000 (DC3000) and Pst DC3000 ΔavrPto ΔavrPtoB (ΔΔ) strains. Samples were taken at two time points (6 and 12 hai). VIN3 (best) or GADPH (worst) reference genes have been used for normalization of the data. Bars represent the mean of three biological replicates and technical replicates with their corresponding standard deviation.
Discussion
RT-qPCR is a powerful technique for gene expression detection and quantification, but the accuracy and reliability of the results highly depend on appropriate data normalization4. In this sense, several reports in different plant species like Arabidopsis54, soybean45, rice55, cotton56 among others, have supported the importance of identifying stably expressed genes for each species, tissue, treatment or condition to be analyzed.
As a new approach for the tomato-Pseudomonas pathosystem, we have taken advantage of previously published RNA-seq data17,18 for the selection of stably expressed genes. In both studies, different infiltrations were performed in tomato leaves aiming at analyzing transcriptional changes during PTI and ETI activation, and the influence of bacterial effectors on plant defenses. To complete this transcriptomic set of information, we performed new experiments that include untreated tomato plants and infiltrations with additional MAMPs (flagellin and non-flagellin derived) and bacterial strains and mutants. Together these data formed a robust set of gene expression information (37 different treatments/time points with an average of 3 biological replicates generated in independent experiments, Supplementary Table S2) that allowed us to select genes with low variation coefficients in the tomato-Pseudomonas pathosystem. To our knowledge, this study uses the largest set of RNA-seq data to date to identify reference genes.
For validation of our set of reference genes using RT-qPCR we selected treatments that involve a strong activation of transcriptomic changes. For example challenges with different bacterial strains at the concentration and time-point used lead to approximately 2,800 and 5,700 genes differentially expressed for PTI17 and ETI18, respectively. With this in mind, we are confident that we tested our candidate genes under rigorous conditions for steady gene expression. In addition, it is worth noting that these infiltration experiments were performed independently from the RNA-seq ones, adding even greater strength to our results. We employed three widely used tools for the evaluation of gene expression stability such as geNorm51, NormFinder52 and Bestkeeper53, to test our reference gene candidates. Our analysis suggests that all 9 selected genes from the RNA-seq data are more stable than the ones commonly used in the literature.
Based on our results, we strongly recommend the use of ARD2 and VIN3 as the most suitable reference genes for gene expression studies in tomato leaf interactions with Pseudomonas (Table 3). Pairwise variation analysis that geNorm program performs, established that the use of these two genes is sufficient to obtain consistent results. These two selected genes were consistently grouped among the most stable ones, and the traditional GADPH and EF1α were included within the least stable group.
The interaction of tomato with Xanthomonas campestris pv. vesicatoria (Xcv) is another commonly used model system for studying plant-pathogen biology57. In a recent work, two genes (PHD and LSM7) were recommended, based on microarray data identification, for normalization in tomato gene expression assays of plants infected with Xcv38. The authors found that GADPH was particularly not a suitable reference gene for this pathosystem, due to its down regulation upon Xcv challenge. In our case we observed this same effect in GADPH transcript levels when AvrPto/AvrPtoB-mediated ETI response was activated (Supplementary Fig. S3). Contrastingly, GADPH was ranked as one of the most stable candidates analyzed for the pathosystem Actinidia deliciosa-Pseudomonas syringae pv. actinidiae58. To test how a tomato-Xcv suitable reference gene would perform in our system and compare its stability to our set of candidates, we included PHD38. In spite of PHD performing better than the traditional reference genes used in our analysis, all the genes we selected based on RNA-seq data were found to be more stably expressed in the tomato-Pseudomonas pathosystem. These findings support the idea that reference genes need to be identified and tested for each specific system.
To put our selected reference genes to test, we performed RT-qPCR experiments to investigate transcript levels of PTI- and ETI-specific reporter genes previously identified18. The comparison of the gene expression values obtained when normalizing the data with the combination ARD2/VIN3 or GADPH was highly discordant in terms of estimated transcript levels and standard deviations (Fig. 5). This result also emphasizes the importance of the selection of appropriate reference genes to avoid misinterpretation of experiments and further confirmed that GADPH is not a good reference gene for expression studies in the tomato-Pseudomonas interaction.
In conclusion, by using a large RNA-seq data set we were able to identify and validate highly stable RT-qPCR reference genes. We recommend the use of these genes for gene expression analyses of tomato tissues infected with the bacteria Pseudomonas syringae or related experiments. Our results strongly support the importance of taking advantage of high-throughput transcriptomic data currently available for the selection of proper reference genes in RT-qPCR experiments.
Methods
Bacterial strains and growth conditions
Bacterial strains used were: Pseudomonas fluorescens 5548, Pseudomonas syringae pv. tomato (Pst) DC300049 and Pst DC3000 ΔavrPto ΔavrPtoB50, Pst DC3000 ΔhopQ1-159, Pst DC3000 ΔhrcQ-U and Pst DC3000 Δ28E24. All of them were grown on King’s B medium at 30 °C. Antibiotics used were: ampicillin (100 μg/ml) for Pseudomonas fluorescens 55 and rifampicin (10 μg/ml) for Pst DC3000 and mutants.
Plant material and treatments
For RNA-Seq analysis, 4-week old Rio Grande (RG-PtoR, prf3 and prf19)60 tomato plants were vacuum or syringe infiltrated with bacterial suspensions and MAMPs, sampled at 30 min., 4 h or 6 h, frozen in liquid N2 and stored at −80 °C until processed. Additionally, non-treated tissue was processed in the same way. A detail of the treatments performed in this work along with those from previous works17,18 is shown in Supplementary Table S1. For RT-qPCR analysis, 4-week old resistant Rio Grande-PtoR plants were syringe-infiltrated with a suspension of 108 cfu/ml P. fluorescens 55, 5 × 106 cfu/ml Pst DC3000, 5 × 106 cfu/ml Pst DC3000 ΔavrPto ΔavrPtoB or 10 mM MgCl2 (Table 1). Three biological replicates per infiltration were used and leaf samples were collected at 6 and 12 h after infiltration (hai), frozen in liquid N2 and stored at −80 °C until processed.
RNA-Seq library preparation and analysis
Total RNA was isolated using TRIzol reagent (Life Technologies, NY, USA) and libraries prepared as described previously18. Barcoded libraries were multiplexed by 8–15 in each lane and sequenced on an Illumina HiSeq 2000 equipment with 45–50 bp single-end read mode. Sequence reads generated in this work have been deposited in the NCBI sequence read archive (SRA) under accession number SRP093524. Analysis of the RNA-seq data was performed as described previously18. Processed data generated in this work are available from the Tomato Functional Genomics Database (Tomato Functional Genomics Database [ http://ted.bti.cornell.edu/]).
Selection of the reference genes using RNA-seq data and primer design
Expression data generated in this work, along with those generated in previous RNA-seq experiments17,18 shown in Supplementary Table S1, were used for the selection of the most stably expressed genes across all the treatments. Nine genes with lower variation coefficient (VC), calculated as the ratio between the standard deviation and the average of each gene expression (RPKM, reads per kilobase of transcript per million mapped reads) across all the treatments and biological replicates, were selected (Supplementary Table S2). Additionally, two traditional reference genes used in tomato (GADPH and EF1α) and PHD, the most stably expressed tomato gene identified in a previous report using Xcv infected tomato plants38 were included for analysis.
The nucleotide sequence of each gene was downloaded from the Sol Genomics webpage47 and primers were designed using the PrimerQuest tool (Integrated DNA Technologies). Primer efficiencies were checked by RT-qPCR using different cDNA dilutions (Supplementary Table S3). Dissociation curves were performed to show amplification specificity (Supplementary Fig. S2).
RNA isolation and cDNA preparation
Total RNA was isolated using the Tri-Reagent (Sigma Aldrich) following the manufacturer’s instructions. RNA integrity was assayed by 1% agarose gel electrophoresis. Total RNA (8 μg) was processed with RQ1 RNase-free DNase (Promega) for 60 minutes at 37 °C to eliminate potential DNA contamination and then purified using a chloroform:octanol mix (24:1). RNA concentration and purity was determined using a NanoDrop 2000 spectrophotometer (Thermo Scientific). 2.4 μg RNA was used to prepare cDNA using M-MLV reverse transcriptase (Promega) with random primers according to the manufacturer’s instructions.
RT-qPCR assay
RT-qPCR was performed as described previously61 in 96-well plates (Thermo Fisher Scientific) on the StepOnePlus system (Applied Biosystems). Primer sequences and characteristics are shown in Supplementary Table S3. The reaction mix was performed using: 5 μl of FastStart Universal SYBR Green Master (Rox) (Roche Life Sciences), 2 μl of 2 μM primer mix, 2 μl of a diluted 1:10 cDNA and water to complete a final volume of 10 μl. Cycling conditions were 95 °C for 10 minutes, and 40 cycles of 95 °C for 15 s, 60 °C for 1 min. All RT-qPCR experiments were performed using three biological and three technical replicates.
Evaluation and validation of reference gene expression stability
Data obtained from the RT-qPCR experiments were analyzed using three statistical programs: geNorm51, NormFinder52 and BestKeeper53.
Expression of one PTI- (Solyc02g069960) and one ETI-specific gene (Solyc09g092500) was analyzed by RT-qPCR as explained above18. The data obtained was normalized using the two best and the worst reference genes and the relative expression was expressed as E−ΔΔCq, where E corresponds to the primer efficiency value.
Additional Information
How to cite this article: Pombo, M. A. et al. Use of RNA-seq data to identify and validate RT-qPCR reference genes for studying the tomato-Pseudomonas pathosystem. Sci. Rep. 7, 44905; doi: 10.1038/srep44905 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Bustin, S. Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): trends and problems. J. Mol. Endocrinol. 29, 23–39 (2002).
Derveaux, S., Vandesompele, J. & Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 50, 227–230 (2010).
Bustin, S. A. et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 55, 611–622 (2009).
Kozera, B. & Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 54, 391–406 (2013).
Huggett, J., Dheda, K., Bustin, S. & Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 6, 279–284 (2005).
Gutierrez, L. et al. The lack of a systematic validation of reference genes: a serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotech. J. 6, 609–618 (2008).
Gutierrez, L., Mauriat, M., Pelloux, J., Bellini, C. & Van Wuytswinkel, O. Towards a systematic validation of references in real-time RT-PCR. Plant Cell 20, 1734–1735 (2008).
Guénin, S. et al. Normalization of qRT-PCR data: the necessity of adopting a systematic, experimental conditions-specific, validation of references. J. Exp. Bot. 60, 487–493 (2009).
Boller, T. & He, S. Y. Innate immunity in plants: an arms race between pattern recognition receptors in plants and effectors in microbial pathogens. Science 324, 742–744 (2009).
Schwessinger, B. & Ronald, P. C. Plant innate immunity: perception of conserved microbial signatures. Annu. Rev. Plant Biol. 63, 451–482 (2012).
Couto, D. & Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 16, 537–552 (2016).
Dodds, P. N. & Rathjen, J. P. Plant immunity: towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet. 11, 539–548 (2010).
Jones, J. D. & Dangl, J. L. The plant immune system. Nature 444, 323–329 (2006).
Maekawa, T., Kufer, T. A. & Schulze-Lefert, P. NLR functions in plant and animal immune systems: so far and yet so close. Nature Immunol. 12, 817–826 (2011).
Navarro, L. et al. The transcriptional innate immune response to flg22. Interplay and overlap with Avr gene-dependent defense responses and bacterial pathogenesis. Plant Physiol. 135, 1113–1128 (2004).
Tao, Y. et al. Quantitative nature of Arabidopsis responses during compatible and incompatible interactions with the bacterial pathogen Pseudomonas syringae . Plant Cell 15, 317–330 (2003).
Rosli, H. et al. Transcriptomics-based screen for genes induced by flagellin and repressed by pathogen effectors identifies a cell wall-associated kinase involved in plant immunity. Genome Biol. 14, R139, doi: 10.1186/gb-2013-14-12-r139 (2013).
Pombo, M. A. et al. Transcriptomic analysis reveals tomato genes whose expression is induced specifically during effector-triggered immunity and identifies the Epk1 protein kinase which is required for the host response to three bacterial effector proteins. Genome Biol. 15, 492, doi: 10.1186/s13059-014-0492-1 (2014).
Gomez-Gomez, L. & Boller, T. FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis . Mol. Cell 5, 1003–1011 (2000).
Zeng, W. & He, S. Y. A prominent role of the flagellin receptor FLAGELLIN-SENSING2 in mediating stomatal response to Pseudomonas syringae pv tomato DC3000 in Arabidopsis . Plant Physiol. 153, 1188–1198 (2010).
Clarke, C. R. et al. Allelic variation in two distinct Pseudomonas syringae flagellin epitopes modulates the strength of plant immune responses but not bacterial motility. New Phytol. 200, 847–860 (2013).
Hind, S. R. et al. Tomato receptor FLAGELLIN-SENSING 3 binds flgII-28 and activates the plant immune system. Nature Plants 2, 16128, doi: 10.1038/nplants.2016.128 (2016).
Xin, X.-F. & He, S. Y. Pseudomonas syringae pv. tomato DC3000: a model pathogen for probing disease susceptibility and hormone signaling in plants. Annu. Rev. Phytopathol. 51, 473–498 (2013).
Cunnac, S. et al. Genetic disassembly and combinatorial reassembly identify a minimal functional repertoire of type III effectors in Pseudomonas syringae . Proc. Natl. Acad. Sci. USA 108, 2975–2980 (2011).
Martin, G. In Effectors in Plant–Microbe Interactions 121–154 (Wiley-Blackwell, 2011).
Martin, G. B. et al. Map-based cloning of a protein kinase gene conferring disease resistance in tomato. Science 262, 1432–1436 (1993).
Tang, X. & Martin, G. B. et al. Initiation of plant disease resistance by physical interaction of AvrPto and Pto kinase. Science 274, 2060–2063 (1996).
Pedley, K. F. & Martin, G. B. Molecular basis of Pto-mediated resistance to bacterial speck disease in tomato. Annu. Rev. Phytopathol. 41, 215–243 (2003).
Mathieu, J., Schwizer, S. & Martin, G. B. Pto kinase binds two domains of AvrPtoB and its proximity to the effector E3 ligase determines if it evades degradation and activates plant immunity. PLoS Pathog. 10, doi: http://dx.doi.org/10.1371/journal.ppat.1004227 (2014).
Mysore, K. S. et al. Comprehensive transcript profiling of Pto- and Prf-mediated host defense responses to infection by Pseudomonas syringae pv. tomato . Plant J. 32, 299–315 (2002).
Expósito-Rodríguez, M., Borges, A. A., Borges-Pérez, A. & Pérez, J. A. Selection of internal control genes for quantitative real-time RT-PCR studies during tomato development process. BMC Plant Biol. 8, 1–12, doi: 10.1186/1471-2229-8-131 (2008).
Dekkers, B. J. W. et al. Identification of reference genes for RT–qPCR expression analysis in Arabidopsis and tomato seeds. Plant Cell Physiol. 53, 28–37 (2012).
González-Aguilera, K. L., Saad, C. F., Chávez Montes, R. A., Alves-Ferreira, M. & de Folter, S. Selection of reference genes for quantitative real-time RT-PCR studies in tomato fruit of the genotype MT-Rg1. Front. Plant Sci. 7, doi: 10.3389/fpls.2016.01386 (2016).
Løvdal, T. & Lillo, C. Reference gene selection for quantitative real-time PCR normalization in tomato subjected to nitrogen, cold, and light stress. Anal. Biochem. 387, 238–242 (2009).
Lacerda, A. L. M. et al. Reference gene selection for qPCR analysis in tomato-bipartite Begomovirus interaction and validation in additional tomato-virus pathosystems. PLoS ONE 10, e0136820, doi: 10.1371/journal.pone.0136820 (2015).
Alfenas-Zerbini, P. et al. Genome-wide analysis of differentially expressed genes during the early stages of tomato infection by a Potyvirus. Mol. Plant Microbe Interact. 22, 352–361 (2009).
Mascia, T., Santovito, E., Gallitelli, D. & Cillo, F. Evaluation of reference genes for quantitative reverse-transcription polymerase chain reaction normalization in infected tomato plants. Mol. Plant Pathol. 11, 805–816 (2010).
Muller, O. A. et al. Genome-wide identification and validation of reference genes in infected tomato leaves for quantitative RT-PCR analyses. PLoS ONE 10, e0136499, doi: 10.1371/journal.pone.0136499 (2015).
Wang, Z., Gerstein, M. & Snyder, M. RNA-Seq: a revolutionary tool for transcriptomics. Nature Rev. 10, 57–63 (2009).
Ansorge, W. J. Next generation DNA sequencing techniques. New Biotechnol. 25, 195–203 (2009).
Haas, B. J. & Zody, M. C. Advancing RNA-Seq analysis. Nature Biotechnol. 28, 421–423 (2010).
Wang, L., Li, P. & Brutnell, T. P. Exploring plant transcriptome using ultra high-throughput sequencing. Brief. Funct. Genomics 9, 118–128 (2010).
Rosli, H. G. & Martin, G. B. Functional genomics of tomato for the study of plant immunity. Brief. Funct. Genomics, doi: 10.1093/bfgp/elv007 (2015).
González-Agüero, M. et al. Identification of two putative reference genes from grapevine suitable for gene expression analysis in berry and related tissues derived from RNA-Seq data. BMC Genomics 14, 1–12, doi: 10.1186/1471-2164-14-878 (2013).
Yim, A. K.-Y. et al. Using RNA-seq data to evaluate reference genes suitable for gene expression studies in soybean. PLoS ONE 10, e0136343, doi: 10.1371/journal.pone.0136343 (2015).
Ma, R., Xu, S., Zhao, Y., Xia, B. & Wang, R. Selection and validation of appropriate reference genes for quantitative real-time PCR analysis of gene expression in Lycoris aurea . Front. Plant Sci. 7, 536, doi: 10.3389/fpls.2016.00536 (2016).
Fernandez-Pozo, N. et al. The Sol Genomics Network (SGN)—from genotype to phenotype to breeding. Nucleic Acids Res., doi: 10.1093/nar/gku1195 (2014).
Huang, H. C. et al. Molecular cloning of a Pseudomonas syringae pv. syringae gene cluster that enables Pseudomonas fluorescens to elicit the hypersensitive response in tobacco plants. J. Bacteriol. 170, 4748–4756 (1988).
Cuppels, D. A. Generation and characterization of Tn5 insertion mutations in Pseudomonas syringae pv. tomato . Appl. Environ. Microbiol. 51, 323–327 (1986).
Lin, N. C. & Martin, G. B. An avrPto/avrPtoB mutant of Pseudomonas syringae pv. tomato DC3000 does not elicit Pto-mediated resistance and is less virulent on tomato. Mol. Plant Microbe Interact. 18, 43–51 (2005).
Vandesompele, J. et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 3, 1–12, doi: 10.1186/gb-2002-3-7-research0034 (2002).
Andersen, C. L., Jensen, J. L. & Ørntoft, T. F. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 64, 5245–5250 (2004).
Pfaffl, M. W., Tichopad, A., Prgomet, C. & Neuvians, T. P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper – Excel-based tool using pair-wise correlations. Biotechnol. Lett. 26, 509–515 (2004).
Czechowski, T., Stitt, M., Altmann, T., Udvardi, M. K. & Scheible, W.-R. Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis . Plant Physiol. 139, 5–17 (2005).
Fang, P. et al. Assessment of reference gene stability in Rice stripe virus and Rice black streaked dwarf virus infection rice by quantitative Real-time PCR. Virol. J. 12, 175, doi: 10.1186/s12985-015-0405-2 (2015).
Wang, M., Wang, Q. & Zhang, B. Evaluation and selection of reliable reference genes for gene expression under abiotic stress in cotton (Gossypium hirsutum L.). Gene 530, 44–50 (2013).
Potnis, N. et al. Bacterial spot of tomato and pepper: diverse Xanthomonas species with a wide variety of virulence factors posing a worldwide challenge. Mol. Plant Pathol. 16, 907–920 (2015).
Petriccione, M., Mastrobuoni, F., Zampella, L. & Scortichini, M. Reference gene selection for normalization of RT-qPCR gene expression data from Actinidia deliciosa leaves infected with Pseudomonas syringae pv. actinidiae . Sci. Rep. 5, 16961, doi: 10.1038/srep16961 (2015).
Wei, C. F. et al. A Pseudomonas syringae pv. tomato DC3000 mutant lacking the type III effector HopQ1-1 is able to cause disease in the model plant Nicotiana benthamiana . Plant J. 51, 32–46 (2007).
Salmeron, J. M., Barker, S. J., Carland, F. M., Mehta, A., Y. & Staskawicz, B. J. Tomato mutants altered in bacterial disease resistance provide evidence for a new controlling pathogen recognition. Plant Cell 6, 511–520 (1994).
Nguyen, H. P. et al. Methods to study PAMP-triggered immunity using tomato and Nicotiana benthamiana . Mol. Plant Microbe Interact. 23, 991–999 (2010).
Acknowledgements
We thank Dr. Pedro M. Civello and Dr. Gustavo Martinez for providing some of the resources used for this work. This research was supported by National Science Foundation grants IOS-1025642 and IOS-1546625 (GBM and Alan Collmer, PIs), Agencia Nacional de Promoción Científica y Técnica - Argentina (PICT2014-1589), Consejo Nacional de Investigaciones Científicas y Técnicas - Argentina (PIP2013-0440 and PIP2014-0314).
Author information
Authors and Affiliations
Contributions
M.A.P., G.B.M. and H.G.R. designed the research, M.A.P. and H.G.R. performed the research, M.A.P., Y.Z., Z.F., G.B.M. and H.G.R. analyzed the data, M.A.P., G.B.M. and H.G.R. wrote the paper. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Pombo, M., Zheng, Y., Fei, Z. et al. Use of RNA-seq data to identify and validate RT-qPCR reference genes for studying the tomato-Pseudomonas pathosystem. Sci Rep 7, 44905 (2017). https://doi.org/10.1038/srep44905
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep44905
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
