
Abstract
Rice Tungro disease poses a threat to rice production in Asia. Marker assisted backcross breeding is the most feasible approach to address the tungro disease. We targeted to introgress tungro resistance locus tsv1 from Matatag 1 into a popular but tungro susceptible rice variety of Bangladesh, BRRI dhan71. The tsv1 locus was traced using two tightly linked markers RM336 and RM21801, and background genotyping was carried out using 7 K SNPs. A series of three back crosses followed by selfing resulted in identification of plants similar to BRRI dhan71. The background recovery varied at 91–95% with most of the lines having 95%. The disease screening of the lines showed moderate to high level of tungro resistance with a disease index score of ≤ 5. Introgression Lines (ILs) had medium slender grain type, and head rice recovery (59.2%), amylose content (20.1%), gel consistency (40.1 mm) and gelatinization temperature were within the acceptable range. AMMI and Kang’s stability analysis based on multi-location data revealed that multiple selected ILs outperformed BRRI dhan71 across the locations. IR144480-2-2-5, IR144483-1-2-4, IR144484-1-2-2 and IR144484-1-2-5 are the most promising lines. These lines will be further evaluated and nominated for varietal testing in Bangladesh.
Similar content being viewed by others


Introduction
Rice provides daily calories and nutrition to the majority of the Asian population, and it is becoming a popular diet in Africa and Latin America as well1. The demand for rice is increasing globally, so there is a dire need to increase rice production and productivity. As per some estimates, we need to produce additional 10 million tons of rice every year to feed the increasing population2,3,4,5,6. However, rice production is constrained by several major diseases such as bacterial blight, blast, sheath blight, and rice tungro7,8,9. Among the more than 17 viral diseases of rice, rice tungro (RT) incidence and severity are high in Asia10. Annually 10% ($1.5 billion) of rice yield losses are attributed to RT, and there have been several incidences of complete crop failures in South and South East Asian countries11,12,13.
RT disease is caused by two viral particles, namely rice tungro spherical virus (RTSV, an RNA virus) and rice tungro bacilliform virus (RTBV, a DNA virus). Both are easily transmitted by the green leaf hopper (GLH; Nephotettix virescens (Distant)), while the transfer of the latter is RTSV dependent12,14,15. It has been reported that RTBV causes the tungro disease and RTSV intensifies the disease, hence most of the research focuses on RTSV resistance16. RT virus infection of rice plants results in stunted growth, leaf discoloration, delayed flowering and maturity, reduction in reproductive tillers, incomplete panicle exertion, improper grain filling and sterility leading to significant yield losses13,17. There have been multiple tungro mitigating measures such as use of chemicals, cultural and agronomic management practices, integrated tungro management approaches along with the use of resistant rice varieties12,18. However, the use of RT resistant rice varieties and the adoption of good agronomic practices are cost-effective, sustainable and environmentally safer18.
In the past, breeding for RT resistance was hampered mainly due to the confusion of whether to breed for resistance against vector carrier GLH or the RT virus8,18,19. But it has been well proven that breeding for RTSV resistance and deployment of RT resistant rice varieties is a practical approach to tackle RT disease in the farmers’ fields17,18,19,20,21. With the availability of high throughput RT virus screening phenotypic protocols coupled with enzyme linked immunosorbent assay (ELISA), it is possible to accurately identify resistance sources and to breed for tungro resistant rice varieties22. RT disease screening of several thousands of rice germplasm including gene bank accessions, breeding lines and released varieties, resulted in identification of many resistant donors for RTSV and only a few donors tolerant to RTBV13,23,24,25,26,27,28. Some of the prominent donors are Utri Merah, Utri Rajapan, Vikramarya ARC11554, IRGC16680, IRGC166682, Kataribhog, Pankhari 203 and many more. These accessions have been widely used in the genetic analyses and breeding for RT resistance in rice.
Identifying QTLs/genes for RT resistance helps in their marker-assisted introgression into popular rice varieties8,13,28,29,30,31. A dominant locus was identified on chromosome 4 that provided resistance to GLH and RTSV, which was later independently validated and fine mapped in a NSICRc138 and ARC138-4-5-5-2-30 derived back cross population19,32. While, several studies reported recessive genes contributing to RTSV resistance29,33,34. Similarly, QTLs were detected on chromosomes 1, 2 and 7 derived from Utri Rajapan and Vikramarya, each of these QTLs explained more than 16% phenotypic variance 35. A major effect loci “tsv1” for RT resistance was mapped between 22.05 and 22.25 Mb on the long arm of chromosome 7 using Utri Merah backcross derived population8,21. A putative translation initiation factor 4G (eIF4Gtsv1) was found to be associated with tsv1 for RTSV resistance, finally, an allele specific SNP was designed for use in MABB8,36. Recently, novel alleles of rice eIF4G were generated by CRISPR/Cas9 genome editing that conferred resistance to RTSV37. Some successful attempts were also made to develop RTBV resistance in the background of Pusa Basmati 1 through genetic engineering20,37,38.
Studies have clearly shown that most semi-dwarf green revolution rice varieties are susceptible to RT disease39. One of the classic examples is that the popular rice mega variety IR64, despite its wider adaptability, high yield potential, good grain quality traits and moderate to high level of resistance to most of the diseases, is highly susceptible to tungro disease8,27,40,41. In a phenotypic and molecular characterization of a set of rice lines along with commercially released rice varieties in Bangladesh, most rice varieties such as BR11, BRRI dhan39, BRRI dhan48, BRRI dhan49 and BRRI dhan71 were susceptible to RT disease13. Therefore, increasing rice crop resilience to sustain rice production is an important research priority42. Introgression or pyramiding of resistance genes into susceptible rice varieties using tightly linked diagnostic markers through marker assisted back cross breeding (MABB) is a fast track approach to control major diseases43,44,45,46. Even though QTLs/genes have been identified for RT resistance, there are not many efforts to systematically introgress them into popular rice varieties. For the first time, tsv1 gene has been successfully introgressed into a japonica rice variety through MABB21.
Here we report the introgression of tsv1 gene into BRRI dhan71, a popular drought tolerant rice variety of Bangladesh mainly grown during Aman season (wet season), it is widely grown in the northern and central parts of Bangladesh, where the farmers used to cultivate another crop between Aman and Boro seasons. It has a yield potential of 3.0–3.5 t/ha under drought and 4.0–5.0 t/ha under irrigated conditions47. But this variety has become highly susceptible to RT disease13,48.
The main objectives of our study were to conduct marker assisted introgression of tsv1 into a popular rice variety of Bangladesh, BRRI dhan71; to characterize introgressed lines for agronomic traits, yield and RT resistance; to identify introgressed lines which were genotypically and phenotypically similar to recipient parent, and to conduct multi-location trials to identify superior lines for varietal nomination in Bangladesh.
Results
Selection of parental materials and tsv1 linked SSR markers
The popular drought tolerant but RT susceptible rice variety BRRI dhan71 from Bangladesh was used as a recipient parent, for which the source of pure seeds came from Bangladesh Rice Research Institute (BRRI), Gazipur, Bangladesh. The well characterized RT resistant Matatag 1 was used as a donor parent. Both the parents grew well with good agronomic performance and yield potential at International Rice Research Institute (IRRI), Los Banos, Philippines. They are medium duration, BRRI dhan71 and Matatag 1 flowered 85 and 96 days after seeding, respectively, which helped in coinciding the flowering for making crosses. The polymorphism survey between the parents using eight simple sequence repeats (SSR) markers tightly linked to tsv1 gene viz; RM336, RM21797, RM21800, RM6152, RM21801, RM21808, RM21822 and RM455, was carried out, of which two SSR markers RM336 and RM21801 were found to be polymorphic. They are located between 21.87 and 22.05 Mb region, RM336 is 180 kb away from tsv1, whereas, RM21801 marker is very tightly linked. These two markers were utilized for introgression of RT resistance gene into BRRI dhan71.
Development of tsv1 introgression lines through MABB
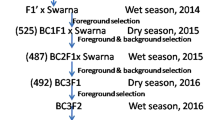
A cross was made using BRRI dhan71 as a female and Matatag 1 as a male parent. Both the parents were cross compatible and successfully produced F1s. The number of seeds produced, genotyped, true F1s and number of plants selected after each cross are provided in Table 1. We genotyped 157 out of the 223 F1 seeds produced, of which 83 were found to be true F1s when tested using SSR markers RM336 and RM21801. Eighteen positive plants were selected and backcrossed to BRRI dhan71 to produce BC1. In each of the backcross generations number of F1s produced and genotyped varied from 136 to 222 and 83–175, respectively, whereas the number of true F1s and those selected for crosses ranged from 43 to 98 and 14–27, respectively. From the 98 BC3F1 plants, 27 plants were selected similar to the recipient parent for plant type and grain type, and selfed them to produce BC3F2 plants. We pooled 10–15 seeds from each of the 27 plants to generate BC3F2 plants. In all 182 BC3F2 plants were genotyped out of which 37 were found to be homozygous for tsv1 gene (Fig. 1). The DF and PH varied 75–98 days and 105–137 cm, respectively. Twenty three plants were selected based on agro-morphological characters resembling the recurrent parent and advanced them to obtain BC3F3 families. The markers segregation analysis was carried out and goodness of fit was applied for each of the back cross and BC3F2 selfing generation. In each of the back cross generations the segregation of markers followed 1:1 and in BC3F2 it was 1:2:1 (Table 2). Both the markers showed clear Mendelian inheritance without any segregation distortion. Overall, three successive MABB followed by selfing resulted in the identification of plants similar to the recurrent parent (Fig. 2).

Gel picture showing RM336 maker segregation in BC3F2 generation. L—Ladder, P1—Susceptible (BRRI dhan71) and P2—Resistant (Matatag 1).

Introgression line with tsv1 gene and recipient parent BRRI dhan71.
Phenotypic and molecular characterization of introgression lines at IRRI-HQ
Agronomic, yield and yield related traits
Twenty three (BC3F3) and 50 ILs (BC3F4) were evaluated in replicated trials during 2021 dry season (DS) and 2021 wet season (WS), respectively. Among the ILs, range and means of DF, PN, PL and YLD were higher in DS, while PH was high in WS. Low to moderate coefficient of variations (CVs) was observed for all the traits during both seasons. There were significant genotypic differences for DF and YLD during both seasons, while PH has significant difference in WS but not in DS. PL has no significant genotypic difference in both seasons, whereas significant genotypic differences were observed for disease resistance reactions. For all the traits, moderate to high heritability (> 53%) were observed in both seasons (Table 3). The DF of BRRI dhan71 was 85 days and Matatag 1 had 96 days. BRRI dhan71 average PH was 121 cm. whereas; PL of BRRI dhan71 exhibited 27 cm with no significant difference. BRRI dhan71 exhibited an average yield of 4.2 ton/ha. Transgressive segregants were also observed for most of the traits. Overall, 62% of the ILs flowered earlier than BRRI dhan71; 14% and 50% of the ILs had longer panicle length and higher yield respectively, than BRRI dhan71. Among the agronomic traits, a significant positive correlation was observed between DF and PH (0.583), PH and PL (0.375), and a significant negative correlation was observed between GL and YLD (-0.292) (Fig. 3).

Correlation among agronomic, yield and grain quality related traits. DF—days to flowering (days), PH—plant height (cm), PN—panicle number, PL—panicle length (cm), YLD—yield (t/ha), BR-brown rice (%), MR-milled rice (%), HR—head rice recovery (%), GL—grain length (mm), GW-grain width (mm), L/B—length and breadth ratio, AC—amylose content (%), GC—gel consistency (mm).
Grain quality assessment
Both the parents were similar for most of the grain quality traits except for HR and with slight differences in GL and LB ratio. Matatag 1 is a long slender grain type with LB ratio of 3 and BRRI dhan71 is a medium slender type with LB ratio of 2.5 (Table 4). Most of the ILs had medium slender grain type with a LB ratio of 2.63 and they were similar to BRRI dhan71. All the ILs, including the parents showed intermediate GT. The BR, MR and HR ranged between 73 and 80%, 64–73% and 58–65%, respectively, whereas BRRI dhan71 exhibited BR, MR and HR with 76%, 68% and 61%, respectively. The majority of ILs, showed 75%, 67% and 60% of BR, MR and HR, respectively. Low to moderate CVs were observed for all the traits among the ILs, and there were significant genotypic differences for GL and LB ratio. Among the grain quality traits significant positive correlations were observed between LB and GL with 0.797 followed by HR and MR (0.515), GC and AC (0.411), and GC and GW (0.300). A significant negative correlation was observed between GL and BR with − 0.496, followed by LB and BR (− 0.442), LB and GW (− 0.374), GC and LB (− 0.364), GL and MR (− 0.344) and GL and YLD (− 0.292) (Fig. 3).
RT resistance screening
The RT disease reactions of the selected 50 ILs evaluated at IRRI-HQ is provided in the Table S1. As expected the donor parent Matatag 1 and the RT resistant check TW16 showed resistant reaction with a disease index (DI) score of 3, while the recipient parent and susceptible check TN1 showed susceptible reaction with DI score of 6 and 7. All the ILs performed better than recurrent parent BRRI dhan71 with a DI of 3–4 (Table 3 and Table S1). Seven ILs viz; IR 144469-1-1-1, IR 144469-1-1-6, IR 144481-1-2-3, IR 144483-1-2-6, IR 144485-1-1-1, IR 144485-1-1-3 and IR 144491-1-2-1 showed resistant reaction with DI of 3. All the selected ILs were healthy without any significant plant height reduction or yellowing of leaves even after 21 days of RT virus infestation (Fig. 4).

Plants are exhibiting symptoms of RT disease after inoculation. ILs are exhibiting resistant reaction against RT disease.
Background recovery
Fifty ILs were subjected for 7 K SNP chip based genotyping, in all 7098 SNPs were successfully called. After filtering out missing (834), nonsense (195), heterozygous (14) and monomorphic (4994) SNPs, finally, 1061 high-quality polymorphic SNPs were extracted with an average marker density of one SNP per 300 kb. The polymorphic SNPs were distributed all over the genome. The number of polymorphic SNPs ranged from 57 on chromosome 9 to 148 on chromosome 1, with more than 100 markers on chromosomes 2, 3 and 4. The lowest numbers of donor allele introgressions were observed on chromosomes 4, 10 and 12 and highest on chromosome 7, on all other chromosomes, introgressions varied from 2.9 to 6.8%. The genetic background recovery of the 50 ILs showing alleles similar to the recipient parent varied from 91 to 95%. Overall in 38 ILs, background recovery was more than 95%. The graphical genotyping of the Recurrent Parent Genome (RPG) recovery of selected ILs showing the position of tsv1 gene is provided in Fig. 5.

RPG recovery estimation of selected ILs.
Multi-location evaluation of introgression lines in Bangladesh
Agronomic performance of selected 50 ILs and their respective parents BRRI dhan71 and Matatag 1 were assessed at three RT disease hot spot locations during Aman season (WS 2021) in Bangladesh. We collected data for DF, PH, PN, PL and YLD in all three locations. There was no variation observed for DF among the lines and BRRI dhan71 across the locations, so it was excluded from the analyses. Results of individual location and combined analyses are provided in Table 5 and Table S2.
In the individual location analysis there were significant genotypic differences for PH and YLD among ILs in all the locations, while PL and PN among ILs and with BRRI dhan71 were almost similar. The mean PH (98 cm) was lowest at HG and the highest mean YLD (5.1 t/ha) was observed at GP. Low CVs were observed for PH (3.9–6.8%) and PL (5.3–6.7%), while moderate to high CVs were observed for PN (16.6–20.2%) and YLD (17.2–22.5%). Moderate to high heritability (> 45%) was observed for all the traits (Table 5).
The AMMI analyses of variance showed significant genotypic effects for PH and YLD, environmental effects for PN and YLD, and significant genotype and environmental (G × E) interactions for PH and YLD. While among the first two PCs, PC1 was significant for all the traits but PC2 was significant for PN and YLD only (Table S2). AMMI biplot analysis showed 76.5% goodness of fit for YLD (Table S2 and Fig. 6) and revealed that the average yield was 5.2 t/ha. The two ILs G1 and G2, yielded higher than BRRI dhan71 with high main additive effects (Fig. 6), whereas the G3 and G6 yielded on par with BRRI dhan71 (G12). G3 and G6 are relatively close to PC1 zero score line, thus have wider adaptation to the test environments. The statistical analysis revealed some of the ILs viz., IR144480-2-2-5 (G1), IR144483-1-2-4 (G2), IR144484-1-2-2 (G3), IR144484-1-2-5 (G4), IR144485-1-1-6 (G5), IR144488-2-1-4 (G6), IR144489-1-1-1 (G7), IR144491-1-1-1 (G8), IR144491-1-2-5 (G9) and IR144491-1-2-6 (G10) performed stably in terms of yield across the three locations in Bangladesh (Table 6).

Adaption map and AMMI biplot. IR144480-2-2-5 (G1), IR144483-1-2-4 (G2), IR144484-1-2-2 (G3), IR144484-1-2-5 (G4), IR144485-1-1-6 (G5), IR144488-2-1-4 (G6), IR144489-1-1-1 (G7), IR144491-1-1-1 (G8), IR144491-1-2-5 (G9), IR144491-1-2-6 (G10), Matatag 1 (G11), BRRI dhan71 (G12), CL (E1), GP (E2) and HG (E3).
Stability analysis was performed for all the traits opting Kang’s stability model. The positive Ysi ranking indicates the stable performance of superior genotypes for different traits across the locations. Among the all, 50% of the ILs exhibited stable performance in at least one key yield trait for PN, PL and YLD and, 48% ILs showed stability for PH in all three locations (Table S3). The ILs IR144483-1-2-3, IR144485-1-1-1, IR144491-1-1-1, IR144491-1-2-3 and IR144492-1-3-2 were superior for PH, PN, PL and YLD traits; IR144480-2-2-5, IR144481-1-2-4, IR144485-1-3-1, IR144488-2-1-2 and IR144492-1-2-6 for PH, PN and YLD traits; and IR144482-1-1-1, IR144485-1-1-3, IR144491-1-2-1 and IR144491-1-2-5 for PH, PL and YLD traits. The Sum of YSi rankings indicates the stable performance of ILs across the locations. IR144491-1-1-1 was identified to have the highest YSi ranking (190) followed by IR144491-1-2-3 (167) and IR144485-1-1-1 (161), while the lowest YSi ranking observed in IR144469-1-1-6 (27) followed by IR144471-2-3-6 (29) and IR144469-1-1-1 (47). Whereas, the selected ILs viz., G1 (136), G2 (132), G3 (119), G4 (123), G5 (140), G6 (145), G7 (119), G8 (190), G9 (143) and G10 (121) exhibited moderate to high YSi ranking with mean yield of 4.33 t/ha across the three locations (Table S3).
Identification of promising introgression lines
Based on the overall performance of ILs for yield and other traits, with good grain quality traits and resistance to RTV, four best promising ILs were identified, namely: IR144480-2-2-5 (G1), IR144483-1-2-4 (G2), IR144484-1-2-2 (G3) and IR144488-2-1-4 (G6). Three of the four lines had genetic background recovery of 95%. The agronomic and yield parameters were better or at par with the recipient parent BRRI dhan71 (Table 6). All these ILs had medium slender grain type with intermediate AC, GT and GC, and HR, BR and MR were almost similar to the BRRI dhan71 indicating their suitability for Bangladesh consumer preferences (Table S4). However, the whole set of 50 ILs are being further evaluated over the same three locations to identify stable lines.
Discussion
Rice production continuously needs to be improved to sustain food security4,49. The increasing biotic stress pressure including the high incidences of RT disease on rice production is a major concern in Asia37,50. The RT is becoming epidemic in Bangladesh and most of the popular rice varieties released in Bangladesh have become susceptible to the disease. Studies have shown 20–100% yield losses, with none of the varieties tested showing complete resistance in the RT disease hotspot location at Cumilla, Bangladesh13,39. Hence, there is an urgent need to incorporate RT resistance into popular rice varieties through MABB or breed newer rice varieties with RT resistance13,18. The current study targeted to improve tungro resistance of a popular rice variety BRRI dhan71 through marker-assisted introgression of RT resistant gene “tsv1”.
Marker assisted introgression of tungro resistance locus tsv1
Marker-assisted introgression of disease resistance genes using tightly linked markers is a fast-track approach to develop disease resistant varieties21,49. The foreground and background selections using molecular markers improve the precision and efficiency of the breeding programs to select desirable plants51. The tsv1 is a recessive locus conferring resistance to RTSV, so it is more practical and appropriate to use MABB to develop RT resistant rice varieties21. It has been reported that RM336 is linked to tsv1 and can be used in MABB8,21. However, this marker is located 180 kb away from the gene and showed a low-level recombination of 1.7% with the gene8. Polymorphism of eight SSR markers around the tsv1 locus was surveyed between the parents BRRI dhan71 and Matatag 1, and polymorphic markers RM21801 and RM336 were used in our introgression work. The RM21801 is located close to tsv1 and showed precise co-segregation with the gene. Interestingly, the study also observed over the different generations 3–4% recombination frequency between RM336 and RM21801 or tsv1. Hence both the marker loci to select positive plants in every generation were used to ensure the introgression of tsv1. It is important to note that an elongation initiation factor (eIFGtsv1) has been identified as a candidate gene and allele specific SNPs have been identified for tsv136. With the availability of 3 K genome sequence better haplotypes can be searched for tsv152. Use of superior haplotypes and tightly linked gene specific markers can further improve the efficiency of MABB53.
As expected, each of the successive backcrosses followed by selfing resulted in the production of plants similar to the recipient parent BRRI dhan71. The seed set, seed germination, and plant growth were normal and markers segregation followed Mendelian inheritance in each generation. These results clearly showed that the tsv1 gene was introgressed and the recipient parent phenotype was successfully recovered without any segregation distortion (Table 2). Similar observations were also made in two populations derived from TW6 showing Mendelian segregation for tsv1 by phenotypic segregation and genotypic segregation based on the progeny test8. Segregation distortion and linkage drag associated with the genes/QTLs derived from distant species of cultivated rice and wild relatives are a major hindrance for their use in breeding54,55. The main advantage of backcross is to retain the most important traits of the recipient varieties, it has been established that 3–4 backcrosses are enough to recover most of the recipient parent genome and thereby most of its traits43,44,49. RPG recovery was tested using genome-wide SNPs only in selected and fixed BC3F4 lines. The genetic RPG recovery varied from 91 to 95%, 38 (76%) out of the 50 ILs having 95% as per the theoretical expectation43,56,57. The stringent selection of progenies resembling recipient parent for agronomic and grain type traits in every back cross generation, and final selection of fixed plants similar to recipient parent resulted in expected genetic background recovery. However, in some ILs more numbers of donor specific alleles were introgressed around the tsv1 gene on chromosome 7 compared to other regions of the genome.
A thorough phenotypic and molecular characterization of a wide range of rice germplasm showed the RT susceptibility of modern rice varieties including rice mega variety IR64, while genetic mapping efforts lead to identification of major loci for RT disease resistance but there are not many efforts to introgress them to popular rice varieties13,23,28,35,41,58. The study is the second report on tsv1 introgression through MAS. In an earlier study tsv1 was successfully combined with photoperiod insensitivity into japonica rice. They were able to identify a set of RT disease resistant breeding lines with high yield and good grain quality traits21. Similarly, a transgene ORFIV introgression into popular rice varieties conferred RT resistance59. However, recognizing the importance of RT resistance for stable rice production in Asia, it has been included as an important component of product profiles, SNPs have also been designed and validated for RT disease resistance and included in 1 K-RiCA assay for routine use in rice breeding60.
Performance of ILs for agronomic, grain quality traits and RT disease resistance
The ILs showed consistently higher mean values and ranges for all the traits except PH during DS than in WS. There were significant genotypic differences for DF and YLD during both seasons, whereas PL has no significant genotypic differences. For all the traits, moderate to high heritability (> 53%) was observed in both seasons (Table 3). These results are in consonance with earlier reports on trait variations, genotypic effects and heritability of ILs for disease resistance, drought and submergence tolerance, and nutritional traits9,21,43,57,61,62. Transgressive ILs with early flowering, longer panicles and higher yield than BRRI dhan71 were observed. Similar observations were also reported on ILs of BB and blast resistance with early flowering, reduced height and increase in number of grains and no changes in panicle length63,64. The appearance of transgressive segregrants in the progenies is due to the accumulation of positive alleles from both the parents65,66. The phenotypic variation present in the ILs provides an opportunity to select superior lines for agronomic and yield related traits along with high level of stress tolerance or nutritional value.
Most of the ILs had medium slender grain type with intermediate AC, GT and GC, while BR, MR and HR were within the acceptable range. It shows that careful selection of lines and MABB resulted in development of ILs with unaltered grain quality parameters and these were similar to the recipient parent BRRI dhan71. Grain quality traits play an important role in consumer acceptance of released rice varieties67.
The RT virus inoculation resulted in stunting and severe yellowing of leaves of susceptible check and BRRI dhan71 (Fig. 4). All the ILs and resistant check showed moderate to high level of resistance compared to BRRI dhan71 (Table 3 and Table S1). All the selected ILs were healthy without any significant plant height reduction or yellowing of leaves even after 21 days of RT virus infestation while susceptible check could not recover. This clearly shows the effectiveness of tsv1 and its successful introgression into BRRI dhan71 through MABB. Efforts are being made for additional controlled screen house based RT disease screening and forced tube inoculation to select the final set of lines. The effectiveness of tsv1 and its successful transfer through MAS and further validation by phenotyping have been previously reported8,21. Similarly, MABB has been used to transfer disease resistance genes of BB, blast, sheath blight, tungro, drought and submergence tolerance etc. in rice, and promising ILs have been successfully released for farmers cultivation21,32,40,43,44,45,46,61,68,69,70.
Tungro resistant ILs with superior agronomic and grain quality traits were identified
Multi-location evaluations of ILs were conducted at RT disease hotspot locations in Bangladesh. Understanding the GxE and identification of stable performing lines is essential for the successful release and adoption of rice varieties71,72,73. Significant genotypic, environment, and G × E effects were observed for YLD. Based on the AMMI and Kang’s stability analyses, ten best performing lines were selected (Table 6). These lines had DF and PH similar to BRRI dhan71, and YLD at par or better than BRRI dhan71. They had moderate to high level of resistance and all the grain quality traits were within the acceptable range. Final set of RT disease resistant lines selected based on Multi-location data will be nominated for varietal testing in Bangladesh.
Conclusion
RT disease is becoming epidemic in several countries of Asia and most of the popular cultivated rice varieties are susceptible to the disease. Breeding for RT disease resistance is an effective approach to tackle the disease. RT disease resistant locus tsv1 was successfully introgressed into a popular but RT susceptible rice variety BRRI dhan71 through MABB. The final set of ILs recovered most of the genetic background of the recipient parent, and showed agronomic, yield and grain quality parameters similar to the BRRI dhan71.
Materials and methods
Plant materials
BRRI dhan71 is a mid-early duration rice variety with promising yields even under drought conditions in Bangladesh but it is highly susceptible to RT disease. Because of its mid-early crop duration farmers are opting for this variety to grow in between Aman (wet) and Boro (dry) season. BRRI dhan71 was imported from Bangladesh Rice Research Institute (BRRI), Gazipur, Bangladesh. Matatag 1 or IR69726-116-1-3 is a Philippine rice variety used as a donor parent for the tungro resistance tsv1 gene, which exhibits moderate resistance against RTSV13,17. The donor parent Matatag 1 is derived from a cross combination of IR52256-84-2-3/IR72//IR1561-228-3*2/Utri Merah (Acc 16682), and seeds were obtained from IRRI gene bank. This study complies with relevant institutional, national, international guidelines and legislation.
Crossing program
A standard MABB program to introgress tsv1 from the donor parent Matatag 1 into BRRI dhan71 was followed. Crosses were made using Matatag 1 as a male parent and BRRI dhan71 was used as a female parent and F1s were produced. The F1s and BCnF1s produced were confirmed using tsv1 gene tightly linked SSR markers RM336 and RM21801. The true F1 plants were backcrossed to the recipient parent three times and selfed to produce BC3F2 plants. The crossing scheme followed to develop BRRI dhan71 RTV resistance lines is provided in Fig. 7. In each of the generations, F1 plants with tsv1 gene and phenotypically resembling recipient parent were selected and used for crosses. Selected BC3F1 was selfed to produce BC3F2 and tsv1 homozygous plants were selected and further advanced through the pedigree method to produce BC3F4 (Fig. 7).

Breeding scheme for developing tungro resistant BRRI dhan71 ILs.
Molecular analysis
DNA isolation and quantification
Young rice leaves (5–7 cm) were collected, lyophilized and each of the samples was grinded to powder in a 2 ml micro tube using GenoGrinder. Genomic DNA was extracted by TPS (a mixture of Tris–HCl, EDTA and KCl) method74. Pre-warmed 700 µl TPS buffer (100 mM Tris–HCl [pH 8.0], 1 M KCl, 10 mM EDTA) was added, well mixed and incubated at 65 °C for 1 h, and then centrifuged. The collected 500 µl supernatant was added with an equal volume of isopropyl alcohol and incubated at 4 °C for 30 min. Pellet was extracted after centrifugation and washed with 70% alcohol. The dried pellet was dissolved in 200 µl of TE buffer (10 mM Tris–HCl [pH 8.0], 1 mM EDTA, RNase A [10 mg/ml]). Quality and quantity of DNA were estimated through 0.8% agarose gel electrophoresis (AGE). We finally adjusted the DNA concentration to 25–50 ng/µl and used in the polymerase chain reaction (PCR). PCR using two SSR markers (RM336 and RM21801) tightly linked to tsv1 gene was carried out 8,13,21. The primers sequences and product sizes are as follows.
RM336:FP-CTTACAGAGAAACGGCATCG; RP-GCTGGTTTGTTTCAGGTTCG; PS:154(bp); RM21801:FP-GCGCACAGCATGTCGAAGTCC; RP-AAACCCGAGGCAAATACGAAACG; PS:191(bp).
Polymerase chain reaction (PCR)
The PCR was performed using a total volume of 10 µl reaction mixture, containing 3 µl of 30 ng of template DNA and 0.5 µl of 5 Pico moles of each primer (Invitrogen), 1 µl of reaction buffer, 1 µl of dNTPs and 0.2 µl of Taq DNA polymerase. The PCR profile used was one cycle of initial denaturation at 95 °C for 5 min, followed by 35 cycles of amplification (each cycle of 1 min denaturation at 95 °C, 30 secs annealing at 55 °C, 30 secs extension at 72 °C), followed by a final extension of 72 °C for 7 min and stored at 4 °C. The amplified products were resolved in 3.5% Agarose gel with 0.5 X TBE buffer containing 0.1 μg/ml of Syber safe (S33102, Invitrogen) and documented in the Gel documentation system21. Gel scoring was carried out based on the donor and recurrent parent banding patterns and their corresponding product sizes. The allele corresponding with Matatag 1 was considered resistant allele. Similarly, allele matched with recipient parent BRRI dhan71 scored as a susceptible allele. Whereas if both the alleles (susceptible and resistant) were observed in one sample, they were considered heterozygotes.
Phenotyping
Agronomic and yield parameters
Field experiments were conducted during 2021DS and 2021WS of 23 and 50 ILs, respectively at Zeigler experimental station (ZES), International Rice Research Institute. Randomized Complete Block Design (RCBD) was followed to lay out the experiments. Standard agronomic practices and plant protection measures to raise a good crop were followed. Data on agronomic and yield traits were collected following the standard evaluation system (SES of IRRI)75. Days to 50% flowering (DF) were calculated based on the sowing date to when 50% of the plants in a plot flowered. Plant Height (PH) was measured from the base of the plant to the tip of the tallest panicle from three selected plants; Panicle length (PL) was measured from the panicle base to the tip of the top most grain. After harvesting grains were well dried and moisture content was adjusted to 14%, grain weight per plot was measured in grams (gm) and converted to tons/ha.
Grain quality parameters
For grain quality (GQ) assessment, 140 g of well dried grains from each of replications were submitted at the Analytical Service Laboratory (ASL), IRRI. The grain morphological, milling and cooking quality data were estimated using standard protocols76. The GQ traits measured were grain length (GL), grain width (GW), brown rice (BR), milled rice (MR), head rice recovery (HR), amylose content (AC), gelatinization temperature (GT) and gel consistency (GC).
Disease screening for RT resistance
Parental lines and ILs for RT resistance were screened by artificial inoculation under controlled conditions. Materials were seeded in a metal tray following Randomized Complete Block Design (RCBD) with three replications under the controlled condition at disease screening facilities of IRRI. The Taichung Native 1 (TN1) and BRRI dhan71 were used as susceptible checks, while TW16 and Matatag 1 were used as resistant checks. Uniformity was maintained between entries in terms of the age of the seedlings and the number (with twenty seedlings) of seedlings per entry by thinning. Ten days old seedlings were subjected to RT virus infestation by releasing viruliferous GLH and allowed them to feed on seedlings. The insects were already given chances to feed on infected plants for 5 days. Seed box that contains the test seedlings were placed inside the water tray and covered with a screen. Viruliferous GLH was released in the cage at an average of 6–7 insects per seedling for 2–3 h of inoculation access. The insects were disturbed every 30 min during the inoculation period to ensure the even distribution among the seedlings. After 21 days of infestation, disease symptoms, incidence and severity were recorded (Fig. 4). Disease severity was evaluated based on a scoring system following SES17,75. Disease index (DI) was estimated based on disease percentage and symptoms. The DI value is determined by the severity of disease symptoms; the higher the DI value, the more severe symptoms75 (Table S5).
DI was estimated as follows
where; n: the number of plants showed the corresponding score; tn: the total number of plants.
Multi-location evaluation of introgression lines
Multi-location yield trials (MLTs) were conducted for selected 50 ILs at IRRI-HQ and at three tungro hot spot locations viz; Gazipur (GP), Cumilla (CL) and Habiganj (HG) in Bangladesh. The ILs along with their respective parents BRRI dhan71 and Matatag 1 were planted in three replications following a RCBD design and evaluated for agronomic traits, yield and disease reactions in the field conditions. Standard agronomic practices were adopted to raise healthier crops in all the field trials. Agronomic replicated data on DF, PH, PN, PL and YLD were collected as per the standard evaluation system of IRRI75.
Statistical analyses
Phenotypic data analyses
All the basic statistical analyses were carried out using PB Tools v1.4 77. The data were curated and means were estimated for each of the replications and used for the analyses. The descriptive statistical parameters such as range, mean, standard error (SE), standard deviation (SD), coefficient of variation (CV) and variance were estimated for each of the trials. Correlation analyses were performed by using R78. From the MLTs data Best Linear Unbiased Prediction (BLUPs) were estimated and used as input files to perform Additive Main-effects and Multiplicative Interaction (AMMI) biplot and genotype main effect analysis. Pairwise t-test of GQ data was performed using Statistical Tool for Agricultural Research (STAR) Version: 2.0.1.
Genotype means were estimated for each trial by using the following mixed model.
here yijk is the performance of the ith genotype in the kth block of the jth replication; μ represents the overall mean; ai represents the effect of ith genotype; r j represents the effect of jth replicate; blj the effect of lth block within the jth replicate; and eijk represents the random error.
Broad-sense heritability (H2) for each trait in each season was calculated as:
where \({\sigma }_{p}^{2}\) is the phenotypic variance, \({\sigma }_{g}^{2}\) is the genotypic variance, \({\sigma }_{e}^{2}\) is the error variance and r is the number of replications in the season.
Stability analysis
The effect of G × E interactions and performance of the genotypes in multi-locations trial in Bangladesh can be explained by stability models like Kang’s stability index. These models will enable to identify stable genotypes.
Genotypic data analyses
The segregation analysis of the tsv1 gene was carried out using the marker segregation data of the RM336 and RM21801. We carried out Chi-square (χ2) analyses for the goodness of fit for each of the data sets from BC generations and BC2F2 using (Excel 2010). The RGP for each of the ILs was estimated based on the number of SNP alleles similar to the recipient parent to that of the total number of alleles amplified and expressed in percentage using Micro soft Excel.
Data availability
The original contributions presented in the study are included in the article/Supplementary Material.
References
Wennberg, A. Food and agriculture organization of the United Nations. Encycl. Toxicol. 3, 628–630 (2014).
Pradhan, P., Fischer, G., van Velthuizen, H., Reusser, D. E. & Kropp, J. P. Closing yield gaps: How sustainable can we be?. PLoS ONE 10, e0129487 (2015).
Patra, B. C., Ray, S., Ngangkham, U. & Mohapatra, T. Rice. In Genetic and Genomic Resources for Grain Cereals Improvement (eds Singh, M. & Upadhyaya, H. D.) 1–80 (Academic Press, 2016). https://doi.org/10.1016/B978-0-12-802000-5.00001-0.
FAO. 2021. Food Outlook – Biannual Report on Global Food Markets. Food Outlook, November 2021. Rome. https://doi.org/10.4060/cb7491en
FAOSTAT. Food and Agriculture Organization of the United Nations: Preliminary Agricultural Production data. http://www.fao.org/economic/est/publications/rice-publications/rice-market-monitor-rmm/en/ (accessed March 20, 2019). (2017).
Tripathy, S. K., Dash, M., Behera, S. K., Ithape, D. M. & Maharana, M. Nutrient rich quality rice—A journey to healthy life. Adv. Plants Agric. Res. 7, 364–367 (2017).
Bunawan, H., Dusik, L., Bunawan, S. N. & Amin, N. M. Rice tungro disease: From identification to disease control. World Appl. Sci. J. 31, 1221–1226 (2014).
Lee, J. H. et al. Single nucleotide polymorphisms in a gene for translation initiation factor (elF4G) of rice (Oryza sativa) associated with resistance to rice tungro spherical virus. Mol. Plant-Microbe Interact. 23, 29–38 (2010).
Ramalingam, J. et al. Gene pyramiding for achieving enhanced resistance to bacterial blight, blast, and sheath blight diseases in rice. Front. Plant Sci. 11, 1–12 (2020).
Hibino, H. Biology and epidemiology of rice viruses. Annu. Rev. Phytopathol. 34, 249–74 (1996)
Warburton, H., Palis, F. L. & Villareal, S. Farmers’ perceptions of rice tungro disease in the Philippines. Pest Manag. Pract. Rice Farmers Asia 129–141 (1997).
Nas, M. & Rae, C. How to overcome rice Tungro virus. Manila Bull. Agric. 2–5 (2011).
Nihad, S. A. I. et al. Genetic variability, heritability, genetic advance and phylogenetic relationship between rice tungro virus resistant and susceptible genotypes revealed by morphological traits and SSR markers. Curr. Plant Biol. 25, 100194 (2021).
Hull, R. Rice tungro bacilliform virus. Encycl. Virol. 65, 1292–1296 (1999).
Dai, S. & Beachy, R. N. Genetic engineering of rice to resist rice tungro disease. Vitr. Cell. Dev. Biol. Plant 45, 517–524 (2009).
Muralidharan, K., Krishnaveni, D., Rajarajeswari, N. V. L. & Prasad, A. S. R. Tungro epidemics and yield losses in paddy fields in India. Curr. Sci. 85, 1143–1147 (2003).
Ladja, F. T., Hidayat, S. H., Damayanti, T. A. & Rauf, A. Responses of Tungro resistant rice varieties and donor parents against five Tungro virus isolates from Indonesia. J. Int. Soc. Southeast Asian Agric. Sci. 22, 18–27 (2016).
Latif, M. A., Rahman, M. M., Ali, M. E., Ashkani, S. & Rafii, Y. Inheritance studies of SSR and ISSR molecular markers and phylogenetic relationship of rice genotypes resistant to tungro virus. C. R. Biol. 336, 125–133 (2012).
Sebastian, L. S. et al. Molecular mapping of resisitance to rice tungro spherical virus and green leafhopper. Phytopathology 86, 25–30 (1996).
Roy, S., Banerjee, A., Tarafdar, J., Senapati, B. K. & Dasgupta, I. Transfer of transgenes for resistance to rice tungro disease into high-yielding rice cultivars through gene-based marker-assisted selection. J. Agric. Sci. 150, 610–618 (2012).
Shim, J. et al. Rice tungro spherical virus resistance into photoperiod-insensitive japonica rice by marker-assisted selection. Breed. Sci. 65, 345–351 (2015).
Henry Sum, M. S., Yee, S. F., Eng, L., Poili, E. & Lamdin, J. Development of an indirect ELISA and dot-blot assay for serological detection of rice tungro disease. Biomed Res. Int. 2017, (2017).
Anjaneyulu, A., Shukla, V. D., Rao, G. M. & Singh, S. K. Experimental host range of rice tungro virus and its vectors. Plant Dis. 66, 54–56 (1982).
Azzam, O. & Chancellor, C. B. The biology, epidemiology, and management of rice tungro disease in Asia. Nat. Resour. Institute, Univ. Greenwich, Chatham Marit. Kent, UK 86, 88–100 (2002).
Hibino, H., Daquioag, R. D., Mesina, E. M. & Aguiero, V. M. Resistances in rice to tungro-associated viruses. Plant Dis. 74, 923 (1990).
Shahjahan, M., Jalani, B. S., Zakri, A. H., Imbe, T. & Othman, O. Inheritance of tolerance to rice tungro bacilliform virus (RTBV) in rice (Oryza sativa L.). Theor. Appl. Genet. 80, 513–517 (1990).
Zenna, N. S. et al. Genetic analysis of tolerance to rice tungro bacilliform virus in rice (Oryza sativa L.) through agroinoculation. J. Phytopathol. 154, 197–203 (2006).
Bandillo, N. et al. Multi-parent advanced generation inter-cross (MAGIC) populations in rice: progress and potential for genetics research and breeding. Rice 6, 1–15 (2013).
Azzam, O., Imbe, T., Ikeda, R., Nath, P. D. & Coloquio, E. Inheritance of resistance to rice tungro spherical virus in a near-isogenic line derived from Utri Merah and in rice cultivar TKM6. Euphytica 122, 91–97 (2001).
Angeles, E. R. & Khush, G. S. Genetic analysis of resistance to green leafhopper, Nephotettix virescens (Distant), in three varieties of rice. Plant Breed. 119, 446–448 (2000).
Sebastian, L. S. et al. Genetic mapping of resistance to rice tungro spherical virus (RTSV) and green leafhopper (GLH) in ARC11554. Genetics https://doi.org/10.1142/9789812814289_0064 (2008).
Waing, F. P. et al. Characterization of the QTL linked to tungro and green leafhopper resistance in rice (Oryza sativa L.) using advance backcross population. Philipp. J. Crop Sci. 41, 20–30 (2016).
Ebron, L. A., Yumol, R. R., Ikeda, R. & Imbe, T. Inheritance of resistance to rice tungro spherical virus in some rice cultivars. Int. Rice Res. Notes. 19, 10–11 (1994).
Imbe, T. et al. Genetic studies in relation to breeding rice varieties resistant to rice tungro disease. in The Development of Stabilization Technology for Double Cropping in the Tropics. (Final Report of the IRRI-Government of Japan Collaborative Research Project., 1995).
Neeraja, C. N. et al. A marker-assisted backcross approach for developing submergence-tolerant rice cultivars. Theor. Appl. Genet. 115, 767–776 (2007).
Cho, J. H. et al. Development of allele specific SNP marker for RTSV resistance gene at the tsv1 locus. Plant Breed. Biotechnol. 6, 309–312 (2018).
Macovei, A. et al. Novel alleles of rice eIF4G generated by CRISPR/Cas9-targeted mutagenesis confer resistance to Rice tungro spherical virus. Plant Biotechnol. J. 16, 1918–1927 (2018).
Tyagi, H., Rajasubramaniam, S., Rajam, M. V. & Dasgupta, I. RNA-interference in rice against Rice tungro bacilliform virus results in its decreased accumulation in inoculated rice plants. Transgenic Res. 17, 897–904 (2008).
BRRI Annual Report. BRRI Annual Report (2018–2019). (2018).
Miah, G. et al. Marker-assisted introgression of broad-spectrum blast resistance genes into the cultivated MR219 rice variety. J. Sci. Food Agric. 97, 2810–2818 (2017).
Mackill, D. J. & Khush, G. S. IR64: A high-quality and high-yielding mega variety. Rice 11, (2018).
News, S. Sustainable Rice Platform. EU retailer survey points to emerging demand for sustainable rice (2021).
Iftekharuddaula, K. M. et al. Development of new submergence tolerant rice variety for bangladesh using marker-assisted backcrossing. Rice Sci. 22, 16–26 (2015).
Balachiranjeevi, C. H. et al. Identification of a novel locus, BPH38(t), conferring resistance to brown planthopper (Nilaparvata lugens Stal.) using early backcross population in rice (Oryza sativa L.). Euphytica 215, 185 (2019).
Hsu, Y. C., Chiu, C. H., Yap, R., Tseng, Y. C. & Wu, Y. P. Pyramiding bacterial blight resistance genes in tainung82 for broad-spectrum resistance using marker-assisted selection. Int. J. Mol. Sci. 21, (2020).
Zheng, J. et al. Marker-Assisted Pyramiding of Genes for Multilocular Ovaries , Self-Compatibility , and Clubroot Resistance in Chinese Cabbage ( Brassica rapa L . ssp . pekinensis ). (2022).
Kader, M. A., Aditya, T. L., Majumder, R. R., Hore, T. K. & Haq, M. E. Early maturing drought tolerant rice variety BRRI dhan71 suitable for drought prone environment in Bangladesh. Int. J. Plant Soil Sci. 32, 1–11 (2020).
BRRI Annual report. BRRI Annual report (2020–2021). (2021).
Hasan, M. M. et al. Review; Agriculture and Environmental Biotechnology Marker-assisted backcrossing: a useful method for rice improvement. Biotechnol. Biotechnol. Equip. 29, 237–254 (2015).
Seck, P. A., Diagne, A., Mohanty, S. & Wopereis, M. C. S. Crops that feed the world 7: Rice. Food Secur. 4, 7–24 (2012).
Collard, B. C. Y. & Mackill, D. J. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. B Biol. Sci. 363, 557–572 (2008).
Juma, R. U. et al. Identification of an elite core panel as a key breeding resource to accelerate the rate of genetic improvement for irrigated rice. Rice 14, 92 (2021).
Bhat, J. A., Yu, D., Bohra, A., Ganie, S. A. & Varshney, R. K. Features and applications of haplotypes in crop breeding. Commun. Biol. 4, 1266 (2021).
Wang, M. et al. A hybrid sterile locus leads to the linkage drag of interspecific hybrid progenies. Plant Divers. 42, 370–375 (2020).
Zhang, F., Shi, Y., Ali, J., Xu, J. & Li, Z. Breeding by selective introgression: Theory, practices, and lessons learned from rice. Crop J. 9, 646–657 (2021).
Biswas, P. S. et al. Development and field evaluation of near-isogenic lines of GR2-EBRRI dhan29 golden rice. Front. Plant Sci. 12, 1–15 (2021).
Balachiranjeevi, C. H. et al. Marker-assisted introgression of bacterial blight and blast resistance into DRR17B, an elite, fine-grain type maintainer line of rice. Mol. Breed. 35, (2015).
Caguiat, X. G. I., Waing, F. P., Millas, R. A., Aquino, J. D. & Tabanao, D. A. Genetic diversity of the coat protein gene of rice tungro bacilliform virus in the Philippines. Philipp. J. Sci. 149, 511–520 (2020).
Kumar, G. et al. Assessment of resistance to rice tungro disease in popular rice varieties in India by introgression of a transgene against Rice tungro bacilliform virus. Arch. Virol. 164, 1005–1013 (2019).
Arbelaez, J. D. et al. 1k-RiCA (1K-Rice Custom Amplicon) a novel genotyping amplicon-based SNP assay for genetics and breeding applications in rice. Rice 12, (2019).
Swamy, B. P. M. et al. Genetic, physiological, and gene expression analyses reveal that multiple QTL enhance yield of rice mega-variety IR64 under drought. PLoS ONE 8, e62795 (2013).
Swamy, B. P. M. et al. Development and characterization of GR2E Golden rice introgression lines. Sci. Rep. 11, 1–12 (2021).
Balachiranjeevi, C. H. et al. Marker-assisted pyramiding of two major, broad-spectrum bacterial blight resistance genes, Xa21 and Xa33 into an elite maintainer line of rice, DRR17B. PLoS ONE 13, e0201271–e0201271 (2018).
Nan, M. S. A. et al. Field evaluation of RD6 introgression lines for yield performance, blast, bacterial blight resistance, and cooking and eating qualities. Agronomy 9, (2019).
Koide, Y. et al. Genetic properties responsible for the transgressive segregation of days to heading in rice. G3 (Bethesda). 9, 1655–1662 (2019).
Swamy, B. P. M. et al. QTL analysis for grain quality traits in 2 BC2F2 populations derived from crosses between Oryza sativa cv. Swarna and 2 accessions of O. nivara. J. Hered. 103, 442–452 (2012).
Sharma, N. & Khanna, R. Rice Grain Quality: Current Developments and Future Prospects. Recent Advances in Grain Crops Research: Edited by Farooq Shah, Zafar Khan, Amjad Iqbal, Metin Turan and Murat Olgun. (2019https://doi.org/10.5772/intechopen.89367.
Sundaram, R. M. et al. Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 160, 411–422 (2008).
Yap, R., Hsu, Y., Wu, Y., Lin, Y. & Kuo, C. Multiplex PCR genotyping for five bacterial blight resistance genes applied to marker-assisted selection in rice (Oryza sativa). Plant Breed. 135, 309–317 (2016).
Luo, W. et al. Marker-assisted selection for rice blast resistance genes Pi2 and Pi9 through high-resolution melting of a gene-targeted amplicon. Plant Breed. 136, 67–73 (2017).
Malosetti, M., Ribaut, J.-M. & van Eeuwijk, F. A. The statistical analysis of multi-environment data: Modeling genotype-by-environment interaction and its genetic basis. Front. Physiol. 4 (2013).
Bishaw, Z. & van Gastel, A. J. G. Variety release and policy options. In Plant Breeding and Farmer Participation (eds Ceccarelli, S. et al.) (FAO, 2009).
Inabangan-Asilo, M. A. et al. Stability and G × E analysis of zinc-biofortified rice genotypes evaluated in diverse environments. Euphytica 215, 1–17 (2019).
Miura, K. et al. A metastable DWARF1 epigenetic mutant affecting plant stature in rice. Proc. Natl. Acad. Sci. USA. 106, 11218–11223 (2009).
IRRI. Standard Evaluation System for Rice. Int. Rice Res. Instiitute 55 (2013).
Juliano, B. O. Physicochemical properties of starch and protein in relation to grain quality and nutritional value of rice. IRRI Rice Breed. 389–405 (1972).
IRRI. Plant Breeding Tools (PB Tools), Version: 1.4, International Rice Research Institute, Los Banos. (2014).
R core Software. A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, Vienna, 2012). (2012).
Acknowledgements
BMZ for funding the project Leveraging Diversity for Ecologically based Pest Management (VERDE). NATP-2, BARC, Farmgate, Dhaka, Bangladesh supported PhD scholarship to TKH. The author(s) acknowledge the plant pathology group, cross-cutting operation (CCO) platform, analytical services laboratory (ASL) and genotyping services laboratory (GSL) at IRRI, Philippines for their enormous help.
Funding
This study was funded by Bundesministerium für Wirtschaftliche Zusammenarbeit und Entwicklung.
Author information
Authors and Affiliations
Contributions
T.K.H. conducted introgression. T.K.H., M.A.I.A. and R.W. conducted screen house and field testing at I.R.R.I. M.A.L. and S.N. conducted multi-location field testing in Bangladesh. B.P.M.S. conceived the idea. T.K.H. and B.C. wrote the manuscript. The manuscript was improved by J.E.H., G.B.G., T.U.D., M.G.Q.D. and B.P.M.S. All authors reviewed and improved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hore, T.K., Inabangan-asilo, M.A., Wulandari, R. et al. Introgression of tsv1 improves tungro disease resistance of a rice variety BRRI dhan71. Sci Rep 12, 18820 (2022). https://doi.org/10.1038/s41598-022-23413-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-23413-4
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
