
Abstract
Global climate change (GCC) can negatively affect freshwater ecosystems. However, the degree to which freshwater populations can acclimate to long-term warming and the underlying molecular mechanisms are not yet fully understood. We used the cooling water discharge (CWD) area of a power plant as a model for long-term warming. Survival and molecular stress responses (expression of molecular chaperones, antioxidants, bioenergetic and protein synthesis biomarkers) to experimental warming (20–41 °C, +1.5 °C per day) were assessed in invasive clams Corbicula fluminea from two pristine populations and a CWD population. CWD clams had considerably higher (by ~8–12 °C) lethal temperature thresholds than clams from the pristine areas. High thermal tolerance of CWD clams was associated with overexpression of heat shock proteins HSP70, HSP90 and HSP60 and activation of protein synthesis at 38 °C. Heat shock response was prioritized over the oxidative stress response resulting in accumulation of oxidative lesions and ubiquitinated proteins during heat stress in CWD clams. Future studies should determine whether the increase in thermal tolerance in CWD clams are due to genetic adaptation and/or phenotypic plasticity. Overall, our findings indicate that C. fluminea has potential to survive and increase its invasive range during warming such as expected during GCC.
Similar content being viewed by others


Introduction
Freshwater ecosystems play a critical role in sustaining life and support a disproportionately high fraction of the Earth’s biodiversity. Because of their extensive use and the landlocked position, the freshwater ecosystems are among the most threatened habitats due to pollution, alterations in land and water use and global climate change1. Global climate change (GCC) is an imminent threat for freshwater ecosystems leading to warming, higher amplitude of thermal fluctuations and increased frequency of extreme weather events such as droughts and heat waves. The consensus scenarios of the Intergovernmental Panel for Climate Change (IPCC) forecast a 1.5–6 °C increase in the mean temperature worldwide (depending on the CO2 emission scenarios), with freshwater bodies warming significantly faster than most terrestrial and marine habitats2. This change can strongly affect freshwater organisms, which are >99% comprised of ectotherms and thus are sensitive to the direct effects of temperature on their physiological and biochemical processes.
Understanding the physiological and molecular mechanisms that limit species’ thermal tolerance in the face of GCC is critical for predicting the potential effects of GCC on aquatic populations3. An important challenge for the development of such physiologically-based mechanistic models is choosing a tractable experimental system that approximates the magnitude and the time scale of warming typical of GCC. Macrophysiological approaches comparing populations from different latitudes provide important insights into the mechanisms involved in the long-term acclimation or adaptation to different thermal regimes; however, such adaptations typically occur on much longer time scale than GCC and may be confounded by different evolutionary history of the populations from different latitudes4,5. On the other end of the spectrum, laboratory studies involving short-term (days to months) acclimations to different thermal regimes may underestimate the long-term effects and/or overestimate the thermal tolerance limits of aquatic organisms facing GCC6. Studies of the model populations sharing a recent common origin and exposed to long-term warming are therefore important to complement and advance our understanding of the potential effects of GCC on aquatic organisms. Cooling water discharge (CWD) areas of electrical power plants can serve as incidental field laboratories to study the effects of long-term warming. The CWD areas are characterized by distinctive thermal regimes with temperatures stably elevated above similar natural reservoirs permitting long-term acclimation of the resident biota7,8. Furthermore, long-term exposure to elevated temperatures in the CWD area occurs in the natural settings that preserve the essential habitat features and species interactions. This makes CWD areas experimental goldmines to study the responses of freshwater organisms to long-term warming in the field.
Freshwater bivalves such as clams and mussels are keystone organisms in freshwater ecosystems playing a key role in the food webs, sediment bioturbation and top-down control of water quality9,10. Due to their sedentary life style and filter-feeding behavior, freshwater bivalves can serve as bioindicators of the water quality and ecosystem health. The Asian clam Corbicula fluminea (Müller, 1774) is a common freshwater bivalve in the United States. It is an invasive species introduced from Asia around 1938 and spread throughout all the major watersheds of both Americas11,12. C. fluminea is a nuisance species that colonizes the pipes of water treatment systems and power stations altering flow and increasing sedimentation rates. Established C. fluminea populations also affect the water quality and sediment mixing through active filtration and burrowing behavior13,14. Due to their broad distribution, important role in freshwater ecosystems and common occurrence in the CWD areas of the power plants, C. fluminea are useful organisms to study the acclimation capacity to long-term warming. In this study, we focused on three populations of C. fluminea from two pristine areas (Lake Norman State Park, or LN, and Catawba River, or CR) and the water discharge channel of the Catawba Nuclear Station (CWD) in North Carolina, USA. Catawba Nuclear Station began commercial operations in 1985, approximately 15 years after C. fluminea colonized Catawba river11 so that the CWD population of C. fluminea was exposed to elevated temperatures near the power station outflow for ~30 years.
To gain insights into the acclimation potential of C. fluminea facing long-term warming, we determined thermal tolerance limits of clams from the three populations (LN, CR and CWD) and assessed their molecular stress responses and bioenergetic capacity during heat stress. We hypothesized that acclimation in the CWD area will result in an upward shift of the thermal tolerance limits of clams, and this shift will be associated with upregulation of stress protection mechanisms at high temperatures. To test these hypotheses, we determined the expression of stress-related genes (including heat shock proteins and antioxidants) and measured levels of oxidative lesions and protein damage during acute temperature rise from 20 to 41 °C (at a rate of 1.5 °C day−1) in C. fluminea from different habitats. We also assessed whether interpopulational differences in heat tolerance are associated with differential ability to maintain energy homeostasis during heat stress. Cellular energy status was assessed by expression of a key energy sensor, AMP-dependent protein kinase α (AMPKα) (Hardie, 2014). Transition to partial anaerobiosis (indicative of insufficient aerobic capacity) was determined by the relative activities of key enzymes at aerobic/anaerobic branchpoint [pyruvate kinase (PK), phosphoenolpyruvate carboxykinase (PEPCK), and lactate dehydrogenase (LDH)], and by stabilization of the hypoxia-inducible factor 1α (HIF-1α)15,16,17,18,19. Effects of acute warming on the protein synthesis was determined by measuring the expression of a key regulator of protein translation, the eukaryotic initiation factor eIF-2α (Larade and Storey, 2002; 2007). This comprehensive assessment of molecular and cellular stress responses provides insights into the mechanisms involved in the heat tolerance of clams exposed to chronically elevated temperatures.
Results
Heat tolerance
Clams from the three studied populations differed in tolerance to the experimental temperature rise (+1.5 °C per day), with CWD clams being the most tolerant and those from the LN site – most susceptible to heat stress (Fig. 1). Thus, >90% of CWD clams survived 48 h of exposure to 38 °C, compared to only 10% of CR clams. All LN clams died at 35 °C. LT10 (temperatures inducing 10% mortality) were 24, 28 and 36 °C in LN, CR and CWD clams, respectively.

Mortality of C. fluminea from the three studied sites during acute temperature rise.
(A) – mortality, (B) – geographic position of the studied C. fluminea populations in the Catawba River basin, map data: © 2016 Google. Populations: LN - Lake Norman site, CR – Catawba River site, CWD – cooling water discharge site. Temperature was raised at a rate of 1 °C per hour for 3 hours, followed by a 48 h acclimation period after every 3 °C rise. LT10 and LT50 are the temperatures causing 10% and 50% mortality in the studied populations, respectively. LT50 could not be determined in the CWD clams due to the low mortality rates at the temperatures ≤38 °C and 100% mortality at 41 °C. Vertical bars represent the standard error of estimate. N = 122 for LN and CR, and 119 for CWD.
Molecular chaperones
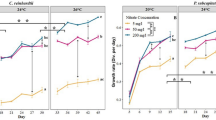
Levels of mRNA encoding molecular chaperones were significantly affected by the site, the experimental temperature and their interaction, except for HSC70 where only the site effect was statistically significant (Table 1). Elevated temperatures (32 °C) led to a significant upregulation of mRNA for the inducible HSP70 (by ~4–5 fold) and HSP90 (by ~2.5–4 fold) in all three studied populations (Fig. 2). Notably, mRNA expression of HSP70 and HSP90 increased slightly at 38 °C in CR clams, but rose dramatically (by ~25 and ~10 fold for HSP70 and HSP90, respectively) in CWD clams (Fig. 2). No data for 38 °C are available for LN clams due to 100% mortality. Expression of the constitutive HSP70 isoform (HSC70) was unaffected by the temperature in clams from the three studied populations (Fig. 2).

Effect of acute warming on expression of mRNA encoding molecular chaperones (heat shock proteins) in C. fluminea from the three studied populations.
X- axis – exposure temperatures, Y- axis – mRNA levels of the target gene relative to the geometric mean of two reference genes (β-actin and RSP9). Different letters indicate significant differences between the means for different populations (p < 0.05). Asterisks indicate values that are significantly different from the respective control group (exposed at 20 °C) within each population (p < 0.05). Arrows on the X-axis indicate LT10 for each of the studied populations. Vertical bars represent the standard error of means. N = 8–9.
Mitochondrial HSP60 was strongly upregulated at 32 °C in the CR population (by ~11 fold) and at 38 °C in the CWD population (by ~30 fold), but did not change with exposure temperature in the LN population (Fig. 2). Expression of HSP60 mRNA expression decreased back to the baseline levels at the extreme temperature (38 °C) in CR clams.
Expression of mRNA for small HSPs (HSP40 and HSP22) did not change in response to acute temperature rise in LN clams (Fig. 2). In CR clams, expression of HSP22 mRNA declined at elevated temperatures, whereas HSP40 expression transiently declined at 32 °C and then increased at 38 °C. In CWD clams, HSP22 expression was not affected by the experimental temperature, while HSP40 levels decreased with warming (Fig. 2).
Antioxidants and glutathione system
mRNA expression of all studied antioxidants (except GSTS1) was significantly affected by the temperature and/or site × temperature interactions (Table 1). Temperature-induced changes in the mRNA expression of antioxidants differed between the clams from the three study sites. In the LN population, exposure to 26 °C led to a significant upregulation of the expression of Mn-SOD, GSTμ and TXNRD1; this increase was transient and mRNA expression of these genes returned to the background levels at higher temperatures (32 and 38 °C) (Fig. 3). mRNA expression of CAT, GPxA, GSTS1, TPX and TXNIP did not change in response to acute temperature rise in LN clams, while expression of Cu, Zn-SOD and GSTM1 declined at 32 °C. The CR population showed the strongest response of the antioxidant expression to experimental heating. In this group, many antioxidants (Mn-SOD, Cu,Zn-SOD, CAT, GPxA, GSTμ and TXRND1) showed a similar non-linear expression pattern with a decrease in the mid-range of the experimental temperatures (26 and 32 °C) followed by the return to the baseline levels at 38 °C (Fig. 3). In the most heat-tolerant CWD population, expression levels of antioxidants generally remained stable during the acute temperature rise except for GPxA, MTs and TPX1 that were upregulated and CAT that was downregulated at the extreme temperature (38 °C).

Effect of acute warming on expression of mRNA encoding antioxidants in C. fluminea from the three studied populations.
X- axis – exposure temperatures, Y- axis – mRNA levels of the target gene relative to the geometric mean of two reference genes (β-actin and RSP9). Different letters indicate significant differences between the means for different populations (p < 0.05). Asterisks indicate values that are significantly different from the respective control group (exposed at 20 °C) within each population. Arrows on the X-axis indicate LT10 for each of the studied populations. Vertical bars represent the standard error of means. N = 8–9.
Oxidative lesions
The levels of MDA and protein carbonyls were significantly affected by the site, but not experimental temperature, while the levels of 4-HNE were affected by the site, experimental temperature and their interaction (Table 1). Under the control conditions, the levels of MDA and carbonyls were similar in clams from all three studied populations, whereas the levels of 4-NHE were lower in the CR group (Fig. 4). Acute warming led to an increase of MDA levels in the CWD population and an increase of 4-HNE levels in the CR population from 26 °C onwards. Levels of 4-HNE were also slightly elevated at 26 and 32 °C in the LN populations, but this trend was only significant at 26 °C (Fig. 4). Elevated temperature had no effect on the protein carbonyl levels in any of the three studied populations (Fig. 4).

Effect of acute warming on expression of mRNA encoding components of thioredoxin system (A,C,E) and oxidative lesions (B,D,F) in C. fluminea from the three studied populations. X- axis – exposure temperatures, Y axis – mRNA levels of the target gene relative to the geometric mean of two reference genes (β-actin and RSP9) (A,C,E) or tissue levels of oxidative lesions (B,D,F). Different letters indicate significant differences between the means for different populations (p < 0.05). Asterisks indicate values that are significantly different from the respective control group (exposed at 20 °C) within each population (p < 0.05). Arrows on the X-axis indicate LT10 for each of the studied populations. Vertical bars represent the standard error of means. N = 8–9 for mRNA expression and 5 for oxidative lesions (MDA, 4-HNE, and carbonyls).
Protein ubiquitination
The baseline levels of ubiquitinated proteins were lower in CWD clams compared to those from the LN and CR sites (Fig. 5). The levels of protein ubiquitination increased with the increasing temperature in CWD clams, did not change in CR clams and decreased with temperature in LN clams (Fig. 5).

Effect of acute warming on protein expression of bioenergetic and protein synthesis biomarkers in C. fluminea from the three studied populations.
X- axis – exposure temperatures, Y - axis – target protein levels in relative units (RU). Different letters indicate significant differences between the means for different populations (p < 0.05). Asterisks indicate values that are significantly different from the respective control group (exposed at 20 °C) within each population (p < 0.05). Arrows on the X-axis indicate LT10 for each of the studied populations. Vertical bars represent the standard error of means. N = 3–4 for all proteins except ubiquitin (N = 5) and HIF-1α from the heat exposed CR groups (26, 32 and 38 °C) where N = 2.
Protein translation
Protein expression levels of the total and phosphorylated elongation factor 2α (eIF-2α and p-eIF-2α, respectively) were affected by the site, experimental temperature and their interaction (Table 1). Under the control conditions, protein levels of eIF-2α and p-eIF-2α were higher in the mollusks from the CWD group. Total levels of eIF-2α increased with increasing temperatures in CR and CWD but not LN clams (Fig. 5). Elevated temperature led to a strong decrease (by 4–7 fold) of the phosphorylated p-eIF-2α in the CWD clams and an increase of p-eIF-2α levels in the LN and CR groups (Fig. 5).
Bioenergetic markers
Basal AMPKα expression levels were highest in the CR population and lowest in CWD clams. In all three populations, AMPK levels increased with increasing temperatures (Fig. 5). Expression of HIF-1α was induced by elevated temperature in the LN and CWD populations (at 26 and 32–38 °C, respectively) (Fig. 5). No temperature-induced change in the HIF-1α protein levels was found in CR clams.
Enzymatic activities of PK and LDH were similar in the control clams from the three studied populations, while the PEPCK activity was lower in LN clams compared to those from the CR and CWD sites (Fig. 6). As a result, PK/PEPCK activity (an index of the relative aerobic capacity) was considerably higher in the LN population compared to CR and CWD. Acute warming had little effect on the activities of PK, PEPCK and LDH (Fig. 6). In LN clams, acute warming led to a significant decrease in PK/PEPCK ratio indicating a decline in aerobic capacity and activation of anaerobic pathways (Fig. 6). In CR and CWD clams, PK/PEPCK ratio remained at the steady-state levels throughout the experimental exposures (Fig. 6).

Effect of acute warming on enzyme activities in C. fluminea from the three studied populations.
(A,B) – population LN, (C,D) – population CR, and (E,F) – population CWD. A, C, D – activities of PK, PEPCK and LDH; B, D, F – PK/PEPCK ratio. X- axis – exposure temperatures, Y - axis – specific activities of the respective enzymes (U g−1 protein). Asterisks indicate values that are significantly different from the respective control group (exposed at 20 °C) within each population (p < 0.05). Arrows on the X-axis indicate LT10 for each of the studied populations. An inset table above the graphs indicates comparisons of the enzyme activities between the control groups from different populations; if the rows do not share a letter, the respective enzyme activities are significantly different (p < 0.05). Vertical bars represent the standard error of means. N = 8.
Data integration
Expression levels of different antioxidants (including Mn-SOD, Cu, ZN-SOD, GPxA, TPX, TXNRD1, CAT, GSTM1 and GSTμ) significantly and positively correlated with each other in C. fluminea across the studied populations and experimental groups (Supplementary Table 1). Tissue levels of oxidative lesions (MDA, 4-HNE and protein carbonyls) positively correlated with each other and negatively correlated with mRNA expression of select antioxidants (CAT in the case of MDA, MTs and GSTM1 in the case of 4-HNE, and GSTM1 and GSTS1 in the case of carbonyls). Notably, expression of HIF-1α positively correlated with the expression of molecular chaperones (HSP70, HSP90, and HSP22), antioxidants (MT, Mn-SOD, GSTM1, GSTμ, GPxA, TPX, and TXNRD1), oxidative lesions (MDA, HNE and carbonyls) and activity of LDH. Expression of total eiF-2α was positively correlated with the expression of its phosphorylated form (p-eiF-2α), as well as AMPK, MTs and molecular chaperones (HSP60, HSP70, HSP90 and HSP22) and negatively correlated with the expression of antioxidants (Mn-SOD, GSTM1, GSTμ, and CAT) (Supplementary Table 1). Expression of p-eiF-2α also negatively correlated with oxidative stress parameters (GSTμ, MDA and carbonyls) and ubiquitin levels (Supplementary Table 1). AMPK expression positively correlated with the activity of PEPCK, and negatively - with the PK activity, expression of antioxidants (GSTμ, TPX1) and oxidative lesions (MDA, HNE and carbonyls) (Supplementary Table 1).
Discriminant analysis showed a clear separation among the three studied populations based on their site of origin (F48,130 = 10.59, p < 0.0000) and the experimental temperature (F72,192 = 10.45, p < 0.0000). The parameters that most significantly contributed to the discriminant function separating different populations included Mn-SOD, HSP22, MDA, 4-HNE, eiF-2α, and AMPK (Fig. 7A). CART analysis (used to identify a single most distinguishing characteristic between the groups) showed that clams exposed to 32 or 38 °C were distinguished from other experimental temperatures by high levels of HSP70, while clams exposed to 38 °C were differentiated by higher HSP22 expression levels from those exposed to 32 °C (Fig. 7B). Clams exposed to 26 °C were distinguished from the control group (20 °C) based on the expression of AMPK and TXNIP.

Separation of the studied populations and/or experimental treatment groups based on the discriminant (A) and CART (B) analyses. A - Population groupings based on the discriminant analysis. Biomarkers that significantly contributed to the discriminant function included Mn-SOD, HSP22, MDA, 4-HNE, AMPK and eiF-2α (p < 0.05). B – Classification and regression Tree (CART) analysis. The main distinguishing characteristic separating each of the temperature groups (across all populations) is shown at the respective nodes of the tree.
Discussion
Long-term acclimation of C. fluminea to a chronically warmed CWD area was associated with a dramatic increase in heat tolerance compared to the clams from the habitats that do not experience thermal pollution. In the CWD population, ~90% clams could withstand 48 h of exposure at 38 °C, the conditions which where lethal to >90% of the CR clams and 100% of the LN clams. Furthermore, LT10 (i.e. the threshold temperature at which 10% of the population died) was 12 °C higher in the CWD clams than in the LN clams, and 8 °C higher compared to the CR clams. While we cannot rule out the possibility that other ecological variables (such as food availability in the field) may have contributed to the variation in thermal tolerance, a strong correlation between the thermal tolerance limits of clams and daytime summer temperatures in their habitats make the thermal regime the most plausible explanation for the observed differences in the thermal thresholds. This plasticity of the upper thermal limits indicates a strong potential of C. fluminea to withstand long-term warming such as expected during GCC. It is worth noting that the upper thermal limit of the most tolerant CWD population is close to the maximum thermal tolerance limits recorded in ectotherms and thus may have a limited capacity to evolve to reach even higher levels20.
A common pattern of heat stress response in all studied populations included upregulation of mRNA for large mitochondrial and cytosolic heat shock proteins (HSP70 and HSP90). Induction of HSP70 and HSP90 is a common response to heat stress in many organisms including mollusks21,22,23,24,25, reflecting the important role of these chaperones in refolding the partially denatured proteins and targeting the irreparably damaged ones for degradation26,27. Temperature-dependent chaperone expression revealed significant interpopulational differences linked to the differences in the heat tolerance among the three studied populations. While the magnitude of HSP70 and HSP90 induction at intermediate temperatures (32 °C) were similar in the three studied populations, only the heat tolerant CWD clams were capable of a strong upregulation (by 10–25 fold) of HSP70 and HSP90 at the extreme temperature (38 °C). Given the protective role of HSP70 and HSP90 against the heat-induced protein damage26,28, high levels of HSP70 and HSP90 expression in the CWD clams may contribute to their ability to survive extremely high temperatures. In marine gastropods from the genus Tegula, acclimation to elevated temperatures led to an upward shift of the temperature, at which the highest HSP expression was observed22. Interestingly, the temperature at which the synthesis of HSPs was inactivated strongly correlated with the species-specific thermal tolerance22. In sea urchins (Strongylocentrotus purpuratus) and fish (Gillichthys mirabilis) long-term acclimation to elevated temperatures resulted in better survival and enhanced transcription of HSP70 at extreme temperatures (32–36 °C) compared to their cold-acclimated counterparts that failed to induce HSP70 mRNA under these conditions29,30. A strong link between the ability to synthesize HSP70 at extreme temperatures and thermotolerance was also found in ascidians, where the more thermotolerant invasive species Diplosoma listerianum expressed high levels of HSP70 at extreme temperatures while the less thermotolerant, native Distaplia occidentalis failed to maintain HSP70 levels at the temperatures above LT5031. In two species of freshwater unionid bivalves, a more thermotolerant species, Villosa lienosa, expressed higher levels of HSP70 and HSP90 transcripts during heat stress compared to the thermally sensitive Villosa nebulosa32. These data indicate that an ability to maintain synthesis of HSP70 and HSP90 at extreme temperatures plays an important role in setting the upper limit of thermal tolerance in aquatic invertebrates, including mollusks.
High heat tolerance of CWD clams was also associated with the ability to maintain high levels of the protein synthesis during acute heat rise as indicated by an increase in the levels of the eukaryotic transcription factor eiF2α and a decrease of its phosphorylated form. In mollusks like in other eukaryotes, eiF2α is an essential translation initiation factor required for protein translation which mediates the binding of tRNA for methionine to the ribosome33,34. eiF2α can be inactivated by reversible phosphorylation which results in suppression of the protein synthesis33,34. The temperature-induced increase in the amount of active, non-phosphorylated eiF2α in CWD clams indicates high translational activity at elevated temperatures. This will support the de novo synthesis of heat shock proteins (such as HSP70, HSP90 and HSP60), which are essential for survival under the conditions of heat stress26,27. In contrast, in the two less heat-tolerant populations (LN and CR) the levels of phosphorylated eiF2α increased with increasing temperatures indicating suppression of the protein translation. Given that HSPs are the group of the proteins predominantly synthesized during heat stress in mollusks21,22,35, suppression of the protein synthesis rates at elevated temperatures may impair heat shock response leading to protein damage and mortality in heat-sensitive clam populations.
Notably, the temperature-dependent mRNA expression of HSP40, a co-chaperone of HSP70, did not correlate with the HSP70 expression profile in C. fluminea. HSP40 regulates HSP70 function and acts in concert with HSP70 to prevent protein aggregation and catalyze re-folding of misfolded proteins36,37. Given the tight functional linkages between HSP40 and HSP7036,37, one might expect coordinated changes in mRNA expression of these two chaperones during acute temperature rise. However, mRNA levels of HSP40 remained stable during the acute temperature rise in LN clams and decreased at elevated temperatures in CR and CWD clams. This indicates that transcriptional regulation of HSP70 and HSP40 is decoupled, and that unlike HSP70, mRNA expression of HSP40 is not a good indicator of heat stress in C. fluminea.
Transcript levels of another small molecular chaperone, HSP22, were temperature-insensitive in LN and CWD clams and suppressed during the temperature rise in CR clams. In marine intertidal mussels Mytilus californianus expression of a small HSP (HSP30) decreased at elevated temperatures38 similar to the pattern seen in the CR population. To date, the functional roles and temperature-dependent regulation of small heat shock proteins (sHSPs) such as HSP22 and HSP30 are not well understood in mollusks. In mammals, sHSPs are involved in regulation of apoptosis, antioxidant defense and copper homeostasis39,40. In a model invertebrate (Drosophila melanogaster) HSP22 localizes to the mitochondria and has been proposed to play a role in protection of the mitochondria from oxidative stress41. Interestingly, heat-induced suppression of HSP22 in CR clams goes hand-in-hand with a decrease in the transcript levels of mitochondrial and cytosolic antioxidants such as Mn-SOD, Cu, Zn-SOD, CAT, GPX and GSTμ. This raises an intriguing possibility that HSP22 may have an antioxidant function in mollusks similar to that in Drosophila and mammals40,41, which requires further investigation. Regardless of the cellular functions of HSP22, mRNA levels of this gene do not show a consistent pattern of change during temperature rise in different clam populations and thus cannot serve as a biomarker for heat stress in C. fluminea.
The highly and intermediately tolerant populations (CR and CWD) strongly overexpressed the mitochondrial chaperone HSP60 at 32 and 38 °C, respectively, whereas no temperature-dependent induction of HSP60 mRNA was observed in heat-sensitive LN clams. Mitochondrial HSP60 is involved in maintaining the mitochondrial function and DNA stability42 and can protect mitochondria against temperature stress42,43,44 as well as other proteotoxic insults such as exposure to toxic metals23,45,46. Inability of LN clams to overexpress HSP60 during acute heat stress may contribute to the damage of their mitochondria and lead to a decrease in the aerobic capacity at elevated temperatures. This hypothesis is supported by a decrease in the PK/PEPCK ratio in LN clams at elevated temperatures (≥29 °C). In molluscs, PK and PEPCK act as a metabolic switch channeling glycolytic substrates to aerobic (PK) or anaerobic (PEPCK) ATP production in mitochondria16,47. Thus, a decrease in PK/PEPCK ratio reflects a decrease in the aerobic capacity for ATP production and activation of anaerobic pathways16,18,47. In contrast to LN clams, CR and CWD clams maintained the steady-state PK/PEPCK ratio at all studied temperatures. This suggests that the mitochondrial metabolism of CR and CWD clams is robust to the acute temperature rise, possibly due to the protective overexpression of the mitochondrial chaperone HSP60 at high temperatures.
Tissue levels of AMPKα increased with increasing temperatures in clams from all three populations of C. fluminea, albeit the trend was only significant in CR and CWD clams. This may indicate cellular energy deficiency at elevated temperatures, since AMPKα is the main cellular metabolic sensor activated in response to the energy deficit48. Earlier studies in marine mollusks and crabs also showed upregulation of AMPK during heat stress resulting in activation of ATP-generating pathways to support energy homeostasis49,50. This hypothesis is also consistent with accumulation of HIF-1α protein in clams at elevated temperatures indicative of tissue hypoxemia. HIF-1α is a constitutively expressed transcription factor which is rapidly degraded in the presence of oxygen but accumulates during hypoxia and/or tissue hypoxemia15,51 triggering an adaptive transcriptional program that enhances oxygen delivery to the tissue, increases glucose uptake, and stimulates glycolysis51,52. Upregulation of AMPKα and stabilization of HIF-1α in C. fluminea tissues indicates heat-induced impairment of energy status and potential hypoxemia. However, this cellular energy stress was not sufficient to induce transition to partial anaerobiosis as shown by the stability of PK/PEPCK ratio at the temperatures that trigger accumulation of AMPKα and HIF-1α in the studied clam populations. Oxidative stress response was activated at the intermediate temperatures (26 °C) in the heat-sensitive LN clams as indicated by the elevated mRNA expression of Mn-SOD, GSTμ and TXNRD1. In the intermediately and highly heat-tolerant CR and CWD clams mRNA expression for antioxidants remained at the background levels or declined at elevated temperatures (with the exception of TPX1 and GPxA mRNA upregulated in the CWD clams at 38 °C). This indicates that transcriptional upregulation of antioxidants does not play a key role in the mechanism of heat tolerance in C. fluminea. Lack of induction and/or suppression of mRNA levels of antioxidants at the elevated temperatures (such that induce molecular chaperones) may indicate antagonistic interactions between the transcriptional activation of antioxidant response and heat shock pathways in C. fluminea. Similar antagonism between the heat shock (HSR) and oxidative stress (OxSR) response has been shown in a model organism C. elegans53. Prioritization of heat shock over antioxidant response may come at a cost of oxidative damage to the tissues, as indicated by accumulation of oxidative lesions (MDA and 4-HNE) and ubiquitinated proteins in the CR and CWD clams at high temperatures (32–38 °C).
Despite the overall positive correlations between the expression levels of different antioxidants, no single antioxidant-encoding gene showed a consistent pattern of change in response to acute warming across all studied populations. This indicates that while acute heat stress affects redox status of C. fluminea, the compensatory response may involve different antioxidants. Oxidative stress is a common response to elevated temperature in aquatic ectotherms including bivalves54,55,56,57,58. However, similar to our present study, earlier studies of temperature-induced OxSR showed that specific players (i.e. genes and proteins involved in the OxSR) and the direction of change (i.e. upregulation or suppression) vary among species, habitats and exposure conditions59,60. This may reflect the molecular complexity and redundancy of the antioxidant protection systems in the cell, as well as the dual role of ROS as important signaling molecules, on one hand, and potentially damaging agents, on the other61. Our findings emphasize that detection of the oxidative stress signature during acute warming requires assessment of a broad range of antioxidants and redox status parameters in aquatic organisms (such as conducted in the present study).
As a corollary, exposure to the chronic, long-term warming in the CWD area strongly affects thermal tolerance of freshwater clams C. fluminea making them capable of surviving extremely high temperatures (up to 38 °C). High heat tolerance in clams from the CWD area is associated with their ability to maintain high aerobic capacity and active protein synthesis at elevated temperatures, and strong induction of cytosolic and mitochondrial molecular chaperones (including HSP70, HSP90 and HSP60) during heat stress. Notably, there appears to be a trade-off between the heat shock response and oxidative stress response in C. fluminea. At extreme temperatures, heat shock response is prioritized over the antioxidant protection, so that oxidative lesions accumulate in clam tissues. An increase in thermal tolerance and the associated molecular traits in the CWD clams (reflecting phenotypic plasticity, genetic adaptation or both) indicate that C. fluminea has a potential to survive global climate change in much of its current distribution range. Future studies are needed to determine whether such physiological flexibility may also increase the invasive spread of C. fluminea and help them outcompete less heat-tolerant indigenous freshwater bivalves as the mean temperatures and thermal fluctiuations (and thus frequencies of the thermal extremes) increase in freshwater habitats.
Materials and Methods
Animal collection and maintenance
Adult C. fluminea (shell length 26.2 ± 0.3 mm, N = 210) were collected in August 2015 from three habitats in North Carolina: 1) Lake Norman State Park (LN; 35 40′18.01 N, 80 55′57.02 W), 2) Catawba River (CR; 35 16′20.65 N, 81 00′41.25 W), and 3) the water discharge channel of the Catawba Nuclear Station (CWD; 35 02′42.42 N, 81 04′10.12 W). All three collection sites are situated in the Catawba River basin and characterized by sandy-clay bottom sediments, lack of underwater macrophyte vegetation and shallow depth (<2 m). The study sites are ~70 km apart with LN being the most upstream, CR – intermediate, and CWD – the most downstream location in the Catawba River basin (Fig. 1B). While the distance between the study locations makes exchange of the larvae unlikely, we cannot exclude the potential larval transport from the upstream to downstream sites. Therefore, the differences in thermal tolerance and molecular responses to the heat stress between clams from the three study sites may reflect genetic adaptation, phenotypic plasticity or both. Physico-chemical parameters of the water (pH, phosphates, nitrites, ammonium, sulfate, chloride, oxidisability, calcium hardness, and iron levels) collected at the three study sites were determined using Multitest Ammonia and Multitest Nitrite and Nitrate (Seachem Laboratories Inc., Madison, GA) for ammonia, nitrate and nitrite or the standard tests recommended by the American Public Health Association (APHA) for all other parameters62. All measured parameters were below the threshold levels established for the protection of freshwater aquatic life (EC Council Directive 98/83/EC; http://www.emwis.org/IFP/law_EU.htm) indicating that all study sites had good water quality and negligible levels of contamination (data not shown). Temperature was measured every other week in summer 2015 (July-September) and in spring 2016 (March-April) near the clam collection sites. In summer, water temperatures at the CWD site were ~8 °C higher than at the LN site and ~3 °C higher than at the CR site (29.7 ± 0.7 °C, N = 3; 33.8 ± 0.4 °C, N = 6; 37.5 ± 0.4 °C, N = 6 for LN, CR and CWD sites, respectively). In spring, water temperatures were similar at the CWD and CR sites (18.2 ± 0.8 °C, N = 4 at each site) and ~3 °C higher than at the LN site (15.8 ± 1.3 °C, N = 4).
Clams were acclimated for 7–10 days in the laboratory in recirculating aquaria (at the density of 50 clams per 30 l) containing dechlorinated, conditioned tap water at 20 °C, with acid-washed sand as a substrate. The conditioned tap water was used to ensure uniform high water quality in experimental treatments. Three replicate aquaria were used for each population. Clams were fed ad libitum using 2 ml of commercial algal blend (DT’s Premium Plankton Blend, DT’s Plankton Farm, Sycamore, IL) per tank every other day. After pre-acclimation, clams were exposed to an acute temperature increase from 20 °C to 41 °C, with the warming rate of 1 °C per hour followed by a 48 h acclimation period after every 3 °C rise. This warming reduces the acute metabolic effects of rapid temperature change but does not allow for full thermal acclimation63,64. Water pH, nitrite, nitrate and ammonia concentrations were measured daily. Ammonia and nitrite levels were <0.01 mg l−1, nitrate levels <2 mg l−1 (Multitest Ammonia and Multitest Nitrite and Nitrate, Seachem Laboratories Inc., Madison, GA), and pH was 7.13 ± 0.02 (N = 14) throughout the exposures. Mortality was recorded, and tissue samples were collected after 48 h exposure at each experimental temperature to assess stress biomarkers and activities of key metabolic enzymes. Due to the limited amount of tissues, different biomarkers were measured in different tissues of clams (Table 2). Oxidative stress biomarkers and molecular chaperones were assessed in the gills, which is the main organ of the gas exchange and thus most subjected to oxidative stress. Markers related to protein turnover and oxidative protein damage (carbonyl and MDA- and HNE-protein conjugates) were measured in the foot muscle. Enzyme activities were measured in the digestive gland, a key organ involved in energy storage and metabolism of bivalves.
Immunoblotting
Foot muscle tissues were homogenized in ice-cold homogenization buffer (100 mM Tris, pH 7.4, 100 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Triton-X 100, 10% glycerol, 0.1% sodium dodecylsulfate, 0.5% deoxycholate, 0.5 μg mL−1 leupeptin, 0.7 μg ml−1 pepstatin, 40 μg ml−1 phenylmethylsulfonyl fluoride and 0.5 μg ml−1 aprotinin), sonicated three times for 10 s each (output 69 W, Sonicator 3000, Misonix, Farmingdale, NY, USA), with cooling on ice between sonications, and centrifuged for 10 min at 14,000 × g and 4 °C. Protein content was measured in the supernatant using Bio-Rad Protein Assay kit (Bio-Rad Laboratories, Hercules, CA, USA), and protein expression was determined using standard immunoblotting techniques. Ubiquitinated proteins were analyzed by dot-blot technique using 96-well Convertible® dot/slot blotter. Protein samples (20 μg per well) were loaded onto pre-wet nitrocellulose membrane under vacuum, and probed with a polyclonal rabbit antibody against ubiquitin (#Z0458, Dako Denmark A/S, Grostrup, Denmark). All other proteins, including total and phosphorylated eukaryotic translation initiation factor (eiF-2α and p-eiF-2α, respectively), hypoxia-inducible factor 1α (HIF-1α), and 5′-AMP-activated protein kinase α (AMPKα) were resolved using SDS-PAGE electrophoresis. Protein samples (50 μg per lane for eiF-2α, p-eiF-2α, or 80 μg for AMPK and HIF-1α) were loaded onto 10% polyacrylamide gels and run at 100 mA for 2–2.5 h at room temperature. The resolved proteins were transferred onto a nitrocellulose (for AMPK and HIF-1α) or PVDF (for eiF-2α and p-eiF-2α) membrane in 96 mM glycine, 12 mM Tris and 20% methanol (v/v) using a Trans-Blot semi-dry cell (Thermo Fisher Scientific, Portsmouth, NH, USA). Equal loading was verified with Amido Black staining (McDonough et al. 2015). The membranes were blocked in 5% non-fat milk in Tris-buffered saline, pH 7.6, and probed overnight with primary monoclonal antibodies against p-EIF-2α (Ser51) (#07–760, Millipore, Temecula, CA, USA), EIF-2α (#AHO1182, Invitrogen, Frederick, MO, USA), AMPKα (#2793 S, Cell Signaling Technology, Danvers, MA, USA), and HIF-1α (#QC49321–42215, Avivasysbio, USA). After washing off the primary antibody, membranes were probed with the respective polyclonal secondary antibodies conjugated with horseradish peroxidase (Jackson Immunoresearch, West Grove, PA, USA) and proteins detected by enhanced chemiluminescence according to the manufacturer’s instructions (Pierce, Rockford, IL, USA). Densitometric analysis of the signal on SDS-PAGE membranes and dot-blots was performed by GelDoc 2000™ System with Quantity One 1D Analysis Software (Bio-Rad Laboratories Inc., Hercules, CA, USA). To ensure that the heat stress-induced differences in the protein levels are not confounded with blot-to-blot variations, each blot included the same control sample as an internal standard. All antibodies produced bands of the expected molecular size as specified by the respective manufacturers (Supplementary Figure S1). All antibodies except HIF-1α produced a single band. Western blots for HIF-1α included two bands: an upper band (~110 kDa) corresponding to HIF-1α and a lower diffuse band of ~70 kDa corresponding to HIF-1α degradation products. Only the upper band corresponding to the intact transcription factor HIF-1α was quantified.
Enzyme activities
Activities of PK (EC 2.7.1.40), PEPCK (EC 4.1.1.31) and LDH (EC 1.1.1.27) were measured at 20 °C in hepatopancreas of mollusks using standard NADH-coupled spectrophotometric assays as described elsewhere8,18. A molar extinction coefficient of 6.22·106 M-1·cm−1 for NADH was used. In mollusks, PK channels glycolytic substrates to aerobic catabolism47,65. PEPCK and LDH were chosen as they are involved in anaerobic production of succinate and lactate, two major anaerobic end-products in freshwater mollusks65. For all enzyme activities, assay conditions were selected to ensure non-limiting concentrations of the substrates and linearity of the reactions over the assay time. The initial reaction velocity was used to estimate Vmax. Enzyme activities (Vmax) were expressed as U g−1 protein.
Quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from the gills tissue of mollusks with TRIzol® reagent (Invitrogen, Carlsbad, USA). Specific primers for target genes were designed against partial mRNA sequences published in the National Center for Biotechnology Information (NCBI) database (Table 1) using PrimerQuest software (Integrated DNA Technologies, Inc., Coralville, IA, USA). RNA amount and quality were assessed using Nanodrop spectrophotometer. RNA concentration in all samples was >200 ng/μL with A260/280 and A260 ⁄230 values of 2.0 ± 0.1 and 2.2 ± 0.1, respectively (N = 130). All RNA samples were subjected to a PCR reaction with gene-specific primers (Table 1) to test for DNA contamination; no amplification was observed after 40 cycles of PCR indicating that RNA samples contained no DNA contamination that could interfere with qRT-PCR. cDNA was then synthesized from RNA samples as described elsewhere66. Transcript levels of target genes as well as ß-actin and 40 S ribosomal protein S9 (RSP9) used as the reference genes were quantified by qRT-PCR using a 7500 Fast Real-Time PCR System (Life Technologies, Carlsbad, CA, USA) and SYBR® Green PCR kit (Life Technologies, Bedford, MA, USA). Expression levels of ß-actin and RSP9 did not significantly differ between the experimental groups (p > 0.05). The normalized expression of target genes was calculated as follows:

Relative mRNA quantities of the target and reference genes were calculated according to Pffafl (2001) using gene-specific amplification efficiencies. Amplification efficiency was determined in each run by amplifying serial dilutions of the internal cDNA standard (Pfaffl, 2001). The qRT-PCR conditions were as described in previous studies58,66; the annealing and read temperatures were 55 °C and 72 °C, respectively, for all primer pairs except HSC70. For HSC70 the annealing and read temperatures were 50 °C and 72 °C, respectively. A single cDNA sample was included in each run to test and correct for the potential run-to-run amplification variability58,67.
Oxidative lesions
Protein conjugates of malondialdehyde (MDA) and 4-hydroxynonenal (4-HNE) were measured in gill tissue homogenates (N = 5) using enzyme immunoassays (MDA OxiSelect™ MDA adduct ELISA Kit and HNE OxiSelect™ HNE-His adduct ELISA Kit, respectively) according to the manufacturer’s protocols (Cell Biolabs, Inc., CA, USA). Protein carbonyl (PC) content was measured in gill tissue homogenates through the reaction with 2,4-Dinitrophenylhydrazine (DNPH)68. The differences in absorbance between the DNPH- and the HCl-treated samples were determined by spectrophotometry at 360 nm, and the amount of carbonyl was determined by using a molar extinction coefficient of 2.2·104 M−1·cm−1. Data were expressed as nmol carbonyl·mg−1 of soluble extracted protein.
Statistical analysis
Data were tested for normality and homogeneity of variance by Kolmogorov-Smirnoff and Levene’s tests, respectively. As needed, data were normalized by Box-Cox common transforming method (Table 1). Fisher’s Least Significant Differences (LSD) tests were used for planned post-hoc comparisons of the differences between the pairs of means of interest. Logit analysis was used to calculate LT10 and LT50, the temperatures causing 10% and 50% mortality, respectively. Pearson’s correlation was used to assess correlations between the studied traits within each individual across all experimental groups. Normalized Box-Cox transformed data were subjected to the discriminant analysis to reduce the dimensionality of the multivariate data set and determine the grouping of the studied treatment groups in the multivariate trait space. The classification tree based on all the studied traits was built using Classification and Regression Tree (CART) analysis (as implemented in Statistica v. 10.0) using raw (non-transformed) data. CART analysis creates a tree based on a set of decision rules that identify homogeneous groups as a function of a set of explanatory variables (e.g. the values of specific biomarkers) and permits identification of a single most distinguishing characteristic at each split of the classification tree69. All statistical analyses were performed with Statistica v. 10.0. Differences were considered significant if the probability of Type I error was less than 0.05. For all traits and all experimental treatment groups, sample size was 8–13 except of immunoblotting analysis where sample size was 5. The data are presented as means ± standard error (SE) unless indicated otherwise.
Additional Information
How to cite this article: Falfushynska, H. I. et al. Long-Term Acclimation to Different Thermal Regimes Affects Molecular Responses to Heat Stress in A Freshwater Clam Corbicula Fluminea. Sci. Rep. 6, 39476; doi: 10.1038/srep39476 (2016).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Woodward, G., Perkins, D. M. & Brown, L. E. Climate change and freshwater ecosystems: impacts across multiple levels of organization. Philosophical Transactions of the Royal Society B: Biological Sciences 365, 2093–2106, doi: 10.1098/rstb.2010.0055 (2010).
IPCC. IPCC - Intergovernmental Panel on Climate Change 5th Assessment Report (http://www.ipcc.ch/, 2014).
Bozinovic, F. & Pörtner, H.-O. Physiological ecology meets climate change. Ecology and Evolution 5, 1025–1030, doi: 10.1002/ece3.1403 (2015).
Munday, P. L., Warner, R. R., Monro, K., Pandolfi, J. M. & Marshall, D. J. Predicting evolutionary responses to climate change in the sea. Ecology Letters 16, 1488–1500, doi: 10.1111/ele.12185 (2013).
Osovitz, C. J. & Hofmann, G. E. Marine macrophysiology: Studying physiological variation across large spatial scales in marine systems. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 147, 821–827, doi: 10.1016/j.cbpa.2007.02.012 (2007).
Peck, L., Clark, M. S., Morley, S. A., Massey, A. & Rossetti, H. Animal temperature limits and ecological relevance: effects of size, activity and rates of change. Functional Ecology 23, 248–256, doi: 10.1111/j.1365-2435.2008.01537.x (2009).
Langford, T. Ecological Effects of Thermal Discharges. 468, pp. (Springer., 1990).
Falfushynska, H. et al. Habitat pollution and thermal regime modify molecular stress responses to elevated temperature in freshwater mussels (Anodonta anatina: Unionidae). Science of The Total Environment 500–501, 339–350, doi: 10.1016/j.scitotenv.2014.08.112 (2014).
Viergutz, C., Linn, C. & Weitere, M. Intra- and interannual variability surpasses direct temperature effects on the clearance rates of the invasive clam Corbicula fluminea. Marine Biology 159, 2379–2387, doi: 10.1007/s00227-012-1902-0 (2012).
Vaughn, C. C. & Hakenkamp, C. C. The functional role of burrowing bivalves in freshwater ecosystems. Freshw Biol 46, doi: 10.1046/j.1365-2427.2001.00771.x (2008).
Fox, R. O. The Corbicula story: chapter three. Fourth Annual Meeting, Western Society of Malacologists. 5, pp (1971).
Sousa, R., Antunes, C. & Guilhermino, L. Ecology of the invasive Asian clam Corbicula fluminea (Müller, 1774) in aquatic ecosystems: an overview. Ann Limnol 44, doi: 10.1051/limn:2008017 (2008).
Atkinson, C. L., First, M. R., Covich, A. P., Opsahl, S. P. & Golladay, S. W. Suspended material availability and filtration–biodeposition processes performed by a native and invasive bivalve species in streams. Hydrobiologia 667, 191–204, doi: 10.1007/s10750-011-0640-5 (2011).
Beaver, J. R., Crisman, T. L. & Brock, R. J. Grazing Effects of an Exotic Bivalve (Corbicula fluminea) on Hypereutrophic Lake Water. Lake and Reservoir Management 7, 45–51, doi: 10.1080/07438149109354253 (1991).
Semenza, G. L. Hydroxylation of HIF-1: Oxygen Sensing at the molecular level. Physiology 19, 176–182 (2004).
van Hellemond, J. J., van der Klei, A., van Weelden, S. W. H. & Tielens, A. G. M. Biochemical and evolutionary aspects of anaerobically functioning mitochondria. Philosophical Transactions of the Royal Society B: Biological Sciences 358, 205–215, doi: 10.1098/rstb.2002.1182 (2003).
de Zwaan, A. Molluscs. In: Metazoan Life without Oxygen. C.Bryant [Ed.]. Chapman and Hall, London etc., 186–217 (1991).
Sokolova, I. M. & Pörtner, H. O. Temperature effects on key metabolic enzymes in Littorina saxatilis and L. obtusata from different latitudes and shore levels. Marine Biology 139, 113–126 (2001).
Sokolova, I. M., Sukhotin, A. A. & Lannig, G. Stress effects on metabolism and energy budgets in mollusks In: Oxidative Stress in Aquatic Ecosystems. Abele, D., Zenteno-Savin, T., Vazquez-Medina, J. (eds). Blackwell Wiley, Boston etc, 263–280 (2011).
Araújo, M. B. et al. Heat freezes niche evolution. Ecology Letters 16, 1206–1219 (2013).
Roberts, D. A., Hofmann, G. E. & Somero, G. N. Heat-Shock Protein Expression in Mytilus californianus: Acclimatization (Seasonal and Tidal-Height Comparisons) and Acclimation Effects. Biol Bull 192, 309–320 (1997).
Tomanek, L. & Somero, G. Evolutionary and acclimation-induced variation in the heat-shock responses of congeneric marine snails (genus Tegula) from different thermal habitats: implications for limits of thermotolerance and biogeography. J Exp Biol 202, 2925–2936 (1999).
Snyder, M. J., Girvetz, E. & Mulder, E. P. Induction of Marine Mollusc Stress Proteins by Chemical or Physical Stress. Archives of Environmental Contamination and Toxicology 41, 22 (2001).
Tomanek, L. The Heat-Shock Response: Its Variation, Regulation and Ecological Importance in Intertidal Gastropods (genus Tegula)1. Integrative and Comparative Biology 42, 797–807, doi: 10.1093/icb/42.4.797 (2002).
Dahlhoff, E. P. Biochemical indicators of stress and metabolism: Applications for marine ecological studies. Annual Review of Physiology 66, 183–207, doi: 10.1146/annurev.physiol.66.032102.114509 (2004).
Sörensen, J. G., Kristensen, T. N. & Loeschcke, V. The evolutionary and ecological role of heat shock proteins. Ecology Letters 6, 1025–1037 (2003).
Pelham, H. R. B. Functions of the hsp70 protein family: An overview. In: Stress Proteins In Biology and Medicine. Morimoto Morimoto, R. I., Tissieres, A., Georgopoulos, C. (eds). Cold Spring Harbor laboratory Press, Cold Spring Harbor. 450, pp, 287–299 (1990).
Huey, R. B. & Bennett, A. F. Physiological adjustments to fluctuating thermal environmnets: An ecological and evolutionary perspective. In: Stress Proteins In Biology and Medicine. Morimoto, R. I., Tissieres, A. & Georgopoulos, C. (eds). Cold Spring Harbor laboratory Press, Cold Spring Harbor. 450, pp, 37–59 (1990).
Osovitz, C. J. & Hofmann, G. E. Thermal history-dependent expression of the hsp70 gene in purple sea urchins: Biogeographic patterns and the effect of temperature acclimation. Journal of Experimental Marine Biology and Ecology 327, 134–143 (2005).
Lund, S. G., Ruberté, M. R. & Hofmann, G. E. Turning up the heat: The effects of thermal acclimation on the kinetics of hsp70 gene expression in the eurythermal goby, Gillichthys mirabilis. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 143, 435–446 (2006).
Zerebecki, R. A. & Sorte, C. J. B. Temperature Tolerance and Stress Proteins as Mechanisms of Invasive Species Success. PLoS ONE 6, e14806 (2011).
Payton, S. L., Johnson, P. D. & Jenny, M. J. Comparative physiological, biochemical, and molecular thermal stress response profiles for two Unionid freshwater mussel species. The Journal of Experimental Biology, doi: 10.1242/jeb.140129 (2016).
Larade, K. & Storey, K. B. Arrest of transcription following anoxic exposure in a marine mollusc. Molecular and Cellular Biochemistry 303, 243–249 (2007).
Larade, K. & Storey, K. B. Reversible suppression of protein synthesis in concert with polysome disaggregation during anoxia exposure in Littorina littorea. Molecular and Cellular Biochemistry 232, 121–127 (2002).
Tomanek, L. & Somero, G. N. Evolutionary and acclimation-induced variation in the heat-shock pesponses of congeneric marine snails (genus Tegula) from differet thermal habitats: Implications for limits of thermotolerance and biogeography. Journal of Experimental Biology 202, 2925–2936 (1999).
Youker, R. T. & Brodsky, J. L. In Cell Stress Proteins (ed Calderwood, Stuart K. ) 209–227 (Springer New York, 2007).
Fan, C.-Y., Lee, S. & Cyr, D. M. Mechanisms for regulation of Hsp70 function by Hsp40. Cell Stress & Chaperones 8, 309–316 (2003).
Roberts, D. A., Hofman, G. E. & Somero, G. N. Heat-shock protein expression in Mytilus californianus: acclimatisation (seasonal and tidal-height comparisons) and acclimation effects. Biological Bulletin 192, 309–320 (1997).
Bakthisaran, R., Tangirala, R. & Rao, C. M. Small heat shock proteins: Role in cellular functions and pathology. Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics 1854, 291–319, doi: 10.1016/j.bbapap.2014.12.019 (2015).
Kim, Y. E., Hipp, M. S., Bracher, A., Hayer-Hartl, M. & Hartl, F. U. Molecular Chaperone Functions in Protein Folding and Proteostasis. Annual Review of Biochemistry 82, 323–355, doi: 10.1146/annurev-biochem-060208-092442 (2013).
Morrow, G., Inaguma, Y., Kato, K. & Tanguay, R. M. The Small Heat Shock Protein Hsp22 of Drosophila melanogaster Is a Mitochondrial Protein Displaying Oligomeric Organization. Journal of Biological Chemistry 275, 31204–31210, doi: 10.1074/jbc.M002960200 (2000).
Sanyal, A. et al. Heat shock protein HSP60 can alleviate the phenotype of mitochondrial RNA-deficient temperature-sensitive mna2 pet mutants. Molecular and General Genetics MGG 246, 56–64, doi: 10.1007/bf00290133 (1995).
Choresh, O., Ron, E. & Loya, Y. The 60-kDa Heat Shock Protein (HSP60) of the Sea Anemone Anemonia viridis: A Potential Early Warning System for Environmental Changes. Marine Biotechnology 3, 501–508, doi: 10.1007/s10126-001-0007-4 (2001).
Yang, Y. n. et al. Characterization and expression of SpHsp60 in hemocytes after challenge to bacterial, osmotic and thermal stress from the mud crab Scylla paramamosain. Fish & Shellfish Immunology 35, 1185–1191, doi: 10.1016/j.fsi.2013.07.029 (2013).
Ivanina, A. V., Cherkasov, A. S. & Sokolova, I. M. Effects of cadmium on cellular protein and glutathione synthesis and expression of stress proteins in eastern oysters, Crassostrea virginica Gmelin. J Exp Biol 211, 577–586, doi: 10.1242/jeb.011262 (2008).
Kammenga, J. E., Arts, M. S. J. & Oude-Breuil, W. J. M. HSP60 as a Potential Biomarker of Toxic Stress in the Nematode Plectus acuminatus. 34, 253–258 (1998).
de Vooys, C. G. N. Anaerobic metabolism in sublittoral living Mytilus galloprovincialis Lam. in the Mediterranean. II. Partial adaptation of pyruvate kinase and phosphoenolpyruvate carboxykinase. Comparative Biochemistry and Physiology 65 B, 513–518 (1980).
Hardie, D. G. AMPK—Sensing Energy while Talking to Other Signaling Pathways. Cell Metabolism 20, 939–952, doi: 10.1016/j.cmet.2014.09.013 (2014).
Han, G.-d., Zhang, S., Marshall, D. J., Ke, C.-h. & Dong, Y.-w . Metabolic energy sensors (AMPK and SIRT1), protein carbonylation, and cardiac failure as biomarkers of thermal stress in an intertidal limpet: linking energetic allocation with environmental temperature during aerial emersion. The Journal of Experimental Biology, doi: 10.1242/jeb.084269 (2013).
Frederich, M., O’Rourke, M. R., Furey, N. B. & Jost, J. A. AMP-activated protein kinase (AMPK) in the rock crab, Cancer irroratus: an early indicator of temperature stress. Journal of Experimental Biology 212 (2009).
Lahiri, S. et al. Oxygen sensing in the body. Progress In Biophysics & Molecular Biology 91, 249–286 (2006).
Wenger, R. H. Cellular adaptation to hypoxia: O2-sensing protein hydroxylases, hypoxia-inducible transcription factors, and O2-regulated gene expression. FASEB J. 16, 1151–1162 (2002).
Crombie, T. A., Tang, L., Choe, K. P. & Julian, D. Inhibition of the oxidative stress response by heat stress in Caenorhabditis elegans. Journal of Experimental Biology, doi: 10.1242/jeb.135327 (2016).
Abele, D., Burlando, B., Viarengo, A. & Pörtner, H. O. Exposure to elevated temperature and hydrogen peroxide elicits oxidative stress and antioxidant response in the antarctic intertidal limpet Nacella concinna. Comp Biochem Physiol 120B, 425–435 (1998).
Abele, D., Heise, K., Portner, H. O. & Puntarulo, S. Temperature-dependence of mitochondrial function and production of reactive oxygen species in the intertidal mud clam Mya arenaria. J Exp Biol 205, 1831–1841 (2002).
Heise, K., Puntarulo, S., Poertner, H. O. & Abele, D. Production of reactive oxygen species by isolated mitochondria of the Antarctic bivalve Laternula elliptica (King and Broderip) under heat stress. Comp Biochem Physiol C 134, 79–90 (2003).
Cherkasov, A. A., Overton, R. A. Jr., Sokolov, E. P. & Sokolova, I. M. Temperature dependent effects of cadmium and purine nucleotides on mitochondrial aconitase from a marine ectotherm, Crassostrea virginica: a role of temperature in oxidative stress and allosteric enzyme regulation. Journal of Experimental Biology 210, 46–55 (2007).
Sanni, B., Williams, K., Sokolov, E. P. & Sokolova, I. M. Effects of acclimation temperature and cadmium exposure on mitochondrial aconitase and LON protease from a model marine ectotherm, Crassostrea virginica. Comparative Biochemistry and Physiology, Part C 147, 101–112 (2008).
Monserrat, J. et al. In Oxidative Stress in Aquatic Ecosystems 317–326 (John Wiley & Sons, Ltd, 2011).
Lushchak, V. I. Environmentally induced oxidative stress in aquatic animals. Aquatic Toxicology 101, 13–30, doi: 10.1016/j.aquatox.2010.10.006 (2011).
Halliwell, B. & Gutteridge, J. M. C. Free Radicals in Biology and Medicine. (Oxford University Press, 1999).
APHA. American Public Health Association. Standard methods for the examination of water and wastewater. 20th edn (American Public Health Association, 1998).
Sokolova, I. M. & Pörtner, H. O. Metabolic plasticity and critical temperatures for aerobic scope in a eurythermal marine invertebrate (Littorina saxatilis, Gastropoda: Littorinidae) from different latitudes. Journal of Experimental Biology 206, 195–207 (2003).
Lannig, G., Eilers, S., Pörtner, H. O., Sokolova, I. M. & Bock, C. Impact of ocean acidification on energy metabolism of oyster, Crassostrea gigas—changes in metabolic pathways and thermal response. Marine Drugs 8, 2318–2339 (2010).
de Zwaan, A. & Wijsman, T. C. M. Anaerobic metabolism in bivalvia (Mollusca) Characteristics of anaerobic metabolism. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology 54, 313–323 (1976).
Ivanina, A. V. & Sokolova, I. M. Effects of cadmium exposure on expression and activity of P-glycoprotein in eastern oysters, Crassostrea virginica Gmelin. Aquatic Toxicology 88, 19–28, doi: 10.1016/j.aquatox.2008.02.014 (2008).
Pfaffl, M. W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research 29, 2002–2007 (2001).
Matoo, O. B., Ivanina, A. V., Ullstad, C., Beniash, E. & Sokolova, I. M. Interactive effects of elevated temperature and CO2 levels on metabolism and oxidative stress in two common marine bivalves (Crassostrea virginica and Mercenaria mercenaria). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 164, 545–553, doi: 10.1016/j.cbpa.2012.12.025 (2013).
Breiman, L., Friedman, J. M., Stone, C. J. & Olshen, R. A. Classification and Regression Trees. (Chapman and Hall/CRC, 1998).
Acknowledgements
This work was supported by the U.S. Civil Research and Development Foundation awards FSCX-15-61168-0, OISE-15-61170-0 and UKB1-7109-TE-13, and Fulbright Scholarship and Ministry of Education and Science of Ukraine (#130B) to H.F. We thank Zachary Hall (Duke Energy) for assisting with the access to the CWD site, Dr. Didier Dréau for assistance with the dot-blot technique.
Author information
Authors and Affiliations
Contributions
I.M.S., H.F. and T.P. designed the experiments, H.F. and T.P. conducted the experimental work, I.M.S. and H.F. carried out the data analysis, I.M.S. wrote the first draft of the manuscript. All authors participated in the discussion of the data and revisions of the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Falfushynska, H., Phan, T. & Sokolova, I. Long-Term Acclimation to Different Thermal Regimes Affects Molecular Responses to Heat Stress in a Freshwater Clam Corbicula Fluminea. Sci Rep 6, 39476 (2016). https://doi.org/10.1038/srep39476
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep39476
This article is cited by
-
Multibiomarker assessment in zebrafish Danio rerio after the effects of malathion and chlorpyrifos
Toxicology and Environmental Health Sciences (2021)
-
Divergence and plasticity shape adaptive potential of the Pacific oyster
Nature Ecology & Evolution (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
