
Abstract
17β-Estradiol receptors have been found in several brain nuclei including the suprachiasmatic nucleus (SCN) of mammalian species. The SCN is believed to act as brain clock regulating circadian and circannual biological rhythms, such as body temperature, sleep, and mood. Here, we examined whether 17β-estradiol (E2) could affect cell excitability and synaptic transmission in the SCN. Bath application of E2 (0.03–3 μM) increased the spontaneous firing frequency and depolarized cell membrane of the SCN neurons significantly. Furthermore, E2 (0.03–3 μM) increased (by about 25–150% of control) frequency of the miniature excitatory postsynaptic currents. Amplitude of the evoked excitatory postsynaptic currents was enhanced (by about 32% of control) after exposure to 1 μM E2. The paired-pulse ratio was reduced by E2. These effects were prevented by the estrogen receptor antagonist, ICI 182780. Exposure to the biologically inactive 17α-estradiol did not cause any significant changes in the parameters mentioned above. These findings are in favor of an implication of estrogen in modulation of neuronal activity in SCN and possibly regulating circadian rhythms.
Similar content being viewed by others
INTRODUCTION
The suprachiasmatic nucleus (SCN), a paired-hypothalamic structure lying dorsal to the optic chiasm is referred as the clock of the brain orchestrating circadian and circannual biological rhythms, such as the rhythms of hormones, body temperature, sleep, and mood (Card and Moore, 1991; Morin, 1994). One prominent feature of neurons in the SCN is the circadian rhythm in spontaneous firing frequency (SFF). The oscillations of the SFF in the SCN have been verified by both in vitro and in vivo extracellular electrophysiological recordings (Inouye and Kawamura, 1979; Green and Gillette, 1982; Groos and Hendriks, 1982; Bos and Mirmiran, 1990). The phase of the rhythm is shown to be set by light signals which cause glutamate release from terminals of retinal ganglion neurons with projections to the ventral part of the SCN (Castel et al, 1993; Shirakawa and Moore, 1994). The output of the SCN is suggested to be translated into synchronization of the metabolic and hormonal activity and subsequently behavioral reactions of the animal to the external light–dark cycle (see Van Esseveldt et al, 2000). Significant variations in membrane potential and input resistance recorded from mammalian SCN neurons during day and night have been reported (Jiang et al, 1997; De Jeu et al, 1998; Kuhlman and McMahon, 2004). It has also been demonstrated that as many as one-third of the neurons afferent to the SCN in some parts of the preoptic area and the corticomedial amygdala are estrogen receptor α-immunoreactive (Kruijver and Swaab, 2002). Some investigations suggested that variations in circadian neuronal activity in the SCN could be due to sex hormonal regulation (Zucker et al, 1980; Kow and Pfaff, 1984; Su et al, 2001). A link between the circadian rhythms and circadian oscillations of neuronal excitability has been proposed by some researchers (eg Bouskila and Dudek, 1995). Furthermore, the presence of estrogen receptor (α and β) among neurons intrinsic to the SCN of human and rat (Su et al, 2001; Kruijver and Swaab, 2002) supports the hypothesis that these receptors may play a crucial role in regulating neuronal activity and rhythmic functions of the SCN neurons. Although clinical and experimental studies support that the gonadal hormone, 17β-estradiol (E2), influences circadian rhythms (Zacharieva et al, 2002; Perrin et al, 2006), it is not well established yet whether this is by a direct effect on neuronal activity in the SCN. Recent studies demonstrated that E2 could affect neuronal excitability and synaptic transmission in some brain regions such as hippocamp and the parabrachial nucleus (Carrer et al, 2003; Fatehi et al, 2006). Therefore, in view of the postulated role of estrogen on circadian rhythms, to determine whether estrogen could affect neuronal activity and neurotransmission in the SCN, the present study was designed. Using whole-cell patch-clamp recording techniques, we tested the hypothesis that in vitro electrophysiological properties of neuronal cells and synaptic transmission in the SCN could be affected by exposure to exogenous E2. The extended objective of this study was to determine the mechanisms of possible actions by which E2 might regulate neuronal activity in the SCN.
MATERIALS AND METHODS
Slice Preparation
The handling and maintenance of animals met the guidelines of the Canadian Council on Animal Care. Efforts were made to minimize the number of animals used and their suffering. Male Sprague–Dawley rats (Charles River, Montreal, QC, Canada) weighing 150–250 g were maintained on a 12-h light/dark (lights on at 8 a.m.) schedule. Animals were deeply anesthetized with isoflurane (Abbott Laboratories, Saint-Laurent, QC, Canada) vapor in a closed environment and then decapitated between 9 and 10 a.m. The brain was removed and placed in ice-cold (2–3°C) artificial cerebrospinal fluid (aCSF) containing (in mM): 125 NaCl, 2.5 KCl, 11 D-glucose, 18 NaHCO3, 1.2 NaH2PO4, 1.2 MgCl2, 2.5 CaCl2, equilibrated with 95% O2–5% CO2 (carbogen), pH 7.2–7.3; osmolality, 285–295 mOsmol/l. Coronal slices (250–300 μm thick) of hypothalamus containing the SCN were prepared from brains maintained in cold aCSF bubbled with carbogen, using a vibratome (Model 1000 plus, Ted Pella Inc., Redding, CA, USA). Slices were incubated in a temperature-controlled chamber (30–32°C) containing aCSF bubbled with carbogen for at least 1 h before patch-clamp recordings. A single slice was then transferred to a 500 μl recording chamber on the stage of an Olympus microscope, held in place with a nylon net mounted on a stainless-steel ring and continuously superfused with aCSF (saturated with carbogen) at a flow rate of 2–3 ml/min. The SCN neurons were visualized using infrared differential interference contrast video-microscopy.
Electrophysiological Recordings and Analysis
Patch micropipettes were pulled from thin-walled (outer diameter, 1.5 mm) glass capillary tubes (KG-33; Garner Glass Co., Claremont, CA, USA) by a Flaming–Brown puller (Model P-87; Sutter Instruments Co., Novato, CA, USA). The composition of the internal solution was (in mM): 130 K-gluconate, 6 NaCl, 10 HEPES, 2.5 Na-ATP, 0.1 Na-GTP; pH and osmolarity adjusted to 7.3 (with KOH) and 280–290 mOsmol/l, respectively. The external solution had the same composition as that used for slice preparation. When filled with internal solution the micropipettes had resistance of 4–6 MΩ. Junction potentials were measured and zeroed by a function key on seal test window of Clampex 9, which control a MultiClamp 700B amplifier. A seal resistance in excess of 2 GΩ was obtained before moving to the whole-cell configuration. All experiments were carried out at temperatures of 32±1°C. The fast electrode capacitance was first compensated before breaking into the cell and access resistance routinely ranged between 15 and 25 MΩ. After whole-cell configuration was achieved, capacitive transients were cancelled by using an automatic compensation function of the MultiClamp 700B (about 70–80%) and were monitored periodically. Access (series) and input resistances of all cells were also monitored and recorded periodically throughout the experiment by applying a 20 mV hyperpolarizing pulse for 20–40 ms. Only those cells that showed less than 10% change in access resistance over the period of experiments were included in the analysis of the data. Synaptic responses of the SCN neurons were evoked by electrical stimulation using a twisted bipolar tungsten electrode connected to a programmable digital pulse generator (Master-8, AMPI) which was placed into the lateral SCN, approximately 150 μm dorsal and lateral to the recording site in the ventromedial SCN. For synaptic current recordings, all cells were voltage clamped at a holding potential (Vh) of −60 mV. Data acquisition and analysis were performed using Clampex and Clampfit 9.2 softwares, respectively (Axon Instruments, Union City, CA, USA). Whole-cell currents and voltages were low-pass filtered at 2–5 kHz and acquired at a sampling rate of 1–10 kHz depending on the signal speed.
Evoked excitatory postsynaptic current (EPSC) amplitudes were measured from baseline to peak and considered as the excitatory synaptic strength at the chosen stimulus intensity. To calculate paired-pulse ratios, the mean amplitude of at least four EPSC2s was divided by the mean amplitude of at least four EPSC1s. Responses were normalized by taking the mean of the last four responses before drug application and dividing the rest of the values by this mean. Therefore, the pooled data are expressed as mean percentage change from control values±standard error of the mean (SEM). Each individual cell served as its own internal control. Results expressed as percentages of control were considered to be nonparametric data and analyzed by employing the Mann–Whitney U-test. Statistical significance was determined at p⩽0.05. Graphing was performed using SigmaPlot® and CorelDraw® softwares.
Drugs
Drugs were applied to the cells by bath perfusion of the slices with aCSF containing the final concentration of the drug. Isoflo (isoflurane) was purchased from Abbott Laboratories (Saint-Laurent). ICI 182780 was obtained from Tocris (Ellisville, MO, USA). α-Amino-3-hydroxy-5-methylisoxazole-4-propionic acid hyrobromide (AMPA), 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX), D(−)-2-amino-5-phosphonopentanoic acid (D-APV), 17β-estradiol, 17α-estradiol, picrotoxin, tetrodotoxin (TTX), and all other chemicals were purchased from Sigma (St Louis, MO, USA). Appropriate stock solutions were made and diluted with aCSF just before application.
RESULTS
Data described here were extracted from either current- or voltage-clamp experiments on neurons located in the ventromedial part of the SCN. Electrophysiological criteria set for accepting cells were resting membrane potential (RMP) more negative than −40 mV, spontaneous action potential amplitude higher than 60 mV measured from the RMP, and EPSC latency less than 5 ms. Under our experimental conditions, 76% of the SCN neurons spontaneously fired action potentials. For voltage-clamp experiments, holding potential was −60 mV. Since we were interested in non-NMDA receptor-mediated synaptic responses, a selective NMDA receptor blocker, D-APV (50 μM), and a GABAA receptor blocker, picrotoxin (50 μM), were present in the perfusing solution through the patch-clamp recording experiments. To verify that the negative current signal following the stimulus artifact recorded under voltage-clamp conditions was a synaptic response, in a few experiments either TTX (1 μM) or cadmium (100 μM) was added to the perfusing solution, resulting in a complete elimination of the signal. Thus, taking the latter observations and disappearance of miniature inward currents after bath application of 10 μM CNQX (n=4, data not shown), we were confident that the recorded currents were glutamate non-NMDA receptor-mediated EPSCs and miniature EPSCs (mEPSCs). Of a total of 98 cells exposed to E2, 79 cells (about 80%) exhibited altered electrophysiological properties.
Effects on Spontaneous Action Potentials and Resting Membrane Potentials
To explore if E2 affect cell excitability and pattern of spontaneous action potential, various concentrations (0.01–3 μM) of the steroid were applied through continuous superfusion of the brain slices.
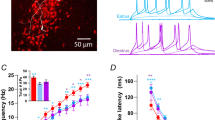
As it has been reported by others (eg Thomson et al, 1984; Kononenko and Dudek, 2004), in the absence of picrotoxin in superfusing aCSF, we observed different firing patterns of spontaneous action potentials recorded from the SCN neurons. Some neurons fired spontaneous action potentials at a clock-like pattern (regular interspike intervals) and others either showed no spontaneous activity or fired spontaneous action potentials at irregular intervals (data not shown). For E2 treatment, we only proceeded with those neurons, which had a regular pattern of interspike intervals. The control level of SFF in these neurons was normally higher than 4 Hz. The representative traces of potentials before and after E2 are shown in Figure 1A. The interspike intervals histograms (Figure 1B) demonstrate that the steroid reduces intervals of spontaneous firings. Current-clamp experiments revealed that E2 (1 μM) increased SFF to 218.7±5.0% of control (Figure 1, p<0.05; n=5). There was no significant difference between changes in firing rate measured in the presence of 1 and 3 μM E2, implying that the maximum effect was achieved at concentration of 1 μM. Figure 1C illustrates time–effect relationship of 1 μM E2 on changes in SFF. A quantitative and summarized presentation of differences in the firing frequency recorded under control conditions and 5 min after exposure to various concentrations of E2 is shown in Figure 1D. We plotted a sigmoidal graph based on changes (percentage of maximum effect) in firing rate induced by various concentrations of E2 to obtain an EC50 value of 0.15 μM for the steroid (Figure 1E). Moreover, application of 1 μM E2 had also a profound effect on the afterhyperpolarization (AHP) part of the action potentials, attenuating by 57.4±6.3% of control (Figure 1F and G). A significant change in the duration of spontaneous action potentials was observed after exposure to E2. For instance, the duration of action potentials in the middle of the signals was increased from 1.2±0.3 to 2.3±0.4 ms when recording was performed in the presence of 1 μM E2 (p<0.05; n=4). To determine whether these effects of the hormone were receptor-mediated, we treated slices with E2 following pretreatment with aCSF containing the estradiol receptor antagonist, ICI 182780 (5 μM, dissolved in 0.05% v/v DMSO). Similar to our previous finding (Fatehi et al, 2006), exposure of the brain slices to DMSO alone (up to 0.1% v/v), or ICI 182780 (5 μM dissolved in 0.05% v/v DMSO) had no significant effect on membrane potentials or on synaptic currents recorded from the SCN neurons (data not shown). The effects of 1 μM E2 on the firing rate and amplitude of AHP were blocked in the presence of 5 μM ICI 182780 (Figure 1A, B, D and G).

Effects of 17β-estradiol (E2) on the frequency and pattern of spontaneous action potentials fired by SCN neurons. (A) Representative traces of potential recordings from two different SCN neurons in the absence (top) and presence (bottom) of 5 μM ICI 182780 under current-clamp conditions before (left panel) and after (right panel) exposure to 1 μM E2. (B) Interspike interval histograms constructed using data obtained from the experiments shown in A illustrate that E2 reduced interspike intervals only in the absence of antagonist ICI 182780. Black and gray bars represent data before and after exposure to E2, respectively. (C) An averaged time–effect plot generated from four cells, which were exposed to 1 μM E2 for the duration indicated by the length of the line. (D) A bar graph summarizing effects of various concentrations of E2 on spontaneous firing frequency (SFF) recorded from the SCN neurons (n=3–4). Note lack of effect in the presence of 5 μM ICI 182780 (1+ICI). (E) A concentration–response relationship on a logarithmic scale constructed to calculate EC50 of 0.15 μM for E2 using data presented in D (n=3–4). (F) The superimposed examples of spontaneous action potentials before (a) and after (b) exposure to 1 μM E2. Note reduction in the afterhyperpolarization potential (AHP) amplitude and increase in the duration of action potential. (G) A bar graph summarizing effects of 1 μM E2 on AHP amplitude in the absence and presence of ICI 182780 (*p<0.05, **p<0.01; n=4).
The RMPs of the SCN neurons measured under control conditions ranged from −44 to −53 mV (−48±4 mV, n=64). Examples of voltage recording from two different cells exposed to 1 μM E2 in the absence and presence of ICI 182780, an estrogen receptor antagonist, are shown in Figure 2a. These recordings were performed while brain slices were superfused by aCSF containing 1 μM TTX. As it could be seen clearly from these traces, a pretreatment with the estrogen receptor antagonist prevented membrane depolarization by E2. E2 (0.03–3 μM) caused a concentration-dependent decrease in RMP. An EC50 of 0.21 μM was found when the percentage of maximum membrane depolarization was plotted against various concentrations of E2 (Figure 2b). Membrane depolarization occurred within 2 min and reached a plateau within 3 min of exposure to the steroid (Figure 2c). The input resistance of the cells before and after exposure to E2 (1 μM) was 994±53 MΩ and 1373±67 MΩ (n=5), respectively.

Effects of 17β-estradiol (E2) on the resting membrane potential (RMP) of SCN neurons. (a) Example traces of continuous potential recordings from two different neurons in the absence (top panel) and presence (bottom panel) of 5 μM ICI 182780. (b) A concentration–response relationship on a logarithmic scale constructed to calculate EC50 of 0.21 μM for E2 (n=3–5). (c) An averaged time–effect plot generated from five cells exposed to 1 μM E2 for the duration indicated by the length of the line.
Effects on Excitatory Postsynaptic Currents
Following our observation on the pattern of spontaneous firing, voltage-clamp experiments were performed to examine if synaptic events were susceptible to increased excitability of neurons induced by E2. mEPSCs were recorded from the SCN neurons before and after various concentrations of E2. These currents could be blocked by application of 10 μM CNQX (non-N-methyl-D-aspartate acid, a non-NMDA receptor blocker). In the presence of 1 μM TTX, 50 μM picrotoxin, and 50 μM 4-APV, E2 (0.03–3 μM) increased frequency of the mEPSCs under holding potential of −60 mV (Figure 3, *p<0.05, **p<0.01; n=3–4). Following exposure to 1 μM E2, frequency of the mEPSCs was increased from 1.8±0.3 Hz in the control to 4.3±0.4 Hz (p<0.05; n=4). It is illustrated in Figure 3 that application of 1 μM E2 increases the frequency of mEPSCs without altering their amplitude and time course. However, when slice preparations were pretreated with 5 μM ICI 182780, exposure to 1 μM E2 did not alter the frequency of mEPSCs (Figure 3e).

Effect of 17β-estradiol (E2) on the miniature excitatory postsynaptic currents (mEPSCs) recorded from SCN neurons. (a) Representative current traces recorded from a cell (Vh=−60 mV) in the presence of picrotoxin (50 μM), TTX (1 μM), and APV (50 μM) show an enhancement of the frequency of mEPSCs after exposure to 1 μM E2. The frequency of mEPSCs was 1.5 Hz in control (left panel) and 3.0 Hz in the presence of E (middle panel). mEPSCs were blocked by the bath application of 20 μM CNQX (right panel). (b) mEPSCs’ interval histograms constructed using data obtained from the experiments shown in A illustrate that E2 reduces the events intervals. Black and gray bars represent data before and after exposure to E2, respectively. (c) The superimposed average mEPSCs before and after exposure to E2 showing no significant change in the amplitude and the time course of mEPSCs is induced by 1 μM E2. (d) mEPSCs’ amplitude histograms before (black bars) and after (gray bars) exposure to E2, data were obtained from the experiment shown in A. (e) A concentration–response bar graph summarizes the effect of various concentrations of E2 on the mEPSCs’ frequency (*p<0.05, **p<0.01; n=3–5). (f) An averaged time–effect plot generated from four cells, which were exposed to 1 μM E2 for the duration indicated by the length of the line.
To further investigate whether the evoked excitatory transmission could also be affected by the hormone, we looked for any possible changes in the synaptic responses to electrical stimulation. Again in this set of experiments, picrotoxin (50 μM) and AVP (50 μM) were added to the aCSF thereby eliminating inhibitory postsynaptic currents and NMDA receptor-mediated currents. At a holding potential of −60 mV and inclusion of picrotoxin, the evoked synaptic events were believed to be predominately non-NMDA receptor-mediated excitatory currents (EPSCs). Synaptic events recorded under voltage clamp at this holding potential were inward currents. The evoked EPSCs were fast-rising and slowly decaying signals at a holding potential more negative than −50 mV. Application of E2 (0.3 and 1 μM) for 3 min increased the amplitude of EPSCs (Figure 4A and B). An analysis of the current–voltage (I–V) relationships (n=3) indicated that the potentiation of EPSCs induced by E2 was stronger at more negative holding potentials (Figure 4B). The onset of action was between 1 and 2 min with a peak effect observed after 3–4 min of application. When slices were pretreated with ICI 182780 (5 μM), exposure to 1 μM E2 caused only nonsignificant changes in amplitude of the EPSCs (Figure 4A and B).

Effect of 17β-estradiol (E2) on the evoked excitatory postsynaptic currents (EPSCs) recorded from SCN neurons. (A) Superimposed representative current traces recorded from two different cells: in the absence (left panel) and presence (right panel) of ICI 182780 (Vh=−60 mV) illustrate an increase in the amplitude of EPSCs induced by E2 in the absence of antagonist ICI 182780; the evoked response before (a), after (b) 1 μM E2, and after (c) 20 μM CNQX. APV (50 μM) and picrotoxin (50 μM) were present throughout the experiments. (B) The I (amplitude of EPSCs)–V (holding potential of the cells) relationships before (•) and after (○) exposure to 1 μM E2 (left panel, n=3), and a concentration–response bar graph for the effects of E2 on the evoked EPSCs’ amplitude (right panel, *p<0.05; n=3–4). (C) The representative traces recorded from SCN neurons when the preparations were subjected to a pair of pulses in the absence (top) and presence (bottom) of antagonist 5 μM ICI 182780 before (left panel) and after (right panel) exposure to 1 μM E. Note increase in the amplitude of response to the first stimulation induced by E2 is more profound compared to that in response to the second stimulation. (D) A bar graph summarizing changes in paired-pulse ratio following exposure to 1 μM E2 (*p<0.05; n=3).
Effects on Paired-Pulse Ratio
Further experiments were designed to investigate the locus (presynaptic or postsynaptic) of action of E2. For these experiments paired EPSCs were recorded in response to electrical stimulation of the SCN at inter-stimulus interval of 100 ms and intensity of strong enough (100–400 μA, 100 μs duration) to produce responses of about 60% of the maximal responses. The paired-pulse ratio is a common index used to determine change in neurotransmitter release probability from the presynaptic terminal. Absolute current amplitudes of both responses (EPSC1 and EPSC2) were measured from the baseline current level 3–4 ms after the stimulus artifact to the maximum current amplitude of each signal. No significant current rundown was observed in cells recorded for about 10 min.
Of a total of fifteen cells examined, the paired-pulse ratio of nine cells exhibited reproducible paired-pulse depression (PPD). This PPD occurred less frequently at interstimuls intervals greater than 350 ms (data not shown). The remaining (six) cells that showed no PPD were not included for analysis. This exclusion was done to avoid further possible variation resulted from heterogeneity observed among intra-SCN synapses (Gompf and Allen, 2004). E2 (1 μM) enhanced the amplitude of the first response more profoundly, resulting in a significant decrease (76±5% of control) in the paired-pulse ratio (Figure 4C and D, n=3, p<0.05). This effect of the steroid was prevented by pretreatment of slices with the antagonist ICI 182780 (5 μM).
Lack of Effect on AMPA-Induced Currents
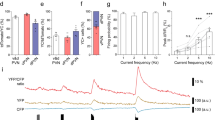
Application of AMPA (10 μM), produced an inward current with averaged amplitude of about 80 pA reaching its peak within 1–2 min. The experiments with AMPA were performed to test if the hormone could affect non-NMDA postsynaptic receptors’ sensitivity. Responses to AMPA were not affected by 1 μM E2 (Figure 5, n=3).

Lack of effect on the exogenous AMPA-induced currents. (a) The example tracings of inward currents induced by 10 μM AMPA in the absence (left) and presence (right) of 1 μM E recorded from a SCN neuron. (b) A summary bar graph showing E had no significant effect on the responses to exogenous application of AMPA (n=3).
The Stereo-Specificity of the Effects
The stereo-specificity for estradiol modulation of neuronal activities and the excitatory neurotransmission in the SCN was tested by exposure of the slice preparations to 17α-estradiol. A total of 17α-estradiol (1 μM) had no significant effect on the RMPs, spontaneous action potentials frequency, and mEPSC frequency (Figure 6).

Lack of effect of 17α-estradiol (17α-E) on the resting membrane potentials (RMP), spontaneous firing frequency (SFF) and mEPSC frequency (mEPSCF). (a–c) Bar graphs summarizing the data related to the experiments examining the effects of 17α-estradiol (0.3 and 1 μM) on RMP, SFF, and mEPSCF.
DISCUSSION
Recognition of multi-facet and complex nongenomic effects of estrogen in the CNS over the past two decades encouraged more sophisticated investigations to understand the underlying mechanisms at cellular and molecular levels (for review see Woolley, 2007). The present study provides a pharmacological profile of E2 for its in vitro effects on the SCN neurons activity. To our best knowledge, so far, no direct observation on the effects of E2 on neuronal excitability and neurotransmission in the SCN has been reported. Considering the presence of estrogen receptors on the SCN neurons, this study may contribute to extension of the current vision on hormonal regulation of circadian rhythms. More specifically, this article describes results obtained from electrophysiological examinations, which confirm that E2 enhances cell excitability and excitatory synaptic transmission in the SCN of rat. Involvement of estrogen receptors in mediating such effects is also discussed here.
Alteration of Cell Excitability and Resting Membrane Potential
We have provided evidence to show that E2 increased cell excitability as revealed by an increase in the frequency of neuronal firing and cell membrane depolarization. It has already been shown that the SCN neurons under in vitro conditions, fire spontaneous action potentials (Inouye and Kawamura, 1979; Green and Gillette, 1982; Jackson et al, 2004; Kononenko and Dudek, 2004). Significant differences between averaged values for the RMPs, input resistance (Rin) and SFF recorded from the SCN neurons during subjective days and nights have been reported frequently (eg Jiang et al, 1997; Kuhlman and McMahon, 2004). There is also a considerable variation among the averaged values of these parameters obtained from recordings performed during the subjective day by various laboratories (RMP ranging from −36 to −60 mV; Rin ranging from 639 to 1820 MΩ; SFF ranging from 3 to 11 Hz for regularly firing neurons) (eg Reynolds and Pinnock, 1997; Cloues and Sather, 2003; Jackson et al, 2004; Pierson et al, 2005; Wang and Huang, 2006). Under our experimental conditions, based on firing rate patterns, we observed three different types of neurons in the SCN, namely silent, regularly, and irregularly firing cells. However, to avoid additional interaction between heterogeneous cell type and effect of the steroid, we proceeded with only those neurons that fired spontaneously at regular intervals. Our averaged control values for RMPs (interspike potentials), input resistance, and frequency of spontaneous firings are in good agreement with those values reported previously by Cloues and Sather (2003), De Jeu et al (2002), and Kononenko and Dudek (2004), respectively. Depolarization induced by E2 may underlie our consistent observation of an increase in the excitability of the SCN neurons. A moderate depolarization (6–7 mV about 15–17% of control RMP) caused by 30 μM strophanthidin, a Na+/K+-ATPase inhibitor, has been shown to be accompanied by a significant increase (about twofold) in the frequency of spontaneous action potentials recorded from the SCN neurons (Wang and Huang, 2006). Interestingly, attenuation of the AHP amplitude caused by E2 (1 μM) is quantitatively comparable to what is reported for the effect of 30 μM strophanthidin on these waveforms recorded from the SCN neurons (Wang and Huang, 2006). Our results on cell excitability is consistent with previous observations on the effects of E2 on hippocampal CA1 pyramidal neurons reported by Carrer et al (2003). The latter study revealed that estradiol-induced suppression of the AHP was paralleled by decreased calcium influx and decreased amplitude of at least one of the calcium-activated potassium currents that might contribute to the AHP. Nevertheless, since it has been shown that E2 occluded an effect of nifedipine to further reduce the AHP, it seems likely that one of proximal targets for E2 is L-type calcium channels (Kumar and Foster, 2002). While the nature of channels involved in circadian regulation of membrane potential is not established yet, several laboratories have identified a number of channels that are involved in circadian regulation of spike frequency. For instances, involvement of the fast delayed rectifier potassium current (Itri et al, 2005) and the large conductance calcium-activated potassium current (Meredith et al, 2006; Pitts et al, 2006) have been documented. We have no direct evidence to support whether these currents in the SCN neurons are modulated by E2. However, attenuation of the AHP by E2 shown in the present study supports the hypothesis that modulation of the ionic currents involved in hyperpolarization after an action potential could contribute to an increase in spike (spontaneous firing) frequency. It is believed that in many cases, an important determinant of neuronal excitability is the AHP (Sah, 1996). The voltage-independent calcium-activated potassium channels mediate some of the underlying potassium currents for the AHP. Nonetheless, another investigation on the effects of various potassium channel blockers on AHP contradicts the hypothesis that blockade of calcium-activated potassium channels by E2 may be a dominant factor for increased rate of the spontaneous firing neurons, because apamin and iberiotoxin apparently do not alter firing frequency in the SCN (Cloues and Sather, 2003). Furthermore, it was shown that E2 (10 μM) had no effect on large-conductance calcium activated-potassium channel activity recorded from cultured vascular endothelial cells (Chiang and Wu, 2001). Qiu et al (2006) found a significant increase in whole-cell T-type current as well as spontaneous firings in arcuate neurons from ovariectomized guinea pigs treated with estrogen for 24 h. They suggested that the increased T-type calcium current and excitability were most likely the results of increased expression of calcium (Cav3.1) channels. Regardless of which ionic channel may be modulated preferentially, demonstrated effects of E2 on the rate and pattern of spontaneous firing, and RMPs altogether are indications of an increased excitability in the SCN neurons.
Facilitation of Excitatory Neurotransmission
Miniature inhibitory and EPSCs have been recorded from the SCN neurons (Kim and Dudek, 1991; Jiang et al, 1997; Lundkvist et al, 2002). Whereas it has been shown that in horizontal slices of rat brain only excitatory synaptic activity in the SCN varied significantly between night and day (Lundkvist et al, 2002), this could not be confirmed in coronal slices (Michel et al, 2002).
Since it was our objective to investigate possible effects of E2 on only excitatory responses, all experiments were performed in the presence of 50 μM picrotoxin. This concentration of picrotoxin has been shown to be sufficient for eliminating GABAA-mediated responses (Glatzer and Smith, 2005). We avoided use of bicuculline, because it was shown recently that bicuculline could also manipulate the SCN neurons excitability in addition to blocking GABA receptors (Kononenko and Dudek, 2004). The increase in frequency of the miniature excitatory currents and in amplitude of the evoked currents presented in this report support the conclusion that E2 facilitates excitatory synaptic neurotransmission within the SCN. These observations suggest that E2 modulates the responsiveness of the SCN neurons to excitatory inputs. We speculate that by acting to enhance the excitatory neurotransmission in the SCN, E2 may influence the function of this nucleus. Understanding such modulatory effects of E2 may be beneficial with respect to hormone replacement. It has been suggested that short-term effects on memory as well as mood regulation by E2 are related to its acute electrophysiological effects in the CNS (Brinton, 2005; see Woolley, 2007). If there is a correlation between production of endogenous E2 and behavior in menopausal women, there will be a greater need for more mechanistic information in order to conclude whether hormone replacement could have a significant impact on behavior.
The observation that neuronal activity and the frequency of mEPSCs did not return to their control levels following washout of estradiol for 10–15 min in this study was consistent with results of some other studies (Woolley et al, 1997; Woolley, 1999). A simple explanation for this phenomenon could be that this hormone might induce some long-lasting changes in the cell including activation of a variety of intracellular signaling pathways (Lee and McEwen, 2001; Simoncini and Genazzani, 2003), which are not reversible within minutes. It is well accepted that estrogen by acting at the membrane can rapidly elicit the activation of various kinases and increase calcium within cells (Falkenstein and Wehling, 2000; Davis et al, 2002). Furthermore, The increased excitability of neurons induced by E2 is blocked by a variety of inhibitors to intracellular signaling cascades such as phospholipase C, protein kinase A, and protein kinase C (Kelly et al, 2002a, 2002b).
An approach to distinguish presynaptic from postsynaptic actions on synaptic transmission is the use of a paired-pulse protocol (Creager et al, 1980; Clark et al, 1994; Glatzer and Smith, 2005; Fatehi et al, 2006). Both facilitation and inhibition of synaptic transmission in various brain regions have been observed after application of E2 (for review, see Woolley, 1999 and Kelly et al, 2003). Some studies seem to indicate that the effect of estradiol on synaptic transmission within central nervous system nuclei is predominantly excitatory (Wong and Moss, 1992; Rudick and Woolley, 2003; Shiroma et al, 2005). However, in a previous study (Fatehi et al, 2006), we provided evidence that E2 at a much higher concentrations (10–100 μM) attenuated excitatory neurotransmission in the parabrachial nucleus of male rats. The latter was in agreement with some other studies, finding an inhibitory action for this hormone in the CNS (Woolley, 1999; Womble et al, 2002; Xue and Hay, 2003).
The fact that postsynaptic responses to AMPA were not affected by pre-exposure to E2 suggests that facilitation of neurotransmission is more likely to be due to a presynaptic action of this neurosteroid. This is supported by the observation of a significant change in the paired-pulse ratio. A postsynaptic effect of the steroid (ie membrane depolarization), which plays a crucial role for cell excitability, seems to be unlikely as a key factor for the facilitation of neurotransmission. It rather appears more intriguing to suggest that presynaptic membrane depolarization possibly induced by the steroid is a prominent cause for the facilitation of neurotransmission in the SCN synapses.
Involvement of Estrogen Receptors and Stereo-Specificity of Actions
The enhancement of neuronal firing and facilitation of the excitatory neurotransmission induced by E2 appeared to be receptor mediated since all these effects were abolished by a pretreatment with the estradiol receptor antagonist, ICI 182780. This is in agreement with other findings, indicating that the modulatory effects of E2 on potassium currents were receptor-mediated since they were blocked by the antagonist ICI 182780 (Li and Hay, 2000). Rapid effects of E2 on various ionic currents recorded from neuronal cells have frequently been reported (eg Kelly et al, 2002a; Fatehi et al, 2005; Kow et al, 2006). Acute (nongenomic) actions of E2 in neurons are probably initiated at cell membrane, but they could trigger or potentiate eventual transcriptional effects. Strong evidence supporting this suggestion comes from a recent study, which demonstrates clearly that a membrane-impermeable estrogen conjugate, 17β-estradiol linked to bovine serum albumin, could potentiate the transcription induced by E2 (Vasudevan et al, 2005).
The lack of effect of 17α-estradiol on the RMPs, spontaneous action potentials and EPSCs recorded from SCN neurons (Figure 6) verified that those effects exerted by 17β-estradiol were not nonspecific steroid effects. Similar stereo-specificity for the modulatory actions of estradiol has been observed in several other studies (eg Li and Hay, 2000; Bryant et al, 2005; Fatehi et al, 2005). This means that the biologically active form of the hormone is capable of controlling or regulating of such vital responses in the neuronal networks.
SUMMARY AND GENERAL CONCLUSION
The important findings of this study are as follows: (1) E2 increases neuronal activity in the SCN. (2) The steroid facilitates release of excitatory neurotransmitter glutamate probably via a presynaptic mechanism of action. (3) These effects of E2 might be attributed to its effect on neuronal membrane potentials. The excitatory effect reported here is in good agreement with the in vitro effects observed in earlier electrophysiological investigations which include prolongation of the excitatory postsynaptic potentials and increased incidence of repetitive firing in response to synaptic stimulation in the striatum radiatum (Wong and Moss, 1992; Woolley et al, 1997). Although it is now well accepted that the SCN play a significant role in controlling mood and behavior, further in vivo and in vitro investigations are required to confirm a robust correlation between increased excitability of the SCN neurons induced by E2 and its clinical use before reaching a positive conclusion regarding this matter.
References
Bos NP, Mirmiran M (1990). Circadian rhythms in spontaneous neuronal discharges of the cultured suprachiasmatic nucleus. Brain Res 511: 158–162.
Bouskila Y, Dudek FE (1995). A rapidly activating type of outward rectifier K+ current and A-current in rat suprachiasmatic nucleus neurones. J Physiol 488: 339–350.
Brinton RD (2005). Investigative models for determining hormone therapy-induced outcomes in brain: evidence in support of a healthy cell bias of estrogen action. Ann N Y Acad Sci 1052: 57–74.
Bryant DN, Bosch MA, Rønnekleiv OK, Dorsa DM (2005). 17β-Estradiol rapidly enhances extracellular signal-regulated kinase 2 phosphorylation in the rat brain. Neuroscience 133: 343–352.
Card JP, Moore RY (1991). The organization of visual circuits influencing the circadian activity of the suprachiasmatic nucleus. In: Klein DC, Moore RY, Reppert SM (eds). Suprachiasmatic Nucleus: the Mind's Clock. Oxford University Press: Oxford. pp 51–76.
Carrer HF, Araque A, Buno W (2003). Estradiol regulates the slow Ca2+-activated K+ current in hippocampal pyramidal neurons. J Neurosci 23: 6338–6344.
Castel M, Belenky M, Cohen OP, Storm-Mathisen J (1993). Glutamate-like immunoreactivity in retinal terminals of the mouse suprachiasmatic nucleus. Eur J Neurosci 5: 368–381.
Chiang HT, Wu SN (2001). Inhibition of large-conductance calcium-activated potassium channel by 2-methoxyestradiol in cultured vascular endothelial (HUV-EC-C) cells. J Membr Biol 182: 203–212.
Clark KA, Randall AD, Collingridge GL (1994). A comparison of paired-pulsed facilitation of AMPA and NMDA receptor-mediated excitatory postsynaptic currents in the hippocampus. Exp Brain Res 101: 272–278.
Cloues RK, Sather WA (2003). Afterhyperpolarization regulates firing rate in neurons of the suprachiasmatic nucleus. J Neurosci 23: 1593–1604.
Creager R, Dunwiddie T, Lynch G (1980). Paired-pulse and frequency facilitation in the CA1 region of the in vitro rat hippocampus. J Physiol 299: 409–424.
Davis PJ, Tillmann HC, Davis FB, Wehling M (2002). Comparison of the mechanisms of nongenomic actions of thyroid hormone and steroid hormones. J Endocrinol Invest 25: 377–388.
De Jeu M, Geurtsen A, Pennartz C (2002). A Ba2+-sensitive K+ current contributes to the resting membrane potential of neurons in rat suprachismatic nucleus. J Neurophysiol 88: 869–878.
De Jeu M, Hermes M, Pennartz C (1998). Circadian modulation of membrane properties in slices of rat suprachiasmatic nucleus. NeuroReport 9: 3725–3729.
Falkenstein E, Wehling M (2000). Nongenomically initiated steroid actions. Eur J Clin Invest 30: 51–54.
Fatehi M, Kombian SB, Saleh TM (2005). 17β-Estradiol inhibits outward potassium currents recorded in rat parabrachial nucleus cells in vitro. Neuroscience 135: 1075–1086.
Fatehi M, Zidichouski J, Kombian SB, Saleh TM (2006). 17β-Estradiol attenuates excitatory neurotransmission and enhances the excitability of rat parabrachial neurons in vitro. J Neurosci Res 84: 666–674.
Glatzer NR, Smith BN (2005). Modulation of synaptic transmission in the rat nucleus of the solitary tract by endomorphin-1. J Neurophysiol 93: 2530–2540.
Gompf HS, Allen CN (2004). GABAergic synapses of the suprachiasmatic nucleus exhibit a diurnal rhythm of short-term synaptic plasticity. Eur J Neurosci 19: 2791–2798.
Green DJ, Gillette R (1982). Circadian rhythm of firing rate recorded from single cells in the rat suprachiasmatic brain slice. Brain Res 245: 198–200.
Groos G, Hendriks J (1982). Circadian rhythms in electrical discharge of rat suprachiasmatic neurons recorded in vitro. Neurosci Lett 34: 283–288.
Inouye ST, Kawamura H (1979). Persistence of circadian rhythmicity in a mammalian hypothalamic ‘island’ containing the suprachiasmatic nucleus. Proc Natl Acad Sci USA 76: 5962–5966.
Itri JN, Michel S, Vansteensel MJ, Meijer JH, Colwell CS (2005). Fast delayed rectifier potassium current is required for circadian neuronal activity. Nat Neurosci 8: 650–656.
Jackson AC, Yao GL, Bean BP (2004). Mechanism of spontaneous firing in dorsomedial suprachiasmatic nucleus neurons. J Neurosci 24: 79–85 -7998.
Jiang ZG, Yang Y, Liu ZP, Allen CN (1997). Membrane properties and synaptic inputs of suprachiasmatic nucleus neurons in rat brain slices. J Physiol 499: 141–159.
Kelly MJ, Qiu J, Ronnekleiv OK (2003). Estrogen modulation of G-protein-coupled receptor activation of potassium channels in the central nervous system. Ann N Y Acad Sci 1007: 6–16.
Kelly MJ, Qiu J, Wagner EJ, Ronnekleiv OK (2002a). Rapid effects of estrogen on G protein-coupled receptor activation of potassium channels in the central nervous system. J Steroid Biochem Mol Biol 83: 187–193.
Kelly MJ, Ronnekleiv OK, Ibrahim N, Lagrange AH, Wagner EJ (2002b). Estrogen modulation of K(+) channel activity in hypothalamic neurons involved in the control of the reproductive axis. Steroids 67: 447–456.
Kim YI, Dudek FE (1991). Intracellular electrophysiological study of suprachiasmatic nucleus neurons in rodents: excitatory synaptic mechanisms. J Physiol 444: 269–287.
Kononenko NI, Dudek FE (2004). Mechanism of irregular firing of suprachiasmatic nucleus neurons in rat hypothalamic slices. J Neurophysiol 91: 267–273.
Kow LM, Devidze N, Pataky S, Shibuya I, Pfaff DW (2006). Acute estradiol application increases inward and decreases outward whole-cell currents of neurons in rat hypothalamic ventromedial nucleus. Brain Res 1116: 1–11.
Kow LM, Pfaff DW (1984). Suprachiasmatic neurons in tissue slices from ovariectomized rats: electrophysiological and neuropharmacological characterization and the effects of estrogen treatment. Brain Res 297: 275–286.
Kruijver FP, Swaab DF (2002). Sex hormone receptors are present in the human suprachiasmatic nucleus. Neuroendocrinology 75: 296–305.
Kuhlman SJ, McMahon DG (2004). Rhythmic regulation of membrane potential and potassium current persists in SCN neurons in the absence of environmental input. Eur J Neurosci 20: 1113–1117.
Kumar A, Foster TC (2002). 17beta-estradiol benzoate decreases the AHP amplitude in CA1 pyramidal neurons. J Neurophysiol 88: 621–626.
Lee SJ, McEwen BS (2001). Neurotrophic and neuroprotective actions of estradiols and their therapeutic implications. Annu Rev Pharmacol Toxicol 41: 569–591.
Li Z, Hay M (2000). 17-beta-estradiol modulation of area postrema potassium currents. J Neurophysiol 84: 1385–1391.
Lundkvist GB, Kristensson K, Hill RH (2002). The suprachiasmatic nucleus exhibits diurnal variations in spontaneous excitatory postsynaptic activity. J Biol Rhythms 17: 40–51.
Meredith A, Wiler SW, Miller BH, Takahashi JS, Fodor AA, Ruby NF et al (2006). BK calcium-activated potassium channels regulate circadian behavioral rhythms and pacemaker output. Nat Neurosci 9: 1041–1049.
Michel S, Itri J, Colwell CS (2002). Excitatory mechanisms in the suprachiasmatic nucleus: the role of AMPA/KA glutamate receptors. J Neurophysiol 88: 817–828.
Morin LP (1994). The circadian visual system. Brain Res Rev 19: 102–127.
Perrin JS, Segall LA, Harbour VL, Woodside B, Amir S (2006). The expression of the clock protein PER2 in the limbic forebrain is modulated by the estrous cycle. Proc Natl Acad Sci USA 103: 5591–5596.
Pierson PM, Liu X, Raggenbass M (2005). Suppression of potassium channels elicits calcium-dependent plateau potentials in suprachiasmatic neurons of the rat. Brain Res 1036: 50–59.
Pitts GR, Ohta H, McMahon DG (2006). Daily rhythmicity of large-conductance Ca2+-activated K+ currents in suprachiasmatic nucleus neurons. Brain Res 1071: 54–62.
Qiu J, Bosch MA, Jamali K, Xue C, Kelly M, Rønnekleiv OK (2006). Estrogen upregulates T-type calcium channels in the hypothalamus and pituitary. J Neurosci 26: 11072–11082.
Reynolds T, Pinnock RD (1997). Neuromedin C decreases potassium conductance and increases a non-specific conductance in rat suprachiasmatic neurones in brain slices in vitro. Brain Res 750: 67–80.
Rudick CN, Woolley CS (2003). Selective estradiol receptor modulators regulate phasic activation of hippocampal CA1 pyramidal cells by estradiol. Endocrinology 144: 179–187.
Sah P (1996). Ca2+-activated K+ currents in neurons: types, physiological roles and modulation. Trends Neurosci 19: 150–154.
Shirakawa T, Moore RY (1994). Glutamate shifts the phase of the circadian neuronal firing rhythm in the rat suprachiasmatic nucleus in vitro. Neurosci Lett 178: 47–50.
Shiroma S, Yamaguchi T, Kometani K (2005). Effects of 17β-estradiol on chemically induced long-term depression. Neuropharmacology 49: 97–102.
Simoncini T, Genazzani AR (2003). Non-genomic actions of sex steroid hormones. Eur J Endocrinol 148: 281–292.
Su JD, Qiu J, Zhong YP, Chen YZ (2001). Expression of estrogen receptor -alpha and -beta immunoreactivity in the cultured neonatal suprachiasmatic nucleus: with special attention to GABAergic neurons. Neuroreport 12: 1955–1959.
Thomson AM, West DC, Vlachonikolis IG (1984). Regular firing patterns of suprachiasmatic neurons maintaind in vitro. Neurosci Lett 52: 329–334.
Van Esseveldt KE, Lehman MN, Boer GI (2000). The suprachiasmatic nucleus and the circadian time-keeping system revised. Brain Res Rev 33: 34–77.
Vasudevan N, Kow LM, Pfaff D (2005). Integration of steroid hormone initiated membrane action to genomic function in the brain. Steroids 70: 388–396.
Wang Y-C, Huang R-C (2006). Effects of sodium pump activity on spontaneous firing in neurons of the rat suprachiasmatic nucleus. J Neurophysiol 96: 109–118.
Womble MD, Andrew JA, Crook JJ (2002). 17β-Estradiol reduces excitatory postsynaptic potential (EPSP) amplitude in rat basolateral amygdala neurons. Neurosci Lett 331: 83–86.
Wong M, Moss RL (1992). Long-term and short-term electrophysiological effects of estradiol on the synaptic properties of hippocampal CA1 neurons. J Neurosci 12: 3217–3225.
Woolley CS (1999). Electrophysiological and cellular effects of estradiol on neuronal function. Crit Rev Neurobiol 13: 1–20.
Woolley CS (2007). Acute effects of estrogen on neuronal physiology. Annu Rev Pharmacol Toxicol 47: 657–680.
Woolley CS, Dlugosch DJ, Schwartzkroin PA (1997). Estradiol increases the sensitivity of hippocampal CA1 pyramidal cells to NMDA receptor-mediated synaptic input. J Neurosci 11: 1848–1859.
Xue B, Hay M (2003). 17β-Estradiol inhibits excitatory amino acid-induced activity of neurons of the nucleus tractus solitarius. Brain Res 976: 41–52.
Zacharieva S, Kirilov G, Kalinov K, Shiqarminova R, Nachev E, Orbetzova M et al (2002). Effect of different hormone replacement therapy regimens on circadian blood pressure profile and active renin in postmenopausal women. Gynecol Endocrinol 16: 461–467.
Zucker I, Fitzgerald KM, Morin LP (1980). Sex differentiation of t-e circadian system in the golden hamster. Am J Physiol 238: R97–R101.
Acknowledgements
We are grateful to the faculties and staff in the Department of Biomedical Sciences at the University of Prince Edward Island, Canada, for their support and providing the opportunity to carry out this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
DISCLOSURE/CONFLICT OF INTEREST
There is no potential conflict of interest among the authors.
Rights and permissions
About this article
Cite this article
Fatehi, M., Fatehi-Hassanabad, Z. Effects of 17β-Estradiol on Neuronal Cell Excitability and Neurotransmission in the Suprachiasmatic Nucleus of Rat. Neuropsychopharmacol 33, 1354–1364 (2008). https://doi.org/10.1038/sj.npp.1301523
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.npp.1301523
Keywords
This article is cited by
-
Impact of sleep patterns upon female neuroendocrinology and reproductive outcomes: a comprehensive review
Reproductive Biology and Endocrinology (2022)
-
Effects of estrogen and progesterone on the neurogenic inflammatory neuropeptides: implications for gender differences in migraine
Experimental Brain Research (2020)
-
L-Type Calcium Channels Modulation by Estradiol
Molecular Neurobiology (2017)
