
Abstract
Hepatocellular carcinoma (HCC) is progressively increasing tumor with lack of accurate prognosis and inadequate systemic treatment approaches. Solanum sp. (such as Solanum melongena) is a folk herb which is reported to possess anticancer properties. In a continuity for our interest in pursuing the anticancer activity of compounds isolated from the fruit peels of Solanum melongena, the HPLC profiling and ESI-MS assessment for the methanolic extract evidenced the presence of bioactive glycoalkaloids (solasonine, solasodine and solamargine). These glycoalkaloids were isolated, purified and proved to possess in vitro cytotoxicity against human liver cancer cell lines (Huh7 and HepG2). Herein, we investigated the potential mechanism of action of these compounds using DNA content flow-cytometry and apoptosis/necrosis differential anaylsis using annexin-V/FITC staining. Solasonine, solasodine and solamargine inducd significant antiproliferative effect against liver cancer cells (Huh7 and HepG2) which was attributed to cell cycle arrest at S-phase. Solamargine, solasodine and solasonine induced significant apoptosis in Huh7 cells. Only solamargine-induced cell cycle arrest, was reflected as apoptotic cell killing effect against HepG2 cells. In conclusion, glycoalkaloids derived from Solanum melongena and particularly, solamargine are promising antiproliferative agents with potential anticancer effects.
Similar content being viewed by others

Introduction
Hepatocellular carcinoma (HCC) is progressively increasing solid tumor type with poor prognosis and inadequate systemic treatment approaches. In human, almost 80% of patients die within one year of HCC diagnosis. In men, it is considered the fifth most common cancer and the third leading cause of cancer related mortality1,2. Chronic inflammatory liver disease due to high-fat diet, alcohol consumption, and chronic infection such as hepatitis virus B and C are the most common leading causes of HCC. Hepatitis C virus infection is considered the principal risk factor for HCC in Egypt3. HCC comprises national health problem; its incidence rate in Egypt alone is significantly larger than those observed in both USA and the rest of Middle Eastern countries4. Tumor development is correlated to both an increase in cell proliferations and a decrease in programmed cell death. It is now clear that development and progression of various liver diseases are accompanied with minimal increase or decrease in hepatocyte apoptosis. This in turn leads to extending hepatocyte cell viability and accumulated genetic mutations5.
Compounds derived from natural origin such as, herbal products and other folk remedies draw great attention as a treatment modality for several illnesses such as, malegnancies6. Several natural compounds and phytochemicals represent milestone chemotherapeutic agents which showed significant anticancer effects such as paclitaxel, doxorubicin, vincristine and others7. Solanum sp. is a folk herb which is abundant in open fields. It is frequently reported for the treatment of several cancers such as, cervical carcinoma, breast cancer, melanoma and most interestingly, liver cancer8,9,10,11,12. In terms of folk use, Solanum sp. was used for the treatment of edema, mastitis, inflammatory disorders13 and fever besides its robust anti oxidant and cytoprotective effects14,15.
Glycoalkaloids are class of steroidal glycosides which are structurally diverse and display broad spectrum of biological activities such as antibacterial, anti-inflammatory, and anticancer activities16. It was found that both non-sugar and sugar moieties are essential for the glycoalkaloids biological activity17. The conjugates of solasodine aglycone showed anticancer activity against human colon and liver cancer cells18. In our previous study, we isolated five steroidal glycosides from the methanolic extract of Solanum melongena fruit peels (MEP). MEP along with the isolated compounds were tested in vitro against five human cancer cell lines; colon cancer cell line HCT116, larynx cancer cell line HEP2, breast cancer cell line MCF7, cervix cancer cell line HeLa and liver cancer cell line HepG2. Solasonine, solasodine, and solamargine demonstrated the most potent activity among the tested compounds. Remarkably, human liver cancer cell line (HepG2) was considerably sensitive to the aforementioned compounds9. Herein, we describe the HPLC profile of MEP to confirm the presence of those glycoalkaloids (solasonine, solasodine and solamargine) in MEP. The identity of the compounds was further confirmed by ESI-MS. Furthermore, we investigated in some details the antiproliferative/cytotoxic, cell cycle interfering and apoptosis inducing profile of the three biologically active glycoalkaloids (solasonine, solasodine and solamargine) against two liver cancer cell lines HepG2 and Huh-7 cells.
Results
HPLC-PDA Detection of solasodine, solasonine, and solamargine in MEP
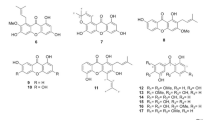
HPLC chromatograms S. melongena fruit peels (MEP) was monitored at 200 nm (Fig. 1-A). Solasodine, solasonine, and solamargine authentics have been injected using the same HPLC conditions to assign the chemical identity of the eluted peaks. Solasodine was first to be eluted from the MEP at 12 min (Fig. 1-B) followed by solasonine at 14.3 min (Fig. 1-C), then solamargine at 15 min (Fig. 1-D). The ESI-MS of the isolated compounds is displayed in Fig. 2(A–C). The structures of the isolated/tested compounds are displayed in Fig. 2-D.

HPLC-PDA chromatogram of the methanolic extract of Solanum melongena fruit peels (A), solasodine standard (B) solasonine standard (C) and solamargine standard (D) monitored at 200 nm.

ESI-MS of solasodine (A), solamargine (B) and solasonine (C) in the positive ion mode. Structures of the isolated compounds from the MEP (D).
Dose response relationship of solasonine, solasodine and solamargine against liver cancer cells (huh-7 and HepG2
SRB-U assay was used to assess the cytotoxicity of the isolated glycoalkaloids against two different liver cancer cell lines (Huh-7 and HepG2) over concentration range of 0.01–100 μM. Tested compounds showed comparable cytotoxicity profile against both liver cell lines under investigation. However, Huh-7 cells were relatively more susceptible to cell killing effect copared to HepG2 cells.
In Huh-7 cells, solasonine, solasodine and solamargine showed comparable cytotoxicity with IC50’s of 10.3 ± 1.5 μM, 11.7 ± 0.3 μM and 9.6 ± 0.5 μM, respectively. In addition, R-values (resistance fraction) for solasonine and solasodine were 19.2 ± 2.6% and 10.8 ± 1.0%, respectively. It is worth mentioning that, the R-value for solamargine was interestingly low (less than 1%) within Huh-7 cells (Fig. 3-A).

Dose response relationship of solasonine, solasodine and solamargine against Huh-7 (A) and HepG2 cells (B). Cells were exposed to serial concentrations of solasonine (●), solasodine (○) or solamargine (▼) for 72 h and viability was assessed by using SRB assay. Data are presented as mean ± SD (n = 3).
With respect to HepG2 cell line, solasonine showed significantly weaker cytotoxicity compared to Huh-7 cells with IC50 of 91.8 ± 9.4 μM. solasodine showed moderate cytotoxicity with IC50 equals 19.4 ± 0.4 μM. Interestingly, solamargine showed comparable potency against HepG2 cells with IC50 of 10.8 ± 0.1 μM. Similar to Huh-7 cells, solamargine did not suffer from any significant resistance (R-value was less than 1%) and none of the other two glycoalkaloids showed resistance value higher than 10% (Fig. 3-B.
Influence of solasonine, solasodine and solamargine on the cell cycle distribution of Huh-7 cells
In our previous work, solasonine, solasodine and solamargine showed promising anti-proliferative/cytotoxic effects against different cancer cell lines at concentration range of 1–10 µM. Herein, we further investigated the interference of these glycoalkaloids to cell cycle phases of Huh-7 liver cancer cells. Solasonoine and solasodine induced moderate cell cycle arrest at S-phase and increased percentage of cells in S-phase from 28.2 ± 1.6% to 32.1 ± 0.7% and 35.4 ± 1.8%, respectively. Despite cells accumulating in S-phase due to treatment with solasonine and solasodine, no significant change was noticed in G2/M-phase after treatment with both glycoalkaloid. On the other hand, S-phase arrest induced in Huh-7 cells due to solamargine (36.8 ± 3.1%) decreased cell propagation to G2/M-phase from 22.1 ± 3.0% to 13.9 ± 5.2% (Fig. 4-A,B). In alignment with the induced S-phase arrest, solamargine and, to a lesser extent, solasonine significantly decreased cell population in SupraG2-phase from 10.4 ± 1.3% to 1.0 ± 0.3% and 7.9 ± 1.2%, respectively (Fig. 4-C). In addition, solasonine, solasodine and solamargine significantly increased dead cell population (Pre-G phase) from 0.7 ± 0.1% to 1.8 ± 0.1%, 3.0 ± 0.1% and 21.1 ± 2.0%, respectively (Fig. 4-D).

Effect of solasonine, solasodine and solamargine on the cell cycle distribution of Huh-7 cells. Cells were exposed to test compounds (10 µM) for 24 h (A–D) or 48 h (E–H)and cell cycle histograms were compared to control cells (A,E); and different cell cycle phases were plotted as percent of total events (B,F). Percentages of cells in pre-G1 phase (C,G) and supra-G2 phase (D,H) were assessed numerically and compared to control treatment. Data are presented as mean ± SD (n = 3); and (*) indicates significantly different from control group at P < 0.05.
Further exposure of Huh-7 cells to solasonine, solasodine and solamargine for 48 h resulted in further significant cell cycle arrest at S-phase; solamargine significantly increased S-phase cell population from 34.4 ± 0.9% to 43.9 ± 0.8% with reciprocal decrease in G0/G1-phase from 44.0 ± 1.3% to 31.5 ± 1.8%. In addition, solasonine and solasodine increased S-phase cell population to 39.6 ± 2.7% and 36.6 ± 0.4%, respectively. Only solamargine induced significant cell cycle arrest at G2/M-phase as well and increased cell population in G2/M-phase from 21.7 ± 0.5% to 24.6 ± 1.9% (Fig. 4-E,F). In addition and due to the induced G2/M-phase arrest, solamargine significantly decreased cell population in SupraG2-phase from 11.3 ± 1.2% to 6.8 ± 0. 8%. On contrary, solasonine increased cells in supraG2 phase to 15.0 ± 0.9% (Fig. 4-G). Moreover, solasodine and solamargine significantly increased dead cell population (Pre-G phase) from 1.3 ± 0.1% to 1.9 ± 0.1% and 8.5 ± 0.1%, respectively (Fig. 4-H).
To confirm our observations about cell cycle interference, more resistant liver cancer cell line (HepG2) was used.
Influence of solasonine, solasodine and solamargine on the cell cycle distribution of HepG2 cells
Besides Huh-7 cells, we further investigated the interference of these glycoalkaloids to cell cycle phases of the more resistant HepG2 liver cancer cells. Similar to Huh-7 cells, solasonine, solasodine and solamargine induced significant S-phase cell cycle arrest and increased percentage of cells in S-phase from 41.0 ± 0.3% to 50.6 ± 3.2%, 50.3 ± 1.0% and 45.0 ± 0.8%, respectively. Reciprocally, treatment with solasonine, solasodine and solamargine induced significant increase in cells in G2/M-phase from 17.7 ± 0.4% to 14.1 ± 1.3%, 14.4 ± 0.8% and 14.8 ± 0.3%, respectively; and significant decrease in G0/G1-phase from 41.4 ± 0.6% to 35.3 ± 2.0%, 35.2 ± 0.2% and 40.3 ± 0.6%, respectively (Fig. 5-A,B). Solasodine significantly increased cell population in SupraG2-phase from 16.1 ± 0.8% to 24.8 ± 1.7%; while solamargine decreased SupraG2-phase cells to 13.4 ± 0.6% (Fig. 5-C). Furthermore, solasonine, solasodine and solamargine significantly increased dead cell population (Pre-G phase) from 1.8 ± 0.1% to 2.3 ± 0.4%, 3.0 ± 0.2% and 3.7 ± 0.3%, respectively (Fig. 5-D).

Effect of solasonine, solasodine and solamargine on the cell cycle distribution of HepG2 cells. Cells were exposed to test compounds ( 10 µM) for 24 h (A–D) or 48 h (E–H) and cell cycle histograms were compared to control cells (A,E); and different cell cycle phases were plotted as percent of total events (B,F). Percentages of cells in pre-G1 phase (C,G) and supra-G2 phase (D,H) were assessed numerically and compared to control treatment. Data are presented as mean ± SD (n = 3); and (*)indicates significantly different from control group at P < 0.05.
Extended exposure of HepG2 cells to solasonine for 48 h resulted in significant cell cycle accumulation at G2/M-phase from 16.4 ± 0.9% to 22.0 ± 2.5%. Solasodine significantly increased G2/M-phase cell population from 16.4 ± 0.9% to 23.7 ± 1.7% with reciprocal decrease in G0/G1-phase from 38.0 ± 1.3% to 34.8 ± 1.1%; and in S-phase from 45.5 ± 1.4% to 41.5 ± 0.6%. (Fig. 5-E,F). The three tested glycoalkaloids showed non-significant change in supraG2 phase cells (Fig. 5-G). In addition, solasodine and solamargine significantly increased dead cell population (Pre-G phase) from 1.5 ± 0.1% to 2.0 ± 0.1% and 2.3 ± 0.1%, respectively (Fig. 5-H).
Further analysis for the mechanism of cell damage (programmed vs. non-programmed) induced by our test compounds were studied using annexin V-FITC/PI differential staining.
Apoptosis/necrosis assessment using flow cytometry
Annexin V-FITC/PI staining coupled with flow cytometry was used to differentially assess proportion of cells dying via necrosis versus cells undergoing apoptosis in both Huh-7 and HepG2 cells. Cells were treated with 10 μM of solasonine, solasodine and solamargine for 24 h and 48 h prior to apoptosis/necrosis differential assessment. In Huh-7 cells, all glycoalkaloids under investigation (solasonine, solasodine and solamargine) significantly increased apoptosis (early and late) by 1.6, 2 and 5.8 folds, respectively compared to control cells; and consequently increased total cell death. Only solasodine induced non-specific necrotic cell death (3.3 folds) in Huh-7 cells (Fig. 6-A,C). Further exposure of huh-7 cells to solasonine, solasodine and solamargine resulted in similar profile of apoptotic cell death; they significantl increased the apoptosis cell fractions (early and late) by 2.3, 3.6 and 7.6 folds, respectively compared to control (Fig. 6-B,D).

Cell death mechanism in Huh-7 cells after treatment with solasonine, solasodine and solamargine. Cells were exposed to 10 µM of solasonine, solasodine and solamargine for 24 (A) and 48 h (B) and percentages of cells undergoing apoptosis/necrosis were assessed using Annexin-V/FITC-PI staining and compared to control cells (C,D). Data are presented as mean ± SD (n = 3); and (*) indicates significantly different from control group at P < 0.05.
With respect to HepG2 cells, only solamargine showed significantly more cells with early apoptosis compared to control untreated cells. In addition,solasodine and solamargine showed significant apoptosis induction (2 and 3 folds more late apoptosis cells compared to control untreated cells, respectively); and marginal overall killing effect (1.6 and 2.8 folds, respectively) after 24 h of exposure. (Fig. 7-A,C). Further exposure of HepG2 cells to solasodine and solamargine to 48 h showed similar results in terms of cells in the late apoptosis phase (1.2 and 2.9 folds, respectively) and overall killing effect (1.5 and 3.3 folds, respectively) (Fig. 7-B,D). It is worth to mention that both solasodine and solamargine induced significant necrosis which was indicative of non-specific killing effect (Fig. 7).

Cell death mechanism in HepG2 cells after treatment with solasonine, solasodine and solamargine. Cells were exposed to 10 µM of solasonine, solasodine and solamargine for 24 (A) and 48 h (B) and percentages of cells undergoing apoptosis/necrosis were assessed using Annexin-V/FITC-PI staining and compared to control cells (C,D). Data are presented as mean ± SD (n = 3); and (*) indicates significantly different from control group at P < 0.05.
Discussion
Developing countries are experiencing increasing rates of liver cancer. HCC is one of the three most commonly diagnosed cancers in Egypt that has a high mortality rate and limited therapeutic opportunities. Current mortality due to liver cirrhosis and cancer is over 40,000/year and is increasing annually19. Glycoalkaloids are interestingly biologically anticancer active compounds which can be found in several natural herbs such as Solanum melongena18. Herein and as a continuation of our previous work9, we explored the detailed antiproliferative/cytotoxic profile of three biologically active glycoalkaloids (solasonine, solasodine and solamargine) isolated from Solanum melongena against resistant and sensitive liver cancer cells (HepG2 and Huh7).
In the current study, we examined the HPLC profile of the total methanolic extract of Solanum melongena in terms of glycoalkaloids abundance. Solasodine, solasonine, and solamargine were observed, identified and quantified in the extract using ESI-MS technique. Yet, the underlying mechanisms of the observed in vitro cytotoxic activity of Solanum melongena total extract and the isolated glycoalkaloids against HepG2 cell lines was unclear9. Potential anticancer property of a compound might be attributed to its antiproliferative effects or direct cytotoxic activity20. We studied these two proposed modes of action for the isolated glycoalkaloids (solasodine, solasonine, and solamargine) using cell cycle analysis and annexin-V apoptosis assessment. In several cases, excessive/prolonged cell cycle arrest results in cell death and ultimately cytotoxicity21,22,23.
In the current work, all isolated glycoalkaloids induced obvious antiproliferative effect against Huh7 cells which could be attributed to the significant cell cycle arrest at S-phase. This cell cycle arrest was extended over 48 h and was most prominent after treatment with solamargine. Many compounds of natural origin exert their anticancer properties via S-phase arrest and cell cycle catastrophe24. Not only S-phase arrest, but also did solamargine induce cell cycle arrest at G2/M phase. This explains the decreased cell population in the supraG2 phase. Extended cell cycle arrest induced by solamargine and to a lesser extent by solasodine induced cell killing effect which could be observed by elevated cell population in the pre-G phase. Besides, the three glycoalkaloids influenced cell cycle progression of HepG2 cells similar to Huh7 cells; solasosonine, solasodine and solamargine induced significant cell cycle arrest at S-phase with compensatory cell accumulation in either G0/G1 and/or G2/M-phases. Yet, cell cycle arrest induced by solasonine and solamargine were relieved after extended exposure for 48 h. This could be explained by eliminated arrested cells by apoptosis cell death25. In addition, accumulation of cells in Pre-G phase, which is indicative of cell death, due to cell cycle arrest in HepG2 cells was much lower than Huh7 cells. HepG2 liver cancer cells were found before to be more resistant to anticancer treatment compared to Huh7 cells26; and other tumor cell lines as well20.
Programmed cell death induced by chemotherapy against cancer cells is preferred than non-pragrammed cell death27. Generally, the three glycoalkaloids under investigation increased pre-G cell cycle phase to different extends which is indicative of cell killing effect. Yet, we examined whether this killing effect is programmed or non-programmed in nature. Interestingly, all the three glycoalkaloids induced significant apoptosis in Huh7 cells; and minimal necrosis (non-programmed cell death) was induced only by solasodine. With respect to the more resisitant liver cancer cell line (HepG2), only solasodine and solamargine induced significant apoptosis and necrosis cell death. It was previously reported that glycoalkaloids (such as solasodine) showed better anticancer activities than their aglycone28. It was explained by the ability of rhamnose sugar moity to mediate binding to the cancer cells receptors29. Yet, the exact apoptosis mechanism and its underlying molecular pathways are strongly recommended for further investigation.
Conclusion
Herein, the composition of the glycoalkaloid-rich extract of Solanum melongena was authenticated by HPLC and purified to be used as a chemotherapeutic agent. The current study confirms the antiproliferative anticancer activity of three naturally occurig glycoalkaloids (solasonine, solasodine and solamargine) isolated from the methaolic extract of the fruit peels of Solanum melongena against two liver cancer cell lines of different susceptibility. Particularly, solamargine was the most potent among the isolated glycoalkaloids followed by solasodine; and the least active anticancer glycoalkaloid was solasonine. The isolated glycoalkaloids induced antiproliferative effect which is attributed to inhibiting cell cycle progression in S-phase; this in turn induced progressive cell apoptosis and ultimately cell death. These compounds worth further investigation as potential therapeutic leads for anticancer drug development.
Materials and Methods
Extraction and isolation
The methanolic extract of the fruits peels (MEP) as well as the compounds; solasonine, solasodine and solamargine were prepared/or isolated by the authors as previously described9.
Analysis of the MEP of Solanum melongena fruit peels (MEP) using RP-HPLC
Sample preparation
The MEP (20 mg) was dissolved in 1 ml of 40% methanol in H2O, using an ultrasonic, and loaded on an EXtrelut® prepacked column (Merck). Elution was carried out using 7 × 1 ml, 50% methanol in H2O, and the volume completed to 10 ml in a volumetric flask.
HPLC method
Samples were analyzed using Waters 2695 HPLC system equipped with: a photodiode array detector (PDA) UV detector, Masslynx V4.1 SCN 714 software, and analytical BDS-Hypersil C18 column (250 × 4.6 mm i.d.; particle size 5 μm; Thermo). The analyses were carried out employing a gradient elution system. Solvent A was sodium phosphate buffer (pH 7.2; 0.01 M) and solvent B was acetonitrile. The gradient elution program was employed at a flow-rate of 1.0 mL/min, consisting of: 0–2 min (20% B, isocratic); 2–26 min (70% B, linear gradient); 26–30 min (85% B, isocratic, washing column); 30–31 min (20% B, linear gradient); 31–35 min (20% B, isocratic, column equilibration). A 10 μL of 5 mg/mL of MEP was injected. The following standards were injected to verify the identity of the eluted peaks: 1 μL of 5 mg/mL of solasodine standard, 5 μL of 4 mg/mL of solasonine standard, and 5 μL of 7 mg/mL of solamargine standard. Additionally, each standard was directly infused into MS to double confirm the identity of the standards. ESI-MS of the isolated compounds was detected from m/z 50 to 1000 using a MS QQQ mass spectrometer equipped with an electrospray ion source in positive ion mode. The following instrument settings were used; nebulizer gas, nitrogen, 40 psi; dry gas, nitrogen, 10 ml/min, 300 °C; capillary, −3000 V (+4000 V); end plate offset, −500 V; funnel 1 RF, 200 Vpp; funnel 2 RF, 200 Vpp.
Cell culture
Human liver cancer cell lines (HepG2 and Huh-7) were obtained from the Vacsera (Giza, Egypt). Cells were maintained in DMEM culture medium, supplemented with 100 µg/ml streptomycin, 100 units/ml penicillin and 10% heat-inactivated fetal bovine serum. Cells were kept passaging in subconfluent state in humidified 5% CO2 (v/v) atmosphere at 37 °C as previously described30.
Cytotoxicity assessment
The cytotoxicity of the isolated glycoalkaloids was tested against Huh-7 and HepG2 liver cancer cells by SRB assay as previously described31. Briefly, exponentially growing cells were trypsinized by 0.25% Trypsin-EDTA and seeded in 96-well plates at 1000–2000 cells/well. Cells were treated with serial concentrations of the isolated compounds for 72 h and subsequently fixed with TCA (10%) for 1 h at 4 °C. After several washings with water, cells were stained with 0.4% SRB solution for 10 minutes at room temperature in dark place and subsequently washed with 1% glacial acetic acid. After drying the plates overnight, Tris-HCl was used to dissolve the SRB stained cells and color intensity was measured at 540 nm with microplate reader (Spectramax® M3, Molecular devices, San Jose, CA, USA. Each concentration was replicated 6 times and the whole experiment was repeated three times; data represents mean ± SD of three replicates.
Data analysis
The dose-response curves were analyzed as previously described20 using Emax model (Eq. 1).
Where [R] is the residual unaffected fraction (the resistance fraction), [D] is the drug concentration used, [Kd] or IC50 is the drug concentration that produces a 50% reduction of the maximum inhibition rate and [m] is a Hill-type coefficient. Absolute IC50 is defined as the drug concentration required to reduce absorbance by 50% of control group (i.e., Kd = absolute IC50 when R = 0 and Emax = 100 − R).
Cell cycle analysis
To assess the effect of isolated alkaloids (solasonine, solasodine and solamargine) on cell cycle distribution, cells were incubated with 5 µM of the test compounds for 24 h and 48 h. Cells were collected by trypsinization; washed twice with ice-cold PBS and re-suspended in 0.5 ml PBS. Two milliliters of 70% ice-cold ethanol was added gently while shaking. Cells were kept in ethanol solution at 4 °C for 1 hour for fixation. Upon analysis, cells were washed and re-suspended in 1 ml of PBS containing 50 μg/mL RNAase A and 10 μg/mL propidium iodide (PI). After 20 minutes incubation in dark place at room temperature, cells were analyzed for DNA contents by ACEA Novocyte™ flowcytometer (ACEA Biosciences Inc., San Diego, CA, USA) and analyzed for PI fluorescent signals using FL2 detector (λex/em 535/617 nm). For each sample, 12,000 events were acquired. Percent of cells in each cell cycle phase was analyzed and calculated using ACEA NovoExpress™ software (ACEA Biosciences Inc., San Diego, CA, USA). Each treatment was repesated three times and data represents mean ± SD of three replicates.
Apoptosis assessment using annexin V-FITC/PI staining coupled with flowcytometry
To assess the effect of isolated alkaloids on programmed cell death, apoptosis and necrosis cell populations were determined using Annexin/V-FITC apoptosis detection kit (Abcam Inc., Cambridge Science Park, Cambridge, UK). Briefly, cells were treated with 5 µM of solasonine, solasodine and solamargine for 24 h and 48 h. After treatment, cells were collected by trypsinization, washed twice with ice-cold PBS, and re-suspended in 0.5 mL of annexin/V-FITC/PI solution for 30 min in dark according to manufacturer protocol. After staining at room temperature, cells were injected through ACEA Novocyte™ flowcytometer (ACEA Biosciences Inc., San Diego, CA, USA) and analyzed for FITC and PI fluorescent signals using FL1 and FL2 signal detector, respectively (λex/em 488/530 nm for FITC and λex/em 535/617 nm for PI). For each sample, 12,000 events were acquired and positive FITC and/or PI cells were quantified by quadrant analysis and calculated using ACEA NovoExpress™ software (ACEA Biosciences Inc., San Diego, CA, USA). Each treatment was repesated three times and data represents mean ± SD of three replicates.
Statistical analysis
Data are presented as mean ± SD. Analysis of variance (ANOVA) followed by Tukey’s post hoc test was used for testing the significance using SPSS for windows, version 17.0.0. p-value of 0.05 was taken as a cut off value for significance.
References
Torre, L. A. et al. Global cancer statistics, 2012. CA Cancer J Clin 65, 87–108 (2015).
Altekruse, S. F., McGlynn, K. A. & Reichman, M. E. Hepatocellular carcinoma incidence, mortality, and survival trends in the United States from 1975 to 2005. J. Clin. Oncol. 27, 1485–1491 (2009).
Strickland, G. T. Liver disease in Egypt: hepatitis C superseded schistosomiasis as a result of iatrogenic and biological factors. Hepatology 43, 915–922 (2006).
Freedman, L. S., Edwards, B. K., Ries, L. A. G. & Young, J. L. Cancer incidence in four member countries (Cyprus, Egypt, Israel, and Jordan) of the Middle East Cancer Consortium (MECC) compared with US SEER. Cancer Incid. four Memb. Ctries. (Cyprus, Egypt, Isr. Jordan) Middle East Cancer Consort. Comp. with US SEER (2006).
El–Serag, H. B. & Rudolph, K. L. Hepatocellular carcinoma: epidemiology and molecular carcinogenesis. Gastroenterology 132, 2557–2576 (2007).
Craig, W. J. Phytochemicals: guardians of our health. J Am Diet Assoc 97, S199–204 (1997).
Mann, J. Natural products in cancer chemotherapy: past, present and future. Nat Rev Cancer 2, 143–148 (2002).
Almehdar, H., Abdallah, H. M., Osman, A. M. M. & Abdel-Sattar, E. A. In vitro cytotoxic screening of selected Saudi medicinal plants. J. Nat. Med. 66, 406–412 (2012).
Shabana, M. M., Salama, M. M., Ezzat, S. M. & Ismail, L. R. In vitro and in vivo anticancer activity of the fruit peels of Solanum melongena L. against hepatocellular carcinoma. J. Carcinog. Mutagen. 2013 (2013).
El-Sayed, M. A. et al. Cytotoxicity of 3-O-(beta-D-glucopyranosyl) etioline, a steroidal alkaloid from Solanum diphyllum L. Z Naturforsch C 64, 644–649 (2009).
Wang, H. C., Wu, D. H., Chang, Y. C., Li, Y. J. & Wang, C. J. Solanum nigrum Linn. water extract inhibits metastasis in mouse melanoma cells in vitro and in vivo. J Agric Food Chem 58, 11913–11923 (2010).
Li, J., Li, Q., Feng, T. & Li, K. Aqueous extract of Solanum nigrum inhibit growth of cervical carcinoma (U14) via modulating immune response of tumor bearing mice and inducing apoptosis of tumor cells. Fitoterapia 79, 548–556 (2008).
Zakaria, Z. A. et al. Antinociceptive, anti-inflammatory and antipyretic effects of Solanum nigrum chloroform extract in animal models. Yakugaku Zasshi 126, 1171–1178 (2006).
Sultana, S., Perwaiz, S., Iqbal, M. & Athar, M. Crude extracts of hepatoprotective plants, Solanum nigrum and Cichorium intybus inhibit free radical-mediated DNA damage. J Ethnopharmacol 45, 189–192 (1995).
Prashanth Kumar, V., Shashidhara, S., Kumar, M. M. & Sridhara, B. Y. Cytoprotective role of Solanum nigrum against gentamicin-induced kidney cell (Vero cells) damage in vitro. Fitoterapia 72, 481–486 (2001).
Sparg, S., Light, M. E. & Van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 94, 219–243 (2004).
Wang, Y. et al. Exploration of the correlation between the structure, hemolytic activity, and cytotoxicity of steroid saponins. Bioorg. Med. Chem. 15, 2528–2532 (2007).
Lee, K.-R. et al. Glycoalkaloids and metabolites inhibit the growth of human colon (HT29) and liver (HepG2) cancer cells. J. Agric. Food Chem. 52, 2832–2839 (2004).
Jemal, A., Center, M. M., DeSantis, C. & Ward, E. M. Global patterns of cancer incidence and mortality rates and trends. Cancer Epidemiol. Biomarkers Prev. 19, 1893–1907 (2010).
Mohamed, G. A., Al-Abd, A. M., El-Halawany, A. M., Abdallah, H. M. & Ibrahim, S. R. M. New xanthones and cytotoxic constituents from Garcinia mangostana fruit hulls against human hepatocellular, breast, and colorectal cancer cell lines. J. Ethnopharmacol. 198, 302–312 (2017).
Kuh, H. J. et al. A computational model for quantitative analysis of cell cycle arrest and its contribution to overall growth inhibition by anticancer agents. Jpn J Cancer Res 91, 1303–1313 (2000).
Sanchez, C. A. et al. Statin-induced inhibition of MCF-7 breast cancer cell proliferation is related to cell cycle arrest and apoptotic and necrotic cell death mediated by an enhanced oxidative stress. Cancer Invest 26, 698–707 (2008).
Mahmoud, A. M. et al. Transcriptional profiling of breast cancer cells in response to mevinolin: Evidence of cell cycle arrest, DNA degradation and apoptosis. Int. J. Oncol. 48 (2016).
Joe, A. K. et al. Resveratrol induces growth inhibition, S-phase arrest, apoptosis, and changes in biomarker expression in several human cancer cell lines. Clin Cancer Res 8, 893–903 (2002).
Pucci, B., Kasten, M. & Giordano, A. Cell cycle and apoptosis. Neoplasia 2, 291–299 (2000).
Al-Abbasi, F. A. et al. Gingerol synergizes the cytotoxic effects of doxorubicin against liver cancer cells and protects from its vascular toxicity. Molecules 21 (2016).
Hannun, Y. A. Apoptosis and the dilemma of cancer chemotherapy. Blood 89, 1845–1853 (1997).
Cui, C. et al. Synthesis of solasodine glycoside derivatives and evaluation of their cytotoxic effects on human cancer cells. Drug Discov. Ther. 6, 9–17 (2012).
Zha, X., Sun, H., Hao, J. & Zhang, Y. Efficient Synthesis of Solasodine, O‐Acetylsolasodine, and Soladulcidine as Anticancer Steroidal Alkaloids. Chem. Biodivers. 4, 25–31 (2007).
Mahmoud, A. M., Al-Abd, A. M., Lightfoot, D. A. & El-Shemy, H. A. Anti-cancer characteristics of mevinolin against three different solid tumor cell lines was not solely p53-dependent. J. Enzyme Inhib. Med. Chem. 27 (2012).
Skehan, P. et al. New colorimetric cytotoxicity assay for anticancer-drug screening. J Natl Cancer Inst 82, 1107–1112 (1990).
Acknowledgements
The authors are grateful to Dr. Hawazen Bin Mahfouz, Faculty of Medicine, King Abdulaziz University, for her technical support during the course of this work.
Author information
Authors and Affiliations
Contributions
Mostafa I. Fekry performed the HPLC analysis. Shahira M. Ezzat and Maha M. Salama performed extraction, purification and shared in the identification of the compounds. Ohoud Y. Alshehri and Ahmed M Al-Abd performed the biological activity evaluation (dose response analysis, cell cycle analysis and apoptosis assessment). Mostafa I. Fekry, Maha M. Salama and Ahmed M. Al-Abd wrote the manuscript. Shahira M. Ezzat and Ahmed M Al-Abd prepared the figures. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fekry, M.I., Ezzat, S.M., Salama, M.M. et al. Bioactive glycoalkaloides isolated from Solanum melongena fruit peels with potential anticancer properties against hepatocellular carcinoma cells. Sci Rep 9, 1746 (2019). https://doi.org/10.1038/s41598-018-36089-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-36089-6
This article is cited by
-
Revisiting the ethnomedicinal, ethnopharmacological, phytoconstituents and phytoremediation of the plant Solanum viarum Dunal
Naunyn-Schmiedeberg's Archives of Pharmacology (2024)
-
Deciphering the effects of kosmotrope and chaotrope salts during aqueous two phase extraction (ATPE) of polyphenolic compounds and glycoalkaloids from the leaves of a nutraceutical plant, Solanum retroflexum, with the aid of UPLC-QTOF-MS
Applied Biological Chemistry (2021)
-
Haimufang decoction, a Chinese medicine formula for lung cancer, arrests cell cycle, stimulates apoptosis in NCI-H1975 cells, and induces M1 polarization in RAW 264.7 macrophage cells
BMC Complementary Medicine and Therapies (2020)
-
Evaluation of biomarkers in liver following Solanum melongena green calyx administration in diabetic rats
Journal of Diabetes & Metabolic Disorders (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
