
Abstract
Blastocystis is a species complex that exhibits extensive genetic diversity, evidenced by its classification into several genetically distinct subtypes (ST). Although several studies have shown the relationships between a specific subtype and gut microbiota, there is no study to show the effect of the ubiquitous Blastocystis ST1 on the gut microbiota and host health. Here, we show that Blastocystis ST1 colonization increased the proportion of beneficial bacteria Alloprevotella and Akkermansia, and induced Th2 and Treg cell responses in normal healthy mice. ST1-colonized mice showed decreases in the severity of DSS-induced colitis when compared to non-colonized mice. Furthermore, mice transplanted with ST1-altered gut microbiota were refractory to dextran sulfate sodium (DSS)-induced colitis via induction of Treg cells and elevated short-chain fat acid (SCFA) production. Our results suggest that colonization with Blastocystis ST1, one of the most common subtypes in humans, exerts beneficial effects on host health through modulating the gut microbiota and adaptive immune responses.
Similar content being viewed by others

Introduction
The human gut comprises thousands of diverse microbes including bacteria, fungi, archaea, viruses, and single cell eukaryotes (SCEs)1. Although numerous studies have focused on the effect of gut bacteria on host health and diseases over the past decades, the role of SCEs in the gut ecosystem has only recently begun to be recognized2. Blastocystis is a common gut SCE found in humans and a wide range of animals with an estimated more than 1 billion people colonized worldwide3. It mainly colonizes the host’s large intestine, where it has a close relationship with gut microbiota4. The relationship between Blastocystis and gut microbiota is an emerging area of research interest, and multiple studies have shown that Blastocystis colonization is asymptomatic and often associated with healthy gut microbiota5,6,7. However, other studies showed Blastocystis was related to decreased cognitive function in humans and colonic hypersensitivity in animal models8,9. The differences observed in previous studies could be attributed to the distinct characteristics of the various Blastocystis subtypes, resulting in differential effects on host health10.
To date, at least 30 subtypes have been identified across humans and animals based on the variations within sequences of the small subunit (SSU) rRNA gene, and more subtypes are emerging11. Blastocystis subtypes were also considered separate species that exhibited distinct biological features12. For example, considerable differences in the number of genomic features like DNA base composition, genome size, and the number of genes and introns were observed between ST1, ST4, and ST713. Blastocystis ST7 can elevate the expression of pro-inflammatory cytokines interleukin (IL)-6, IL-1β, and tumor necrosis factor-alpha (TNFα) in murine macrophages, while Blastocystis ST4 did not show such an effect14. Besides, Blastocystis ST3 and ST4 colonization showed inverse correlations to the proportion of beneficial bacteria Akkermansia in a large cohort in Europe7. Akkermansia may play crucial roles in the prevention of several metabolic diseases, such as obesity, type 2 diabetes or hypertension15.
It has been reported that Blastocystis ST4 colonization triggers potent Th2 and Treg immune responses, which help experimentally colonized mice recover faster from experimentally-induced colitis16. Similarly, long-term colonization with Blastocystis ST3 could promote faster recovery from colitis and decrease the expression of TNFα and IL-1β in dinitrobenzene sulfonic acid (DNBS)-induced colitis mice17. In contrast, Blastocystis ST7 infection worsens the severity of colitis induced by dextran sulfate sodium (DSS) in a mouse model18. These data suggest that it is necessary to investigate the effects of Blastocystis on host health at the subtype level. In this study, we investigated a specific subtype, ST1, one of the most common subtypes in humans worldwide19. We firstly investigated the effect of Blastocystis ST1 colonization on gut microbiota and adaptive immune responses, and then further explored the role of ST1-altered gut microbiota in the development of experimentally-induced colitis. Our data provide valuable insights into host-parasite interactions.
Results
Effects of Blastocystis ST1 colonization on normal healthy mice
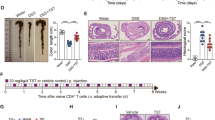
Multiple epidemiologic studies have shown that Blastocystis ST1 is one of the most common subtypes in humans19, while the effect of ST1 colonization on host health remains largely unknown. The mice were orally gavaged with Blastocystis ST1 for two weeks (Fig. 1a). Blastocystis colonization was confirmed by SEM and qPCR (Fig. 1b, c). Clinical manifestations were assessed by evaluating weight changes and DAI. There were no significant changes in the weight and DAI of mice colonized with ST1 compared to non-colonized mice (Fig. 1d, e). Similarly, ST1-colonized mice did not show any colonic histological damage and the colon lengths were comparable to control mice (Fig. 1f). These data suggest that ST1 colonizes asymptomatically in normal healthy mice, which is similar to the effects of Blastocystis ST416.

a Experimental design. b Scanning electron microscopy of colon and cecum tissues from ST1-colonized and un-colonized mice, ST1 are indicated with a red asterisk (*). Scale bar = 1 μm. c Blastocystis ST1 cells per milligram of stool in ST1-colonized mice. Weight changes (d) and DAI (e) between control and ST1-colonized mice. f Representative micrographs of H&E-stained colon sections, colon length, and colonic histological scores from un-colonized and ST1-colonized mice. Scale bar = 100 μm. Data are shown as the mean ± SEM. Significance was determined by two-sided unpaired Student’s t test (ns non-significant).
Effect of Blastocystis ST1 colonization on gut bacterial composition
Next, we performed 16 S rRNA gene sequencing on fecal samples from ST1-colonized and control mice to determine the effect of Blastocystis ST1 colonization on the gut bacterial community. The rarefaction curves showed sufficient sequencing depth was obtained for further diversity analysis (Supplementary Fig. 1a). There were no significant changes in the alpha diversity based on Shannon, Pielou’s evenness, and Chao1 indices measurements, when compared to the baseline in both ST1-colonized and control mice (Fig. 2a). At the start of the experiment (day 0), PCoA plots showed ST1-colonized and control mice clustered together, suggesting those mice showed similar microbiota composition before Blastocystis ST1 colonization (P > 0.05, Fig. 2b and Supplementary Table 1). Notably, we observed significant changes in the gut microbial compositions in both ST1-colonized and control groups by PERMANOVA analysis (P < 0.05, Fig. 2b and Supplementary Table 1). The fluctuations observed in the control group could be attributed to the complex and dynamic nature of the gut microbiota, which is known to change over time20. At the phylum level, the abundance of Bacteroidota was elevated in both ST1-colonized and non-colonized groups, while the proportion of Firmicutes was found to be decreased in these groups (Fig. 2c). At the genus level, Alloprevotella and Akkermansia were significantly enriched in ST1-colonized mice (Fig. 2d). Alloprevotella are SCFA-producing bacteria and are positively correlated with the anti-inflammatory cytokine IL-1021,22. It has also been determined that the administration of Akkermansia muciniphila can ameliorate DSS-induced colitis in mice23, and increase mucus production to prevent intestinal inflammation24. In contrast, the level of Candidatus Saccharimonas and Bacteroides was significantly higher in control mice when compared to the baseline, and the commensal bacteria Anaeroplasma declined in un-colonized mice (Fig. 2d). Muribaculum, Alistipes, and Clostridia vadinBB60 group showed similar changes in both ST1-colonized and non-colonized mice (Supplementary Fig. 1b). Similarly, LefSe analysis also showed that the Bacteroides and Alloprevotella were enriched in non-colonized and ST1-colonized mice respectively (Supplementary Fig. 1c). Altogether, our data showed Blastocystis ST1 colonization could alter the gut microbiota composition and was positively associated with the beneficial bacteria Alloprevotella and Akkermansia.

a Alpha diversity was measured by Shannon, Pielou’s evenness, and Chao1 indices (n = 8 for each group). b PCoA of fecal gut microbiota in ST1-colonized and un-colonized mice. c Relative abundance of the top 5 phyla. d Heatmap of the 40 most abundant genera and those genera showing significant differences in Blastocystis-free and ST1-colonized mice. Data are presented as boxplots, with the center line representing the median, the boundary of the whiskers representing the minimum and maximum values of the dataset, and the boundary of the box representing the 25th and 75th percentile of the dataset (a, c, d). Significance was determined using the Wilcoxon rank-sum test. (ns non-significant; *P < 0.05; **P < 0.01; ***P < 0.001; and ****P < 0.0001).
Effect of Blastocystis ST1 colonization on colonic T helper cells
Host adaptive immune responses play a crucial role in maintaining intestinal homeostasis, and inappropriate T-cell responses against the microbiota are implicated in the pathogenesis of inflammatory bowel disease (IBD)25. We next investigated whether ST1 could also alter the functions of colonic T helper cells during colonization. Interestingly, the expansion of IL-4-producing CD4+ Th2 cells and IL-10-producing CD4+ Treg cells was observed in ST1-colonized mice (Fig. 3a, b). Forkhead box P3 (Foxp3) is a crucial regulator of the development and function of Treg cells26. We also observed an elevated proportion of Foxp3-expressing CD4+ T cells in the ST1-colonized mice (Fig. 3c, d). The accumulation of Th2 and Treg cells is commonly found in extracellular parasite infection, which plays a significant role in immunity, tissue repair, and tolerance of the parasite at tissue sites27. In contrast, the percentage of IFNγ-, and TNF-α-producing CD4+ Th1 cells and IL-17A-producing CD4+ Th17 cells did not expand in both ST1-colonized and control mice (Fig. 3a–d). These results were consistent with our previous report that Blastocystis ST4 colonization promotes Th2 and Treg cell differentiation to maintain immune homeostasis in normal healthy mice16.

a Colored contour plots show staining for IL-4, IL-10, and IL-17A within CD4+ cells. b Bar charts show the percentage of IL-4, IL-10, and IL-17A expressing CD4+ T cells. c Colored contour plots show staining for IFN-γ, TNF-α, and Foxp3 within CD4+ cells. d Bar charts show the percentage of IFN-γ, TNF-α, and Foxp3-expressing CD4+ T cells. Data are shown as the mean ± SEM. Significance was determined by two-sided unpaired Student’s t test (ns non-significant; *P < 0.05; **P < 0.01).
Effect of Blastocystis ST1 colonization on the severity of DSS-induced colitis
The gut microbiota is believed to contribute to the pathogenesis of several diseases, such as IBD28,29. We then sought to determine whether changes in gut microbiota induced by Blastocystis ST1 were correlated with susceptibility to experimentally-induced colitis. ST1-colonized mice were treated with 2% DSS for one week to develop colitis (Fig. 4a). Blastocystis ST1 colonization status was assessed using SEM and qPCR, which indicated the presence and the number of Blastocystis ST1 cells in the mouse intestine respectively (Fig. 4b, c). Notably, ST1-colonized mice showed lower severity of colitis with less weight loss and lower DAI than control mice (Fig. 4d, e). The results of the colon histological examinations indicated that un-colonized mice experienced significant crypt disruption and infiltration of inflammatory cells (Fig. 4f). Similarly, ST1-colonized mice exhibited significantly longer colon length and lower colonic histological damage scores than un-colonized mice (Fig. 4g, h). These results indicated that ST1 colonization exerts a protective effect on DSS-induced colitis.

a Experimental design. b Scanning electron microscopy of colon and cecum tissues from ST1-colonized and un-colonized mice, ST1 are indicated with a red asterisk (*). Scale bar = 1 μm. c Blastocystis ST1 cells per milligram of stool in ST1-colonized mice. Weight changes (d) and DAI (e) between ST1-colonized and un-colonized mice during DSS treated for 7 days. f Representative micrographs of H&E-stained colon sections from un-colonized and ST1-colonized mice. Colon length (g) and colonic histological (h) in un-colonized and ST1-colonized group. Scale bar = 100 μm. Data are shown as the mean ± SEM. Significance was determined by two-sided unpaired Student’s t test (*P < 0.05; **P < 0.01).
The protective effect against DSS-induced colitis associated with Th2 and Treg immune responses
Several studies have shown that infection with helminth prevents experimental colitis by inducing the accumulation of Th2 and Treg cells30,31. We next investigated whether the Th2 and Treg cells are involved in the protective effects of DSS-induced colitis. Colonization with Blastocystis ST1 increases the production of IL-4, IL-10, and the proportion of Foxp3+CD4+ T cells (Fig. 5a, b). In contrast, the control mice showed a substantial increase in the percentage of pro-inflammatory cytokine TNF-α-producing T cells (Fig. 5c, d). IL17-A-producing CD4+ T cells in the colonic mucosa were comparable between ST1-colonized and control mice (Fig. 5a, b). These data suggest that colonization with Blastocystis ST1 prevents experimental colitis associated with the accumulation of the Th2 and Treg cells.

a Colored contour plots show staining for IL-4, IL-10, and IL-17A within CD4+ cells. b Bar charts show the percentage of IL-4, IL-10, and IL-17A expressing CD4+ T cells. c Colored contour plots show staining for IFN-γ, TNF-α, and Foxp3 within CD4+ cells. d Bar charts show the percentage of IFN-γ, TNF-α, and Foxp3-expressing CD4+ T cells. Data are shown as the mean ± SEM. Significance was determined by two-sided unpaired Student’s t test (ns non-significant; *P < 0.05; **P < 0.01; ***P < 0.001).
Effect of Blastocystis ST1-altered gut microbiota on the severity of DSS-induced colitis
To determine whether Blastocystis ST1-altered gut microbiota can influence the susceptibility of DSS-induced colitis, the mice were treated with a cocktail of the antibiotic for two weeks and then were orally gavaged with Blastocystis-altered gut microbiota (Fig. 6a). The fecal samples used for FMT were taken from both ST1-colonized and control mice (Fig. 1a). To disrupt the live Blastocystis ST1 cells, the samples were subjected to a freeze-thaw process (Supplementary Fig. 2). Mice that received fecal transplants from the control donor treated with DSS showed progressive weight loss and higher DAI compared to ST1-altered gut microbiota recipient mice (Fig. 6b). SEM and feces culturing did not reveal Blastocystis cells in the caecum or colon, suggesting the protective effects are associated with ST1-altered gut microbiota (Fig. 6c). Examination of colon tissue from DSS treated recipient mice found that control mice exhibited a higher degree of inflammation and tissue damage than the ST1-altered gut microbiota recipient mice treated with DSS (Fig. 6d). A shorter colon length was observed in control mice when compared to ST1-altered gut microbiota recipient mice (Fig. 6e). These results suggest that transplantation of Blastocystis-altered gut microbiota could exert the similar protective effects on the severity of colitis.

a Experimental design. b Weight changes and DAI between mice with ST1-altered gut microbiota and control donor during DSS treated for 7 days. c SEM showed no Blastocystis cells in both mice with ST1-altered gut microbiota and control donor. Scale bar = 1 μm. d Representative micrographs of H&E-stained colon sections from mice with control donor and ST-1 altered gut microbiota. e Colon length and colon histology in mice with control donor and ST-1 altered gut microbiota. Scale bar = 100 μm. Data are shown as the mean ± SEM. Significance was determined by two-sided unpaired Student’s t test (*P < 0.05).
Effect of Blastocystis ST1-altered gut microbiota on T helper cells and SCFA production
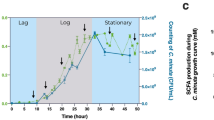
Since our previous study showed Blastocystis ST4-altered gut microbiota increased SCFA production and the number of CD4+ cells expressing IL-10 to suppress the DSS-induced colitis16, we sought to evaluate the effect of the ST1-altered gut microbiota on colonic T helper cells and SCFA production. Similarly, ST1-altered gut microbiota recipient mice showed a higher proportion of anti-inflammatory cytokine IL-10 and Foxp3+-Treg cells (Fig. 7a–d). In contrast, the pro-inflammatory cytokine IFN-γ and TNF-α-producing Th1 cells were increased significantly in control recipient mice (Fig. 7c, d). Interestingly, the levels of butyric, isobutyric, valeric acid and 2-methylbutyric acid in recipient mice of ST1-altered gut microbiota exhibited significantly higher concentrations compared to control mice (Fig. 7e). These findings suggest that the gut microbiota altered by Blastocystis ST1 have a beneficial effect on host health by regulating adaptive immune response and SCFA production.

a Colored contour plots show staining for IL-4, IL-10, and IL-17A within CD4+ cells. b Bar charts show the percentage of IL-4, IL-10, and IL-17A expressing CD4+ T cells. c Colored contour plots show staining for IFN-γ, TNF-α, and Foxp3 within CD4+ cells. d Bar charts show the percentage of IFN-γ, TNF-α, and Foxp3-expressing CD4+ T cells. e SCFA concentration in mice with ST1-altered gut microbiota and control donor. Data are shown as the mean ± SEM. Significance was determined by two-sided unpaired Student’s t test (ns non-significant; *P < 0.05; **P < 0.01).
Discussion
The associations between Blastocystis and gut microbiota have been investigated in several studies, but the pathogenicity of Blastocystis remains controversial4,32. Several studies report that Blastocystis colonization was associated with healthy gut microbiota and the organism was thus considered a commensal6,33,34. On the other hand, several studies reported that Blastocystis could cause dysbiosis and is associated with decreased cognitive function8,18. In the present study, we utilized a specific subtype (ST1), which is one of the most common subtypes found in humans. We observed that ST1 colonization was positively associated with the beneficial bacteria Alloprevotella and Akkermansia, and could prevent experimentally-induced colitis through the accumulation of Treg cells.
Alterations to the gut microbiota are implicated in the pathogenesis of IBD25. Alloprevotella is generally considered to be an SCFA producer, and the decreased abundance of Alloprevotella was correlated with reduced acetic acid and propionic acid concentrations in a mouse model21. Moreover, a positive correlation was observed between Alloprevotella and the anti-inflammatory cytokine IL-10 in mice with DSS-induced colitis, indicating a potential link to the protective effect against inflammation22. The increased level of Alloprevotella and other taxa is associated with reduced intestinal inflammation35. Multiple studies have shown that Akkermansia is associated with a healthy intestinal condition36,37,38. It has been determined that administration of Akkermansia muciniphila can ameliorate experimentally caused colitis23, and a lower proportion of Akkermansia has been identified in both fecal samples and mucosal biopsies of ulcerative colitis patients39,40. A. muciniphila can adhere to human colonic cell lines (Caco-2 and HT-29) and enhance barrier function in vitro, suggesting its ability to strengthen the compromised intestinal barrier41. Besides, A. muciniphila can also induce T-cell immune responses during homeostasis42, and increased abundance was observed in time-restricted feeding rats43. Although a previous study showed ST4 colonization increases Akkermansia level7, this is the first study to show a causal relationship between Blastocystis colonization and Akkermansia levels.
Previous studies had indicated that Blastocystis ST1 is involved in host innate immune responses. For example, co-incubation of ST1 with the cell lines HT-29 and T-84 increased the release of IL-8 and GM-CSF44. ST1 can also induce intestinal epithelial cells to secrete antimicrobial peptides (LL-37)45. The present study is the first study to show that Blastocystis ST1 can activate Th2 and Treg cell immune responses to prevent mice from DSS-induced colitis. Th2 cells are involved in type 2 immune responses, which are important for the eradication of extracellular parasites and bacterial infection46. Th2 cells were shown to suppress INF-γ-producing Th1 cells to inhibit pro-inflammatory immune responses47. Gut microbiota has been shown to induce intestinal Foxp3+ Treg cells and to enhance the expression of IL-10 in Treg cells48. IL-10 can suppress the secretion of pro-inflammatory cytokines, such as TNFα, to improve colonic inflammation in the DSS-induced colitis model49. IL-10 knockout mice are prone to develop spontaneous intestinal colitis, suggesting it is an essential immunoregulator in the intestinal tract50. Secretion of the anti-inflammatory cytokine IL-10 can suppress Th1 and Th17 effector functions, both of which are key mediators of IBD51.
Blastocystis ST1 strains exhibit extensive genetic variations, suggesting that this subtype may exert strain-dependent differential effects on host health52. For example, Blastocystis ST1 was regarded as a pathogenic subtype since it was significantly associated with IBS Diarrhea (IBS-D) in Indonesian adolescents53. Blastocystis ST1 was also the most predominant subtype in colorectal carcinoma (CRC) patients and has a strong significant association risk with CRC54. Besides, ST1 isolated from a symptomatic patient led to lethargy and intestinal inflammation in rats55. On the other hand, ST1 isolated from asymptomatic human donors did not result in any clinical symptoms and can colonize asymptomatically for more than one year56, and numerous studies revealed that ST1 mainly colonized healthy individuals19. Our data showed that ST1 isolated from a healthy individual during health screening in NUH did not cause any pathology in mice colon tissues, and instead, ST1 colonization played a protective role in the development of DSS-induced colitis. However, the effects of ST1 on the host should be expanded to different ST1 isolates to better explain its exact effects on host health.
While our study demonstrated that colonization with Blastocystis ST1 led to significant changes in the gut microbiota and altered host susceptibility to DSS-induced colitis, the precise mechanism behind this effect is still unclear. The effect may be mediated through direct interaction between ST1 and the gut bacteria/archaea, leading to changes in the microbial community structure and function. Alternatively, ST1 could be producing metabolites that affect the gut microbiota and host immune system. Future studies using targeted metagenomic and metabolomic analyses could help shed light on the specific mechanisms underlying the effect of ST1 on the gut microbiota and host health.
The germ-free mouse model is an important tool for understanding the complex interactions between microorganisms and host health, and has been used to investigate the role of microorganisms in various diseases, including IBD57. Using this mouse model would eliminate the confounding factors associated with the presence of other microorganisms in the gut, providing a clear picture of the impact of Blastocystis ST1 on the gut microbiota and the host immune response. Besides, adding a non-Blastocystis colonized control microbiota along with Blastocystis to see if the effect on the host could be “rescued,” could allow for a more targeted investigation of the role of Blastocystis on the host.
In conclusion, we report that Blastocystis ST1 colonization in a mouse model was associated with a higher proportion of the beneficial bacteria Alloprevotella and Akkermansia, and induced protective immune responses. Furthermore, ST1-colonized mice and recipient mice of ST1-altered gut microbiota were refractory to DSS-induced colitis via increasing the anti-inflammatory cytokine IL-10 and SCFA production, indicating that Blastocystis-mediates healthy gut physiology through positive alteration in the microbiota bacterial flora. A summary of these findings is illustrated in Fig. 8.

Blastocystis ST1 alters intestinal microbiota composition resulting in increases of beneficial bacteria Alloprevotella and Akkermansia. These species are associated with modulation of intestinal inflammation through Th2 and Treg immune responses, which suppress Th1-associated inflammation. ST1 mediates increases in SCFA in colonized mice, through increases in SCFA producers, which also play a role in suppressing inflammation through Treg responses.
Methods
Culture of Blastocystis
ST1-NUH9 was originally isolated in 2007 from a patient undergoing routine screening at the National University Hospital58. Permission from the National Healthcare Group Institutional Review Board was given before project commencement. Axenization was performed to obtain purified Blastocystis isolates59. Briefly, ST1-NUH9 isolate was cultured in Jones’ medium under aerobic conditions for 7 days, and then inoculated into pre-reduced Iscove’s Modified Dulbecco’s Media (IMDM) supplemented with 10% horse serum (Gibco) and antibiotics (2000 μg/ml claforan, 500 μg/ml ampicillin, 100 μg/ml streptomycin, and 100 UI/ml penicillin). Cultures were subcultured every 7 days until Blastocystis cells equaled or outnumbered bacterial ones. Blastocystis was then grown in 3.6% Bacto Agar, and Blastocystis colonies differentiated from bacterial ones by microscopy. Blastocystis colonies were isolated and transferred into pre-reduced IMDM supplemented with horse serum and antibiotics to obtain purified Blastocystis cells, using a Pasteur pipette. Axenized Blastocystis ST1 was maintained in 10 ml of pre-reduced IMDM supplemented with 10% heat-inactivated horse serum. Cultures were incubated under anaerobic conditions in an AnaeroJar (Oxoid) with a gas pack (Oxoid) at 37 °C. The number of Blastocystis cells was manually calculated using a hemocytometer (Kova International). Before each experiment in which it was included as an inoculum, the Blastocystis ST1 isolate was assessed for purity through microscopy. For quality control, supernatants from Blastocystis cultures were plated on Trypticase soy agar with defibrinated sheep blood (BD) to observe for bacteria colonies.
Mice and treatments
C57BL/6 mice, aged 8–12 weeks, were maintained in isolated ventilated IsoCages in the animal facility of the National University of Singapore (NUS). Littermates of the same sex and age were randomly assigned to different experimental groups. Mice were orally gavaged with 5 × 107 live Blastocystis ST1 cells16. In total, 2% DSS (MP Biomedicals) in drinking water was administrated to mice to induce acute colitis. Mice were administrated with a broad-spectrum antibiotic cocktail (0.5 g/L of vancomycin and 1 g/L of ampicillin, neomycin, and metronidazole), before the microbiota transplant experiments60. The disease activity index (DAI), assessed by weight changes, stool consistency, and the presence of fecal blood, was used to determine the severity of colitis61. All animal experiments were performed under the Singapore National Advisory Committee for Laboratory Animal Research guidelines and approved by the Institutional Animal Care and Use Committee of NUS (R19-1259).
Fecal microbiota transplantation
Fresh fecal samples were collected from ST1-colonized and non-colonized mice and frozen at –80 °C for 1 week. The live Blastocystis ST1 cells were ruptured by freeze-thaw to exclude their effect on experimentally caused colitis. The presence or absence of live Blastocystis cells was determined by culturing the feces in Jones’ medium (Supplementary Fig. 2). Fecal microbiota transplantation (FMT) was performed by diluting frozen feces with pre-reduced PBS (50 mg/ml) and administering the mixture to recipient mice via oral gavage (10 mg/mice) three times a week62.
Scanning electron microscopy
Scanning electron microscopy (SEM) was performed to determine the colonization status of Blastocystis ST163. The cecum/colon tissues were cut longitudinally without disturbing the intestinal contents. The opened tissues were pinned down to a silicone mat and fixed in 2.5% glutaraldehyde at 4 °C overnight. The tissues were washed with PBS and processed by post-fixing in 1% osmium tetroxide for 1 h, followed by dehydration with increasing concentrations of ethanol and critical-point drying. The dried samples were coated with 25 nm gold and imaged on a field emission JSM-6701F SEM at a voltage of 10 kV.
DNA extraction and real-time quantitative PCR
Genome DNA from fecal samples was extracted using QIAamp Fast DNA Stool Mini Kit (Qiagen) according to the manufacturer’s protocol. Real-time quantitative PCR (qPCR) was used to estimate the number of Blastocystis cells in fecal samples64. The extracted DNA was combined with a mixture of 5 µl of SsoAdvanced Universal SYBR Green Supermix, along with 0.5 μM forward primer BL18SPPF1 (5′- AGTAGTCATACGCTCGTCTCAAA-3′) and 0.5 μM reverse primer BL18SR2PP (5′-TCTTCGTTACCCGTTACTGC-3′) specific for the SSU rRNA gene of Blastocystis. The quantification of Blastocystis was carried out on an ABI 7500 real-time PCR system instrument (Life Technologies) with the following thermal cycling conditions: initial denaturation at 95 °C for 5 min, followed by 45 cycles of 95 °C for 15 s, 68 °C for 10 s, and 72 °C for 15 s. ST1-NUH9 cultures and IMDM medium were used as positive and negative control, respectively.
Colon histology
Colonic tissues were collected at the end of experiments and fixed in 4% neutral buffered formalin, and then processed and embedded in paraffin. Sections of 4.5 μm were prepared and stained with hematoxylin and eosin (H&E). Histology scoring was performed in a blinded fashion, whereby changes in intestinal crypt architecture, degree of inflammation, and epithelium damage were scored65.
16 S rRNA gene sequencing and bioinformatics analysis
The 16 S rRNA gene sequencing library was constructed using a MetaVX Library Preparation Kit (South Plainfield, NJ). Briefly, 20–30 ng of DNA was used to generate amplicons that cover V3 and V4 hypervariable regions of the 16 S rRNA gene of bacteria. The forward primer contains the sequence ‘CCTACGGRRBGCASCAGKVRVGAAT’, and the reverse primers contain the sequence ‘GGACTACNVGGGTWTCTAATCC’. The 25 µl PCR mixture was prepared with 2.5 µl TransStart buffer, 2 µl dNTPs, 1 µl of each primer, 0.5 µl TransStart Taq DNA polymerase, and 20 ng template DNA. The PCR was performed with the following program: 3 min of denaturation at 94 °C, 24 cycles of 5 s at 95 °C, 90 s of annealing at 57 °C, 10 s of elongation at 72 °C, and a final extension at 72 °C for 5 min. Indexed adapters were added to the ends of the amplicons by limited cycle PCR. Finally, the library was purified using magnetic beads. DNA concentration was determined with a microplate reader (Tecan, Infinite 200 Pro) and the fragment size was determined by 1.5% agarose gel electrophoresis, which was expected at ~600 bp. Next-generation sequencing was conducted on an Illumina Novaseq Platform (Illumina, San Diego, USA) at the laboratory. Automated cluster generation and 250 paired-end sequencing with dual reads were performed according to the manufacturer’s instructions.
Paired-end sequences of positive and negative reads were filtered, denoised, and chimeras removed to obtain amplicon sequence variants (ASVs) through DADA2 using the Quantitative Insights into Microbial Ecology 2 (Qiime2) plugin66. The Silva 138 database was used for the taxonomic analysis of the representative ASV sequences67. Shannon and Chao1 were used to estimate the bacterial diversity and richness respectively. Beta diversity was assessed by permutational multivariate analysis of variance (PERMANOVA). Principal coordinates analysis (PCoA) plots were constructed based on Bray–Curtis dissimilarity to illustrate the differences in community structure between different groups. Heatmaps were used to show the different taxa between groups. Linear discriminant analysis (LDA) effect size (LEfSe) analysis was performed to detect bacterial taxa with significantly different abundance among different groups with P value <0.05 and LDA score >4 (https://huttenhower.sph.harvard.edu/galaxy/).
Isolation of colon lamina propria cells
To analyze colonic lymphocytes, the colon tissues were longitudinally opened and washed with ice-cold PBS to remove luminal contents. The tissues were cut into 1 cm pieces and incubated in Roswell Park Memorial Institute (RPMI) 1640 medium (Sigma-Aldrich) containing 1 mM EDTA (Sigma-Aldrich) and 1 mM DTT (Sigma-Aldrich) at room temperature for 20 min under slow rotation and spun down to remove the supernatant. The remaining tissue pieces were incubated in RPMI containing 25% HEPES, 10% fetal calf serum (FCS), 1 mM EDTA, and 1 mM DTT at 37 °C for 1 h under slow rotation and then washed by PBS to remove epithelial cells and intraepithelial lymphocytes. Tissue pieces were digested with Liberase (Sigma-Aldrich) at 37 °C for 30 min under slow rotation. The digested tissue pieces and supernatants were filtered with 70-μm cell strainers and glass wool separately. After centrifugation, pellets containing the lamina propria (LP) lymphocytes were harvested.
Flow cytometry analysis
Lymphocytes were stimulated for 6 h with a cell stimulation cocktail of PMA (50 ng/ml), ionomycin (750 ng/ml), and GolgiStop (monensin, BD Biosciences). Live/dead stain was used to evaluate the viability of the cells using live/dead fixable viability stain kits. For surface staining, stimulated cells were stained with TCR beta monoclonal antibody (BUV510, eBioscience), and anti-CD4 (BUV395; Biolegend). Fixation and permeabilization buffers (Biolegend) were used for intracellular cytokine staining. Fixed and permeabilized cells were stained with fluorochrome-conjugated anti-mouse antibodies against IL-4 (BUV421; Biolegend), IL-10 (PE; Biolegend), IL-17A (APC; eBioscience), interferon-gamma (IFN-γ) (BUV711; Biolegend), tumor necrosis factor (TNF-α) (APC; eBioscience), and Foxp3 (PE, Biolegend) at 4 °C for 10 h. Flow cytometry analysis was performed on a Fortessa X-20 (BD biosciences) flow cytometer and the data were analyzed using FlowJo V10 software. Gating strategy of immune populations in the colonic LP was shown in Supplementary Fig. 3.
LC/MS/MS assay
Liquid chromatography/tandem mass spectrometry (LC/MS/MS) was performed for analysis of short-chain fatty acid (SCFA) in derivatized stool extracts68. Briefly, 500 μl of ice-cold extraction solvent containing 10 μM of d5-benzoic acid as internal standard (IS) was added to 250 mg of stool samples. The mixture was then spun down, and the supernatant was discarded. Subsequently, an aliquot of 100 μl was derivatized using aniline and EDC. The derivatization reaction was quenched using succinic acid and 2-mercaptoethanol. All samples were saved at 4 °C until analysis on the same day. Analysis was carried out using an Agilent 1290 Infinity LC system (Agilent Technologies, Santa Clara, CA, USA).
Statistical analysis
Statistical analysis was performed in Prism 8 (Graphpad Software Inc.). Two independent replicates were performed for each experiment. A two-sided unpaired Student’s t test was used for comparisons of two groups. Error bars on graphs display the mean and SEM. P values of <0.05 were considered significant; the following symbols were used to indicate significance levels: ns, non-significant; *P < 0.05; **P < 0.01; ***P < 0.001; and ****P < 0.0001.
Data availability
The datasets generated and analyzed in the current study are available in the Sequence Read Archive (SRA) database at NCBI under BioProject ID PRJNA891885. All other data are available from the corresponding author upon request.
References
Rowan-Nash, A. D., Korry, B. J., Mylonakis, E. & Belenky, P. Cross-domain and viral interactions in the microbiome. Microbiol. Mol. Biol. Rev. 83, e00044–18 (2019).
Lukeš, J., Stensvold, C. R., Jirků-Pomajbíková, K. & Wegener Parfrey, L. Are human intestinal eukaryotes beneficial or commensals? PLoS Pathog. 11, e1005039 (2015).
Stensvold, C. R., Tan, K. S. W. & Clark, C. G. Blastocystis. Trends Parasitol. 36, 315–316 (2020).
Stensvold, C. R. & van der Giezen, M. Associations between gut microbiota and common luminal intestinal parasites. Trends Parasitol. 34, 369–377 (2018).
Chabé, M., Lokmer, A. & Ségurel, L. Gut protozoa: friends or foes of the human gut microbiota? Trends Parasitol. 33, 925–934 (2017).
Audebert, C. et al. Colonization with the enteric protozoa Blastocystis is associated with increased diversity of human gut bacterial microbiota. Sci. Rep. 6, 25255 (2016).
Tito, R. Y. et al. Population-level analysis of Blastocystis subtype prevalence and variation in the human gut microbiota. Gut 68, 1180–1189 (2019).
Mayneris-Perxachs, J. et al. Presence of Blastocystis in gut microbiota is associated with cognitive traits and decreased executive function. ISME J. 16, 2181–2197 (2022).
Defaye, M. et al. Fecal dysbiosis associated with colonic hypersensitivity and behavioral alterations in chronically Blastocystis-infected rats. Sci. Rep. 10, 9146 (2020).
Deng, L. et al. Colonization with two different Blastocystis subtypes in DSS-induced colitis mice is associated with strikingly different microbiome and pathological features. Theranostics 13, 1165–1179 (2023).
Maloney, J. G. et al. Identification and molecular characterization of four new Blastocystis subtypes designated ST35-ST38. Microorganisms 11, 46 (2022).
Andersen, L. O. & Stensvold, C. R. Blastocystis in health and disease: are we moving from a clinical to a public health perspective? J. Clin. Microbiol. 54, 524–528 (2016).
Gentekaki, E. et al. Extreme genome diversity in the hyper-prevalent parasitic eukaryote Blastocystis. PLoS Biol. 15, e2003769 (2017).
Lim, M. X. et al. Differential regulation of proinflammatory cytokine expression by mitogen-activated protein kinases in macrophages in response to intestinal parasite infection. Infect. Immun. 82, 4789–4801 (2014).
Depommier, C. et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat. Med. 25, 1096–1103 (2019).
Deng, L. et al. Experimental colonization with Blastocystis ST4 is associated with protective immune responses and modulation of gut microbiome in a DSS-induced colitis mouse model. Cell. Mol. Life. Sci. 79, 245 (2022).
Billy, V. et al. Blastocystis colonization alters the gut microbiome and, in some cases, promotes faster recovery from induced colitis. Front. Microbiol. 12, 641483 (2021).
Yason, J. A., Liang, Y. R., Png, C. W., Zhang, Y. & Tan, K. S. W. Interactions between a pathogenic Blastocystis subtype and gut microbiota: in vitro and in vivo studies. Microbiome 7, 30 (2019).
Popruk, S., Adao, D. E. V. & Rivera, W. L. Epidemiology and subtype distribution of Blastocystis in humans: a review. Infect. Gene. Evol. 95, 105085 (2021).
Langille, M. G. et al. Microbial shifts in the aging mouse gut. Microbiome 2, 50 (2014).
Qu, W. et al. Dietary advanced glycation end products modify gut microbial composition and partially increase colon permeability in rats. Mol. Nutr. Food. Res. 61, 1700118 (2017).
Li, A. L. et al. Effect of cinnamon essential oil on gut microbiota in the mouse model of dextran sodium sulfate-induced colitis. Microbiol. Immunol. 64, 23–32 (2020).
Bian, X. et al. Administration of Akkermansia muciniphila ameliorates dextran sulfate sodium-induced ulcerative colitis in mice. Front. Microbiol. 10, 2259 (2019).
Everard, A. et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 110, 9066–9071 (2013).
Belkaid, Y. & Hand, T. W. Role of the microbiota in immunity and inflammation. Cell 157, 121–141 (2014).
Lu, L., Barbi, J. & Pan, F. The regulation of immune tolerance by FOXP3. Nat. Rev. Immunol. 17, 703–717 (2017).
Bouchery, T., Kyle, R., Ronchese, F. & Le G.ros, G. The differentiation of CD4(+) T-helper cell subsets in the context of helminth parasite infection. Front. Immunol. 5, 487 (2014).
Caruso, R., Lo, B. C. & Núñez, G. Host-microbiota interactions in inflammatory bowel disease. Nat. Rev. Immunol. 20, 411–426 (2020).
Anand, S. & Mande, S. S. Host-microbiome interactions: Gut-Liver axis and its connection with other organs. NPJ Biofilms Microbiomes. 8, 89 (2022).
Kang, S. A. et al. Adoptive transfer of Trichinella spiralis-activated macrophages can ameliorate both Th1- and Th2-activated inflammation in murine models. Sci. Rep. 9, 6547 (2019).
Shan, W. et al. Schistosoma japonicum peptide SJMHE1 inhibits acute and chronic colitis induced by dextran sulfate sodium in mice. Parasit Vectors 14, 455 (2021).
Deng, L., Wojciech, L., Gascoigne, N. R. J., Peng, G. & Tan, K. S. W. New insights into the interactions between Blastocystis, the gut microbiota, and host immunity. PLoS Pathog. 17, e1009253 (2021).
Beghini, F. et al. Large-scale comparative metagenomics of Blastocystis, a common member of the human gut microbiome. ISME J. 11, 2848–2863 (2017).
Stensvold, C. R. et al. Stool microbiota diversity analysis of Blastocystis-positive and Blastocystis-negative individuals. Microorganisms 10, 326 (2022).
Cui, H. X., Hu, Y. N., Li, J. W. & Yuan, K. Hypoglycemic mechanism of the berberine organic acid salt under the synergistic effect of intestinal flora and oxidative stress. Oxid. Med. Cel. Longev. 2018, 8930374 (2018).
Belzer, C. & de Vos, W. M. Microbes inside–from diversity to function: the case of Akkermansia. ISME J. 6, 1449–1458 (2012).
Bae, M. et al. Akkermansia muciniphila phospholipid induces homeostatic immune responses. Nature 608, 168–173 (2022).
Luo, Y. et al. Rational consideration of Akkermansia muciniphila targeting intestinal health: advantages and challenges. NPJ Biofilms Microbiomes. 8, 81 (2022).
James, S. L. et al. Abnormal fibre usage in UC in remission. Gut 64, 562–570 (2015).
Png, C. W. et al. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am. J. Gastroenterol. 105, 2420–2428 (2010).
Reunanen, J. et al. Akkermansia muciniphila adheres to enterocytes and strengthens the integrity of the epithelial cell layer. Appl. Environ. Microbiol. 81, 3655–3662 (2015).
Ansaldo, E. et al. Akkermansia muciniphila induces intestinal adaptive immune responses during homeostasis. Science 364, 1179–1184 (2019).
Palomba, A. et al. Time-restricted feeding induces Lactobacillus- and Akkermansia-specific functional changes in the rat fecal microbiota. NPJ Biofilms Microbiomes. 7, 85 (2021).
Long, H. Y., Handschack, A., König, W. & Ambrosch, A. Blastocystis hominis modulates immune responses and cytokine release in colonic epithelial cells. Parasitol. Res. 87, 1029–1030 (2001).
Yason, J. A., Ajjampur, S. S. R. & Tan, K. S. W. Blastocystis isolate B exhibits multiple modes of resistance against antimicrobial peptide LL-37. Infect. Immun. 84, 2220–2232 (2016).
Paul, W. E. & Zhu, J. How are TH2-type immune responses initiated and amplified? Nat. Rev. Immunol 10, 225–235 (2010).
Butcher, M. J. & Zhu, J. Recent advances in understanding the Th1/Th2 effector choice. Faculty Rev. 10, 30 (2021).
Hagihara, Y. et al. Epigenetic regulation of T helper cells and intestinal pathogenicity. Semin. Immunopathol. 41, 379–399 (2019).
Tomoyose, M., Mitsuyama, K., Ishida, H., Toyonaga, A. & Tanikawa, K. Role of interleukin-10 in a murine model of dextran sulfate sodium-induced colitis. Scan. J. Gastroenterol. 33, 435–440 (1998).
Kühn, R., Löhler, J., Rennick, D., Rajewsky, K. & Müller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 75, 263–274 (1993).
Clough, J. N., Omer, O. S., Tasker, S., Lord, G. M. & Irving, P. M. Regulatory T-cell therapy in Crohn’s disease: challenges and advances. Gut 69, 942–952 (2020).
Wakid, M. H., Aldahhasi, W. T., Alsulami, M. N., El-Kady, A. M. & Elshabrawy, H. A. Identification and genetic characterization of Blastocystis species in patients from Makkah, Saudi Arabia. Infect. Drug. Resist. 15, 491–501 (2022).
Kesuma, Y., Firmansyah, A., Bardosono, S., Sari, I. P. & Kurniawan, A. Blastocystis ST-1 is associated with irritable bowel syndrome-diarrhoea (IBS-D) in Indonesian adolescences. Parasite Epidemiol. Cont. 6, e00112 (2019).
Mohamed, A. M. et al. Predominance and association risk of Blastocystis hominis subtype I in colorectal cancer: a case control study. Infect. Agent. Cancer 12, 21 (2017).
Li, J. et al. A rat model to study Blastocytis subtype 1 infections. Parasitol. Res. 112, 3537–3541 (2013).
Růžková, J. et al. Evaluating rodent experimental models for studies of Blastocystis ST1. Exp. Parasitol. 191, 55–61 (2018).
Grover, M. & Kashyap, P. C. Germ-free mice as a model to study effect of gut microbiota on host physiology. Neurogastroenterol. Motil. 26, 745–748 (2014).
Wong, K. H. et al. Predominance of subtype 3 among Blastocystis isolates from a major hospital in Singapore. Parasitol Res 102, 663–670 (2008).
Ng, G. C. & Tan, K. S. Colony growth as a step towards axenization of Blastocystis isolates. Parasitol. Res. 85, 678–679 (1999).
Belda, E. et al. Impairment of gut microbial biotin metabolism and host biotin status in severe obesity: effect of biotin and prebiotic supplementation on improved metabolism. Gut 71, 2463–2480 (2022).
Cooper, H. S., Murthy, S. N., Shah, R. S. & Sedergran, D. J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 69, 238–249 (1993).
Burrello, C. et al. Therapeutic faecal microbiota transplantation controls intestinal inflammation through IL10 secretion by immune cells. Nat. Commun. 9, 5184 (2018).
Mishra, A. et al. Microbial exposure during early human development primes fetal immune cells. Cell 184, 3394–3409.e3320 (2021).
Poirier, P. et al. Development and evaluation of a real-time PCR assay for detection and quantification of Blastocystis parasites in human stool samples: prospective study of patients with hematological malignancies. J. Clin. Microbiol. 49, 975–983 (2011).
Melgar, S. et al. Validation of murine dextran sulfate sodium-induced colitis using four therapeutic agents for human inflammatory bowel disease. Intern. Immunopharmacol. 8, 836–844 (2008).
Callahan, B. J. et al. DADA2: high-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583 (2016).
Quast, C. et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids. Res. 41, D590–D596 (2013).
Chan, J. C., Kioh, D. Y., Yap, G. C., Lee, B. W. & Chan, E. C. A novel LCMSMS method for quantitative measurement of short-chain fatty acids in human stool derivatized with (12)C- and (13)C-labelled aniline. J. Pharm. Biomed. Anal. 138, 43–53 (2017).
Acknowledgements
We thank Dr. Thet Tun Aung and Dr. Benoit Malleret for the help in scanning electron microscopy experiments. We would like to thank Chua Yen Leong for assistance in animal work. This study was supported by the Singapore Ministry of Education NUHS seed grants T1-NUHS Joint Grant Call FY17-1st call-03 (to K.S.W.T.), NUHSRO/2019/049/T1/SEED-MAR/02 (to NRJG), and R-148–000-277–114 (to E.C.Y.C.). KSWT also acknowledges the Blastocystis Under One Health Grant (A-8000685-00-00) for supporting this study. L.W. was the recipient of an NUSMed Postdoctoral Fellowship. The images in Fig. 1a, Fig. 4a, Fig. 6a, and Fig. 8 were created using BioRender.
Author information
Authors and Affiliations
Contributions
L.D. performed experiments, analyzed and interpreted the data. L.W. and L.D. performed immunological experiments. C.W.P. performed histological analyses. L.W. and L.D. performed immunological experiments and analyses. Y.Q.D.K. and E.C.Y.C. performed LC/MS/MS assays. Y.Z., N.R.J.G., and K.S.W.T. designed, conceived, and supervised the study.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Deng, L., Wojciech, L., Png, C.W. et al. Colonization with ubiquitous protist Blastocystis ST1 ameliorates DSS-induced colitis and promotes beneficial microbiota and immune outcomes. npj Biofilms Microbiomes 9, 22 (2023). https://doi.org/10.1038/s41522-023-00389-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41522-023-00389-1
This article is cited by
-
Effect of minocycline, methyl prednisolone, or combination treatment on the colonic bacterial population in a state of colonic inflammation using the murine dextran sulfate sodium model
Microbial Cell Factories (2023)
-
Prevalence of Blastocystis and Dientamoeba fragilis in diarrheal patients in Corum, Türkiye
Parasitology Research (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
