


Introduction
The foraging ecology and choice of feeding habitat of New Zealand’s rarest breeding bird (Hansen Reference Hansen2005), the New Zealand Fairy Tern (hereafter: NZFT) Sternula nereis davisae (Gould Reference Gould1843), have not been described to date. With a current breeding population of only nine pairs (four-year average 2008/2009–2011/2012; DoC internal reports) at only four known breeding sites (summary of Ferreira et al. Reference Ferreira, Hansen, Parrish, Pierce, Pulham and Taylor2005, Lagnaz Reference Lagnaz2010, Meagher Reference Meagher2010, Smith Reference Smith2010, Zimmerman Reference Zimmerman2010), the New Zealand taxon is currently ranked as nationally Critically Threatened (Miskelly et al. Reference Miskelly, Dowding, Elliot, Hitchmough, Powlesland, Robertson, Sagar, Scofield and Taylor2008); with the species classified globally as “Vulnerable” (IUCN 2011). In spite of intensive protection, breeding monitoring and egg and chick management by the New Zealand Department of Conservation (Parrish and Honnor Reference Parrish and Honnor1997, Hansen Reference Hansen2005) and community volunteer efforts, the NZFT still faces an extinction risk of 39% over the next 50 years (Ferreira et al. Reference Ferreira, Hansen, Parrish, Pierce, Pulham and Taylor2005).
While the factors identifying suitable breeding habitat have recently been assessed (Brooks et al. Reference Brooks, Davis, Baird, Bellingham and Zimmerman2011), and feeding observations at nest sites indicate the potentially crucial prey spectrum during chick rearing (Parrish and Pulham Reference Parrish and Pulham1995a), no data on foraging habitat choice and dependency, or foraging behaviour of the NZFT have been published to date.
Reproductive success in small terns has been shown to depend heavily on proximity of productive foraging areas (Perrow et al. Reference Perrow, Gilroy, Skeate and Tomlinson2011a, for Little Tern Sternula albifrons), and availability of high quality prey for chick provisioning (Dänhardt et al. Reference Dänhardt, Fresemann and Becker2011, for Common Tern Sterna hirundo). The conspecific Australian Fairy Tern (AFT) S. n. nereis has been reported to rarely travel beyond 2 km for foraging during the breeding season (Paton and Rogers Reference Paton and Rogers2009). Investigation of the feeding ecology of the NZFT is warranted to guide long-term conservation actions for the taxon.
We conducted a field study at Mangawhai Harbour, Northland, New Zealand during the 2010/2011 chick-rearing period (November–January) to address this gap in knowledge. We assessed the foraging ecology of the largest remaining breeding population at Mangawhai Harbour, where five out of the 10 breeding pairs nested on the neighbouring sand spit in the 2010/2011 season (DoC internal reports). Our study combined a specifically designed visual tracking regime that collated 140 hours of direct field observations to establish NZFT foraging habitat use, with sampling by seine-net and analyses of potential prey items from 10 identified NZFT feeding sites to describe the potential NZFT prey spectrum. Stable isotope analyses offer a non-invasive tool to assess dietary composition, circumventing disruption of nest sites, as would be risked by collection of pellets and dropped prey items. We therefore used stable isotope analyses of 52 samples of feathers as well as potential prey tissues, to assess the importance of the most common estuarine fish and shrimps in NZFT diet.
Materials and methods
Study species and study area
The subspecies S. n. davisae (Mathews and Iredale Reference Mathews and Iredale1913) of Sternula nereis (Gould Reference Gould1843) only breeds in New Zealand, and is genetically distinct from its Australian and New Caledonian conspecifics (Baling and Brunton Reference Baling and Brunton2005). In contrast with both the Australian subspecies S. n. nereis and the New Caledonian subspecies S. n. exsul, which form breeding colonies (Bransbury Reference Bransbury1992, Carter and Mustoe Reference Carter and Mustoe2007, Baling et al. Reference Baling, Jeffries, Barre and Brunton2009), breeding behaviour under current ecological conditions suggests the NZFT is a solitary breeder (Falla et al. Reference Falla, Sibson and Turbott1979) that defends its breeding territory against conspecifics (Parrish and Pulham Reference Parrish and Pulham1995a). NZFT winter in Kaipara Harbour, New Zealand (Goffin Reference Goffin1978, Chamberlin and Dowding Reference Chamberlin and Dowding1985, Parrish and Pulham Reference Parrish and Pulham1995b). Immature and non-breeding birds (Pulham, Habraken, Vaughan, Riegen, unpubl. data, Ornithological Society of New Zealand and New Zealand Wader Study Group) and failed east coast breeding pairs (Reference Baird, Ismar, Wilson, Plowman, Zimmerman and BellinghamBaird et al. in press) have also been found to feed and roost in this harbour during the austral summer months.
The breeding season of the NZFT at its four current breeding locations at Waipu (35°59’S, 174°29’E), Mangawhai (36°06’S, 174°36’E), Pakiri (36°15’S, 174°44’E), and Papakanui (36°26’S, 174°12’E) on New Zealand’s North Island (Figure 1) spans from September, when birds have returned to their breeding locations, to an extended post-fledging tuition period in February–March (Preddey Reference Preddey2008) in the vicinity of their breeding sites. The nesting period typically encompasses October to January (Shaw Reference Shaw and Robertson1985). NZFT nest on vegetation-free areas of sand and shell above the mean high-water mark (Parrish and Pulham Reference Parrish and Pulham1995a). Three of the above breeding locations are prominent sand spits, the fourth, Pakiri, is on a small sand spit adjacent to a river mouth. All are characterised by an estuary on one side, and coastal shallows on the other. Mangrove Avicennia marina var. resinifera vegetation occurs in parts of all four estuaries and river inlets. At Mangawhai Harbour, a breach of the sand spit was closed by constructing a bund through the historic estuary channel in June 1996, and this created an oxbow lagoon. This lagoon was replenished by spring tides from the coastal side until 2009.

Figure 1. Breeding range of the New Zealand Fairy Tern Sternula nereis davisae; yellow symbols: the four remaining breeding locations at Waipu, Mangawhai, Pakiri and Papakanui Spit; inset: NZFT feeding a goby Favonigobius sp. to its chick, Mangawhai 2008/2009 breeding season (Photo credit: Ian Southey; satellite images: Data SIO, NOAA, U.S. Navy, NGA, GEBKO, 2010 DigitalGlobe, TerraMetrics, GoogleTMEarth).
Assessment of foraging habitat use
Observations of NZFT foraging activity were collated over eight sampling events during the 2010/2011 nesting season. Sampling took place on 16, 17 and 30 Nov, on 1 and 17 Dec 2010, and on 6, 7, and 20 Jan 2011; all observation sessions lasted four hours and were timed during daylight hours spanning two hours either side of low tide. This time covered the peak NZFT foraging activity and provisioning at nests previously seen by wardens at nest sites. Given the critically endangered status of the NZFT in New Zealand, deploying biologging devices to record foraging behaviour could not be considered. Visual tracking constitutes an effective means of recording tern behaviour from the nest sites to the foraging grounds and back to the nests (Perrow et al. Reference Perrow, Skeate and Gilroy2011b).
The local topography allowed us to design a cohesive observation regime (Figure 2a), monitoring the range of potential NZFT foraging habitats. Strategic observer positions were chosen that gave clear views of the mid-estuary, the lower harbour, the coastal shallows, and the lagoon, allowing simultaneous observation of any active nests. With the aid of 8 x 40 binoculars, and 60 mm zoom spotting scopes with 15-45x eye-pieces, a team of experienced observers were able to record NZFT behaviour from stationary positions at > 30 m from the nearest foraging habitat, which precluded any risk of displacing the focal birds or interfering with their behaviour (minimum observer distances of 20–30 m are commonly maintained for nest watches; Parrish and Pulham Reference Parrish and Pulham1995a). Contact between observers was maintained using mobile phones, pre-notifying the next observer station as a NZFT left an observation field, aiming to follow NZFT from the time they left the nest site, to the time they returned to feed their chicks. Sighting times of NZFT and numbers of dives were quantified for each monitored habitat type, and dives were plotted using ArcMap 10 (ESRI Inc).

Figure 2. (a) New Zealand Fairy Tern foraging habitats monitored in the Mangawhai Harbour (shaded); grey symbols: observer positions; asterisks: NZFT nest sites; black circles: prey sampling sites A-J; (b) symbols: foraging dives of NZFT recorded in the 2010/2011 nesting season; (c) kernel density distribution of foraging NZFT at Mangawhai 2010/2011 (Land Information New Zealand (LINZ) topographic maps).
Prey sampling
Samples of potential prey were collected from areas where feeding activity of NZFT had been observed, using a fine-mesh beach seine net 5 m long and of 1.5 m high and mesh size of 1 mm with a weighted lead line that was pulled a distance of 5–30 m; the effective sampling width averaged 4 m. All fish samples were collected during daylight hours between 06h00 and 18h00 and within three hours of low tide.
Three replicate tows of the seine net were taken on sampling dates between 17 Nov 2010 and 20 Jan 2011 at sampling sites A-I (Figure 2a), with sites A, B, C and I sampled twice during the austral summer and sites E and G sampled three times. As NZFT were observed flying upstream of the upper data collection site (observer position 5; Figure 2a) on eight occasions, an opportunistic single replicate sample was taken at site J in the upper estuary on 26 Feb 2012.
All sampled fish were identified to genus and where possible to species level, and standard length (SL) was measured to the nearest 1 mm for all or up to 300 specimens per taxon and haul. Fish were retained as voucher material at the Auckland Museum, New Zealand. All fish were counted and abundances standardised per 10 m of haul length. As fish numbers were non-normally distributed across our samples, median and interquartile ranges of the abundance of all fish taxa were calculated for each sampling site. Size-distributions of the three most abundant fish species were assessed, using pooled data from replicate hauls for each site and sampling date.
Stable isotope sampling and analyses
Potential prey sources that occurred with a median abundance of greater than zero at least at one sampling site were assessed with stable isotope analyses. These were gobies Favonigobius sp. (n = 9), flounder Rhombosolea sp. (n = 4), and shrimp (n = 6) caught at confirmed NZFT foraging sites; a single juvenile parore Girella tricuspidata, was included as an illustrative assessment of an oceanic prey signature. Stable isotope analyses were conducted on homogenised whole prey.
Reference prey items were snap-frozen in liquid nitrogen upon collection, and subsequently stored at -80 °C until freeze-drying to constant weight and homogenising. Breast feathers of 29 19–20 day-old NZFT chicks (collected at the four breeding sites during New Zealand Department of Conservation banding operations 2007–2011; one adult wing under-covert feather from a predated female tern; and feathers from two adult and two immature specimens from the Natural History Museum Vienna, Austria (NMW 48.943 ♂ ad., NMW 48.946 ♂ immat., NMW 48.945 ♀ ad., July 1882, Manukau, A. Reischek; NMW 48.944 unsexed immat., collected prior to July 1866, exact locality not known, probably surroundings of Christchurch, J. v. Haast), were used for stable isotope analyses. All feathers were cleansed with 70% ethanol and subsequently dried and homogenised. Approximately 0.7 mg of sample were weighed and encapsulated in standard tin cups for stable isotope analyses.
Nitrogen (as an indicator of trophic position) and carbon (as an indicator of the primary carbon source) stable isotope ratios were determined at the National Institute for Water and Atmospheric Research (NIWA), Wellington, utilising a NA1500 (Fisons Instruments, Rodano, Italy) CHN elemental analyser linked to a DeltaPlus (Thermo-Fisher Scientific, Bremen, Germany) fully automated continuous flow isotope ratio mass spectrometer. Ratios of the rare to the common carbon (13C, 12C) and nitrogen (15N, 14N) isotopes were reported using delta notation, written as δ, in units of parts per thousand (‰), calculated against a limestone standard originally calibrated against the international standard Pee Dee Belemnite (for C) and against Nitrogen in air (for N). Isotope values were normalised and corrected against a suite of National Institute of Standards and Technology (NIST) standards. Repeat analysis of NIST and internal DL Leucine working standards produced data accurate to within 0.5‰ for δ15N and 0.4‰ for δ13C and a precision of better than 0.25‰ for δ15N and 0.29‰ for δ13C.
Data analyses
Core NZFT foraging areas were calculated as kernel density distributions of all recorded dives using the ArcGIS 10 Spatial Analyst Tool (ESRI Inc). Density contours were set to display the top 90% of spatial use.
Mann-Whitney Rank Sum Tests were performed in SigmaPlot version 11.0 (Systat Software Inc.) to compare abundance of fish per 10 m haul length in the mid-estuary with mangrove-vegetated shorelines, to those in the lower harbour. Kruskal-Wallis Tests were subsequently run to compare abundance between all sampling sites, followed by Dunn’s all pairwise multiple comparisons procedures. Kruskal-Wallis analyses were also run to test for differences in distributions of standard lengths, (a) including samples pooled from three replicate hauls at sites A-E taken on 1 Dec 2010, and (b) between samples from three replicate hauls taken at estuary sites A, B, C, and E on 07 Jan 2011. All tests were performed at a significance threshold of α = 0.05.
Likely dietary composition of adult and chick NZFT was first assessed using standard mass balance equations to determine prey proportions, assuming that only the two predominant fish taxa, goby and flounder, were consumed. Calculations followed Phillips (Reference Phillips2001):
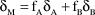 $${\rm{\delta }}_{\rm{M}} = {\rm{f}}_{\rm{A}} {\rm{\delta }}_{\rm{A}} + {\rm{f}}_{\rm{B}} {\rm{\delta }}_{\rm{B}}$$
$${\rm{\delta }}_{\rm{M}} = {\rm{f}}_{\rm{A}} {\rm{\delta }}_{\rm{A}} + {\rm{f}}_{\rm{B}} {\rm{\delta }}_{\rm{B}}$$
and
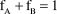 $${\rm{f}}_{\rm{A}} + {\rm{f}}_{\rm{B}} = 1$$
$${\rm{f}}_{\rm{A}} + {\rm{f}}_{\rm{B}} = 1$$
where δ represents the isotopic signatures for the mixture M and for sources A and B; fA and fB are the proportions of A and B in M (Phillips and Gregg Reference Phillips and Gregg2001). In our case:
 $${\rm{f}}_{{\rm{goby}}} = \left( {{\rm{\delta }}_{{\rm{feather}}} - {\rm{\delta }}_{{\rm{flounder}}} } \right)/\left( {{\rm{\delta }}_{{\rm{goby}}} - {\rm{\delta }}_{{\rm{flounder}}} } \right)$$
$${\rm{f}}_{{\rm{goby}}} = \left( {{\rm{\delta }}_{{\rm{feather}}} - {\rm{\delta }}_{{\rm{flounder}}} } \right)/\left( {{\rm{\delta }}_{{\rm{goby}}} - {\rm{\delta }}_{{\rm{flounder}}} } \right)$$
and
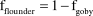 $${\rm{f}}_{{\rm{flounder}}} = 1 - {\rm{f}}_{{\rm{goby}}}$$
$${\rm{f}}_{{\rm{flounder}}} = 1 - {\rm{f}}_{{\rm{goby}}}$$
for feather signatures from NZFT adults and chicks, respectively.
Means and standard errors of stable isotope signatures of NZFT feathers from chicks and adults, and prey items were calculated. A juvenile parore Girella tricuspidata caught in the oceanic shallows was included to provide the signatures of an oceanic prey item. Prey δ13C and δ15N signatures were tested for normality and equal variance applying Shapiro-Wilk Tests, and differences between goby and flounder were assessed in unpaired two-sample t-tests. Subsequently, as shrimp also occurred in high numbers at several of our sampling sites, these were included as a third potential prey source, and compared with flounder and goby signatures in one-way ANOVAs. A prey polygon was constructed (Figure 4), assuming average assimilation rates of 3.4‰ δ15N per trophic level, and 0.8‰ δ13C from prey to predator, to account for fractionation during digestion and assimilation (Phillips and Gregg Reference Phillips and Gregg2003). All possible dietary compositions were calculated using IsoSource 1.3.1 software with 1% increments of possible source contributions and a tolerance level of 0.1 (Phillips and Gregg Reference Phillips and Gregg2001, Reference Phillips and Gregg2003) for NZFT chicks and adults respectively.
Results
Foraging habitat choice
From 140 hours of observation effort we collated data from 30.4 h of NZFT attendance to nesting, foraging and roosting areas (surveying up to five active nest sites). Monitored behaviour included foraging, roosting and transit between habitats, collating 139 sightings of NZFT (Table 1). In total, 21 complete tracks from nest site to the foraging areas and back to the nest were recorded. We recorded 405 foraging dives, and observed NZFT foraging in the oxbow lagoon on the Mangawhai sand spit, the shallows of the mid-estuary with mangrove shorelines, the shallow tidal pools on flats in the lower harbour, the shallows of the channels, and the shallows of the ocean (Table 1, Figure 2b). Their dives comprised a downward plunge with an opening of the wings so that generally the bill and head were submerged. The mid-estuary, which is characterised by at least one mangrove-vegetated shoreline and by extensive shallow channels at low tide, and the oxbow lagoons on the sand spit, in particular, were found to be core areas of NZFT foraging (Figure 2c), as well as hosting most roosting behaviour (Table 1).
Table 1. Summary of attendance, including foraging, roosting and transit behaviour of the New Zealand Fairy Tern (NZFT) population at NZFT habitats at Mangawhai in the 2010/2011 nesting season.

Prey abundance
Seine net sampling yielded 4,368 fish of at least 11 species, encompassing eight families, and shrimps including thousands of carids and mysids (Table 2). For two fish species, the bridled goby Arenigobius bifrenatus and the glass goby Gobiopterus semivestitus, these are the first records from Mangawhai Harbour. Estuarine gobies Favonigobius sp. contributed 92.8% of fish samples; and shrimps constituted the second most abundant taxon with roughly 2,000–3,000 individuals. The glass goby and bridled goby were the second and the third most abundant fishes, but each occurred at only a single sampling site. The bridled goby was only found in the lagoon and the glass goby at site J in the upper estuary. Small sand flounders (which were predominantly yellowbelly flounder R. leporina) regularly occurred in small numbers at sampling sites throughout the estuary.
Table 2. Survey summary of potential NZFT prey at identified foraging sites in Mangawhai Harbour and Harbour, the lagoon on the sand spit, and its adjacent shallows; medians, 25% and 75% quartiles shown for all fish species. All fish abundances standardised /10 m haul length.

Favonigobius sp. were highly significantly more abundant at sampling sites within the mid-Mangawhai Harbour (pooled A-C, E, F), than at sampling sites in the lower harbour (pooled D, G) (Mann-Whitney U statistic 1.00, T 12,30 = 79.00, P < 0.001). Kruskal-Wallis analyses revealed significant differences in numbers of Favonigobius sp. between sampling sites (H 8 = 34.13, P < 0.001), particularly when only sites within Mangawhai Harbour were included (H 6 = 28.93, P < 0.001). Subsequent Dunn’s pairwise multiple comparisons showed that B (diff of ranks = 19.89, Q = 3.08, P < 0.05), C (diff of ranks = 27.56, Q = 4.26, P < 0.05), and E (diff of ranks = 24.28, Q = 4.20, P < 0.05) had significantly higher Favonigobius sp. abundance than site G. There were no significant differences in Rhombosolea sp. abundance between sampling sites included in our study overall (H 8 = 9.70, P = 0.29), nor within the Mangawhai Harbour (H 6 = 3.89, P = 0.69).
Size distributions of Favonigobius sp. ranged from 18–38 mm SL (A), 24–40 mm (B), 20–40 mm (C), 23–41 mm (E), and 24–35 mm (F) in the mid-estuary, from 35–41 mm (D) in the lower harbour, and from 18–44 mm (I) in the oxbow lagoon during Nov-Dec 2010. In 7 Jan samples, smaller size classes were collected: 8–20 mm SL at C (n = 227, SL 7-19 mm), and low numbers of small specimens occurred at sites A (n = 1, 12 mm SL), B (n = 4, 12–18 mm), and E (n = 3, 12–15 mm SL) (Figure 3).

Figure 3. Size-distributions of Favonigobius sp., Arenigobius bifrenatus, and Gobiopterus semivestitus at sampling sites A-J, Mangawhai, plotted in order from upstream to downstream. No gobies were caught in the ocean (Site H).

Figure 4. Carbon and nitrogen stable isotope signatures (means and standard deviations) of New Zealand Fairy Tern feathers of adults and chicks, and potential prey taxa identified from seine net sampling (black symbols). Prey signatures, plus the fractionation factor of 3.4‰ for N and 0.8‰ for C, are indicated by blue symbols, where the dotted lines show the fractionation increment from the measured prey tissue isotope values.
Differences in SL distributions of Favonigobius were highly significant amongst sampled sites (A-E) in Mangawhai Harbour on 1 Dec 2010 (H 4 = 112.80, P < 0.001) and on 7 Jan 2011 (A-C, E; H 3 = 374.07, P < 0.001; Table S1 in the online supplementary material).
Dietary composition
Feather stable isotope signatures of NZFT chicks were 14.37 ± 0.90‰ δ15N and -15.61 ± 1.65‰ δ13C, those of adult birds 14.82 ± 1.85‰ δ15N and -15.53 ± 1.52‰ δ13C (Figure 4). Stable isotope signatures of museum specimens are additionally depicted separately in Figure 4; of these the lowest signature (12.61‰ δ15N, -18.25‰ δ13C) originated from an unsexed immature specimen which was probably collected in the surroundings of Christchurch, South Island.
Mass balance equations determined NZFT chick feather signatures were most likely the result of feeding on 80% by mass of gobies and 20% by mass of flounder, with proportions of 78% by mass of gobies and 22% by mass of flounders for the diet of adult birds. Flounder Rhombosolea sp. were significantly enriched in 13C compared to gobies Favonigobius sp. (T 11 = -7.87, P < 0.001; Figure 4). Nitrogen isotope signatures, in contrast, did not differ significantly between gobies and flounders (T 11 = -1.45, P = 0.17). Shrimp δ13C signatures were not significantly different from other potential prey (ANOVA F 2 = 3.41, P = 0.06); the differences in mean δ15N values were significant between all prey groups when shrimp were included in the analyses (ANOVA F 2 = 12.94, P < 0.001).
IsoSource modelling showed that gobies constituted the most substantial part of chick diet (with a likely range of 59–82% by mass), followed by flounders (18–22%) and may have included shrimps as potential prey (0–19% mass). The diet of adult NZFT was modeled to comprise 47–74% goby, 19–32% flounder, and 0–21% shrimp.
Discussion
Weighing only c.70 g, the NZFT is the smallest tern found in New Zealand (Falla et al. Reference Falla, Sibson and Turbott1979). In contrast to sympatric larger tern species, such as White-fronted Tern Sterna striata (Bräger Reference Bräger1998), the NZFT is not a plunge diver, but commonly feeds within the upper few centimeters of water. The NZFT typically breaks the momentum of its descending aerial dive just above the water surface by spreading its tail and wings, submerging usually just the bill, and part of its head, to seize prey (Fig. S1 in the online supplementary material). We estimate, based on culmen and head morphometrics, that if only the bill and head are immersed, as commonly observed in this study, these terns can feed on approximately the upper 5–8 cm of the water. This foraging method also enables the bird to utilise extremely shallow waters, such as tidal pools. Indeed, the breeding distribution of NZFT indicates that this subspecies only persists in shallow estuarine environments, which is consistent with the generally reported breeding habitat of the species (Higgins and Davies Reference Higgins and Davies1996).
NZFT had previously been observed to feed gobies Favonigobius lentiginosus, elvers Anguilla sp., and flounder Rhombosolea sp. to their chicks, with provisioning rates of ∼ 0.5–2 fish/h (Parrish and Pulham Reference Parrish and Pulham1995a) to peak provisioning bouts of four fish per 8-minute period during post-fledging parental care (Preddey Reference Preddey2008). The core 90% of NZFT foraging habitat fell almost exclusively on the oxbow lagoon on the sand spit, and in the mid-estuary that had at least one mangrove-vegetated shoreline. The shallows of the main channel in the lower harbour, and the shallows of the ocean were within the least densely used 10% of NZFT foraging habitats during our time of study. Our results indicate that those areas where suitable prey was most abundant were frequented most by NZFT for foraging, and we note that within those, the shallows were used most of all. Favonigobius sp., which according to our stable isotope source apportioning results constitute the main prey of NZFT, shelter in the pneumatophore zones provided by prop roots along the edges of mangroves at high tide. Consequently, highest goby abundance is associated with mangrove edges, with their distributions extending into mangrove stands and onto adjacent mudflats (Hindell and Jenkins Reference Hindell and Jenkins2005). As the tide lowers and the mangrove-vegetated areas and adjacent tidal flats fall dry, gobies move with the water into the channels and tidal pools of the upper and mid-estuary. Small-sized estuarine gobies can be flushed into the ocean where tidal currents are strong (Young and Potter Reference Young and Potter2003), but adult fish are able to largely stay within their preferred estuarine habitats when subject to tidal ranges as found at our study site (Gill and Potter Reference Gill and Potter1993, Potter and Hyndes Reference Potter and Hyndes1999). This is consistent with the low numbers of gobies overall, and the absence of specimens < 35 mm in the lower Mangawhai Harbour in our study. The recorded fish were suitably sized for NZFT prey (Figure 3), with only one specimen exceeding 9 cm, the maximum prey size to date observed to be taken by a NZFT (G. Pulham unpubl. data).
The bimodal size distribution of Favonigobius sp. (Figure 3) at site C in January showed settlement of a new size class. Minimum sizes of 11 mm and 9 mm are reported respectively for the two species of Favonigobius reported in New Zealand, F. lentiginosus and F. exquisitus (Chargulaf et al. Reference Chargulaf, Krück and Tibbetts2011), indicating recent settlement of Favonigobius larvae at mid-estuary site C during our January sampling at Mangawhai. These parts of the Mangawhai Harbour currently provide the suitable flow regimes (sensu Strydom et al. Reference Strydom, Whitfield and Wooldridge2002) to enable settlement and retention of Favonigobius larvae. No recruitment was evident in the lagoon by January 2010.
Our results highlight gobies as the most important prey for NZFT chick rearing, and indicate that most NZFT prey is likely to be of estuarine, rather than oceanic, origin. Yet, large variation in chick δ13C values may indicate a variation in the contribution of oceanic prey to offspring provisioning in NZFT between different breeding sites and years.
Nitrogen stable isotope data showed flounders and gobies to be at a similar trophic level. The 13C-enriched signatures of flounders compared to other fish taxa that were sampled, indicates a strictly benthic feeding habit of Rhombosolea spp. Favonigobius sp., in contrast, had significantly lower δ13C values than flounders, indicating a more planktonic diet. This is consistent with recent findings that Favonigobius spp. selectively feed on copepods and decapod shrimps (Chargulaf et al. Reference Chargulaf, Krück and Tibbetts2011), which occurred in our samples from the mid-estuary and lagoon, and which may also aggregate in mangrove pneumatophore systems at high tide (Modlin Reference Modlin1990, Buskey Reference Buskey1998, Reference Buskey2000). The stable isotope signatures we measured for shrimp suggested that these are indeed likely prey for gobies in the Mangawhai Harbour and oxbow lagoon, although smaller size-classes of shrimp would not have been captured quantitatively by our seine net sampling (the mesh size of 1 mm would have under-sampled mysids that have a body width and height < 1 mm).
These integrative findings highlight the nursery role of the mid-estuary (Chang et al. Reference Chang, Wang, You and Tzeng2006, Strydom et al. Reference Strydom, Whitfield and Wooldridge2003) for the most quantitatively important NZFT prey. The shallow topography, together with the structure provided by mangrove roots during the flood phase of the tidal cycle, would reduce current flow, enabling the successful transition in gobies from pelagic larvae to benthic juveniles. Mangrove roots also provide a refuge for adult gobies at high tide (Hindell and Jenkins Reference Hindell and Jenkins2005), and additionally increase goby abundance by providing sites for aggregations of their prey (Buskey Reference Buskey1998, Reference Buskey2000).
Our stable isotope results indicate that shrimps could be a substantial diet component for NZFT, potentially contributing up to 21% of the mass. In the 2011/2012 breeding season, adult NZFT were observed taking shrimp from the oxbow lagoon at Mangawhai (R. Zimmerman pers. obs.), but shrimp have never been recorded as food items delivered to chicks. Shrimps are likely to be of lower nutritional value than fish for chick rearing, and some foraging adult terns have been shown to distinguish between food items to be ingested by themselves, and those suitable for feeding to their offspring (Dänhardt et al. Reference Dänhardt, Fresemann and Becker2011). In many cases it was impossible to ascertain whether prey was captured by foraging NZFT during our study. Shrimps are less conspicuous than fish to human observers using binoculars or a spotting scope to monitor bird behaviour, and may, if ingested immediately upon capture, go undetected by the observer.
Productive foraging grounds in the vicinity of the breeding sites are vital in enabling the high chick provisioning rates observed in NZFT (Parrish and Pulham Reference Parrish and Pulham1995a, Preddey Reference Preddey2008). Without productive feeding grounds close to the nesting locations of NZFT, sufficiently frequent nuptial feeding of the incubating female and of the chicks at the nest, or local post-fledging tuition of offspring will not be possible. Reproductive failure in the conspecific AFT has been found to be associated with lack of suitable prey in the proximity of the breeding area in the Coorong (Paton and Rogers Reference Paton and Rogers2009), and recent declines in the Coorong AFT population coincided with a dramatic distribution contraction of small-mouthed hardyhead Atherinosoma microstoma as suitable prey for AFT (Paton et al. Reference Paton, Rogers, Hill, Bailey and Ziembicki2009).
Striking evidence of the direct linkage between impeded local foraging success near the breeding site and reduced reproductive output also comes from the congeneric Little Tern Sternula albifrons at its most important UK colony (Perrow et al. Reference Perrow, Skeate, Lines, Brown and Tomlinson2006, Reference Perrow, Gilroy, Skeate and Tomlinson2011a). In this close relative of the Fairy Tern (Bridge et al. Reference Bridge, Jones and Baker2005), construction of an offshore wind farm has led to a significant decline in the abundance of young-of-the-year clupeids, a major part of the diet for these Little Terns. This decline was associated with reduced foraging success in the birds, and unprecedented rates of egg abandonment and hatching failure (Perrow et al. Reference Perrow, Gilroy, Skeate and Tomlinson2011a).
Reduced foraging success as a result of major feeding habitat modification could similarly result in a decline in reproductive success of the NZFT at its most important breeding site at Mangawhai, New Zealand. This could cause a significant increase in the (already high) extinction risk of this Critically Endangered bird.
Apart from high prey abundances, a range of other factors are likely to additionally benefit the foraging activity of NZFT in the mid-Mangawhai Harbour. These include low disturbance rates (Jeffries Reference Jeffries2000) by walkers and dogs along currently mangrove-vegetated shores of this part of the estuary, no disturbance by motor-driven vessels at low tide, when the main channels are currently not navigable in this part of the harbour, and improved water clarity that may facilitate detection of prey.
Further research is needed to assess pivotal NZFT foraging habitats at other breeding sites, and across years. A crucial future research direction will also be assessment of prey selectivity per se: NZFT very likely select prey taxon-specifically by size, for example only relatively small flatfish can be ingested due to their body shape, while relatively large gobies would be preferable, offering a more substantial meal size and better detectability. As we had to avoid possible disruption and displacement of foraging birds, we could not select observer distances that would have allowed us to gather sufficient data on what size of prey was taken at the foraging grounds. Our results present a first basis for understanding, protecting and managing the food sources and the feeding and breeding habitats of the NZFT, which will be crucial to ensure this bird’s survival.
Supplementary Material
The supplementary materials for this article can be found at journals.cambridge.org/bci
Acknowledgements
This project was carried out under New Zealand Department of Conservation Research Permit AK-29785-RES and Ministry of Fisheries Special Permit 422. Funding was provided by the Faculty of Science of the University of Auckland to SMHI, and by the Royal Forest & Bird Protection Society of New Zealand Inc. and a BirdLife International Research Grant to SMHI and KB. We thank Alison Davis, Bev Woods, Darryl Jeffries, Jane Vaughan, Mandy Herrick, Jessa Cochrane and Bailey Lovett for help in the field, Gwenda Pulham, Rangi Zimmerman, Linda Wilson and Ernst Bauernfeind for helpful advice, Das Naturhistorische Museum Wien for generous provision of samples, and Leandro Bugoni and one anonymous referee for helpful comments. We gratefully acknowledge the Department of Conservation, Warkworth and Whangarei Offices, and About Tern Volunteers for their advice throughout our project.








